平成 28 年度
学位論文(博士)
ニチニチソウ栽培時の光環境制御による
抗がん剤成分 vinblastine の効率的な生産方法に関する研究
玉川大学大学院農学研究科
福山 太郎
1
目次
第 1 章 序論 ... 3 第 2 章 光質および光強度が生育と vinblastine 前駆体濃度に与える影響 ... 12 第 1 節 序論 ... 12 第 2 節 材料および方法 ... 14 実験 1 異なる光質が生育と vindoline および catharanthine 濃度に与える影響 .... 14 実験 2 異なる光強度の赤色光が生育と vindoline および catharanthine 濃度に与え る影響 ... 16 第 3 節 結果 ... 20 実験 1 異なる光質が生育と vindoline および catharanthine 濃度に与える影響 .... 20 実験 2 異なる光強度の赤色光が生育と vindoline および catharanthine 濃度に与え る影響 ... 20 第 4 節 考察 ... 22 第 3 章 赤色光下で生育したニチニチソウに対する紫外線 A 領域および 青色光照射が vinblastine 生産に与える影響 ... 39 第 1 節 序論 ... 39 第 2 節 材料および方法 ... 40 第 3 節 結果 ... 42 第 4 節 考察 ... 45 第 4 章 赤色光の光強度、UVA 強度、葉の老化が vinblastine 生産に与える影響 ... 56 第 1 節 序論 ... 56 第 2 節 材料および方法 ... 57 実験 1 異なる光強度の赤色光下で生育したニチニチソウに対する UVA 照射が vinblastine生産に与える影響 ... 57 実験 2 UVA照射強度が赤色光で生育したニチニチソウの葉における vinblastine 生産に与える影響 ... 57 実験 3 葉の老化が UVA 照射による vinblstine 蓄積に対して与える影響 ... 58 実験 4 UVA光源との位置・高さ・角度が UVA の強度に与える影響 ... 58 第 3 節 結果 ... 60 実験 1 異なる光強度の赤色光下で生育したニチニチソウに対する UVA 照射が vinblastine生産に与える影響 ... 602 実験 2 UVA照射強度が赤色光で生育したニチニチソウの葉における vinblastine 生産に与える影響 ... 61 実験 3 葉の老化が UVA 照射による vinblstine 蓄積に対して与える影響 ... 62 実験 4 UVA光源との位置・高さ・角度が UVA の強度に与える影響 ... 62 第 4 節 考察 ... 63 第 5 章 総合考察... 72 引用文献 ... 75 謝辞 ... 86
3
第1章 序論
ニチニチソウ(Catharanthus roseus L (G) Don.)には、100 種を超える monoterpenoid indole
alkaloids(MIA)が含まれることが知られている(van der Heijden et al., 1989)。この MIA の
中には、抗がん剤として利用されている vinblastine および vincristine(Fig. 1-1, Carter and
Livingston, 1976)、昇圧剤として利用されている ajmalicine が、それぞれ葉と根に含まれてい
る(Schmeller and Wink, 1998)。
vinblastineは、単量体 MIA の vindoline と catharanthine が縮合し、3‘, 4’-anhydrovinblastine
を介して合成される二量体 MIA である(Fig. 1-1)。また、vincristine は、vinblastine に含ま れる vindoline の N-メチル基がアルデヒド基に酸化された MIA である。 日本において、vinblastine および vincristine は、それぞれエクザールⓇおよびオンコビンⓇ の名称で抗がん剤として利用されている。また、これらのアルカロイドを元に vindesine や vinorelbineといった半合成アルカロイドが開発されており、それぞれフィルデシンⓇとナベ ルビンⓇの名称で抗がん剤として利用されている。これらの物質は、細胞分裂時の tubulin に 結合することで細胞分裂を阻害する作用を持っており、広範囲のがん患者に対して処方さ れている(Prakash and Timasheff, 1991)。また、vinblastine、vincristine、vinorelbine は、WHO
model list of essential medicines(2015)に掲載されており、抗がん剤の中でも重要性が高い薬
剤である。
しかし、4 種の抗がん剤の原料である vinblastine と vincristine の植物体に含まれる濃度は 極めて低いことが知られており、総じてこれらのアルカロイド由来の薬価は比較的高価で ある。過去にこの問題を解決するため、vinblastine と vincristine を全合成または半合成する 方法がいくつか報告されているが、効率やコスト面から実用化はされていない(Kutney et al., 1988; Misawa et al., 1988; Kuehne et al., 1991; Yokoshima et al., 2002; Shirahama et al., 2006;
Ishikawa et al., 2009)。また、細胞培養による大量生産方法も研究はされたものの成功してい
ない(De Carolis et al., 1990; Moreno et al., 1995; Croteau et al., 2000)。そのため、これらのア ルカロイドの供給はもっぱら圃場で大量栽培し、大量の植物体から抽出精製することで賄 われている。 ニチニチソウは、マダガスカルが原産の植物であり。熱帯および亜熱帯地方に自生してい る多年草で、栽培適温は昼間 24 ℃以上、夜間が 18 から 20 ℃とされている。日本は多くの 地域が温帯に属するため、11 月から 3 月までの平均気温が 18 ℃以下に低下する時期は、屋 外で圃場栽培することが難しいためか、アルカロイド生産用のニチニチソウの栽培は行わ れていない。そのため、日本ではニチニチソウ由来の抗がん剤およびその原料の供給を輸入 によって賄っている。 しかし、天然物由来成分を使用した薬品の輸入には、大きなリスクがあることが問題視さ れている。例として、多くの漢方薬の材料になる生薬のカンゾウ(甘草)がある。甘草は、 Glycyrrhiza属の根を乾燥させたもので、日本は平成 20 年に 1,267,395 kg を中国から輸入し
4 て利用している(日本漢方生薬製剤協会生薬委員会,2011)。近年世界的な漢方薬ブームの ため、甘草の需要が世界的に増加している。しかし、中国では環境保全のために野生のカン ゾウの採集規制を行っており、野生のカンゾウ生産量は限界があると予想される。そのため、 甘草の価格高騰や十分な供給が出来なくなる可能性が挙げられる。加えて、輸入品の甘草に 含まれる主要成分の glycyrrhizin 濃度のばらつきが大きいことも問題となっている。これら の問題を解決するため、カンゾウを国内で安定的に栽培する技術の開発が積極的に行われ ている(Afreen et al., 2005; kojoma et al., 2011; Sun et al., 2012; Yamamoto et al., 2002, 2003)。 ニチニチソウは、カンゾウと異なり栽培されたものを用いているものの、生産をおこなっ ている地域が限定されているため、カンゾウと同様の問題が起こると予想される。特に、抗 がん剤は発展途上国が豊かになるにつれ、使用量が増加することが予想される。そのため、 価格の高騰や供給不足を予防する観点から vinblastine および vincristine を安定的に生産する ためのニチニチソウ栽培技術が必要である。vinblastine および vincristine の安定生産を達成 するには、これらの MIA を生産する環境でニチニチソウを栽培することで可能になる。 近年、光環境を含めた高い環境調節能力を持つ植物生産システムとして植物工場が注目 されている。植物工場は人工光型と太陽光型の大きく 2 つに大別される。1 つ目の人工光型 植物工場は、閉鎖空間において人工光源と空調制御システムを用いることで外界の環境変 動の影響を受けない植物の栽培システムである。そのため、栽培する場所を選ばずに植物を 安定かつ周年生産することが可能である。また、光源を自由に設置できるため、栽培面を多 段化することが可能であり、土地面積あたりの栽培面積を圃場栽培と比べて大きくできる。 2つ目の太陽光型は、ガラス・プラスチック温室に高度な環境調節技術を組み込んだ栽培シ ステムで、光源は主に太陽光を利用する。人工光型に比べると環境調節能力および土地面積 あたりの栽培面積は劣るものの、様々な植物の周年生産が可能である。そのため、植物工場 は、vinblastine および vincristine が蓄積するニチニチソウの栽培環境を作り出すことが期待 できる。
二量体 MIA 前駆体である vindoline と catharanthine の植物体中の濃度は、二量体 MIA と 比較して極めて高いことから、二量体 MIA 合成における律速段階は vindoline と catharanthine のカップリング反応であることが知られている。vinblastine および vincristine を生産するに は、二量体 MIA 合成時のカップリング反応を促進できるかが重要であると考えられる。過 去の研究から、このカップリング反応が律速される要因として、2つの要因が挙げられる。 1つ目の要因は、このカップリング反応を触媒する酵素の遺伝子発現のタイミングと
vindolineおよび catharanthine の蓄積タイミングが異なる点である。vindoline と catharanthine
のカップリング反応には少なくとも 5 つの peroxidase が関与していることが確認されてい る(Endo et al., 1988;Sottomayor and Ros Barcelo, 2003;Sottomayor et al., 1998)。これらの
peroxidaseの中でも 3’, 4’-anhydrovinblastine 合成酵素として CrPRX1 が知られている(Costa
et al., 2008)。葉における CrPrx1 の遺伝子発現量は、葉の展開が完了していない段階と葉が
5
catharanthineの濃度は、完全展開時に最も高く、葉が老化するにつれ減少していく傾向にあ
る(Roepke et al., 2010)。そのため、CrPRX1 の酵素量と基質である vindoline および
catharanthine濃度の変化に不一致が生じる。
2つ目の要因として、二量体 MIA 前駆体である vindoline と catharanthine が葉の異なる細
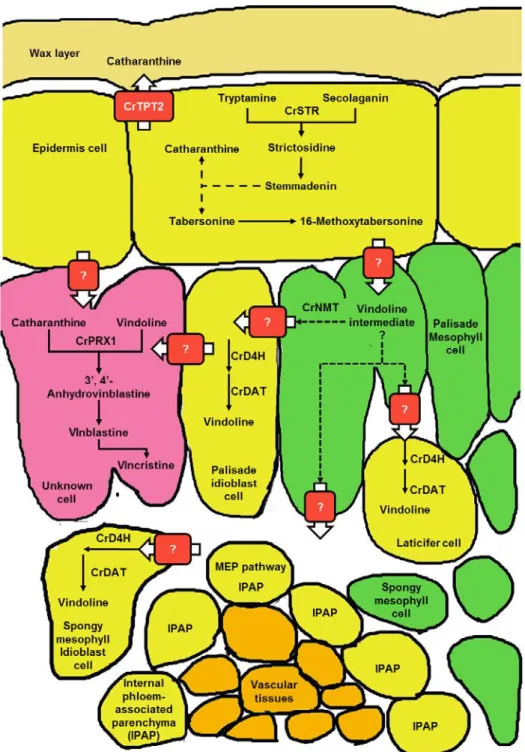
胞に蓄積していることである。vindolien は葉肉細胞中の異型細胞(idioblast cell)または乳 管細胞(laticifer cell)に局在している(Fig. 1-2, Mersey and Cutler, 1986)。一方の catharanthine は表皮細胞で合成され、その後 ATP binding cassette(ABC)トランスポーターの CrTPT2 に よって表皮細胞から葉表面のワックス層に移動する(Yu and De Luca, 2013)。そのため、二 量体 MIA 生合成を行うには、異なる細胞中に蓄積している vindoline と catharanthine が同一 の細胞に輸送され、カップリング反応を起こす必要がある。このカップリング反応が起こる 細胞や輸送体については、まだ明確なことはわかっていないが、Yamamoto et al.(2016)は、
Imaging MS と Single-cell MS を用いて茎の異型および乳管細胞から strictosidine と
catharanthine を検出している。そのため、二量体 MIA はこれらの細胞で合成されている可
能性が高い。加えて、Roepke et al.(2010)は、この輸送反応が自然条件ではほとんど起こ っていないことを指摘している。
植物の代謝物は生育に直接関与する一次代謝物と、生育には直接関与しない二次代謝物 に分類される。また、二次代謝物は大きく次の 3 つに分類される:terpenoid、alkaloid、
phenylpropanoidとその関連 phenol 化合物である(Croteau et al. 2000)。これらの二次代謝産
物は、植物が環境適応するために生産されていると考えられている。例としては、anthocyanin などの polyphenol 類は、高い抗酸化能力を有しており細胞内の恒常性の維持に利用されて いると考えられている(reviewed by Pietta, 2000)。一方で、ナス科植物の多くが生産する
alkaloid、アブラナ科植物の多くが生産する glucosinolate、シソ科植物が生産する monoterpen
などは、菌の侵入、昆虫や草食動物からの食害を防ぐための防御物質として機能していると 考えられている(Croteau et al., 2000; reviewed by Stamp 2003; reviewed by Wittstock and Halkier, 2002)。
ニチニチソウに含まれる vindoline、catharanthine、vinblastine は、菌や虫に対しても高い毒 性を持っており、tubulin に対する結合度から毒性の強さは vinblastine、catharanthine、vindoline の順であると推定されている(Prakash and Timasheff, 1991)。Catharanthine は、表皮細胞で生 合成された後ワックス層へ輸送されることが知られているが、これは菌や食害虫に対して 葉表面での防御に寄与していると考えられている(Roepke et al., 2010)。また、食害により 細胞が破壊されることで、葉肉細胞中の異型または乳管細胞に局在している vindoline と表 皮細胞で合成される catharanthine が混じりあうことでカップリング反応が起こり、
vinblastine が合成されることで外敵からの防御能力を高めているとも考えられている
(Roepke et al., 2010)。加えて、葉が傷つくと、MIA 量や MIA 生合成に関わる酵素の活性、 遺伝子発現量を増加させることが知られている(Frischknecht et al., 1987; Vázquez-Flota et al.,
6
(strictosidine synthase)、Tdc(tryptophan decarboxylase)、MIA 生合成を制御する転写因子の
Orca3(Octadecanoid-responsive Catharanthus AP2/ERF-domain)発現量を増加させることが報
告されている(Menke et al., 1999; Pasquali et al., 1992; Pauw et al., 2004)。これらの外敵から の防御反応には、MIA 生合成遺伝子の発現を一括制御するマスター制御転写因子が重要な 役割をしている。Orca3 は、マスター制御転写因子の一つであり、Tdc、Str、D4h (deacetoxyvundoline-4-hydroxylase)などの MIA 生合成酵素の遺伝子発現を制御しているこ とが知られている(van der Fits and Memelink, 2000)。特に、Tdc および Str は、それぞれ一 次代謝物の tryptophan を脱炭酸し、二次代謝の tryptamine に変換する酵素と、tryptamine と
secologanineをカップリングさせて MIA の骨格である strictosidine を合成する酵素をコード
しており、共に MIA 生合成の鍵酵素として知られている。
他方で、外敵とは関係の無い環境の変化に対して、MIA 量や MIA 生合成遺伝子発現量が 変化することがいくつか報告されている。特に二量体 MIA は、紫外領域の光に対して反応 を起こすことが知られている。紫外領域は、波長によって大きく 4 つに分けられており、
320から 380 nm の A 領域(UVA)、280 から 320 nm の B 領域(UVB)、100 から 280 nm の
C領域、100 nm 以下の真空領域である(Niu and Fujiwara, 2016)。
Hirata et al.(1991, 1992)は、紫外蛍光灯(ピーク波長 370 nm)を用いて UVA をマルチプ
ルシュートカルチャーに照射すると、二量体 MIA である leurosine および vinblastine が蓄積 されることを確認している。また、leurosineは青色光によっても蓄積する。Asano et al.(2010) は、野外で栽培したニチニチソウの切離葉に UVA を照射したところ、照射 5 日で二量体
MIA量(3‘, 4’-anhydrovinblastine 、vinblastine、leurosine の総量)は最大になり、その後減少
することを報告している。この結果から、UVA によって起こる二量体 MIA の蓄積は比較的 早い時間で完了すると考えられている。また、屋外で栽培したニチニチソウに UV カットフ ィルムを用いて太陽光の光質を変化させて栽培した実験では、380 nm 以下の光をカットす るフィルムを使用すると leurosine と vinblastine 濃度はフィルムを使用しないときに比べて 著しく低下した(Hirata et al., 1993)。一方で、290 nm 以下の光をカットするフィルムを使用 すると2つのアルカロイド濃度は、フィルムを使用するときに比べて高かった。この結果か ら、vindoline と catharanthine のカップリング反応には UVA が適していると考えらている。
UVA以外では、UVB を照射した研究例がいくつかある。Ouwerkerk et al.(1999)は、3 cm2
の葉片に UVB(ピーク波長 312 nm)の光を 2 分間照射し、その後 24 時間培養を行ったと ころ、CrTdc、CrStr の遺伝子発現量が増加した。また、strictosidine が 161 %、tabersonine が
79 %、3’, 4’-anhydrovinblastine が 54 %増加した。一方で、vindoline、catharanthine、vinblastine
は、それぞれ 11、14、8 %とそれぞれの前駆体が増加したにも関わらずあまり増加しなかっ た。Guo et al.(2014)は、1 ヶ月栽培した植物体に窒素の 50 %窒素追肥処理と UVB 処理(ピ ーク波長:305 nm)を同時に行ったところ、コントロール、窒素追肥のみの処理(コントロ ールと比べてアルカロイド量が低下)、UVB のみ(コントロールと比べてアルカロイド量は 増加)の処理に比べて vindoline、catharanthine、vinblastine 量が増加したことを報告してい
7
る。これらの結果から、UVB の照射は MIA の総量や単量体の増加に効果があると考えられ る。
光以外では、乾燥(Frischkecht et al., 1987)、浸透圧(Chang et al., 2014)、肥料中窒素濃度 (Abdolzadeh et al., 2006; Gholamhoss et al., 2011)や硝酸・アンモニア態窒素比(Guo et al.,
2012)の研究がされている。これらの処理は、UVB 処理と同様に MIA の全体量が増加する
傾向にある。
UVBの反応を含めた MIA の全体量を変化させる反応の多くは、ストレス応答によるもの
と推測されている。UVB の場合、UVB は可視光に比べて光子あたりのエネルギーレベルが 高く、細胞酸化や DNA を損傷することが知られている。Ramani and Cheliah(2007)は、ニ チニチソウの培養細胞に UVB を照射すると 4 分間でおよそ 7 倍程度活性酸素種(ROS)が 増加することを報告している。また、UVB 照射された培養細胞に含まれる catharanthine 量 は、非照射時に比べて 3 倍程度増加する。これらの UVB に対する応答は mitogen activated
protein kinase(MAPK)カスケードの関与が示唆されている。MAPK カスケードは、生物が
ストレスを受けた時に働くカスケードとして知られている(reviewed by Zhang and Klessig,
2001)。このカスケード下流にアルカロイド生合成を促進する因子があるため、アルカロイ
ド生合成が促進されたものと考えられる。実際に Raina et al.(2012)は、ニチニチソウの
MAPKの一つである CrMpk3 が UV 照射によって発現量が上がること、および CrMpk3 を過
剰発現させると CrStr などのアルカロイド生合成関連遺伝子の発現量を増加させ、vindoline、
catharanthine、vincristine 量が増加することを報告している。
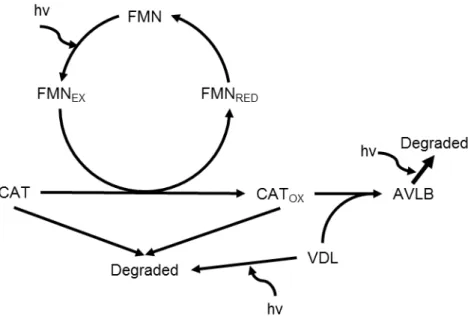
一方で、UVA による二量体 MIA 合成反応は非酵素的に起こる vindoline と catharanthine の カップリング反応が関係している可能性が高いことが指摘されている。Hirata et al.(1993) は、vindoline と catharanthine の標準品、flavin mononucleotide(FMN)および塩化マンガンを
Tris-HClバッファーに溶解させた混合液に 370 nm の UVA を照射することで、非酵素的に
vindolineと catharanthine が縮合し、3‘, 4’-anhydrovinblastine が合成されることを確認してい
る。また、UVA より合成量は少ないが 450 nm の青色光でも 3‘, 4’-anhydrovinblastine の合成 が確認されている。屋外で栽培したニチニチソウの葉に対して、4 または 25 ℃の条件下で
UVA を照射したところ、両方の条件で同じように二量体 MIA が検出された(Asano et al.,
2010)。4 ℃の低温条件下では酵素反応が極めて起こりにくいため、UVA による二量体化反
応は非酵素的な反応が大きく関与していると考えられている。この非酵素的カップリング 反応は、FMN が UVA または青色光によって励起され、励起された FMN を介して
catharanthine が酸化されることでカップリング反応が生じるため起こると考えられており、
生体内でも同様の反応が起こっていると予想される(Fig 1-3; Hirata et al. 1993;1999)。 本研究では、この UVA または青色光による vindoline と catharanthine カップリング反応促 進効果に着目した。Hirata et al.(1993;1999)の研究結果から、UVA または青色光による
vindolineと catharanthine のカップリング反応は化学反応によって起こることから、基質とな
8
とで、大量の vinblastine や vincristine を合成させることが可能であると予想される。また、 効率的に vinblastine や vincristine を生産するには、アルカロイド生産者であるニチニチソウ のバイオマスを増加させることで、これらのアルカロイドの収量を増やすことが可能であ ると考えられる。加えて、光環境の制御は、近年の light emitting diode(LED)の発達などに より光質や光量など制御が容易なため応用性が高い。他方、過去にニチニチソウを完全に人 工照明のみで栽培した例は組織培養など限定的である。そのため、ニチニチソウの生育やア ルカロイドに光環境が及ぼす影響については、十分に明らかではない。
そこで本研究では、玉川大学内にある LED 植物工場研究施設 Future Sci Tech Lab(FST
Lab)にてニチニチソウを様々な光条件下で栽培し、vinblastine が効率的に生産される光条
件の探索を行った。さらに、光環境制御による二量体 MIA 生産の可能性について考察し、 植物工場による効率的な MIA 生産の方法について提案した。
9
Fig. 1-1 Biosynthesis of 3’, 4’- anhydrovinblastine, vinblastine, vincristine, and leurosine from monomeric precursors catharanthine and vindoline (Asano et al. 2010).
10
Fig. 1-2 The spatial intracellular organization of MIA pathway in different plant cells of C. roseus. Broken arrows represent multi-step or uncharacterized reactions. Bold arrows represent transporters. Question marks on a bold arrow represent uncharacterized transporter. Biosynthetic enzymes are abbreviated as follows: CrSTR, strictosidine synthase; CrD4H, deacetoxyvundoline-4-hydroxylase; CrNMT, N-methyltransferase; CrDAT, deacetoxyvundoline 4-O-acetyltransferase; CrPRX1, peroxidase. Figure was modified from Facchini and De Luca (2008). Localization of the enzymes was described according to St-Pierre et al. (1999), Murata and De Luca. (2005).
11
Fig. 1-3 Reaction model of the coupling reaction of catharanthine between vindoline for the synthesis of 3’, 4’-anhydrovinblastine. The substrates, reaction intermediates and products are abbreviated as follows: hν, UVA light; FMN, flavin mononucleotide; FMNEX, FMN excited by UVA
light; FMNRED, FMN reduced by the oxidation of catharanthine; CAT, catharanthine; VDL, vindoline;
CATOX, oxidized catharanthine; AVLB; 3’, 4’-anhydrovinblastine. Figure was modified from Hirata
12
第2章 光質および光強度が生育と vinblastine 前駆体濃度に与える影響
第1節 序論 植物は、光を自身の生育にあわせて様々な方法で利用している。その1つが光合成で、独 立栄養生物である植物は生存に必要なエネルギーを光合成により獲得している。一方で、動 物のように自身の生存に適した環境に移動することが出来ない植物は、自身の生存環境に 適応するために、光を生存環境について知るための情報として用いている。光が持つ特性の 中でも光質は、植物の発芽や花芽形成など様々な影響を与えることが知られている (reviewed by Timmermans et al., 2010)。植物は、光を光質に応じた光受容体で受光し、この光受容体からのシグナルに応じて転写 因子の発現が促進され、それぞれの光質に応じた反応が起こる。現在までに、赤・遠赤色光 受容体として phytochrome(PHY)、青色光・UVA 受容体として cryptochrome(CRY)および
phototropin(PHOT)、UVB 受容体として UV response locus 8(UVR8)が確認されている
(reviewed by Timmermans et al., 2010;Rizzini et al., 2011)。一方で、光受容体を介さない反 応も知られている。UVR8 は、低光量であれば UVB を受光し、シグナル伝達が可能である が、光量が高い場合 UVB は細胞内に活性酸素種を発生させるため酸化ストレスが生じる。 このストレス因子が MAPK カスケード等を通してシグナルが送られることで、植物の応答 が起こる。 光質の影響を受けて植物の形態や生育に大きな影響を及ぼすことが知られている。大嶋 ら(2015)は、様々な赤青混合比の下でレタスを栽培したところ青色光を 50 μmol m-2 s-1以 上照射すると同じ光量の赤色光に比べて乾物重量が低下することを報告している。著者ら は、青色光による葉の展開の阻害が受光面積を低下させたこと、および青色光は赤色光に比 べて光合成効率が悪い点を青色光による乾物重量の低下の要因に挙げている。前者は、CRY からのシグナルが、葉を展開させる作用のある gibberellin 含有量の低下を引き起こすため、 葉の展開阻害が起こったと推定している。後者は、青色光が carotenoid や anthocyanin とい った色素に吸収されてしまうため、赤色光での栽培に比べて光合成に利用できる光子の量 が少なかったと推定している。 一方で、光質の変化は、植物の二次代謝産物にも影響を与えることが知られている。例と しては、シロイヌナズナやレタスでは、青色光や UVA が照射されると anthocyanin 合成が誘 導され anthocyanin 量が増加する(Lin et al, 1996;海老澤ら,2008;庄子ら,2010)。この反 応は、CRY が青色光や UVA を受光し、anthocyanin 合成酵素の遺伝子発現を誘導するために 起こる。また、外敵からの防御に関わる物質に対しても効果があることが知れており、タイ ムの実生において抗菌作用の持つ terpen 類の thymol や γ-terpinen が赤色光照射により増加 し、遠赤色光が照射されることで効果が打ち消されることから、PHY が生合成の制御に関 与していると予想されている(Tanaka et al., 1989)。
13
ていることが知られている(Aerts and De Luca, 1992)。他方、光受容体を介さない反応とし ては、第 1 章でも述べたが、UVA や青色光のような波長の短い光を照射することで二量体
MIAが合成される反応が挙げられる(Hirata et al., 1991;1992;1993)。
ニチニチソウの研究において、可視光域の光質がニチニチソウに与える影響について調べ た論文は Aerts and De Luca(1992)と Hirata et al.(1991;1992;1993)の報告程度しか存在 しない。特に、実生や組織培養でない植物体に対して行われた研究は、Hirata et al.(1993) および本研究室で行われてきたもののみである。本研究室で、過去行った研究では青色光処 理を行ったニチニチソウでは草丈が、赤単色、赤青混合光および白色蛍光灯に比べて大きく なったが、バイオマスや葉数が減少したことを報告している。一方で、vinblastine および vincristine は、すべての試験区で検出されなかった。この結果から、vinblastine および vincristineを可視光のみで生産させることは難しいと考えられる。他方で、ニチニチソウの バイオマスを増加させることには利用が可能であると考えられた。また、vinblastine 前駆体 である vindoline と catharanthine に対する可視光の影響についてはほとんどわかっていない。 そこで本章では、LED を利用して赤単色、青単色、赤青混合、白色蛍光灯下でニチニチソウ を栽培し、光質がニチニチソウの生育と vindoline および catharanthine に与える影響につい て調査を行った。加えて、最もアルカロイド生産に適した光質における光強度の影響を調査 した。
14 第2節 材料および方法 実験1 異なる光質が生育と vindoline および catharanthine 濃度に与える影響 1. 材料 材料はニチニチソウ ‘タイタン’ ダークレッド((株)M&B フローラより購入。)を使用 した。 2. 育苗条件 水道水を十分に給水させた 23 mm╳23 mm╳27 mm のウレタンマットに 200 粒播種を行 った。播種 3 日後まで暗処理を行い、その後光合成有効光量子束密度(PPFD)で 100 μmol m-2 s-1の白色蛍光灯下で栽培を行った。PPFD の測定は光量子メーター(LI-190SA、LI-COR) を用いた。光周期は、明期 18 時間および暗期 6 時間とした。室温は 23 ℃に設定した。播
種 7 日後、電気伝導率(EC)1.0 dS m-1および pH 6.0 の OAT ハウス A 処方(OAT アグリオ
株式会社、Hanyu and Shoji, 2002)の循環式の湛液水耕(DFT)装置に移した。DFT 装置は、
635 mm╳410 mm╳80 mm のポッドに水耕液の注入口と排出口が着いており、このポッド に 200 個の穴の開いた発泡スチロール板を設置したものである。この発泡スチロール板の 穴に一つ飛ばしに苗を 100 本植えた。 3. 光質処理条件 播種 35 日後、子葉および初成葉を除き下部から数えて第 3 葉対が出葉した段階で、異な る 4 つの光処理区を設定した薄膜水耕(NFT)装置に移植した。光処理区は白色蛍光灯(FL)、 LEDによる赤単色(R)、青単色(B)、赤青混合光(RB、R:B 比は PPFD で 2:1)を設定し た。光強度はすべて PPFD 150 μmol m-2 s-1とした。Fig. 2-1 に各光源のスペクトルを表記し た。各光源のスペクトルは英弘精機製の分光放射計(MS-720)で測定を行った。水耕液の 条件は EC 1.5 dS m-1およびpH 5.6 とした。光周期および室温は、育苗時と同様の設定とし た。 光処理で用いた LED は昭和電工アルミ販売(株)製の水冷式 LED パネルを使用した。こ の LED パネルは 119 mm×1000 mm で、赤色および青色 LED がそれぞれ 2 個づつ合計 4 個 セットが、長さ方向に 30 個 2 列配置されていた。 4. vindolineおよび catharanthine の抽出 凍結保存された全葉を液体窒素中で乳棒および乳鉢を用いて凍結粉砕を行った。収穫し た葉と同じ重量のメタノールを加え、ボルテックスした後、超音波による抽出を 30 分行っ た。混合液を 3000 ×g、室温で 5 分間遠心分離を行い、上清を回収した。抽出液を 100 ml にメスアップし、2 ml 採集した。ロータリーエバポレーターを持ちいて、この粗抽出液を 40 ℃で乾固するまで減圧濃縮した。残渣に 0.5N 硫酸および酢酸エチルをそれぞれ 2 ml 加 溶解させた。この混合液を分画し、水層を回収した。残った有機層に 2 ml の 0.5N 硫酸を加
15 え抽出操作を行った。これを二度繰り返した。回収した水層を蒸留水で 10 ml にメスアップ した後、メタノールおよび蒸留水で平衡化させた固相抽出カラム(Sep-Pak C18,Waters)に 試料を加えて、10 ml の蒸留水で洗浄後、10 ml のメタノールで溶出した。溶出液を減圧下、 45 oCで濃縮乾固し、1 ml のメタノールに再溶解後,0.45 µm の PTFE シリンジフィルター でろ過したものをアルカロイド抽出試料液とした。この試料液を高速液体クロマトグラフ ィー(HPLC)で分析した。 5. HPLC条件 HPLC条件は実験 1 および 2 で共通とした。HPLC システムは Chromaster(日立ハイテク ノロジーズ)を用いた。カラムは CapcellpacC18 MGII(4.6 × 250 mm,5 μm,資生堂)を 用いた。注入量は 10 µl、流速は 1.0 ml min-1、カラムオーブン温度は 40 oCに設定した。ア ルカロイドはダイオードアレイで検出し、測定波長は 250 nm に設定した。移動層は 0.02 M リン酸緩衝液(pH7.0,溶媒 A)とアセトニトリル(溶媒 B)を用いた。グラジエント条件 (溶媒 A の容積:溶媒 B の容積)は 0-3 min 70 : 30、3-8 min 40 : 60、8-25 min 40 : 60、25-27
min 25 : 75、27-30 min 25 : 75 とした。vindloine および catharanthine の同定と定量は、vindoline
標準品と catharanthine sulfate(ともに LKT Laboratories Inc.より購入)を用いて保持時間(RT) と吸収スペクトルから同定を行い、定量はメタノールに溶解した標準品を分析サンプルと 同条件で HPLC による分析を行い作成した検量線を用いて行った。
6. vindoline/catharanthine比
vindoline/catharanthine比は Equation 1 で算出した。
vindoline/catharanthine ratio
=catharanthine concentration 336.43vindoline concentration 456.53⁄ ⁄ ∗ ∗ (Equation 1) *vindoline および catharanthine の分子量
7. 株あたりアルカロイド収量
株あたり vindoline および catharanthine 収量は Equation 2 で算出した。 alkaloid yield
16 総アルカロイド収量は Equation 3 で算出した。
total alkaloid yield
= vindoline yield + catharanthine yield (Equation 3)
実験2 異なる光強度の赤色光が生育と vindoline および catharanthine 濃度に与える影響 1. 材料 材料は実験 1 と同じ品種を用いた。 2. 育苗条件 水道水を十分に給水させた 23 mm╳23 mm╳27 mm のウレタンマットに 200 粒播種を行 った。播種 3 日後まで暗処理を行い、その後光強度 100 μmol m-2 s-1の白色蛍光灯下で栽培 を行った。播種 7 日後、EC を 1.5 dS m-1および pH が 5.6 の OAT ハウス A 処方の水耕液が 含まれた DFT 装置に移した。DFT の水耕液は 7 日に 1 度交換を行った。また、終始エアレ ーションを行った。光周期は、明期 18 時間および暗期 6 時間とした。室温は 23 ℃に設定 した。播種 35 日後、4 つの異なる光処理区を設定した DFT 装置に移植した。 3. 赤色光強度処理 処理 1 処理区の光強度は、75、150、300、600 μmol m-2 s-1の赤単色光とした。各試験区 6 株ずつ 光処理を行った。光処理中の光周期、水耕液および室温条件は育苗時と同じにした。播種 63 日後、収穫を行った。収穫時の生育調査として草丈、株あたり新鮮重および全葉新鮮重を測 定した。測定後、第 4 葉をサンプリングしビニルパックに入れ−35 ℃で保存し、これをアル カロイド分析のサンプルとした。 処理 2 処理区の光強度は、150、250、300、350 μmol m-2 s-1の赤単色光とし、各試験区 15 株ずつ 処理を行った。光処理中の光周期、水耕液および室温条件は育苗時と同じにした。播種 63 日後、収穫を行った。収穫時の生育調査としてすべての株で株あたり新鮮重を測定した。15 株のうち、5 株を草丈、全葉新鮮重および乾物重を測定するのに用いた。他の 5 株から第 4 葉をサンプリングしビニルパックに入れ−35 ℃で保管し、アルカロイド濃度の測定に用い た。残りの 5 株の第 4 葉から直径 7 mm のリーフディスクを 30 mg 作成し、2 ml のマイク ロチューブに入れ液体窒素で凍結させた。凍結したマイクロチューブは−80 ℃で保管され、 遺伝子発現解析のサンプルとして利用した。 4. 乾物重の測定
17 収穫したニチニチソウの葉、根および残りを分割して茶封筒に入れ、85 ℃で 48 時間以上 乾燥をさせた。その後、デシケーターで放熱し、重量を測定した。乾燥、放熱、重量測定を 2回以上繰り返し、数値の減少が無くなった時点の値を乾物重とした。 5. shoot/root比 shoot/root比は、Equation 4 を用いて算出した。
shoot/root ratio =shoot dry weightroot dry weight (Equation 4)
6. 地上部含水率
地上部含水率は、Equation 5 を用いて算出した。 water content
=shoot fresh weight − shoot dry weightshoot fresh weight × 100 (Equation 5)
7. vindolineおよび catharanthine の抽出 処理 1 アルカロイドの抽出は Asano et al.(2010)の方法を改良した。凍結保存された葉を液体窒 素中で乳鉢と乳棒を用いて粉砕した。この試料にメタノールを 10 ml 加え、30 分間、超音波 抽出した。混合液を 3000 ×g、室温で 5 分間遠心分離を行い、上清を回収した。この残渣 に 10 ml のメタノールを加えて、抽出、上清を回収する操作を 2 回繰り返した。回収した上 清を減圧下、45 oCで濃縮乾固し、この残渣を 1 ml のメタノールに再溶解させた。再溶解液 を 0.45 µm の PTFE シリンジフィルターでろ過したものをアルカロイド抽出試料液とした。 この試料液を HPLC で分析した。 処理 2 凍結保存した葉を 48 時間以上凍結乾燥し、乾燥した葉を乳鉢と乳棒を用いて粉砕した。 粉末サンプルを秤量し、2 ml マイクロチューブに移した。これに 10 倍量のメタノールを加 え、30 分間超音波抽出を行った。その後、4 oC 、15,000×g で遠心分離を行い、上清を回収 した。残渣に再びメタノール 500 μl を加え、30 分間超音波抽出し、その後 4 oC、15,000×g で遠心分離、上清の回収をする作業を 2 回繰り返した。集めた上清を 0.45 µm の PTFE シリ ンジフィルターでろ過したものをアルカロイド抽出試料液とした。この試料液を HPLC で 分析した。
18
8. HPLC条件
HPLC条件は実験 1 と同様にした。
9. RNA抽出
凍結保存した葉を乳鉢と乳棒で粉砕後、RNeasy Plant Mini Kit(Qiagen)および RNase free
DNase (Qiagen)を用いてマニュアルに従って総 RNA を抽出した。抽出した RNA は、分光光
度計で 260 nm および 280 nm の吸光度を測定し、抽出した RNA の濃度および A260/280 か ら RNA の純度を求めた。また、総 RNA のアガロース電気泳動を行い 18S および 28S ribosome
RNA のバンドの有無から総 RNA が分解されていないか確認を行った。アガロース電気泳
動は、総 RNA 1 µl、loading buffer 1 µl、RNase free H2O 8 µlの混合液を 65 oCで 5 min 保温し
たのち、氷上で急冷却し RNA の変成を行った。この変成液を 1 %アガロースゲルと TAE
bufferで電気泳動を行った。
10. cDNA 合成
RNAの純度が 1.9 以上でかつ、RNA の分解がされていない RNA 300 ng を cDNA に逆転
写した。逆転写には、PrimeScriptTM RT regent Kit (Perfect Real Time)(タカラバイオ(株))を
使用し、マニュアルに従って逆転写を行った。 11. 定量 PCR
DNase free H2O で 10 倍希釈した cDNA を鋳型 DNA とした。測定には、Thermal Cycler
Dice Real Time System Single(タカラバイオ(株))を使用した。反応液は、SYBER Premix
EX Taq II(タカラバイオ(株))5 µl、forward プライマー(50 µM)0.08 µl、reverse プライ
マー(50 µM)0.08 µl、鋳型 DNA 1 µl、DNase free H2O 3.84 µlとした。PCR の条件は、95 oC
で 30 秒間の加温した後、95 oCで 5 秒、60 oCで 30 秒の反応をを 40 サイクル行った。各サ ンプルを 3 連で測定し、その平均値を結果として採用した。サイクル数の決定は二次微分極 大法で行った。 プライマーの特異性を確認するため、PCR 反応終了後、融解曲線分析を行った。加えて、 PCR増幅産物の確認のため、2 %アガローゲルで電気泳動を行い、バンドから PCR 増幅産 物の塩基数を確認した。 測定を行った遺伝子と各プライマーのシーケンスを Table 2-1 に示した。定量は、各測定 遺伝子の検量線を作成して行った。 リファレンス遺伝子は、Wei(2010)が報告している発現量の安定度が比較的高い CrEf1
α、CrUbq6、CrUbq11 の測定を行った。Vandesompele et al. (2002)および Pfaffl et al. (2004)の
方法を参考にして、安定度の高い CrEf1αおよび CrUBQ11 をリファレンス遺伝子とし、
19 corrected expression value
= target gene expression value
�𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶1𝛼𝛼 expression value × 𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶11 expression value
2 (Equation 6)
12. 統計処理
本実験で得られたデータは、統計解析ソフト JMP8(SAS Institute, Japan)を用いて解析し た。各サンプル間の差について Tukey’s HSD または Tukey-Kramer 検定を用いて多重比較を 行った。多重比較の有意水準は 5 %とした。また、実験 1 の草丈、最大葉長、最大葉幅は、 測定日ごとに分散分析を行った。
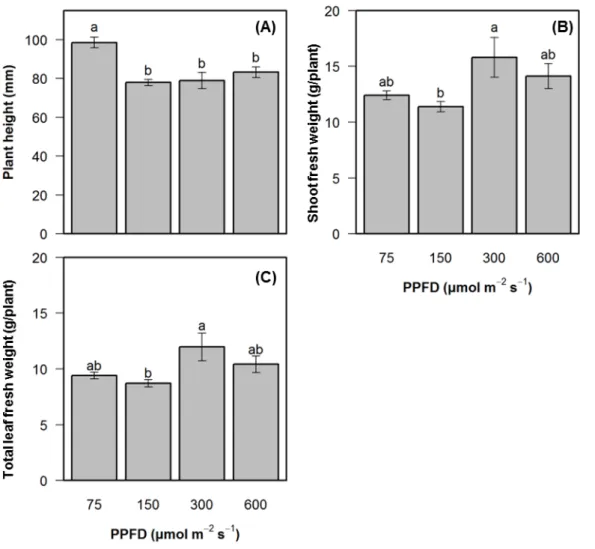
20 第3節 結果 実験1 異なる光質が生育と vindoline および catharanthine 濃度に与える影響 異なる光質下で 28 日間栽培を行ったニチニチソウは、R 処理区で他の試験区に比べ、地 上部の成育が大きく、葉が大きく展開している様子が見られた(Fig. 2-2)。一方で、大きく 葉巻を起こしていた。B 処理区で生育した株の茎は、他の試験区に比べて赤く着色していた。 Fig. 2-3に草丈、最大葉長、葉幅の経日変化を示した。これらのパラメーター間には、相 関係数 0.8 以上の高い相関が見られた。草丈は、処理 28 日後の R 処理区で最も大きかった。 また、処理 14 日後から光質間で差が見られた(Fig. 2-3 A)。また、最大葉長も草丈と同様に 処理 14 日後から光質間で差が見られ、R 処理区の処理 28 日後に最も大きく、次に RB、FL、 B処理区の順であった(Fig. 2-3 B)。最大葉幅も収穫時 R 処理区で最も大きく、その後の順 番も同じであった。また、処理後 14 日から光質間で差が表れた(Fig. 2-3 C)。 地上部新鮮重は、R 処理区で最も大きく、次に FL、RB、B 処理区の順であった(Fig. 2-4)。 葉部新鮮重は、地上部新鮮重と同様に R 処理区が最も大きかった。しかし、その後の順番 は RB、FL、B 処理区の順であった。この順番は、試験区に含まれる R 処理区の光量と一致 した。FL 処理区は、他の試験区と比べて地上部新鮮重に対して葉部新鮮重が少ないことか ら、茎部の重量が他の試験区に比べて大きかった。 全葉に含まれる vindoline および catharanthine 濃度は、有意差は無いものの RB 処理区で 最も大きく、次に R、FL、B 処理区の順であった(Fig. 2-5)。vindoline/catharanthine 比は、 R処理区で最も低く、FL 処理区で高くなる傾向が見られた。 vindoline、catharanthine、総アルカロイド収量は、濃度に大きな変化がなかったことから、 葉部新鮮重量の影響を大きく受けた。そのため、R 処理区で最大であった(Fig. 2-6)。 実験2 異なる光強度の赤色光が生育と vindoline および catharanthine 濃度に与える影響 処理 1 光強度処理 28 日後のニチニチソウの様子を Fig. 2-7 に示した。600 μmol m-2 s-1で生育し た株の下位の葉に黄化または白化が観察された(Fig. 2-7 D)。この症状は葉脈に近い部分か ら起こっていた。75 μmol m-2 s-1で生育した株の葉は葉脈間が盛り上がる傾向が見られた(Fig. 2-7 A)。300 μmol m-2 s-1で生育した株は、他の試験区と比べて葉数が増加している傾向がみ られた(Fig. 2-7 C)。 草丈は、75 μmol m-2 s-1で最も大きかった(Fig. 2-8)。地上部および全葉部新鮮重は、共に 300 μmol m-2 s-1で最も大きかった。
第 4 葉に含まれる vindoline および catharanthine 濃度は、150 μmol m-2 s-1で最も高かった
(Fig. 2-9)。また、光強度が高くなるにつれ濃度が低下した。 処理 2
21
s-1で最も小さく、光強度の増加に伴って重量が増加した(Fig. 2-10 B)。全葉部新鮮重は、
350 μmol m-2 s-1で最も大きかったものの、他の処理区と比べて大きな差は見られなかった
(Fig. 2-10 C)。全葉部、根、地上部、全乾物重は、光強度の増加に伴って増加する傾向が見
られた(Fig. 2-11)。shoot/root 比は、150 μmol m-2 s-1で最も大きく、光強度の増加に伴って
減少する傾向が見られた(Fig. 2-12)。地上部含水率は、shoot/root 比と同じ傾向を示した(Fig. 2-12)。
第 4 葉に含まれる vindoline および catharanthine 濃度は、150 μmol m-2 s-1で最も高かった
(Fig. 2-13)。濃度は光強度が高くなるに従って低下した。
Fig. 2-14にアルカロイド生合成関連遺伝子発現量の測定結果を示した。CrTdc 発現量は、
150から 300 μmol m-2 s-1にかけて増加する傾向があった。一方で、350 μmol m-2 s-1で最小値
を示した(Fig. 2-14 A)。CrStr 発現量は、150 μmol m-2 s-1で最も発現量が多く、光強度の増
加に伴って減少する傾向を示した(Fig. 2-14 B)。CrD4h 発現量は、250 μmol m-2 s-1で最も発
現量が高く、光強度が増加に伴って発現量が減少した(Fig. 2-14 C)。CrDat 発現量は、150
から 300 μmol m-2 s-1にかけて減少する傾向があった(Fig. 2-14 D)。Orca3 発現量は、150 μmol
22 第4節 考察 1. 異なる光質が生育に与える影響 他の光質に比べて R 処理区で生育が大きくなったのは、葉の受光面積の増大が総光合成 量を増加させたことが要因であると考えられた(Fig. 2-4)。特に最大葉長や葉幅は、光処理 14日後から差が表れており R 処理区での葉面積の拡大が光処理の早い段階で起こったと推 定される(Fig. 2-3)。また、葉の生育に関わるパラメーターの大小関係が試験区に含まれる 赤色光の光量の大小関係と一致することから、葉の生育は照射光中に含まれる赤色光の光 量によって制御されていると考えられた(Fig. 2-3 および Fig. 2-4)。赤色光による葉面積の 増加と生育量の増加は、レタスで報告されている(大嶋ら,2015)。また、ピーマン(枡田 ら,2004)、セントジョンーズワート(西村ら,2006)では、赤色光で他の光質と比較して 葉面積と生育の増加が観察されたが、光質間では光合成速度に差がなかったことを報告し ており、この結果は受光面積の増大が生育の増大の要因になっていることを示している。本 実験では、光合成速度の測定を行っていないため、光合成速度については議論できないが、 赤色光照射による葉の拡大は、生育の促進に繋がったと考えられた。 一方で、B 処理区で茎において anthocyanin 蓄積と考えられる色素の沈着があったことか ら、青色光が含まれる処理区区(FL、B、RB 処理区)では葉でも多くの anthocyanin が合成 されていると考えられた(Fig. 2-2)。これらの色素により葉に入射した光が吸収または反射 されてしまうことで光合成効率が低下することを大嶋ら(2015)は指摘しており、ニチニチ ソウでも同様の現象が起こっている可能性は否定できない。特に、本研究で用いている品種 の花色は、濃赤色でありanthocyaninを合成するポテンシャルは低くはないと推定される(Fig. 2-2)。 2. 異なる光質が vindoline および catharanthine 濃度および株あたり収量に及ぼす影響 本実験の結果から、可視光域の光質は vindoline と catharanthine 濃度にほとんど影響を与 えないことが明らかになった(Fig. 2-5)。そのため株あたりの vindoline および catharanthine 収量は、葉部新鮮重の結果が大きく反映された(Fig. 2-4)。また、R 処理区は試験区の中で 最も放射照度が低い試験区であることから、エネルギーあたりのアルカロイド生産効率が 最も高かった。そのため、vindoline および catharanthine 生産には赤単色での栽培が最も適し ていると考えられた。 3. 赤色光の光強度がニチニチソウの生育におよぼす影響 実験 2 処理 2 の結果から、ニチニチソウのバイオマスは PPFD に依存することが示され た(Fig. 2-10)。一方で、実験 2 処理1において地上部と葉部新鮮重が、PPFD の最も高い 600
μmol m-2 s-1に比べて 300 μmol m-2 s-1で高い結果になった(Fig. 2-8)。これは下位葉が黄化し
たことによる生理傷害よって引き起こされたと考えられた(Fig. 2-7)。高光量の赤色光によ る、葉の黄化・白化する現象はコムギ(Tripathy and Brown, 1995; Sood et al., 2004; Sood et al.
23
2005; Gupta et al., 2010)やイネ(Roy et al., 2013)の実生で起こることが知られている。Tripathy
and Brown(1995)は、この現象が chlorophyll 合成が阻害されることで起こることを報告し
ており、本研究でも同様の現象が生じたと考えられた。chlorophyll 合成阻害によって生じる
chlorophyll前駆体の protoporphyrin IX は、光を吸収することで一重項酸素を発生させる作用
がある(Tripathy et al., 2007)。この一重項酸素は、細胞膜を含む細胞小器官に対してダメー ジを与えるため、生育を抑制や葉の黄化・白化を引き起こす(Tripathy and Chakraborty, 1991;
Chakraborty and Tripathy, 1992)。加えて、Roy et al.(2013)は、PHY がこの反応に関わって
いることを報告している。 実験 2 処理 1 において 75 μmol m-2 s-1と 150 μmol m-2 s-1の新鮮重の結果も PPFD の増加に 従わなかった。実験 2 処理 2 において含水率が PPFD 依存的な減少傾向にあることから、 75 μmol m-2 s-1では含水率が高いため、新鮮重が 150 μmol m-2 s-1より大きい結果になった と考えられた。また、草丈が 75 μmol m-2 s-1で他の試験区と比較して一つだけ高いことか ら、光量不足による徒長が生じていると考えられる。 4. 赤色光の光強度が vindoline および catharanthine 濃度に与える影響
第 4 葉に含まれる vindoline および catharanthine 濃度は、150 μmol m-2 s-1から 350 μmol m-2
s-1の間では、PPFD 依存的な生育の増加とは反対に減少する傾向が確認された(Fig. 2-11 お
よび Fig. 2-13)。光学フィルターと蛍光灯を組み合わせて行ったセントジョーンズワートの
実験では、250 μmol m-2 s-1と 500 μmol m-2 s-1の試験区間で、光強度の増加に伴った生育の増
加と二次代謝物である hypericine、pseudoypericine、hyperforine 濃度の減少が確認された (Nishimura et al., 2007)。また、Cai et al.(2009)は、太陽光をネットで遮光することによ る、異なる光強度下で栽培した Rauvolfia 属でも、生育と二次代謝物の間にセントジョーン ズワートと同じ関係があったことを報告している。
Herms and Mattoson(1992)は、生育と植物の防御に関連する二次代謝産物はトレードオ
フの関係にあることを指摘している。このトレードオフ現象は遺伝子発現レベルから制御 されることが報告されている。CrStr 発現量は、vindoline および catharanthine 濃度の変化と およそ同じ傾向にあることから、CrStr 発現量が光強度を変化させたときのアルカロイド濃 度に強く影響している可能性が考えられた(Fig. 2-14 B)。CrStr は、vindoline および
catharanthine に共通の前駆体である strictosidine を合成する酵素をコードする遺伝子である
(Fig. 1-2)。そのため、光強度の変化が strictosidine 生合成量を変化させたことで、vindoline および catharanthine 濃度が同じ様に変動したと考えられる。
Yang et al.(2012)は、jasmonic acid シグナル伝達経路に関わる JAZ と gibberellin シグナ
ル伝達に関わる DELLA の相互作用が、生育と防御のバランスに関わっていることを報告し ている。他方で、CrDat および Orca3 の遺伝子発現は傷害応答を伝達する植物ホルモンの
jasmonic acidによって制御されている(van der Fits and Memelink, 2000)。高光量の処理区で
24
よる生育への応答が強くなり、jasmonic acid の応答が抑制されたと考えられた(Fig. 2-14 B および D)。
しかしながら、実験 2 処理 1 の 300 μmol m-2 s-1から 600 μmol m-2 s-1では、150 μmol m-2 s
-1から 350 μmol m-2 s-1に見られた生育とアルカロイド濃度が反対になる関係が見られなかっ
た。これは、高光量の赤色光で葉が黄化・白化する反応が、MIA 生合成経路に含まれる酵素 等にダメージを与えたためアルカロイド濃度を低下させたと考えられた。MIA 生合成は、 様々な細胞小器官に局在する酵素によって生合成が行われおり、特に MIA の初発物質であ る tryptophan および geraniol を生合成する shikimate および methylerythritol phosphate(MEP) 経路は、葉緑体に存在している(reviewed by Verma et al., 2012)。また、vindoline 生合成酵素 の一つである NMT も葉緑体に局在していることが知られている。そのため、これらの生合 成酵素は、protoporphyrin IX が発生させる一重項酸素による酸化の影響を最も受けやすく、 タンパク質の活性等が低下した可能性が考えられた。 光強度が高くなると炭素分配が地下部に優先される傾向があった(Fig. 2-12)。vindoline 生 合成酵素の一つである NMT は葉緑体に局在しているため、vindoline は地上部にのみ蓄積す る。そのため、地上部でアルカロイド生産に利用できる炭素量は光強度の増加により制限さ れた可能性が考えられた。 5. 光環境が vindoline と catharanthine 生産に与える影響 実験 1 で他の光質に比べて、R 処理区では生育量が大きいながら、vindoline および
catharanthine濃度が低下しなかった(Fig. 2-4 および Fig. 2-5)。この結果は、実験 2 でのバイ
オマスとアルカロイド濃度が反対の傾向を示す結果と矛盾する。この現象の原因には、二つ の可能性が考えられた。
1つ目は、青色光が入ることで catharanthine が FMN を介した酸化が起こり、分解または
他のアルカロイドに変換された可能性である。FMN は、370 nm と 450nm 付近に吸収ピーク を持っており、青色光でも vindoline と catharanthine のカップリング反応を促進させる(Hirata
et al., 1993)。UVA が FMN を介して catharanthine を酸化する反応において、酸化された
catharanthineは、一部が vindoline と結合することで 3‘, 4’-anhydrovinblastine に変換されるも
のの、多くが分解されることが報告されている(Hirata et al., 1999)。青色光でも、FMN を介 した酸化反応は UVA と同じ機構であると考えられている。実際に、vindoline/catharanthine 比 は、青色光が含まれる処理区では、青色光が含まれない処理区(R 処理区)と比較して
vindolineより相対的に catharanthine 量が少ない傾向があった(Fig. 2-5)。そのため、青色光
の照射はカップリング反応の促進と同時に catharanthine の分解を生じていると予想された。
2つ目は、赤色光によって MIA 生合成が促進された可能性である。MIA 生合成を制御す
る jasmonic acid のシグナリングに PHY が関与していることがシロイヌナズナ(Robson et al.,
2010;Chico et al., 2014)やミヤコグサ(Suzuki et al., 2011)で知られている。これらの研究
25 そのため、赤色光が含まれない、または赤色光の光量が少ない試験区では、不活性型の PHY が増加し、jasmonic acid のシグナリングが抑制されたと予想される。その結果、青色光が含 まれる試験区に比べて、赤色光が多く含まれる試験区では、vindoline と catharanthine 生合成 が促進された可能性がある。加えて、青色光が少ない試験区では anthocyanin 等の合成に利 用される炭素量が少ないため、アルカロイド合成に対して分配できる炭素量が、青色光が含 まれる試験区に比べて豊富であると考えられた。
26
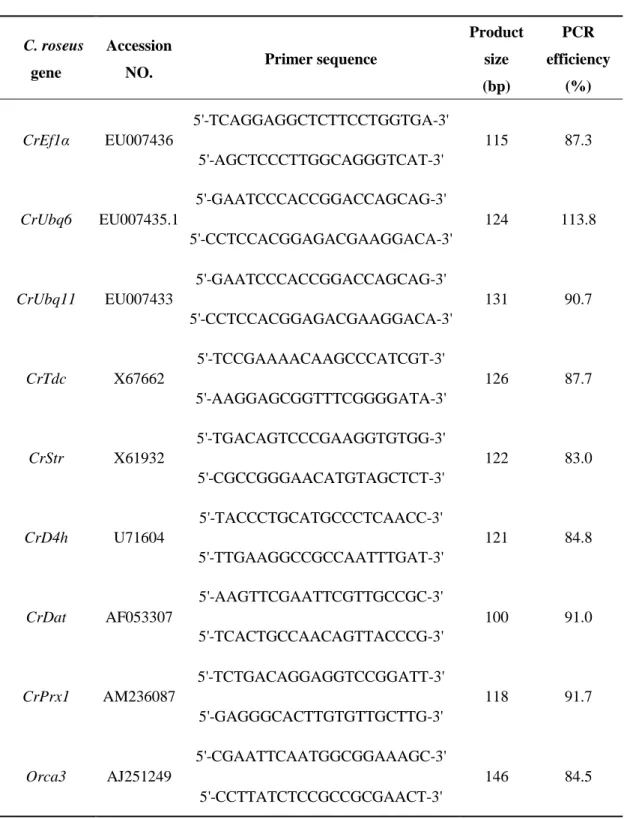
Table 2-1 Primers for qRT-PCR analyses of C. roseus reference and target genes
C. roseus
gene
Accession
NO. Primer sequence
Product size (bp) PCR efficiency (%) CrEf1α EU007436 5'-TCAGGAGGCTCTTCCTGGTGA-3' 115 87.3 5'-AGCTCCCTTGGCAGGGTCAT-3' CrUbq6 EU007435.1 5'-GAATCCCACCGGACCAGCAG-3' 124 113.8 5'-CCTCCACGGAGACGAAGGACA-3' CrUbq11 EU007433 5'-GAATCCCACCGGACCAGCAG-3' 131 90.7 5'-CCTCCACGGAGACGAAGGACA-3' CrTdc X67662 5'-TCCGAAAACAAGCCCATCGT-3' 126 87.7 5'-AAGGAGCGGTTTCGGGGATA-3' CrStr X61932 5'-TGACAGTCCCGAAGGTGTGG-3' 122 83.0 5'-CGCCGGGAACATGTAGCTCT-3' CrD4h U71604 5'-TACCCTGCATGCCCTCAACC-3' 121 84.8 5'-TTGAAGGCCGCCAATTTGAT-3' CrDat AF053307 5'-AAGTTCGAATTCGTTGCCGC-3' 100 91.0 5'-TCACTGCCAACAGTTACCCG-3' CrPrx1 AM236087 5'-TCTGACAGGAGGTCCGGATT-3' 118 91.7 5'-GAGGGCACTTGTGTTGCTTG-3' Orca3 AJ251249 5'-CGAATTCAATGGCGGAAAGC-3' 146 84.5 5'-CCTTATCTCCGCCGCGAACT-3'
27
Fig. 2-1 Spectral energy distribution of the each light sources and sunlight. Measurements are made using a spectroradiometer. Relative values are shown for each peak wavelength.
Fig. 2-2 Periwinkles at the harvest. The plants were grown under white fluorescent lamp (FL), monochromatic red (R) and blue (B), or mixture of red and blue (RB) light of PPFD at 150 µmol m-2
28
Fig. 2-3 Changes in plant height (A), maximum leaf length (B) and width (C) grown under different light treatments. Periwinkles were grown under white fluorescent lamp (■, FL), monochromatic red (●, R) and blue (▲, B), or mixture of red and blue (◆, RB) light at PPFD of 150 µmol m-2 s-1 for 28
days. Bars indicate the standard error (n = 9-10). Asterisks denote significant differences of each day according to analysis of variance (n = 9-10, ** p < 0.01, ***p < 0.001).
29
Fig. 2-4 Shoot fresh weight (left) and total leaf fresh weight (right) at the harvest. Periwinkles were grown under white fluorescent lamp (FL), monochromatic red (R) and blue (B), or mixture of red and blue (RB) light at PPFD of 150 µmol m-2 s-1 for 28 days. Bars indicate the standard error (n = 9-10).
30
Fig. 2-5 Vindoline (A) and catharanthine (B) concentrations in total fresh leaf and vindoline/catharanthine ratio (C) at the harvest. Periwinkles were grown under white fluorescent lamp (FL), monochromatic red (R) and blue (B), or mixture of red and blue (RB) light at PPFD of 150 µmol m-2 s-1 for 28 days. Bars indicate the standard error (n = 3). Means with different letters are significantly
31
Fig. 2-6 Vindoline (A) and catharanthine (B), total alkaloid yields (C) per plant at the harvest. Periwinkles were grown under white fluorescent lamp (FL), red light alone (R), blue light alone (B), or mixture of red and blue light (RB) at PPFD of 150 µmol m-2 s-1 for 28 days. Bars indicate the
standard error (n = 3). Means with different letters are significantly different at the 5 % level by Tukey’s HSD test.
32
Fig. 2-7 C. roseus at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 75 (A), 150 (B), 300 (C), 600 (D) μmol m-2 s-1. Black bar
33
Fig. 2-8 Plant height (A), fresh weights of shoot (B) and total leaf (C) at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 75, 150, 300, 600 μmol m-2 s-1. The bars indicate the standard error (n = 6). Means with different letters within each
34
Fig. 2-9 Vindoline (left) and catharanthine (right) concentrations in the fourth leaves at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 75, 150, 300, 600 μmol m-2 s-1. The bars indicate the standard error (n = 3). Means with different letters
35
Fig. 2-10 Plant height (A), fresh weights of shoot (B) and total leaf (C) at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 150, 250, 300, 350 μmol m-2 s-1. The bars indicate the standard error (n = 5~15). Means with different letters
36
Fig. 2-11 Dry weights of total leaf (A), root (B), shoot (C), and plant (D) at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 150, 250, 300, 350 μmol m-2 s-1. The bars indicate the standard error (n = 5). Means with different letters within
37
Fig. 2-12 Shoot/root ratio (left), and water content (right) at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 150, 250, 300, 350 μmol m-2 s-1. The bars indicate the standard error (n = 5). Means with different letters within each
panel are significantly different at the 5 % level by Tukey’s HSD test.
Fig. 2-13 Vindoline (left) and catharanthine (right) concentrations in the fourth leaves at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 150, 250, 300, 350 μmol m-2 s-1. The bars indicate the standard error (n = 5). Means with
38
Fig. 2-14 CrTdc (A), CrStr (B), CrD4h (C), CrDat (D), Orca3 (E) expressions in the fourth leaves at 63 days after germination. The plants were grown for 28 days under four different monochromatic red light PPFDs: 150, 250, 300, 350 μmol m-2 s-1. The bars indicate the standard error (n = 3). Means
39
第3章 赤色光下で生育したニチニチソウに対する紫外線 A 領域および青色光
照射が vinblastine 生産に与える影響
第1節 序論 第 2 章の結論として、赤単色でのニチニチソウの栽培は、他の光質に比べてバイオマスの 増加に有効でかつ、アルカロイド濃度を低下させないことを示した。そこで本章では、Hirata et al.(1991; 1992; 1993)が報告する UVA または青色光照射によって促進される非酵素的な カップリング反応を赤色光下で栽培したニチニチソウの葉内で起こすことで vinblastine 蓄 積させることが可能か調査を行った。 UVA や青色光のような比較的エネルギーレベルの高い光を細胞が受けると、細胞内にはROSなどのさまざまな酸化物が生じる(Baier et al., 2006)。細胞内での恒常性の維持には、
酸化物の消去を行うことが重要な機能の一つとして知られている。その機能を担う酵素の 一つに ascorbic acid を酸化させて過酸化水素消去を行う ascorbate peroxidase 1(Apx1)が存 在する。Davletova et al.(2005)は、シロイヌナズナにおいて光ストレスが生じると Apx1 発 現量が高まることを報告しており、Apx1 発現量を増やすことでこのストレスに対応してい ると考えられている。vindoline と catharanthine のカップリング反応を触媒する酵素 CrPRX1 は、APX1 と同じペルオキシダーゼであり、過酸化水素を分解する作用がある。エネルギー レベルの低い R で栽培されたニチニチソウに UVA や青色光を照射することは、突然エネル ギーレベルの高い光にさらされることになり、酸化状態の急激な上昇が生じると予想され る。この時、CrPrx1 発現量を高めることで、Apx1 と同様に酸化状態の急激な上昇に対応す る可能性が予想される。Costa et al.(2008)は、CrPrx1 発現量および活性の増加は、3‘, 4’-anhydrovinblastine 量の増加を起こすことを報告していることから、UVA や青色光照射によ って CrPrx1 発現量増加にともなう二量体 MIA 量の増加が期待できる。 植物の二次代謝産物は、いくつかのマスター制御転写因子が複数の生合成酵素遺伝子の 発現制御を行っており、生合成が促進されるような刺激が加えられると代謝経路上の生合 成酵素遺伝子の発現量が同時に増加することが知られている(Gonzalez et al., 2008;Hirai et
al., 2007;Kato et al., 2007;Shoji et al., 2010;Yamada et al., 2012)。ニチニチソウでもマスタ
ー制御転写因子として Orca2(Menke et al., 1999)、Orca3(van der Fits and Memelink, 2000)、
Bis1(Van Moerkercke et al., 2015)が確認されている。そのため、カップリング反応発生時に
マスター制御転写因子が活性化することで単量体 MIA 生合成も高まっている可能性がある。 しかし、UVA や青色光照射行ったときの生合成関連遺伝子発現量を測定した研究例は過去 にない。そこで、本章の実験ではアルカロイド濃度と共にアルカロイド生合成関連遺伝子の 発現量を調査した。
40 第2節 材料および方法 1. 材料および育苗条件 材料は第 2 章と同じ品種を用いた。育苗条件は第 2 章実験 2 と同じにした。 2. 赤色光照射条件 播種 35 日後、150 μmol m-2 s-1の赤単色光に設定した DFT に移植した。光周期、DFT の水 耕液および室温は育苗と同じ条件とした。播種 63 日後、異なる 3 つの光処理区にそれぞれ 32株移植した。 3. UVAまたは青色光照射条件
光処理区は、150 μmol m-2 s-1の青単色区(B)、5 W m-2の UVA に 150 μmol m-2 s-1の赤色
光を補光した区(UVA+R)、コントロールの 150 μmol m-2 s-1の赤単色区(R)とした。可視 光は、第 2 章で用いた LED を使用した。UVA の光源はウシオ電機(株)から提供された UV 蛍光灯(XEFL340、ピーク波長 370 nm 付近)を用いた。UV 蛍光灯は 320 mm 間隔で設置し た。UVA 強度は UV 放射強度計(x9-6, Gigahertz-Optik GmbH)を用いて測定した。光周期、 DFTの水耕液および室温は育苗と同じ条件とした。 4. 収穫および生育調査 播種 63 日後の第 4 葉対および全葉を処理 0 日後サンプルとして、それぞれ異なる株から 4株ずつ収穫した。その後、播種 64、66、68、70 日後の株から第 4 葉対および全葉を、処 理 1、3、5、7 日後としてそれぞれ異なる株から 4 株ずつ収穫した。 収穫後、株あたり地上部新鮮重および全葉新鮮重を測定した。測定後、第 4 葉および全葉 をビニルパックに入れ−35 oCで保存した。その後、凍結した全葉は 48 時間凍結乾燥し、こ れを株あたり全葉乾物重とした。 5. アルカロイド抽出 アルカロイドの抽出は第 2 章実験 2 処理2の方法で行った。 6. HPLC条件 HPLC システムは Chromaster((株)日立ハイテクノロジーズ)を用いた。カラムは CapcellpacC18 MGII(4.6 mm× 150 mm,5 μm,(株)資生堂)を用いた。注入量は 10 µl、 流速は 1.0 ml min-1、カラムオーブン温度は 40 oCに設定した。アルカロイドはダイオードア レイで検出し、測定波長は 250 nm に設定した。移動層は 0.01 M リン酸緩衝液(pH7.0,溶 媒 A)とアセトニトリル(溶媒 B)を用いた。グラジエント条件(溶媒 A の容積:溶媒 B の 容積)は 0-3 min 70 : 30、3-8 min 40 : 60、8-20 min 40 : 60 とした。vindloine および catharanthine の同定と定量は、vindoline と catharanthine sulfate 標準品(ともに LKT Labolatries, Inc.より購