Morphological and gene expression changes in
Arabidopsis thaliana under clinorotated
conditions;with special reference to WRKY46
著者
Soh Hyuncheol
学位授与機関
Tohoku University
学位授与番号
11301乙第9408号
Dissertation
Morphological and gene expression changes in
Arabidopsis thaliana under clinorotated conditions;
with special reference to WRKY46
Hyuncheol Soh
i
Contents
Summary
………...vAcknowledgments
………...viiGeneral introduction and objectives of this study
………..1I. Importance of microgravity simulators...………...…….………..2
I.i. 3-D clinorotation for the study of plant growth and development in simulated microgravity...2
I.ii. Necessity of long-term experiments under 3-D clinorotation conditions……...5
II. Comprehensive studies on gene expressions………...6
III. In silico analysis of Arabidopsis promoter sequence………...7
IV. WRKY protein in Arabidopsis thaliana………....8
V. Analysis of T-DNA inserted mutant ...………..9
VI. Objectives of this study………...9
VII. Literature cited...28
Chapter 1 Gene expression changes in Arabidopsis
seedlings during short- to long-term exposure to 3-D
clinorotation
………...34ii
1.1. Abstract………...35
1.2. Introduction………...35
1.3. Materials and Methods………..37
1.3.1. Plant materials and exposure to 3-D clinorotation………...37
1.3.2. RNA preparation, probe labeling, and microarray hybridization...38
1.3.3. Acquisition and analysis of microarray data...39
1.3.4. Quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR)...40
1.4. Results………....41
1.4.1. Changes in gene expressions under 3-D clinorotation and their classification by gene ontology (GO)...41
1.4.2. Gene expression validated by real-time quantitative PCR...43
1.5. Discussion………..45
1.6. Literature cited...99
Chapter 2 Identification of unique cis-element pattern on
3-D clinorotation treated Arabidopsis by in silico and gene
expression
………...1052.1. Abstract……….106
2.2. Introduction………...106
2.3. Materials and Methods………...109
iii
2.3.2. RNA preparation and qRT-PCR...……...109
2.3.3. In silico analysis of cis-elements...………....110
2.4. Results………..110
2.4.1. Genes highly expressed under 3-D clinorotation...………...110
2.4.2. WRKY46 expression responsive to 3-D clinorotation...111
2.4.3. cis-element analysis of the promoter regions of 40 selected genes...112
2.5. Discussion……….114
2.6. Literature cited...129
Chapter 3 Morphological and gene expression pattern
changes in wrky46 mutant Arabidopsis thaliana under
altered gravity conditions
………...………...1343.1. Abstract………...135
3.2. Introduction………...135
3.3. Materials and Methods………...139
3.3.1. Plant materials and altered gravity treatment………..139
3.3.2. Root cap amyloplast starch staining and light microscopy observation ……….139
3.3.3. RNA preparation and quantitative real-time PCR (qRT-PCR)………..139
3.3.4. T-DNA insertion confirmation……….140
3.4. Results………..140 3.4.1. Confirmation of T-DNA insertion in the wrky46 mutants and their
iv
phenotypic difference under 3-D clinorotation treatment………...140
3.4.2. The phenotypes of WT and wrky46 identical under normal gravity ………...141
3.4.3. Expression patterns of genes related to polar auxin transport and auxin response...141
3.4.4. Root gravitropism of wrky46 mutant and WT seedlings.…………...142
3.4.5. Amyloplast morphology in root cap cells of mutant and WT seedlings grown vertically...143
3.5. Discussion………143
3.6. Literature cited...159
Conclusion and future perspectives
...164v
Summary
Gravity is an important environmental factor that controls plant growth and development. However, the mechanistic aspect of plant response to gravity is far from being fully understood at present. Microgravity conditions are now available by utilizing space environment, but spaceflight opportunities for biological research are very limited. On the ground, clinorotation facilities have been used for simulating plant growth in microgravity. Clinorotation experiments could bring about novel results and hypothesis that should be ultimately verified under ‘real microgravity’ conditions in space. In this study, I examined morphology and gene expression changes of
Arabidopsis thaliana seedlings under short-term (1 hour) and long-term (1 to 6 days)
3-D clinorotation.
To understand the 3-D clinorotation effect on the gene expression pattern of
Arabidopsis roots, using 44k Arabidopsis microarray (Agilent) I analyzed transcript
abundance of genes in seedlings 1 h and 6 days after the start of 3-D clinorotation. The microarray data analysis showed that Arabidopsis seedlings respond to 3-D clinorotation by dynamic up- and down-regulations of genes. Gene ontology and qRT-PCR analyses of the genes investigated showed their expression profiles and patterns similar to those observed in stress-challenged plants. To find genes specifically responsive to 3-D clinorotation, I selected 10 genes that were highly expressed under short-term (1 h) 3-D clinorotation, and their expression patterns were validated by qRT-PCR analysis. Nine out of the 10 genes were up-regulated due to not only clinorotation but also touch and/or wounding in 3-week-old Arabidopsis seedlings. However,
WRKY46 expression was elevated only by 3-D clinorotation. Also, to clarify the
characteristics of the genes expressed at high level in response to 3-D clinorotation, 20 cis-elements in the promoters of the 40 selected genes, including the 10 3-D clinorotation-inducible genes, six WRKY genes, and abiotic stress-inducible genes, were analyzed, and their spatial positions on each promoter were determined. Four
vi
TATABOX5, and POLASIG1) showed a unique spatial arrangement in most 3-D clinorotation-inducible genes including WRKY46. It was suggested that the M/T-G-T-P cis-element pattern identified in the promoter of WRKY46 could play a role in regulating gene expression due to 3-D clinorotation.
To understand the role of the WRKY46 gene in morphological change under clinorotated conditions, the responses of wrky46 knockout mutants, constructed by T-DNA insertion, to 3-D clinorotation and gravistimulation by reorientation were investigated. Morphological changes in roots, such as multidirectional growth patterns, were observed in the wild type (WT) seedlings from day 3 to day 4 after the start of 3-D clinorotation, while in the wrky46 mutant seedlings the changes observed were minimal. Interestingly, the expressions of genes involved in auxin transport, such as
AUX1, PIN2, PIN3, PIN4, PIN7, and ARG1, appeared to be lower in WT than wrky46
mutant seedlings under 3-D clinorotation. In addition, the gravitropic curvature of
wrky46 roots at 10 to 15 h following gravistimulation was smaller than that of WT, and
it became the same in WT and wrky46 by 20 h after gravistimulation. Because auxin-resistance genes AXRs are known to play a role in gravitropic bending of roots, I analyzed their expressions. The results showed that AXR1, AXR3, and AXR4 tended to be higher in WT seedlings compared to those in wrky46 mutant seedlings after gravistimulation.
Thus, this study showed that roots of Arabidopsis seedlings display multiple-directional growth together with coiling under 3-D clinorotated conditions. These morphological changes due to clinorotation could result from loss of gravitropic response, phenocopying root growth in microgravity. Analyses of gene expression with microarray and qRT-PCR identified a clinorotation-responsive gene WRKY46 that could play a role in the clinorotaion-inducible changes of root growth pattern. The results of functional analysis of WRKY46 suggested its involvement in root bending by gravitropic response or by multi-directional growth during 3-D clinorotation, probably via regulating auxin transport and/or auxin response.
vii
Acknowledgments
It would not have been possible to write this doctoral dissertation without the help and support of many people around me, to only some of whom it is possible to acknowledge here. During the completion of this dissertation, I have deepened true studies of my life as well as academic achievement. Especially it was the greatest teaching in my life that I learned patience and courage to finish without any frustration. First and foremost, I would like to express my deep and sincere gratitude to Professor Sukchan Lee, Department of Genetic Engineering, Sungkyunkwan University, who kindly hosted me in his lab and provided me with all the equipment and chemicals for performing the experiments described in chapter 1, 2 and 3. He also graciously guided me and provided a lot of support.
I wish to express my respect and sincere thanks to Late Professor Atsushi Komamine, who enabling me to continue my research. My sincere thanks are also to Dr. Krishnamoorthy Srikanth, my brother and my best friend at Chonbuk National University, who helped me in drafting manuscripts and also in the drafting of this dissertation.
Thanks, are also due for Professor Kyeongsik Han; Woosuk university for providing me with the 3-D clinorotation equipment, Professor Sung Soo Whang; Chonbuk National University for kindly taking me in his lab and for providing me with facilities the experiments described in chapter 3, Professor Chungkyun Auh; Mokpo National University, for kindly providing the fund for performing the microarray experiments, Yongsang Choi; Chonbuk National University, for supporting me in performing the experiments described in chapter 2, Professor Emeritus Woong-Young Soh; Chonbuk National University for providing expert opinion throughout my study, Dr Yong Rhee, Sungkyunkwan University who helped me draft the microarray paper. And finally, Professor Donggiun Kim; Shilla University, Dr Taek-Kyun Lee; Korea Ocean Research and Development Institute, Professor Up-Dong Yeo; Chonbuk
viii
National University for providing various lab facilities during my study.
I wish to express special gratitude to the reverend of Wonbuddism Hang-Sim Choi, who is family to me and helped me overcome many difficult moments with the power of pray. I also acknowledge and express my gratitude to Dr Jinsang Yoo for constantly encouraging me during this journey.
Lastly, I give my infinite gratitude to my parents, Professor Emeritus Woong-Young Soh and Emeritus Professor Duckyee Cho for watching their son's research with endless love and patience for a long period of time. Without their patience, love and support, I would never have reached this position. I would like to dedicate this dissertation to my beloved grandmother in heaven who always cared for me and showered endless love.
1
2
I. Importance of microgravity simulators
Gravity has a significant effect on plant growth and development. In recent decades, numerous experiments have been performed with different types of gravity simulators to help understand gravity-dependent phenomena of various plants (1, 2, 3, 4).
Research under the conditions of microgravity during space missions has contributed to advancing our knowledge of the impact of gravity on biological processes, gravity sensing mechanisms, and gravity-mediated orientation of organisms in their spatial environment (5, 6). These processes, however, are far from being fully understood because of limited access to flight opportunities and insufficient experiments. Therefore, few studies have conducted experiments on ground by simulating microgravity conditions and have compared results with the ‘‘real’’ microgravity achieved using spaceflight mission (7).
Using experimental platforms such as two-dimensional (2-D) clinostats, and random positioning machines (RPMs), and so on, researchers mainly use the term ‘‘simulated microgravity’’ or ‘‘simulated weightlessness’’ to describe the state of acceleration, which is assumed to be achieved with such machines (8). The pictures of 2-D clinostats and random positioning machines (RPMs) are shown in Fig. I-i.
Even though the term ‘‘microgravity’’ (‘‘micro-g environment,’’ or ‘‘μg’’) is frequently used as a synonym of ‘‘weightlessness’’ or ‘‘zero-g,’’ the former should be distinguished from the latter. While the g-forces in simulators are not actually zero but just ‘‘very small”, “true” weightlessness for more than a few seconds, can be achieved only in space. In general, terms such as ‘‘weightlessness’’ or ‘‘zero-g’’ should be avoided since one can never achieve such a level for more than a few seconds with ground-based simulations (7).
3
microgravity
Real microgravity conditions can be produced by free fall parabolic flight and by sounding rockets sent to a lower earth orbit. However, the microgravity conditions achieved by these methods are generally too brief to induce obvious changes in plant growth and morphogenesis (9). Indeed, the best way to study plant growth and development under microgravity is to conduct experiments in space. However, access to flight opportunities is scarce, and a series of succeeding experiments has been realized only sporadically (7). Thus, alternatives to simulate microgravity conditions on earth are necessary for studying plant growth and development.
Various ground-based simulators have been constructed and used to simulate microgravity including simulators, unidirectional clinostats, random positioning machines (RPMs), rotating wall vessels and diamagnetic levitation instruments. Among them, 3-D clinostats have been developed in Japan and the Netherlands (10, 11). The 3-D clinostat is based on the principle of changing the direction of gravity with respect to the sample mounted in the rotation axis, and has two independently rotating frames running at constant speed and constant direction. When two motors are rotated at different constant rates, the position of plants is repeatedly reversed with respect to the gravity vector and the plants cannot detect the direction of the gravity vector. Consequently, the effect of gravity on plants growth is in essence eliminated.
The growth and development under microgravity or under 3-D clinorotation has been named automorphosis or automorphogenesis (10, 12, 13). One of the most important phenomena observed in treatment by 3-D clinostat is the position of the nucleus in statocyte. In microgravity conditions, the amyloplasts are closer to the center of the statocytes, than in the statocytes differentiated at 1G (8). As a result, clinorotation leads to a difference in amyloplast distribution.
Previous studies have reported that the change in position of amylopast under 3-D clinostat was similar to changes observed under real microgravity. A 3-D cell
4
reconstruction done by Smith et al (14) demonstrated that the distribution of the amyloplast was not random in statocytes of Trifolium repens in microgravity. They were grouped near the center of the cells under clinostat, but were scattered under microgravity (0 G) (14) (Fig. I-ii). Kraft et al. (15) reported that position of amyloplast assayed in columella cells in the roots of Arabidopsis thaliana seedlings were measured under the conditions of 2-D clinostat, 3-D clinostat, and in space (true microgravity). In their results, the amyloplast position was not significantly different between spaceflight and RPM conditions but was significantly different between spaceflight and 2-D clinostat (Fig. I-iii).
Plants are dependent on the gravity vector during vegetative growth, in stages such as morphogenesis, direction of organ growth of Pisum, Vigna and Zea, peg formation of Cucumis, leaf senescence of Avena, are shown to be influenced by rotation on the 3-D clinostat in comparison with 1G controls (1, 2, 16, 17, 18). However, the rate of germination of Pisum, Vigna and Oryza and growth correlation among organs of Pisum and Oryza are not influenced by clinorotation (8). The peg formation in cucumber hypocotyl under 3-D clinorotation is changed; that is, the proportion of seedlings without a peg or with two pegs increases in comparison with normal hypocotyl with one peg (1, 19) suggesting that gravity vector also has an effect on polar auxin transport.
When plants are continuously treated with 3-D clinorotation for a long time, they cannot detect the direction of gravity vector. This response is similar to the phenomena observed in microgravity conditions, and 3-D clinorotation strongly reduces the activity of polar auxin transport (12, 20).
Studies have suggested that polar auxin transport is substantially affected by long exposure (6 days) to 3-D clinorotation (20, 21). The responses of higher plants to microgravity conditions seem to be different not only among the species of plants but also in their organs and/or tissues (20). Therefore, there is a necessity for more information from a variety of plants to understand the growth and development of plants under altered gravity, 3-D clinorotation. Table I-i presents a compilation of important studies conducted on the response of plants under altered gravity over the
5
years.
I.ii. Necessity of long-term experiments under 3-D clinorotation conditions
Plants have evolved under the gravitational force on earth. During the evolution process, plants have acquired the ability to use gravity in regulating their growth and development. Therefore, understanding the role of gravity on developmental processes in the life cycle of plants is important for devising mechanism to grown them in space (22).
Gravitropic curvature response occurs within short time from gravity perception through transduction and transmission around 10 minutes (4, 23, 24, 25). Gravitropic response can broadly be grouped under four phases: a) the perception of the stimulus. b) the transduction of this stimulus c) the transmission of this stimulus, and d) the differential growth of the upper and lower sides of the organ (26) (Fig. I-iv).
Although several studies state that, plants adapt to the absence of gravity, the delay in seed development in orbit makes it difficult to state that we fully understand the adaptation of plants to microgravity, because high seed production, is considered to be the major factor in plant adaptation to new conditions. (27).
Plants response to microgravity can be classified into two phases, short term and long term. When higher plants are exposed to short term (seconds to hours) microgravity, such as onboard parabolic flights and sounding rockets, their cells usually exhibit abiotic stress responses. For example, Ca2+-, lipid-, and pH- signaling are rapidly enhanced, and then the production of reactive oxygen species and other radicals increase dramatically along with changes in metabolism and auxin signaling (28, 29, 30). Under long-term (days to months) microgravity exposure, plants acclimatize to the stress by changing their metabolism and oxidative response and by enhancing other tropic responses (31).
6
During spaceflight the living organisms should stay in microgravity or near zero gravity conditions for a long period. Thus, long-term exposure of plants under 3-D clinorotation is desirable in ground-based experiments to obtain results comparable to the information from real spaceflight and to draw generalization about plant gene response to the space environment. Therefore, a systematic analysis of regulatory networks at the molecular level of plants is needed to understand the acclimatization and adaptation of plants to microgravity. Long term, uninterrupted, multigenerational, plant experiments under microgravity will provide us with the information needed for cultivating cereals and vegetables on board the International Space Station, moon bases and other microgravity environments (3, 25, 27, 31).
Most of the ground-based research has been conducted under short-term exposure of plants to microgravity conditions, except one case under 3-D clinorotation (27, 28, 31, 36, 40). It is supposed that the response of plants to long-term exposure under 3-D clinorotation is different from that of the short-term exposure. Therefore, long-term studies with clinorotation are necessary to understand the mechanism of plant adaptation to long-term spaceflight at cellular and molecular level (27).
II. Comprehensive studies on gene expressions
Gene expression studies analysis using microarray have enabled the discovery of many genes involved in stress response and tolerance. DNA chip-based technology are arrays with cDNA sequences on a glass slide at a density > 1,000 genes/cm2. These arrayed sequences are hybridized simultaneously to a two-color fluorescently labelled cDNA probe pair prepared from RNA samples of different cell or tissue types, allowing direct and large-scale comparative analysis of gene expression. This technology was first demonstrated by analyzing 48 Arabidopsis genes for differential expression in roots and shoots (32). Microarrays were used to study 1,000 randomly chosen clones from a human cDNA library for identification of novel genes responding to heat shock and protein kinase C activation (33). In another study,
7
expression profiles of inflammatory disease-related genes were analyzed under various induction conditions by this chip-based method (34). Furthermore, the yeast genome comprising more than 6,000 coding sequences has been analyzed for dynamic expression by the use of microarrays (35, 36).
Many studies have reported that use of microarrays in understanding global gene expression. Using Arabidopsis full-length cDNA microarray, has several advantages. It is easier to do further functional analysis by isolating full length cDNAs and then comparing the Arabidopsis full-length cDNA sequences with the Arabidopsis genomic sequences, the promoter sequences and cis-acting elements of each microgravity induced gene can be studied (37, 38).
III. In silico analysis of Arabidopsis promoter sequence
Transcription factors (TFs) control the expression of tens or hundreds of target genes (39). While the similarity in gene expression patterns can be used to infer modular gene programs (or regulatory networks), identifying cis-regulatory elements requires integrating gene expression and sequence data. These functional elements are responsible for the timing and location of transcriptional activity (40, 41).
Gene expression studies using microarray data have shown that genes showing similar expression pattern are generally enriched for similar functions. Transcriptional regulation controls genes that respond to diverse biological processes, including abiotic stress responses, hormone responses, and developmental processes. TFs are Sequence-specific DNA-binding elements that control or influence many biological processes such as growth, cell division and responses to environmental stimuli. By activating or repressing the transcription of multiple target genes, they have a significant effect on metabolism and cellular response to environmental stress (42).
Stress-responsive genes can be regulated by the interactions between cis-regulatory elements and TFs that bind to these elements in the promoter regions of
8
these genes. Therefore, cis-element analysis of newly isolated stress responsive genes can give some insight into their putative function. Previous studies have only reported the location and number of cis-elements, it is necessary to supplement this information with information on the effect of the position and pattern of cis-element on the spatiotemporal expression of genes (42).
In silico promoter analysis can enhance our understanding of gene expression
in plants exposed to various physiological stresses. Characterization of highly conserved regulatory elements in promoters may help to elucidate the regulatory interactions between distinct cis-elements and trans-factors in response to stress. Critical analysis of cis-element organization combined with in silico analysis might allow for the prediction of transcriptional control and a more defined genetic engineering.
IV. WRKY protein in Arabidopsis thaliana
WRKY transcription factors are one of the largest families of transcriptional regulators in plants and modulate many plant processes (43). The name of the WRKY family is derived from a 60 amino-acid long region (WRKY domain) that is highly conserved amongst family members. The WRKY domain is defined by the conserved amino acid sequence WRKYGQK at its N-terminal, together with a novel zinc-finger-like motif (44) (Fig. IV-i, IV-ii). The clear binding preference of all characterized WRKY proteins for the same DNA motif suggests that it must constitute a DNA-binding domain. Recent studies have shown that an isolated WRKY domain has sequence-specific DNA-binding activity.
All known WRKY proteins contain either one or two WRKY domains and are classified on the basis of the number and feature of the WRKY domains. WRKY proteins with two WRKY domains belong to group I, whereas those with one WRKY domain belong to either group II or group III (43).
9
Many WRKY proteins regulates response to pathogen infection and other environmental stresses. WRKY proteins also regulates secondary-response genes, whose products carry out protective and defensive reactions (43, 44).
The two WRKY domains of group I members appear to be functionally distinct. Table IV-i lists all identified WRKY family genes in Arabidopsis thaliana. The name, locus ID, group, and relation to stresses and functions of the WRKY genes are indicated through a literature survey (43, 44).
V. Analysis of T-DNA inserted mutant
Insertional mutagenesis is a method for disrupting gene function and is based on the insertion of a foreign DNA into the gene of interest. In Arabidopsis, this involves the use of either transposable elements or T-DNA (45, 46, 47) which not only disrupts the expression of the gene into which it is inserted but also acts as a marker for subsequent identification of the mutation.
Polymerase chain reaction (PCR) allows one to easily isolate individual plants that carry a particular DNA mutation of interest (48, 49). An advantage of using T-DNAs as the insertional mutagen, as opposed to transposons (50), is that T-DNA insertions will not transpose subsequent to integration within the genome and the host cells are therefore chemically and physically stable through multiple generations.
VI. Objectives of this study
The main objective of this study was to identify global transcriptome response to 3-D clinorotation in Arabidopsis seedlings. In chapter 1, I studied the effect of short to long-term (from 1 h to 6 days) exposure of Arabidopsis to 3-D clinorotation. I
10
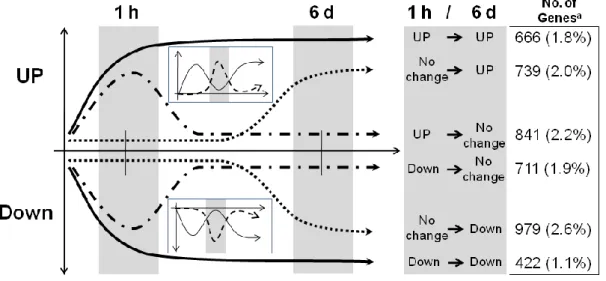
investigated the time-course expression patterns of genes significantly differentially expressed (DE) (over |2| fold change & p < 0.05) under 3-D clinorotation using microarray. Subsequently in chapter 2, the spatial orientation, numbers, and sequential orders of cis-elements in the promoter regions of ten genes that showed ten folds increase in expression under 3-D clinorotation were characterized. The response of these ten genes to other abiotic stresses such as drought, touch and wound was analyzed and WRKY46 was found to be highly induced under 3-D clinorotation. In chapter 3, detailed functional study of WRKY46 under 3-D clinorotation was carried out using T-DNA inserted Arabidopsis mutant (wrky46) to better understand the role of WRKY46 under altered gravity conditions. The phenotypical differences of these plants under 3-D clinorotation and gravistimulation by reorientation were studied, and the expression patterns of genes that play an important role in auxin transport and root gravitropism were analyzed to explore if
11
Fig. I-i A. 2-D clinostats. B. Random positioning machines (RPMs). The panels A and B were reproduced with permission from Herranz et al. (7) with modification.
12
Fig. I-ii Comparison of the position of the amyloplasts in the various gravity condition. Representative medial longitudinal section through clover root caps. (a) ground control (b) microgravity (c) after 3 days clinorotation (d) after 2 days of growth in 1G followed by a third day of clinorotation. Scale bar = 25 µm. This figure was reproduced from Smith et al. (14) with permission.
13
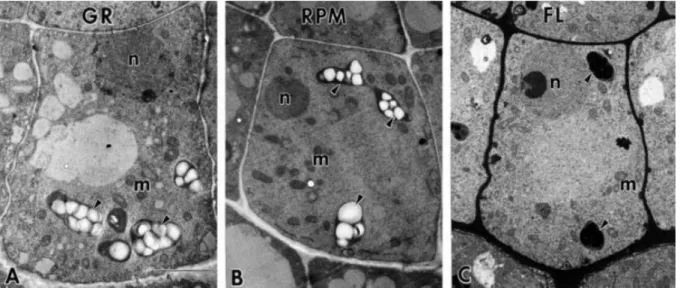
Fig. I-iii Electron micrographs of central columella cells of Arabidopsis seedlings. (GR; A) on the ground condition, (RPM; B) on the RPM (random positioning machine refers 3-D clinorotation), and (FL; C) on the spaceflight. Arrowhead (amyloplast) indicates that sedimented toward the distal cell wall in the ground condition and dispersed in the RPM (3-D clinorotation) and FL (spaceflight). This figure was reproduced from Kraft et al. (15) with permission.
14
Fig. I-iv The different phases of the gravitropic curvature of the root. Four phases are generally distinguished. The perception is the physical phase of the gravitropic reaction and corresponds to the movement of the statoliths in the gravisensing cells located in the root cap. It is followed by the transduction of the stimulus, I, e., the transformation of the mechanical effect of gravity into a biochemical factor. Both phases occur within the gravisensing cells. The transmission of gravistimulus to the reaction zone consists in an asymmetrical hormonal message (downward transport of auxin). It is responsible for a differential growth (curvature) that occurs far away from the perception zone. This figure was modified from Boonsirichai et al. (26) with permission.
0 s
1 s
10 s
10 min
Perception
Transduction
Transmission
Curvature
amyloplast
15
Fig. IV-i The WRKY domain. (a) The WRKY domain consensus for each WRKY subfamily in higher plants. The WRKY motif is highlighted in yellow and the cysteines and histidines that form the zinc finger are shown in blue. The four b-strands are shown in red. I CT and I NT denote the N-terminal and C-terminal WRKY domains from Group I WRKY proteins. (b) Two views of a spacefill structural model of the C-terminal WRKY domain from AtWRKY4. This figure was reproduced from Rushton et al. (43) with permission.
16
Fig. IV-ii Comparison of WRKY domain sequences from AtWRKY proteins. Gaps (dots) have been inserted for optimal alignment. Residues that are highly conserved within each of the major groups are in red and potential zinc ligands are highlighted in black boxes. For each (sub) group, the position of a conserved intron is indicated by an arrowhead. This figure was reproduced from Eulgem et al. (44) with permission.
17
Table I-i A compilation of important studies conducted to understand the response of various plants to altered gravity conditions. Listed are the type of experiment, plant and plant organ used, important finding and the year of publication.
Types of experiment Plant materials Reported results References
microgravity
(spaceflight, sounding rocket, parabolic)
mutant and WT of
Arabidopsis seedlings growth process observation Kiss et al. 1999
etiolated pea epicotyls growth process observation and
Ueda et al. 1999 and maize coleoptiles analysis of polar auxin transport
etiolated pea epicotyls growth process observation and
Ueda et al. 2000 and maize coleoptiles analysis of polar auxin transport
Arabidopsis seedlings
growth process observation and
Centis-Aubay et al. 2003 gene expression investigation
mutant and WT of
root waving assay Piconese et al. 2003
Arabidopsis seedlings
18
analysis of polar auxin transport
thalli of Chara globularis ovservation of statoliths positioning Limbach et al. 2005
Arabidopsis seedlings
and plants
plant cultivation and gene expression
patterns investigation by microarray Paul et al. 2005
Arabidopsis seedlings plant cultivation and gene expression
patterns investigation by microarray Paul et al. 2005 and plants
wheat
growth process observation and
Stutte et al. 2006 gene expression investigation
Ceratopteris richardii
gene expression patterns investigation
Salmi and Loux. 2008 by microarray
Arabidopsis seedlings growth process observation Millar et al. 2010
Arabidopsis seedlings
growth process observation and
Kwon et al. 2015 gene expression investigation by
19
3-D clinorotation
(random positioning machines)
cress, pea ,azuki bean,
growth process observation Hoson et al. 1992 caryopses of maize, rice
maize roots maize root curvatures observation Hoson. 1994
maize seedlings maize root curvatures observation Hoson et al. 1995
oat leaf senescence of oat leaf segments Miyamoto et al. 1995
Arabidopsis plants analysis of polar auxin transport Oka et al. 1995
Arabidopsis seedlings
and plants growth process observation Ishii et al. 1996
Chara rhizoids ovservation of displacement of statoliths Cai et al. 1997
Arabidopsis plants growth process observation Miyamoto et al. 1999
Arabidopsis seedlings
gene expression patterns investigation
Kittang et al. 2004 by microarray
etiolated pea seedlings
growth process observation and
Miyamoto et al. 2005 analysis of polar auxin transport
20
Arabidopsis callus
expression patterns of
Babbick et al. 2007 12 transcription factors
Arabidopsis callus 2-D gel electrophoresis, LC-IT-MS Barjaktarovic et al. 2007
Arabidopsis seedlings
growth process observation and
Soh et al. 2011 gene expression investigation by
microarray
Arabidopsis seedlings
gene expression investigation and
Soh et al. 2012 promoter in silico analysis
mutant and WT of
pea seedlings epicotyl and root curvatures observation Miyamoto et al. 2014
gravistimulation by reorientation
fruit trees investigation of growth and flowering Wareing et al. 1958
fruit trees
investigation of growth and
Wareing et al. 1961 polar auxin transport
mutant and WT of
growth process observation Lincoln et al. 1990
Arabidopsis seedlings
21
analysis of polar auxin transport
cucumber seedlings peg formation Takahashi and Scott. 1994
Arabidopsis seedlings growth process observation Fukaki et al. 1997
cucumber seedlings peg formation observation Takahashi. 1997
rice and barley growth process observation Abe et al. 1998
mutant and WT of root waving assay and
Luschnig et al. 1998
Arabidopsis seedlings gene expression investigation
mutant and WT of
root waving assay Marchant et al. 1999
Arabidopsis seedlings
Arabidopsis seedlings
gene expression investigation by
Moseyko et al. 2002 microarrays
mutant and WT of
root waving assay Boonsirichai et al. 2003
Arabidopsis seedlings
Arabidopsis, radish
root curvatures observation,
Takahashi et al. 2003 columella cells and amyloplast observation
22
microarrays
etiolated pea seedlings
growth process observation and
Hoshino et al. 2006 analysis of polar auxin transport
mutant and WT of
Arabidopsis seedlings mutagenesis and root waving assay Stanga et al. 2009
mutant and WT of rice seedlings
growth process observation and
Song and Xu. 2013 gene expression investigation
mutant and WT of
mutagenesis and root waving assay Zou et al. 2013
Arabidopsis seedlings
microgravity and 3-D clinorotation
sweet clover roots calcium redistribution investigation Hilaire et al. 1995
Arabidopsis seedlings
(dark-grown) plastid position observation Kraft et al. 2000
Chara rhizoids ovservation of statoliths positioning Braun et al. 2002
Arabidopsis callus gene expression investigation by
23
3-D clinorotation and
gravistimulation by reorientation
mutant and WT of
Arabidopsis seedlings
growth process observation and
Soh et al. 2015 gene expression investigation
24
Table IV-i A list of all WRKY family TFs identified in Arabidopsis thaliana. Listed are name, locus id, group (sub-family), function and the type of stress to which they respond. This table was modified from Eulgem et al. (44) with permission.
Gene name Locus ID Group
Function and stress type they respond
AtWRKY1 At2g04880 I salicylic acid
AtWRKY2 At5g56270 I heat
AtWRKY3 At2g03340 I regulation of transcription, DNA-templated, transcription
AtWRKY4 At1g13960 I drought, salt
AtWRKY5 At5g35738 Unclassified pseudogene
AtWRKY6 At1g62300 II cold, drought, salt
AtWRKY7 At4g24240 II calmodulin, salt
AtWRKY8 At5g46350 II hydrogen peroxide, bacterium, fungus
AtWRKY9 At1g68150 II regulation of transcription, DNA-templated, transcription
AtWRKY10 At1g55600 I endosperm development
AtWRKY11 At4g31550 II bacterium, chitin
AtWRKY12 At2g44745 II regulation of transcription, DNA-templated, transcription
AtWRKY13 At4g39410 II salicylic acid
AtWRKY14 At1g30650 II regulation of transcription, DNA-templated, transcription
AtWRKY15 At2g23320 II drought, salt
AtWRKY16 At5g45050 II defense response
AtWRKY17 At2g24570 II bacterium, chitin
25
AtWRKY19 At4g12020 I defense response protein phosphorylation
AtWRKY20 At4g26640 I wounding
AtWRKY21 At2g30590 II glucosinolate metabolic process
AtWRKY22 At4g01250 II leaf senescence, chitin
AtWRKY23 At2g47260 II auxin distribution
AtWRKY24 At5g41570 II regulation of transcription, DNA-templated, transcription
AtWRKY25 At2g30250 I cold, heat, osmotic, salt
AtWRKY26 At5g07100 I heat
AtWRKY27 At5g52830 II bacterium, nitric oxide, gibberellin
AtWRKY28 At4g18170 II cold, drought, salt
AtWRKY30 At5g24110 III drought, salt
AtWRKY30 At5g24110 III drought, salt, salicylic acid
AtWRKY33 At2g38470 I 3-D clinorotation, drought, salt
AtWRKY35 At2g34830 II embryo development ending, pollen development
AtWRKY36 At1g69810 II regulation of transcription, DNA-templated, transcription
AtWRKY38 At5g22570 III defense response, salicylic acid
AtWRKY39 At3g04670 II regulation of transcription, DNA-templated, transcription
AtWRKY40 At1g80840 II 3-D clinorotation, bacterium, fungus, drought, salt, wound
AtWRKY41 At4g11070 III regulation of transcription, DNA-templated, transcription
AtWRKY42 At4g04450 II regulation of transcription, DNA-templated, transcription
AtWRKY43 At2g46130 II regulation of transcription, DNA-templated, transcription
26
AtWRKY45 At3g01970 I phosphate ion transport
AtWRKY46 At2g46400 III 3-D clinorotation, aluminum-induced malate secretion, osmotic
AtWRKY47 At4g01720 II regulation of transcription, DNA-templated, transcription
AtWRKY48 At5g49520 II drought, salt
AtWRKY49 At5g43290 II regulation of transcription, DNA-templated, transcription
AtWRKY50 At5g26170 II bacterium, fungus, jasmonic acid
AtWRKY51 At5g64810 II bacterium, cold
AtWRKY52 At5g45260 III defense response
AtWRKY53 At4g23810 III drought
AtWRKY54 At2g40750 III defense response
AtWRKY55 At2g40740 III regulation of transcription, DNA-templated, transcription
AtWRKY56 At1g64000 II regulation of transcription, DNA-templated, transcription
AtWRKY57 At1g69310 II drought, salt
AtWRKY58 At3g01080 I regulation of transcription, DNA-templated, transcription
AtWRKY59 At2g21900 II abscisic acid
AtWRKY60 At2g25000 II abscisic acid, defense response
AtWRKY61 At1g18860 II cytokinin responses
AtWRKY62 At5g01900 III defense response, jasmonic acid
AtWRKY63 At1g66600 III abscisic acid, drought
AtWRKY64 At1g66560 III regulation of transcription, DNA-templated, transcription
AtWRKY65 At1g29280 II regulation of transcription, DNA-templated, transcription
27
AtWRKY66 At1g80590 III regulation of transcription, DNA-templated, transcription
AtWRKY67 At1g66550 III regulation of transcription, DNA-templated, transcription
AtWRKY68 At3g62340 II regulation of transcription, DNA-templated, transcription
AtWRKY69 At3g58710 II regulation of transcription, DNA-templated, transcription
AtWRKY70 At3g56400 III bacterium, fungus, jasmonic acid
AtWRKY71 At1g29860 II shoot branching
AtWRKY72 At5g15130 II defense response, salicylic acid
AtWRKY73 ESTs
(AV526908) I Undefined
AtWRKY74 At5g28650 II regulation of transcription, DNA-templated, transcription
28
VII. Literature cited
1. Takahashi H. Gravimorphogenesis: gravity-regulated formation of the peg in cucumber seedlings. Planta 1997; 203:S164-S169.
2. Hoson T. Automorphogenesis of maize roots under simulated microgravity conditions. Plant Soil 1994; 165:309-314.van Loon JJWA
3. Ishii Y, Hoson T, Kamisaka S, Miyamoto K, Ueda J, Mantani S, Fujii S, Masuda Y, Yamamoto R. Plant growth processes in Arabidopsis under microgravity conditions simulated by a clinostat. Biological Sciences in Space 1996; 10:3-7.
4. Babbick M, Dijkstra C, Larkin OJ, Anthony P, Davey MR, Power JB, Lowe KC, Cogoli-Greuter M, Hampp R. Expression of transcription factors after short-term exposure of Arabidopsis thaliana cell cultures to hypergravity and simulated microgravity (2-D/3-D clinorotation, magnetic levitation). Advances in Space Research 2007; 39:1182-1189.
5. Kiss JZ, Edelmann RE, Wood PC. Gravitropism of hypocotyls of wild-type and starch-deficient Arabidopsis seedlings in spaceflight studies. Planta 1999; 209:96-103. 6. Paul AL, Popp MP, Gurley WB, Guy C, Norwood KL, Ferl RJ. Arabidopsis gene expression patterns are altered during spaceflight. Space Life Sciences:
Gravity-Related Effects on Plants and Spaceflight and Man-Made Environments on Biological Systems 2005; 36:1175-1181.
7. Herranz R, Anken R, Boonstra J, Braun M, Christianen PC, de Geest M, Hauslage J, Hilbig R, Hill RJ, Lebert M, Medina FJ, Vagt N, Ullrich O, van Loon JJWA, Hemmersbach R. Ground-based facilities for simulation of microgravity: organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013; 13:1-17.
8. Hoson T, Kamisaka S, Masuda Y, Yamashita M, Buchen B. Evaluation of the three-dimensional clinostat as a simulator of weightlessness. Planta 1997; 203:S187-S197.
29
9. Hoson T. Plant growth and morphogenesis under different gravity conditions: Relevance to plant life in space. Life 2014; 4:205-216.
10. Hoson T, Kamisaka S., Masuda Y, Yamashita, M. Changes in plant growth processes under microgravity conditions simulated by a three-dimensional clinostat.
Botanical Magazine Tokyo 1992; 105:53–70.
11. van Loon JJWA. Some history and use of the random positioning machine, RPM, in gravity related research. Advances in Space Research 2007; 39:1161–1165. 12. Miyamoto K, Hoshino T, Yamashita M, Ueda J. Automorphosis of etiolated pea seedlings in space is simulated by a three-dimensional clinostat and the application of inhibitors of auxin polar transport. Physiologia Plantarum 2005; 123:467-474.
13. Miyamoto K, Yamasaki T, Uheda E, Ueda J. Analysis of apical hook formation in Alaska pea with a 3-D clinostat and agravitropic mutant ageotropum. Frontiers in Plant
Science 2014; 5: 1-8.
14. Smith JD, Todd P, Staehelin LA. Modulation of statolith mass and grouping in white clover (Trifolium repens) growth in 1-g, microgravity and on the clinostat. Plant Journal 1997; 12:1361-1373.
15. Kraft TF, van Loon JJWA, Kiss JZ. Plastid position in Arabidopsis columella cells is similar in microgravity and on a random-positioning machine. Planta 2000; 211:415-422.
16. Yamashita M, Yamashita A, Yamada M. Three-dimensional (3D-) clinostat and its operational characteristics. Biological Sciences in Space 1997; 11:112-118.
17. Hoson T, Kamisaka S, Yamamoto R, Yamashita M, Masuda Y. Automorphosis of maize shoots under simulated microgravity on a three-dimensional clinostat.
Physiologia Plantarum 1995; 93:346-351.
18. Miyamoto K, Oka M, Ueda J, Hoson T, Kamisaka S. The senescence of oat leaf segments is promoted under simulated microgravity condition on a three-dimensional clinostat. Biological Sciences in Space 1995; 9:327-330.
30
19. Takahashi H, Scott TK. Gravity-regulated formation of the peg in developing cucumber seedlings. Planta 1994; 193:580-584.
20. Ueda J, Miyamoto K, Yuda T, Hoshino T, Fujii S, Mukai C, Kamigaichi S, Aizawa S, Yoshizaki I, Shimazu T, Fukui K. Growth and development, and auxin polar transport in higher plants under microgravity conditions in space: BRIC-AUX on STS-95 space experiment. Journal of Plant Research 1999; 112:487-492.
21. Oka M, Ueda J, Miyamoto K, Yamamoto R, Hoson T, Kamisaka S. Effect of simulated microgravity on auxin polar transport in inflorescence axis of Arabidopsis
thaliana. Biological Sciences in Space 1995; 9:331-336.
22. Miyamoto K, Yamamoto R, Fujii S, Soga K, Hoson T, Shimazu T, Masuda Y, Kamisaka S, Ueda J. Growth and development in Arabidopsis thaliana through an entire life cycle under simulated microgravity conditions on a clinostat. Journal of Plant
Research 1999; 112:413-8.
23. Kimbrough JM, Salinas-Mondragon R, Boss WF, Brown CS, Sederoff HW. The fast and transient transcriptional network of gravity and mechanical stimulation in the
Arabidopsis root apex. Plant Physiology 2004; 136:2790–2805.
24. Moseyko N, Zhu T, Chang HS, Wang X, Feldman LJ. Transcription profiling of the early gravitropic response in Arabidopsis using high-density oligonucleotide probe microarrays. Plant Physiology 2002; 30:720-8.
25. Centis-Aubay S, Gasset G, Mazars C, Ranjeva R, Graziana A. Changes in gravitational forces induce modifications of gene expression in A. thaliana seedlings.
Planta 2003; 218:179–185.
26. Boonsirichai K, Guan C, Chen R, Masson PH. Root gravitropism: an experimental tool to investigate basic cellular and molecular processes underlying mechanosensing and signal transmission in plants. Annual Review of Plant Physiology 2002; 53:421-447.
31
Biology 2014; 16 Suppl 1:79-90.
28. Toyota, M., Furuichi, T., Tatsumi, H., Sokabe, M. Cytoplasmic calcium increases in response to changes in the gravity vector in hypocotyls and petioles of Arabidopsis seedlings. Plant Physiology 2008; 146:505-154.
29. Toker, A. Phosphoinositides and signal transduction. Cellular and Molecular Life
Sciences 2002; 59:761–779.
30. Fasano JM, Swanson SJ, Blancaflor EB, Dowd PE, Kao TH, Gilroy S. Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 2001; 13:907–921.
31. Zheng HQ, Han F, Jie L. Higher Plants in Space: Microgravity Perception, Response, and Adaptation. Microgravity Science and Technology. 2015; 27:377–386. 32. Schena M, Shalon D, Davis RW, Brown PO. Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 1995; 270: 467– 470.
33. Schena M, Shalon D, Heller R, Chai A, Brown PO, Davis RW. Parallel human genome analysis: Microarraybased expression monitoring of 1,000 genes.
Proceedings of the National Academy of Sciences of USA 1996; 93:10614–10619.
34. Heller RA, Schena M, Chai A, Shalon D, Bedilion T, Gilmore J, Woolley DE, Davis RW. Discovery and analysis of inflammatory disease-related genes using cDNA microarrays. Proceedings of the National Academy of Sciences of USA 1997; 94:2150–2155.
35. DeRisi JL, Iyer VR, Brown PO. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997; 278:680–686.
36. Wodicka L, Dong H, Mittmann M, Ho MH, Lockhart DJ. Genome-wide expression monitoring in Saccharomyces cerevisiae. Nature Biotechnology 1997; 15:1359–1367. 37. Seki M, Narusaka M, Abe H, Kasuga M, Yamaguchi-Shinozaki K, Carninci P,
32
Hayashizaki Y, Shinozaki K. Monitoring the expression pattern of 1,300 Arabidopsis genes under drought and cold stresses using a full-length cDNA microarray. Plant Cell 2001; 13:61–72.
38. Shinozaki K, Yamaguchi-Shinozaki K, Seki M. Regulatory network of gene expression in the drought and cold stress responses. Current Opinion in Plant Biology 2003; 6:410–417.
39. Wellmer F, Riechmann JL. Gene network analysis in plant development by genomic technologies. International Journal of Developmental Biology 2005; 49:745– 759.
40. Haberer G,Mader MT, Kosarev P, Spannagl M, Yang L, Mayer KFX. Large-scale
cis-element detection by analysis of correlated expression and sequence conservation
between Arabidopsis and Brassica oleracea. Plant Physiology 2006; 142:1589–1602. 41. Ma S, Gong Q, Bohnert HJ. An Arabidopsis gene network based on the graphical Gaussian model. Genome Research 2007; 17:1614–1625.
42. Mochida K, Yoshida T, Sakurai T, Yamaguchi-Shinozaki K, Shinozaki K, Tran LS.
In silico analysis of transcription factor repertoire and prediction of stress responsive
transcription factors in soybean. DNA Research 2009; 16:353-369.
43. Rushton PJ, Somssich IE, Ringler P, Shen QJ. WRKY transcription factors. Trends
in Plant Science 2010; 15:247-258.
44. Eulgem T, Rushton PJ, Robatzek S, Somssich IE. The WRKY superfamily of plant transcription factors. Trends in Plant Science 2000; 5:199-206.
45. Krysan PJ, Young JC, Sussman MR. T-DNA as an Insertional Mutagen in
Arabidopsis. Plant Cell 1999; 11:2283–2290.
46. Parinov S, Sevugan M, Ye D, Yang, W-C, Kumaran M, Sundaresan V. Analysis of Flanking Sequences from Dissociation Insertion Lines: A database for reverse genetics in Arabidopsis. Plant Cell 1999; 11:2263–2270.
33
47. McKinney EC, Ali N, Traut A, Feldmann KA, Belostotsky DA, McDowell JM, Meagher RB. Sequence-based identification of T-DNA insertion mutations in
Arabidopsis: actin mutants act2-1 and act4-1. Plant Journal 1995; 8:613-622.
48. Krysan PJ, Young JC, Tax F, Sussman MR. Identification of transferred DNA insertions within Arabidopsis genes involved in signal transduction and ion transport.
Proceedings of the National Academy of Sciences of USA 1996; 93:8145–8150.
49. Martienssen RA. Functional genomics: Probing plant gene function and expression with transposons. Proceedings of the National Academy of Sciences of USA 1998; 95:2021–2026.
50. Wisman E, Hartmann U, Sagasser M, Baumann E, Palme K, Hahlbrock K, Saedler H, Weisshaar B. Knock-out mutants from an En-1 mutagenized Arabidopsis thaliana population generate phenylpropanoid biosynthesis phenotypes. Proceedings of the
34
Chapter 1
Gene expression changes in Arabidopsis seedlings
during short- to long-term exposure to 3-D
clinorotation
Hyuncheol Soh, Chungkyun Auh, Woong-Young Soh, Kyeongsik Han, Donggiun Kim, Sukchan Lee, Yong Rhee. Gene expression changes in Arabidopsis seedlings during short- to long-term exposure to 3-D clinorotation. 18 March 2011, Planta 234:255-70.
Author contribution:
Hyuncheol Soh designed and conducted the experiments, analyzed data, and wrote the paper. Chungkyun Auh provided funds for performing the microarray experiments.
Woong-Young Soh and Donggiun Kim helped with their critical review of this manuscript. Kyeongsik Han provided 3-D clinorotation equipment.
Yong Rhee helped in analyzing the data and writing this manuscript.
Sukchan Lee hosted me in his lab and provided all the equipment and chemicals for performing the experiments as well as supported in conceiving and designing experiments.
35
1.1. Abstract
Seedlings of Arabidopsis thaliana (cv. Columbia) were used to evaluate dynamic transcriptional-level genome responses to 3-D clinorotation. The microarray data analysis showed that the plant may respond to 3-D clinorotation by dynamic induction (up- and down-regulations) of the responsive genes in the genome. The qRT-PCR results on the investigated genes showed that the expression patterns of the genes (molecular response) were generally similar to the physiological response patterns detected in stress-challenged plants. Expression patterns were categorized into short or continual up- or down-regulated patterns, as well as stochastic changes from short- to long-term under 3-D clinorotation. The induced genes are then assumed to establish a new molecular plasticity to the newly adjusted genome status in the basic milieu of maintaining homeostasis during the process of adaptation to 3-D clinorotation.
1.2. Introduction
The growth patterns of higher plants show bi-directionality with downward orientation in the root and upward orientation in the leaves under the normal 1g force field of Earth. Such gravitropic growth of plants assumes the evolutionarily best-selected morphological/physiological outcomes for the photosynthetic apparatus to maximize uptake of water/nutrients (by the roots) from the soil, as well as solar photons (by the leaves). So, changes of normal gravity force can collectively constitute a novel stress which, similarly to other environmental stresses, serves as a challenge to growth and development in plants. Especially, minimization or reduction of normal gravity (i.e. microgravity) could be created during true space flight or by machine-based simulation (clinorotation) for ground-machine-based experiments, and considerable amounts of documentation have been accumulated about responses of plants under minimized gravity conditions.
The majority of traditional research works have focused on developmental and physiological responses of plants to minimized gravity such as morphological changes
36
or gravitation-related hormone/calcium redistribution (1, 2, 3, 4). On the other hand, continual progress has been also made toward a deep understanding of plant responses to minimized gravity conditions at the global genome level. For instance, microgravity effects were investigated on gene expression (at transcription level) as well as morphological and metabolic changes for dwarf wheat under true microgravity condition in space flight; the collective results showed that there were no significant alterations on such examined traits (5). However, apart from it, the significant alterations of gene expression under minimized gravity conditions have been occasionally reported in many research works so far. An example comes from a study with the fern Ceratopteris richardii. The authors reported that transcriptional changes of the genes could be induced in this fern species under microgravity condition in space flight (6). Even though minimized gravity effect was investigated at the single cell level in this case, this work provided a promising clue to extrapolate the results to multi-cellular organisms in response to altered (minimized) gravity condition. In fact, most of interesting reports have been accumulated mainly for an easily manipulated weed plant species with small genome size, Arabidopsis. The gene expression change was reported from Arabidopsis seedlings exposed to a true microgravity environment during space flight (7). In this case, 182 genes in Arabidopsis genome showed differential expression in response to the space flight condition by more than 4-folds in comparison of ground control plants. In addition, Arabidopsis cell cultures were exposed to true microgravity on a sounding rocket and altered changes of the gene activity at transcription level were profiled via DNA chip (8).
From substantial documentation so far, it is now obvious from the above researches that plant genome is responsive to altered (minimized) gravity condition and furthermore, its sensitivity is likely to be dependent upon their genome size. In fact, it might be quite difficult to screen out significant changes of gene activity in polyploid wheat genome with multiple copies of the genes under minimized gravity condition which could not be a potentially strong stress trigger to disturb stable genomic milieu.
In general, the living organisms should stay in the altered (zero or minimized) gravity conditions for a long period during real space flight. So, long term exposure of
37
plants to 3-D clinorotation conditions would be desirable in ground-based experiments in order to obtain comparable results to the information from real space flight in order to draw generalization about plant gene response to space environment. However, apart from one exceptional case in which the global re-setting of gene expression in
Arabidopsis genome was detected under 3-D clinorotation environment via
clinorotation for a prolonged period up to 6 days (9), most of ground based research works have been conducted under relatively short term exposure of plants to minimized gravity conditions. Kittang et al. performed DNA chip experiments for profiling of the genes with altered expressions under 3-D clinorotation condition (1 or 24 h) and found out that about 2.2 % of total 8000 genes screened showed differential expression levels (10). The experiment was done for cultured Arabidopsis cells to monitor the contents of translational gene products in response to 3-D clinorotation (59, 60). In this case, proteome of the exposed undifferentiated plant cells (calli) mounted a significant response to microgravity conditions within a period of 10 min to 16 h. Furthermore, these earlier studies provide no information about time-course based response pattern of the plant genes from short- to long-term exposure to 3-D clinorotation. Actually, this information would valuable because they provide us with important clues on the prediction of molecular tolerance or adaptation of the plant genome in real space environment.
The main objective of this study was to identify the responsive genes during a long period (from 1 h to 6 days) exposure of Arabidopsis under 3-D clinorotation and, subsequently, investigate time-course based gene expression patterns for them. The results are discussed in the context of long-term based plant gene responses in relation to their possible functions for tolerance and adaptation to 3-D clinorotation.
1.3. Materials and Methods
1.3.1. Plant materials and exposure to 3-D clinorotation
All experiments were conducted using 6-day-old seedlings of Arabidopsis
thaliana (L.) Heynh. (ecotype Columbia). The seeds (Arabidopsis Biological Resource
38
plating in Petri dishes for germination and initially surface-sterilized with 70% (v/v) ethanol for 5 min. After treating the seeds with a 20% commercial bleach solution, they were subsequently washed five times in sterile distilled water. The sterilized seeds were finally germinated on 1.5% (w/w) agar plates containing full-strength Murashige and Skoog nutrient medium supplemented with 3% sucrose. The seedlings were cultivated in a growth chamber with 18 h of fluorescent light (40 W m–2) and a constant temperature of 24°C.
The 3-D clinostat consisted of two independently rotating frames, one rotating within the other (but in a different plane) to maximize the microgravity effect. All manipulations were carried out in accordance with the instructions of manufacturer (Watanabe Machine Design Associates, Japan; 30) and refer to the citations (13, 14, 15) for details and specifications of the machine. Under the same environmental conditions as were employed for seedling germination, the selected seedlings were exposed to 3-D clinorotation with a 3-D clinostat, and the intact seedlings (the control) were maintained separately on the shelf during the experiments. All of the seedlings were frozen in liquid nitrogen within a few seconds immediately after harvesting. This step excluded any possibility of 1G interference during the handling procedure for 3-D clinorotation -treated seedlings.
1.3.2. RNA preparation, probe labeling, and microarray hybridization
The harvested seedlings were immediately frozen in liquid nitrogen within a few seconds after harvesting and stored in a deep freezer at -70℃. RNA was extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA), and the RNA integrity was confirmed via separation on 1.2% (w/w) agarose-formaldehyde gels with ethidium bromide staining. For the microarray analysis of under 3-D clinorotation and control seedlings, the synthesis of target cRNA probes and hybridization was conducted using Agilent’s Low RNA Input Linear Amplification Kit (Agilent Technology, Palo Alto, CA, USA), in accordance with the manufacturer’s instructions. In brief, each 1 µg of total RNA and T7 promoter primer were mixed and incubated at 65℃ for 10 min. The cDNA master mix (5X first strand buffer, 0.1M DTT, 10mM dNTP mix, RNase-Out, and
MMLV-39
RT) was prepared and added to the reaction. The samples were incubated for 2 h at 40℃ and then the reverse transcription (RT) and double strand DNA synthesis was completed for 15 min of incubation at 65℃. The transcription master mix was prepared according to the manufacturer’s recommendations (4X transcription buffer, 0.1M DTT, NTP mix, 50% PEG, RNase-Out, inorganic pyro-phosphatase, T7 RNA polymerase and Cyanine 3/5-CTP). Transcription of dsDNA was carried out by the adding the transcription master mix to the dsDNA reaction samples, followed by 2 h of incubation at 40℃. Amplified and labeled cRNA was purified on the cRNA Clean-up Module (Agilent Technology) according to the manufacturer’s recommended protocols. Labeled target cRNA was quantified with an ND-1000 spectrophotometer (Nano Drop Technologies, Wilmington, DE, USA). After checking the labeling efficiency, cRNA fragmentation was conducted via the addition of 10X blocking agent and 25X fragmentation buffer, followed by 30 min of incubation at 60℃. The fragmented cRNA was then re-suspended with 2X hybridization buffer and pipetted directly onto the assembled Arabidopsis Gene Expression Microarray (44K) (Agilent Technology). The arrays were hybridized at 65℃ for 17 h using a hybridization oven (Agilent Technology). The hybridized microarrays were then washed as per the manufacturer’s washing protocols.
1.3.3. Acquisition and analysis of microarray data
The hybridized images were scanned using a DNA microarray scanner and quantified with Feature Extraction Software (Agilent Technology). Probe sets with signal values lower than the detectable range were adjusted to 75, and probe sets with the values of 75 for all conditions were removed from subsequent analysis. All data normalization and selection of fold-changed genes were conducted using Gene Spring GX 7.3 (Agilent Technology); scanned DNA arrays were normalized to a baseline array with median overall expression using dCHIP (16). The averages of normalized ratios were calculated by dividing the average of the normalized signal channel intensity by the average of the normalized control channel intensity. The standard deviation of the
40
vertical controls (four biological replicates) was employed to identify genes of significant changes relative to the vertical controls (P value < 0.05). Functional annotation of the identified genes was conducted by gene ontology analysis with the EasyGO program (http://www.bioinformatics.cau.edu.cn/easygo) (17).
1.3.4. Quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR)
Highly sensitive quantitative real-time reverse transcription polymerase chain reaction was employed to confirm the expression of the selected genes. At least two independent biological replicates of each sample and three technical replicates of each biological replicate were used for real-time PCR analysis. The genes and their qRT-PCR primers are presented in Table 1-1.
The 18S rRNA (8) was amplified as an internal reference for all qRT-PCRs using the following primers: forward (5’-GATGAGCCTGCGTAGTATTAGG-3’) and reverse (5’-AGTCATTCCGAAGAACACTTGC-3’). Reverse transcription was conducted to synthesize the complementary DNA by mixing the following components in order: 5 µg total RNA, 5 µl 5X RT buffer, 1.14 µl oligo-dT (18-mer) (final conc. 1.5 pmoles/µl), 1.25 µl 10 mM dNTPs (final conc. 0.5 mM), 0.05 µl MMLV reverse transcriptase (200 U/µl) (Promega, Madison, WI, USA) (final conc. 10 U), and distilled water (DNase-free) up to a total volume of 25 µl. After the reaction was incubated for 1 h at 37℃, real-time PCR was conducted by adding the following reagents: 3 µl synthesized cDNA, 1 µl primer (final conc. 0.25 mM), and 10 µl 2X SYBR mixture solution (Qiagen, Valencia, CA, USA), and distilled water up to a total volume of 20 µl. The cDNA amplification was conducted in a Rotorgen 3000 Real-time Amplification System (Corbett Research, Australia) with the following thermal profile: 40 cycles at 94℃ (10 s), 59℃ (20 s), 72℃ (15 s), and, finally, 95℃. The PCR products were analyzed on 1.5% ethidium bromide-stained agarose gels. Amplification of all cDNAs with specific primers yielded PCR products of the predicted sizes (150~200 bp) from the respective gene sequences. The data were normalized using a reference (the ribosomal 18S transcript), and the relative quantity of mRNA was calculated by the