Ikeya, 1988), 中 ノ 海 (Ishizaki, 1969), 能 登 九 十 九 湾 (Kamiya et al., 2001), 大 阪 湾 (Yasuhara and Irizuki,
2001), 播 磨 灘 ( 入 月 他,2009), 浦 ノ 内 湾(Ishizaki, 1968; Irizuki et al., 2008),浦内湾 (Irizuki et al., 2006) 等 において,多くのデータが蓄積されてきている。一方, 干潟の貝形虫相の研究例はごく少ない。これは,調査と なる干潟そのものが減少していること,さらに,干潟に 生息する貝形虫の多くは,浅海性種と比べて背甲の石灰 化の度合いが低いため,化石として保存されにくく,古 環境指標として用いられる機会が少なかったことに起因 する。しかし,干潟に生息する貝形虫類が古環境復元に 重要な情報をもたらした例もある (中尾ほか,2008)。ま た,河川汽水域は,河川流量,地形,底質といった要素 が相互に作用し,底生生物に様々な環境を与える場であ 1 .はじめに 貝形虫類は,その多くが体長1mm以下の微小な底生 甲殻類で,淡水域から汽水域,海水の浅海から深海と多 様な環境に生息している。また,分類群によっては環境 に応じて棲み分けていること,背甲とよばれる石灰質の 二枚殻の形態が分類形質として有用であり,それらが化 石として堆積物に保存されることから,古環境復元の指 標として用いられてきた。特に海水準変動を記録する更 新統から完新統を対象とした古環境復元には,現生の沿 岸域の貝形虫相のデータが不可欠である。内湾域を含む 日本沿岸の貝形虫相については,青森湾 (Ishizaki, 1971), 仙台湾 (Ikeya and Itoh, 1991),館山湾 (Frydl, 1982),浜 名 湖 (Ikeya and Hanai, 1982), 伊 勢 湾 (Bodergat and
上山紗也加
*・中尾有利子
**・小沢 広和
***Brackish-water ostracod fauna was studied in the intertidal zone of the Tama River Estuary, Tokyo Bay, central Japan. Nineteen species belonging to 12 genera of Ostracoda were identified, of which two, Ishizakiella miurensis and Nipponocy-there sp., were living. It was revealed that the living ostracod fauna of this study area had extremely low species diversity and was dominated by I. miurensis. The life cycle of I. miurensis was studied from May 2012 to February 2013, and the fol-lowing characteristics were observed: older juvenile instars (A-1, A-2) and adults passed the low water temperature sea-son as lone individuals; eggs and younger juvenile instars (A-7, A-8) remained in female carapaces for the low water tem-perature season and ceased development in winter; in spring, wintering adults produced a new generation, and over-wintering juvenile instars recommenced development to produce the next generation; and the eggs of the next generation that was deposited in spring developed into the subsequent generation. Consequently, the entire population during sum-mer and autumn consisted of plural generations.
Keywords: Ostracoda,Tokyo Bay, Tama River Estuary, tidal flat, brackish-water, population dynamics
多摩川河口干潟の貝形虫相とその季節変化
Ostracod Fauna and Its Seasonal Change in the Tidal Flat of the Tama River Estuary,
Tokyo Bay, Central Japan
Sayaka KAMIYAMA
*, Yuriko NAKAO
**and Hirokazu OZAWA
***(Accepted November 11, 2016)
* Kayama System Machinery Co., Ltd: 9th Floor, Sumitomo Fudosan Shibadaimon Building, 2-5-5, Shibadaimon, Minato-ku, Tokyo, 105-0012, Japan
** Depar tment of Ear th & Environmental Sciences, College of Humanities & Sciences, Nihon University: 3-25-40, Sakurajosui, Setagaya-ku, Tokyo, 156-8550, Japan
*** Earth Science Laboratory, College of Bioresource Sciences, Nihon University: 1866, Kameino, Fujisawa, Kanagawa, 252-0880, Japan * カヤバシステムマシナリー株式会社: 〒105-0012 東京都港区芝大門 2 丁目 5 番地 5 号 住友不動産芝大 門ビル9 階 ** 日本大学文理学部地球科学科: 〒156-8550 東京都世田谷区桜上水3-25-40 *** 日本大学生物資源科学部一般教養地球科学研究室: 〒252-0880 神奈川県藤沢市亀井野1866
り,そこに生息する貝形虫相は比較的狭い範囲内ながら も,場所毎に変化に富むと考えられる。東京湾では東側 の小櫃川河口干潟からのみ貝形虫相の報告があるが(Na-kao and Tsukagoshi, 2002;中尾・塚越,2008),この 1 地域の報告のみで 東京湾の干潟の貝形虫相 を代表さ せるのは不十分といえる。そこで,本研究は,東京湾沿 岸の貝形虫相の解明をめざし,東京湾の西側に位置する 多摩川の河口に形成された干潟を調査地とし,貝形虫相 とその季節変化を明らかにする。また,湾の東側に位置 する小櫃川河口干潟の貝形虫相との比較と,多産種の生 活史の解析を行う。さらに,現在の東京湾の沿岸環境を 記録し,底生生物のモニタリング,環境評価,古環境復 元のための基礎データを蓄積することを目的とする。 2 .試料と方法 多摩川河口域左岸の干潟を調査地 (図 1) とし,2012 年5月から2013年 2月の干潮時に 5 回の堆積物試料採取 を行った。多摩川の流軸方向約3kmの範囲で,川岸に 沿って干出する干潟上に,流軸に直交する4 つの調査ラ インを設定した。最も下流のラインをTM1 (河口原点よ り約0.8km下流),そこから上流に向かってTM2,TM3, TM4とした (図 1)。それぞれのラインで最も陸側の地 点を番号1 として,河川流軸に直角方向に20mおきに試 料 採 取 地 点 を 設 定 し, 陸 側 か ら 順 に, 番 号 を つ け た (例:TM1-1,TM1-2,TM1-3;図1右下枠内)。試 料採取時の潮位により,各ラインで採取した試料数は異 なる。また,ラインTM3付近の護岸の下部から,定常 的に水が流れ出ており,堆積物上に流路を作っていた。 その流路の中に地点TM3-1,流路から外れた場所に地 点TM3-1’を設定した (図2D)。ラインTM3 は低潮線ま での距離が他のラインと比較すると短かった (60m以 下)。汀線まで可能な限り近づき,汀線の陸側で試料を 採取し,地点TM3-4’とした。その他,護岸に埋められ た排水管から定常的に水の流れ込みがある場所を試料採 取地点Aとした (図 1,図2C)。また,2012年 7月の調 査の際,干潮時の汀線より海側で1 サンプル採取し,地 点Bとした (図 1)。試料は2012年 5月 7日に17地点,7 月21日に19地点,9月15日に17地点,12月2日に10地 点,2013年 2月27日に16地点から,合計79個を採取し た。 試料は堆積物表層を約200cm3 (面積10cm×20cm,表 層1cm) を定量して採取した。1mmと0.063mmの篩を 用いて水洗し,0.063mmの篩に残った堆積物を60℃で 乾燥させ,四塩化炭素で浮選し,顕微鏡下で貝形虫類を 拾い出した。これらの貝形虫類は60マスファウナルス ライド上で整理し,分類・同定・計数した。貝形虫個体 数が多く含まれる試料は,200個体を含む程度に縮分し, 縮分した試料をすべての貝形虫類を拾いきるようにし た。分析に用いた試料とそれぞれの縮分率を表1 に示 す。合弁殻 (背甲),片殻ともに 1 個体として計数し, 背甲の中に軟体部が残っているものを生体,それ以外を 遺骸とした。 また,試料採取時に塩濃度 (塩濃度計 ATC-S/Mill-E, アタゴ),水温 (水温計SK-250WPⅡ-R,佐藤計量器製 図1 調査地点の地図
Haneda Airport
TM1
TM2
TM3
TM4
A
Tama R.
50mB
500m ● River mouth←
sandy bottom low-water lineTM1-1
TM1-2
TM1-3
TM1-4
10km NTokyo Bay
Tama R.
TokyoObitsu R.
35°20’ 139°40’ 140°20’ 35°70’study area
Tokyo Bayた。ただし,堆積物の深さ1cm以内で水質計測に十分 な量の水が得られなかった場合は,計測しなかった。ま た,溶存酸素は計測に十分な水深がない場合は計測しな かった。 作所),pH (pH計PH71,横河電機),溶存酸素 (溶存酸 素計ID-100,飯島電子工業) を計測した。これらは,試 料を採取した堆積物の直上の水を測定し,試料採取地点 が干出していた場合は,堆積物の間隙水の水質を測定し 図2 調査地点の写真 A,ラインTM1の岸から流軸方向をみたところ;B,ラインTM2の岸から流軸方向をみたところ.満潮時近くに撮影したため,汀線が近い;C,地点Aの様 子.右下にある折り尺の一辺が約20cm;D,地点TM3-1とTM3-1’の様子.バケツの高さが約26cm;E,ラインTM3で下流側をみたところ.満潮時にも水 没しない高まりがある;F,地点TM4-1付近から下流方向をみたところ.
物表面を覆う状態で,所々水深約5cmの水たまりが あった (図2A)。 ラインTM2の底質は泥混じりの砂であったが,ぬか るみがラインTM1より強くなり,泥の含まれる割合が 高かった。干潮時はラインTM1同様,海水がひたひた にある状態で,表層はデトリタスに覆われており,陸側 にはしばしば礫が堆積していた (図2B)。 ラインTM3の底質は泥混じりの砂であった。上流側 と下流側に,満潮時にも水没しない高まりがある (図 2E)。また,干潟の岸よりには,葦原がある。ライン TM3の岸側は,堆積物が固くしまった状態で,表面が 平らであった。地点TM3-1’は平らな砂の表層,地点 図示された標本 (SUM-CO-2393からSUM-CO-2419) は,静岡大学キャンパスミュージアムに登録,保管され ている。 3 .結果 3-1)試料採取地点の詳細 以下に試料採取地点の底質,試料採取時の状態を示 す。ここでは,堆積物の表面を覆う,藻類等の有機物を 多く含む流動的な茶色い物質をデトリタスと表現する。 ラインTM1の底質は,泥混じりの砂で,水洗した 際,すべての試料の中で,篩に砂が最も多く残った。表 層がデトリタスに覆われ,干潮時は水がひたひたと堆積 表1 試料採取地点の特徴と分析に用いた試料の縮分率 採取地点の底質と特徴 各試料の縮分率
May, 2012 Jul., 2012 Sep., 2012 Dec., 2012 Feb., 2013
B 泥混じりの砂。潮下帯で,試料採取時の水深は20cm程度。 1 TM1-1 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。時折波をかぶる。 1 1 1/4 1 1 TM1-2 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。時折波をかぶる。 1 1 1/4 1 TM1-3 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。時折波をかぶる。 1 1 1 TM1-4 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。時折波をかぶる。 1 A 泥混じりの砂。護岸を貫く排水管から流れ出る水の流路。定常的に流れがあるが,穏やかで,表層に泥がたまる。 1/2 1/4 1/2 1 1 TM2-1 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。 1/16 1/32 1/128 1 1/2 TM2-2 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。 1/16 1/32 1/64 1/32 1/2 TM2-3 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。 1/8 1/16 1/16 1/4 TM2-4 泥混じりの砂。堆積物表面はデトリタスに覆われ,所々に水たまりや澪筋があり,水がひたひたとした状態。 1 1/8 1/16 1/4 TM3-1 泥混じりの砂・礫。護岸の下からしみ出る水の流路で定常的な水流がある。 1 1/2 1/4 1 1 TM3-1 泥混じりの砂。水たまり,澪筋はない。泥が多く,水分が多い所ではぬかるむ。 1 1/4 1/2 1 1 TM3-2 泥混じりの砂。砂がしまった状態でデトリタスが少ないこともあれば,デトリタスに覆われ,水がひたひたとした状態の事もある。 1/2 1/64 1/16 1/4 1/8 TM3-3 泥混じりの砂。われ,所々に水たまりや澪筋があり,水がひたひたとした状態。TM3-2 よりぬかるむ。堆積物表面はデトリタスに覆 1 1/4 1/2 1 TM3-4' 泥混じりの砂。泥が多くぬかるむ。所々に水たまりや澪筋があり,水がひたひたとした状態。干潮時にも時折波をかぶる。 1 1 1 1/2 TM4-1 泥混じりの砂。泥が多くぬかるむ。澪すじ,水たまりがある。 1/16 1/16 1/4 1/2 1/2 TM4-2 泥混じりの砂。水たまり,澪筋はない。泥が多く,水分が多い所ではぬかるむ。 1 1 1/4 1 1 TM4-3 泥混じりの砂。水たまり,澪筋はない。泥が多く,水分が多い所ではぬかるむ。 1 1 1 1 1 TM4-4 泥混じりの砂。水たまり,澪筋はない。泥が多く,水分が多い所ではぬかるむ。 1 1 1 1/4 縮分率の空欄は試料を採取していないことを示す.
のラインTM3とTM4は変化の幅が大きかった。調査期 間中に測定された塩濃度の最小値は2ppt (2012年 5月地 点TM4-4,2012年 7月 地 点TM3-1), 最 大 値 は 30ppt (2013年 2月地点TM1-1,TM1-2)であった。 水温 季節,天候などにより変化した。2015年5月は19.2– 29.3℃,7月は23.0–25.4℃,9月は27.8–34.2℃,12月は 12.5–15.2℃,2013年2月は8.3–10.7℃であった (表 3)。 調査期間中に測定された水温の最小値は8.3℃ (2013年 2 月 地 点TM2-1,TM3-3), 最大 値 は34.2 ℃ (2012年 9月 地点TM3-2) であった。 pH 2012年 5月は7.35–8.70,7月は7.71–8.47,9月は7.68– 8.38,12月 は7.79–8.44,2013年 2月 は8.03–8.55で あ っ た (表 4)。調査期間中に測定されたpHの最小値は7.35 (2012年 5月の地点TM3-1),最大値は8.70 (2012年 5月 地点TM4-1) であった。 溶存酸素 溶存酸素は2012年 5月が2.4–5.8 mg/l,7月が2.9–6.4 mg/l,9月が3.7–5.9 mg/l,12月が3.8–4.8mg/l,2013年 2月は計器の不調により計測できなかった (表 5)。調査 期間中に測定された溶存酸素の最小値は2.4mg/l (2012 年5月地点TM3-1),最大値は6.4mg/l (2012年 7月地点 TM4-1) であった。 TM3-1は護岸から定常的に流れ出る水の流路であり, どちらも表面にデトリタスはほとんどなかった(図 2D)。地点TM3-2より川側は,水がひたひたとした状態 で,堆積物表面はデトリタスに覆われていた (図2E)。 他のラインと比較すると,護岸から汀線までの距離が短 く,幅の狭い干潟である。 ラインTM4の底質は非常にぬかるんだ状態で,泥を 多く含み,陸側に礫が所々堆積し,澪筋があった (図 2F)。他のラインと比べると澪筋以外に水たまりがほと んど見られず,デトリタスは4 つのラインの中で最も少 なかった。 地点Aは河川の外部から,堤防をつきぬけて,排水を 川に誘導する直径約80cmの土管の前に設けた採取地点 で,土管からは定常的に水が流れていた (図2C)。この 水流は比較的ゆるやかで,堆積物表面はデトリタスに覆 われていた。 3-2)水質 以下はすべて試料採取時(干潮時)の値である。 塩濃度 塩濃度は,ライン毎に特徴があった。すなわち,ライ ンTM1は17–30ppt,ラインTM2は13–29ppt,ラインTM3 は2–28ppt,ラインTM4は2–25ppt,地点Aは8–24pptで あった (表 2)。最も下流のラインTM1が調査期間を通 して,比較的高い塩濃度で安定していた。反対に上流側 表2 試料採取地点の塩濃度
地点 May, 2012 Jul., 2012 Sep., 2012 Dec., 2012 Feb., 2013
B − − − − TM1-1 20 20 24 25 30 TM1-2 20 20 23 − 30 TM1-3 20 21 25 − − TM1-4 − 17 − − − A 11 8 22 24 19 TM2-1 17 13 20 21 27 TM2-2 19 16 24 21 29 TM2-3 21 19 − 27 TM2-4 16 25 − 20 TM3-1 6 2 10 5 TM3-1’ 11 4 4 TM3-2 16 17 20 18 28 TM3-3 15 16 19 − 25 TM3-4’ 6 19 − TM4-1 4 11 12 25 TM4-2 20 14 TM4-3 7 15 15 16 TM4-4 2 6 15 − 16 単位はppt.試料採取時に計測できなかった場合は空欄,試料採取をして いない場合は−で示す. 表3 試料採取地点の水温
地点 May, 2012 Jul., 2012 Sep., 2012 Dec., 2012 Feb., 2013
B − − − − TM1-1 13.8 8.6 TM1-2 25.2 25.4 29.2 − 8.7 TM1-3 25.0 25.3 28.8 − − TM1-4 − 24.6 − − − A 20.3 23.3 ß 13.3 9.0 TM2-1 26.8 24.1 31.5 13.8 8.3 TM2-2 25.6 23.8 30.2 9.0 TM2-3 25.4 − 9.0 TM2-4 30.5 − 9.0 TM3-1 19.2 23.0 27.8 15.2 TM3-1’ TM3-2 29.0 24.6 34.2 8.7 TM3-3 27.3 24.7 31.6 − 8.3 TM3-4’ − 24.5 30.0 − 9.2 TM4-1 29.3 25.1 12.5 8.8 TM4-2 32.0 13.0 TM4-3 25.3 13.0 10.7 TM4-4 23.5 30.5 − 10.7 単位は℃.試料採取時に計測できなかった場合は空欄,試料採取をして いない場合は−で示す.
た。よって,属レベルでは12属が確認された。産出し た貝形虫種と個体数 (生体/遺骸) を表 6 から表10に示 す。生体はIshizakiella miurensis と Nipponocythere sp. の 2 種のみであった。すべての試料で I. miurensisがほぼ 独占し,産出個体数には差があるものの,すべての試料 から生体が産出した。Nipponocythere sp. の生体は,2012 3-3)産出した貝形虫 全79試料のすべてから貝形虫が産出し,19種が確認 された (図 3)。このうち,4 種は産出個体数が極端に少 なく,また標本の状態も悪かった。そのため,3 種につ いてはDarwinulid sp., Candonid sp., Cypridid sp.と科レ
ベルの同定にとどめ,その他分類群が不明な1 種とし
表4 試料採取地点の pH
地点 May, 2012 Jul., 2012 Sep., 2012 Dec., 2012 Feb., 2013
B − − − − TM1-1 7.81 7.85 8.15 TM1-2 7.90 7.85 7.90 − 8.28 TM1-3 7.71 7.80 7.92 − − TM1-4 − 7.86 − − − A 8.31 8.02 7.88 8.44 8.23 TM2-1 7.91 8.16 7.94 8.30 TM2-2 8.17 7.72 7.96 8.15 TM2-3 7.64 − 8.27 TM2-4 7.84 − 8.30 TM3-1 7.35 7.83 7.68 7.87 TM3-1’ TM3-2 8.03 7.94 8.38 7.87 8.17 TM3-3 7.71 7.78 − TM3-4’ 7.80 7.94 − 8.30 TM4-1 8.70 8.47 8.11 8.03 TM4-2 8.13 7.85 TM4-3 7.89 7.79 8.55 TM4-4 7.98 7.86 8.05 − 8.55 試料採取時に計測できなかった場合は空欄,試料採取をしていない場合 は−で示す. 表5 試料採取地点の DO
地点 May, 2012 Jul., 2012 Sep., 2012 Dec., 2012 Feb., 2013
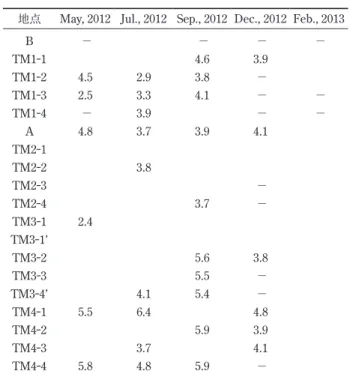
B − − − − TM1-1 4.6 3.9 TM1-2 4.5 2.9 3.8 − TM1-3 2.5 3.3 4.1 − − TM1-4 − 3.9 − − A 4.8 3.7 3.9 4.1 TM2-1 TM2-2 3.8 TM2-3 − TM2-4 3.7 − TM3-1 2.4 TM3-1’ TM3-2 5.6 3.8 TM3-3 5.5 − TM3-4’ 4.1 5.4 − TM4-1 5.5 6.4 4.8 TM4-2 5.9 3.9 TM4-3 3.7 4.1 TM4-4 5.8 4.8 5.9 − 単位はmg/l.試料採取時に計測できなかった場合は空欄,試料採取を していない場合は−で示す. 表6 2012年 5月に多摩川河口干潟で産出した貝形虫種と個体数 B 1-1 1-2 1-3 1-4 A 2-1 2-2 2-3 2-4 3-1 3-1’ 3-2 3-3 3-4’ 4-1 4-2 4-3 4-4 Darwinulid sp. – – /1 Candonid sp. – – /1 Heterocypris salina – – /1 Cypridopsis vidua – – Cypridid sp. – – /1 Perissocytheridea? japonica – – Pontocythere miurensis – /2 /2 – /1 Pontocythere xiphoidea – – Ishizakiella miurensis – 35/3 35/2 5/2 – 408/10 227/8 172/3 219/15 83/6 68/2 85/1 386/24 66/ 5/ 148/15 24/1 20/5 49/2 Spinileberis pulchra – – Spinileberis quadriaculeata – – Aurila sp. – – /1 Angulicytherura miii – – Hemicytherura sp. – – Loxoconcha kosugii – – Loxoconcha pulchra – /3 /1 /1 – Cytheromorpha acupunctata – – Nipponocythere sp. – /2 – 不明 – – –は試料採取をしていないことを示す.生体/遺骸として示す.
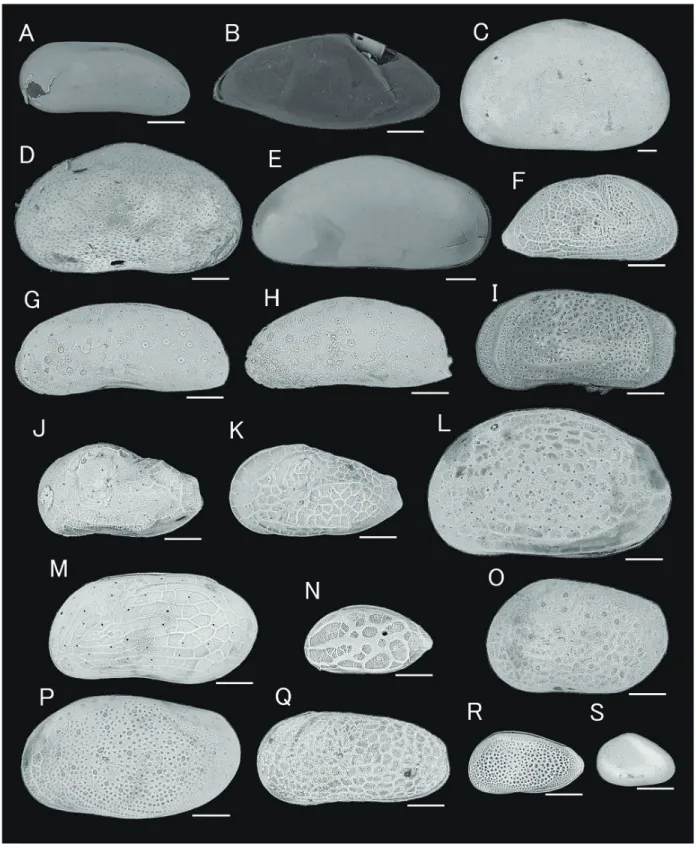
図3 産出した貝形虫類のSEM画像
スケールはそれぞれ100μmを示す.標本は静岡大学キャンパスミュージアムに登録・保管されている.A, Darwinulid sp., carapace in right lateral view (SUM-CO-2393), point TM4-4; B, Candonid sp., carapace in left lateral view (SUM-CO-2394), point TM3-4’; C, Heterocypris salina, left valve in external lateral view (SUM-CO-2395), point A; D, Cypridopsis vidua, right valve in external lateral view (SUM-CO-2396), point TM4-3; E, Cypridid sp., carapace in right lateral view (SUM-CO-2397), point TM3-1; F, Perissocytheridea? japonica, right valve in external lateral view (SUM-CO-2398), point B; G, Pontocythere miurensis, carapace in left lateral view (SUM-CO-2399), point B ; H, Pontocythere xiphoidea, carapace in left lateral view (SUM-CO-2400), point B ; I, Ishizakiella miurensis, carapace in left lateral view (SUM-CO-2401), point TM1-2; J, Spinileberis pulchra, carapace in left lateral view (SUM-CO-2402), point TM1-2; K, Spinileberis
quadriaculeata, carapace in left lateral view (SUM-CO-2403), point TM3-4’; L, Aurila sp. carapace in left lateral view (SUM-CO-2404), point TM1-1; M,
Angulicytherura miii, carapace in left lateral view (SUM-CO-2405), point TM1-1; N, Hemicytherura sp., carapace in left lateral view (SUM-CO-2406), point TM1-3;
O, Loxoconcha kosugii, carapace in left lateral view (SUM-CO-2407), point B; P, Loxoconcha pulchra, carapace in left lateral view (SUM-CO-2408), point B; Q,
Cytheromorpha acupunctata, carapace in left lateral view (SUM-CO-2409), point TM3-3; R, Nipponocythere sp., carapace in left lateral view (SUM-CO-2410), point
Angulicytherura miii, Loxoconcha kosugii, L. pulchra が 産 出 し た( 表7)。9月はP. miurensis, Spinileberis pulchra, A. miii, Hemicytherura sp., L. pulchra, Cytheromorpha acupunctata と同定不能の種が産出した(表 8 )。12 月は I. miurensis と Nipponocythere sp. 以 外 は 産 出 し な か っ た (表 9)。2013年 2月はCypridopsis vidua, Aurila sp., A. miii, C. acupunctata が産出した (表 10)。
年7月に地点 TM1-1で 5 個体,9月に地点 TM1-1で 6 個
体,地点Aで2個体,12月に地点 Aで 1 個体が産出し
た。I. miurensis と Nipponocythere sp. 以外の種はすべて 遺骸で,個体数はごく僅かで,5月はPontocythere miu-rensis, Aurila sp., Loxoconcha pulchra と淡水生 4 種が産 出した (表 6)。7月は,Perissocytheridea? japonica, Ponto-cythere miurensis, P. xiphoidea, Spinileberis quadriaculeata,
表8 2012年 9月に多摩川河口干潟で産出した貝形虫種と個体数 B 1-1 1-2 1-3 1-4 A 2-1 2-2 2-3 2-4 3-1 3-1’ 3-2 3-3 3-4’ 4-1 4-2 4-3 4-4 Darwinulid sp. – – Candonid sp. – – Heterocypris salina – – Cypridopsis vidua – – Cypridid sp. – – Perissocytheridea? japonica – – Pontocythere miurensis – /1 /1 /5 – /2 Pontocythere xiphoidea – – Ishizakiella miurensis – 186/68 189/47 98/54 – 154/29 152/18 139/13 153/14 205/56 149/16 331/13 222/20 206/13 64/41 170/5 90/8 4/2 8/3 Spinileberis pulchra – /1 – Spinileberis quadriaculeata – – Aurila sp. – – Angulicytherura miii – – /1 Hemicytherura sp. – /1 – Loxoconcha kosugii – – Loxoconcha pulchra – /3 – Cytheromorpha acupunctata – – /1 Nipponocythere sp. – 6/21 – 2/ /1 /5 不明 – – /1 –は試料採取をしていないことを示す.生体/遺骸として示す. 表7 2012年 7月に多摩川河口干潟で産出した貝形虫種と個体数 B 1-1 1-2 1-3 1-4 A 2-1 2-2 2-3 2-4 3-1 3-1’ 3-2 3-3 3-4’ 4-1 4-2 4-3 4-4 Darwinulid sp. Candonid sp. Heterocypris salina Cypridopsis vidua Cypridid sp. Perissocytheridea? japonica /2 /1 Pontocythere miurensis /4 /1 /1 /1 /2 Pontocythere xiphoidea /2 /1 Ishizakiella miurensis 7/2 182/53 281/46 79/22 9/3 241/30 185/32 181/13 222/24 171/19 170/9 171/15 197/19 123/26 31/4 229/20 135/11 20/1 12/7 Spinileberis pulchra Spinileberis quadriaculeata /1 Aurila sp. Angulicytherura miii /1 /1 Hemicytherura sp. Loxoconcha kosugii /1 Loxoconcha pulchra /8 /2 /1 /1 Cytheromorpha acupunctata Nipponocythere sp. 5/18 /1 /2 不明 –は試料採取をしていないことを示す.生体/遺骸として示す.
ponocythere 属は,東京湾およびその周辺では,N. bicari-nata が,神奈川県川崎市の完新統 (池谷ほか,1987), 埼玉県三郷市の完新統 (中尾ほか,2008;堀越ほか, 2009),埼玉県菖蒲町の更新統 (入月ほか,2011),神奈 川県横浜市の更新統 (Ozawa, 2009),千葉県木更津市と その周辺の更新統 (Yajima, 1978, 1982) から化石が報告 されている。また,N. hastata が千葉県館山市の完新統 (Frydl, 1982; 田中ほか,2012) から化石が報告されてい 4 .考察 4-1)多摩川河口干潟の貝形虫相 多摩川河口の生体貝形虫相は,年間を通してI. miu-rensis が独占し,Nipponocythere sp. が数個体産出する種 多様性の低い群集であることがわかった。I. miurensis は,九州から本州の河口干潟等汽水域の砂泥底の表層に 普 遍 的 に み ら れ る 種 で あ る (Tsukagoshi, 1994)。Nip-表10 2013年 2月に多摩川河口干潟で産出した貝形虫種と個体数 B 1-1 1-2 1-3 1-4 A 2-1 2-2 2-3 2-4 3-1 3-1’ 3-2 3-3 3-4’ 4-1 4-2 4-3 4-4 Darwinulid sp. – – – Candonid sp. – – – Heterocypris salina – – – Cypridopsis vidua – – – /1 Cypridid sp. – – – Perissocytheridea? japonica – – – Pontocythere miurensis – – – Pontocythere xiphoidea – – – Ishizakiella miurensis – 56/12 42/5 – – 70/12 160/51 117/13 143/25 140/25 25/1 14/ 189/20 137/7 160/4 209/31 12/ 76/4 136/6 Spinileberis pulchra – – – Spinileberis quadriaculeata – – – Aurila sp.. – /1 – – Angulicytherura miii – /1 /1 – – Hemicytherura sp. – – – Loxoconcha kosugii – – – Loxoconcha pulchra – – – Cytheromorpha acupunctata – – – /1 Nipponocythere sp. – /3 – – 不明 – – – –は試料採取をしていないことを示す.生体/遺骸として示す. 表9 2012年12月に多摩川河口干潟で産出した貝形虫種と個体数 B 1-1 1-2 1-3 1-4 A 2-1 2-2 2-3 2-4 3-1 3-1’ 3-2 3-3 3-4’ 4-1 4-2 4-3 4-4 Darwinulid sp. – – – – – – – – – Candonid sp. – – – – – – – – – Heterocypris salina – – – – – – – – – Cypridopsis vidua – – – – – – – – – Cypridid sp. – – – – – – – – – Perissocytheridea? japonica – – – – – – – – – Pontocythere miurensis – – – – – – – – – Pontocythere xiphoidea – – – – – – – – – Ishizakiella miurensis – 75/16 – – – 70/34 7/3 196/6 – – 25/19 50/7 149/13 – – 132/10 13/3 1/ – Spinileberis pulchra – – – – – – – – – Spinileberis quadriaculeata – – – – – – – – – Aurila sp. – – – – – – – – – Angulicytherura miii – – – – – – – – – Hemicytherura sp. – – – – – – – – – Loxoconcha kosugii – – – – – – – – – Loxoconcha pulchra – – – – – – – – – Cytheromorpha acupunctata – – – – – – – – – Nipponocythere sp. – /13 – – – 1/5 – – – – – 不明 – – – – – – – – – –は試料採取をしていないことを示す.生体/遺骸として示す.
gii は塩濃度の変化に耐性がある広塩性種であるといえ る (中尾・塚越,2008)。一方,溶存酸素は,多摩川河口 干潟では3.8–6.4mg/l (調査地全体では2.5–6.4 mg/l), 小櫃川河口干潟では4.4–13.8mg/l (中尾・塚越,2008) であった。多摩川河口干潟の溶存酸素の方が小櫃川河口 干 潟 の そ れ よ り 低 く,I. miurensis は L. kosugii, D. mu-kaishimensis, A. miii より低溶存酸素の環境に耐性をもつ 可能性が考えられる。多摩川河口域では貧酸素水塊がし ばしば観測されており (八木ほか,2007),また,大井人 工干潟では,貧酸素水が干潟に侵入したという報告もあ る (中村ほか,2010)。多摩川河口干潟でも,河口域の貧 酸素水が干潟に侵入し,干潟の溶存酸素を低下させ,貝 形虫相に影響を与えている可能性がある。I. miurensis の多摩川河口干潟での独占は,本種の貧酸素状態への耐 性の高さが一因としてあげられる。 今回遺骸のみ確認されたP.? japonica, P. miurensis, P. xiphoidea, S. pulchra, S. quadriaculeata, A. miii, L. kosu-gii, L. pulchra, C. acupunctata は , 小櫃川河口干潟では生 体 が 報 告 さ れ て い る (Nakao and Tsukagoshi, 2002; 中
尾・塚越,2008)。多摩川河口域での調査範囲を広げる ことで,生体個体が見つかる可能性があり,多摩川河口 干潟の貝形虫相がI. miurensis と Nipponocythere sp. のみ で構成されると結論づけるには,今後のさらなる調査が 必要である。底生生物相の分布を規制する主な要因とし て,汚濁,底質の含泥率,酸化還元電位,強熱減量, COD,硫化物量,干潟の傾斜,水域の違いが報告され ている (例:北園ほか,2001;小林ほか,2003;安藤・ 川井,2007;石井ほか,2012;堀越ほか,2012)。本研究 ではデータがないため,貝形虫相とそれらの要因との関 係には言及できなかった。しかし,現在の沿岸環境と今 後の変化を記録するためにも,貝形虫相とこれらの環境 因子との関係を明らかにし,調査範囲を広げ,継続して 調査していく事が重要である。 4-3)I. miurensisの分布 多摩川河口干潟において,I. miurensis はどの地点で も一年を通して生息していたが,産出個体数は地点に よって大きく異なった。本種の干潟での分布の特性を明 らかにするため,面積10cm×20cm,深さ1cm (200cm3) あたりに換算した各地点の産出個体数を図4 に示す。河 川流軸方向の違いに直目すると,ラインTM2が年間を 通して最も個体数が多く,I. miurensis の主要な分布域 といえる。ラインTM2は,干潮時でも水たまりや澪筋 があり,水がひたひたとした状態で,堆積物表面がデト リタスに覆われている (表 1,図 2 のB)。I. miurensisは, る。多摩川河口干潟で見つかった種は,この2 種と殻の 形態が明確に異なり,また,他のNipponocythere 属の既 知種とも殻の形態が異なっており,未記載種と考えられ るが,これについては稿を改めて報告する。 Nipponocy-there sp. の 生 態 は 不 明 で あ る が, 生 体 が 下 流 の 地 点 (TM1-1,A)からのみ産出したことから,調査地より 海側に生息する可能性が高い。さらに,補足的にライン TM-1より海側の干潟上に設定した地点Bからは産出し なかった事を考慮すると,本種の生息域は,潮下帯より 深い場所であり,潮汐や波の作用によって調査地点に生 体が偶然入り込んだ可能性が高いといえる。その他の種 は,遺骸が数個体のみ産出し,年間を通して生体が一度 も確認できなかった事から,調査地域外で生息していた ものが死後に運搬され,流れ込んだものであるといえ る。 4-2)小櫃川河口干潟の貝形虫相との比較 次に,東京湾を挟み対岸に位置する小櫃川河口干潟の 貝形虫相と比較する。小櫃川河口干潟では13属16種の
生体が産出しており (Nakao and Tsukagoshi, 2002),わ
ずか2 属 2 種の多摩川河口の貝形虫相の種多様性は,小 櫃川河口干潟のそれと比較すると著しく低い。多摩川河 口干潟が,川岸に沿った比較的小規模の干潟であるのに 対し,小櫃川河口干潟は川岸に沿った干潟のみならず, いわゆる後浜と呼ばれる塩性湿地や,砂質前浜干潟,葦 原といった変化に富む地形からなる。二つの干潟の種多 様性の違いは,このような地形的な変化の豊富さの違い よると考えられ,干潟全体の貝形虫相を比較するのは適 切ではない。しかしながら,小櫃河口干潟では,I. miu-rensis が多く産出する地点でも,Loxoconcha kosugii, Dol-erocypria mukaishimensis, 季節によっては Angulicytherura miii が生息しており (中尾・塚越,2008),I. miurensis 1 種が独占することはない。さらに,多摩川河口右岸側の 干潟では,泥干潟や葦原の典型的な生物相が良好な状態 で保たれていること (西・田中,2006),調査地である左 岸側においても葦原が存在していることから,I. miu-rensis の独占状態の説明を,単に地形の単調さだけに求 めることはできない。 多摩川河口干潟でI. miurensis の個体数が特に多く産 出した地点 (TM2-1, 2, 3, TM3-2, TM4-1) と,中尾・塚 越 (2008) によって報告された小櫃川河口干潟でI. miu-rensis が年間を通して寡占した地点 (中尾・塚越,2008 の 地 点OB-24, OB-25, OB-A) の 塩 濃 度 は, そ れ ぞ れ 4–29ppt,2.5–25pptで,ともに塩濃度の変化の幅が大き い。このような場所に多く生息するI. miurensis, L.
kosu-も時折波をかぶる事がラインTM2と異なる点として挙 げられる。小櫃川河口干潟でも汀線に近い地点には貝形 虫類の産出個体数が少ないことが報告されている (中 尾・塚越,2008)。ラインTM1とラインTM2の個体数の 違いは,波の営力というエネルギー環境の違いが原因の 一つとしてあげられる。すなわち,波の営力が大きい場 所では,堆積物が水流で動きやすく不安定で,デトリタ スが容易に移動するため,I. miurensis の個体数が少な くなる。反対に波の影響を受けず,堆積物が動きにくく 安定した場所ではデトリタスも動きにくく,I. miurensis 水深1m未満で,フロッキュレント層を伴う泥質砂底 で,河口付近の低塩分域に生息することが報告されてい る (例:Tsukagoshi, 1994)。このフロッキュレント層と デトリタスは,同じものをさす。これらの事から,多摩 川河口干潟でのI. miurensis の産出個体数の違いは,干 潟上でのデトリタスの分布に左右されると考えられる。 ラインTM1もラインTM2と同様にデトリタスに覆わ れ,水がひたひたとした状態にあるが (表 1,図2A),ラ インTM2と比較するとI. miurensisの産出個体数は少な かった。ラインTM1は干潮時の汀線に近く,干潮時で N TM4 TM1 TM2 TM3 4-1 4-4 4-2 4-3 3-1’ 3-1 3-33-2 3-4’ 2-22-1 2-3 2-4 A 1-1 1-2 1-3 1-4 20m 200m
Sampling points
0May, 2012
6000 3000Jul., 2012
~ 12608 0 6000 3000Sep., 2012
19456 8896 0 6000 3000Dec., 2012
6272 0 6000 3000Feb., 2013
0 6000 3000 図4 多摩川河口干潟におけるIshizakiella miurensis の分布 柱の高さは堆積物試料200cm3あたりの産出個体数を示す.柱の上の数字は,6000個体以上産出した場合の個体数を,点線は最大干潮時の おおよその汀線を示す.白抜きの四角は,試料を採取していない事を示す.ここには地点Bは示していない.4-4)I. miurensisの個体群動態 貝形虫類は脱皮成長するため,背甲の大きさによって 脱皮齢がわかる。慣例的に,成体をA (Adult) とし,そ こから順にさかのぼってA-1齢,A-2齢,A-3齢……と 呼 ぶ。I. miurensis で は A-8齢 ま で 識 別 さ れ て い る が (Tsukagoshi, 1994),今回の堆積物試料からは,成体か らA-7齢まで,8 つの脱皮齢のみが確認された (図 5)。
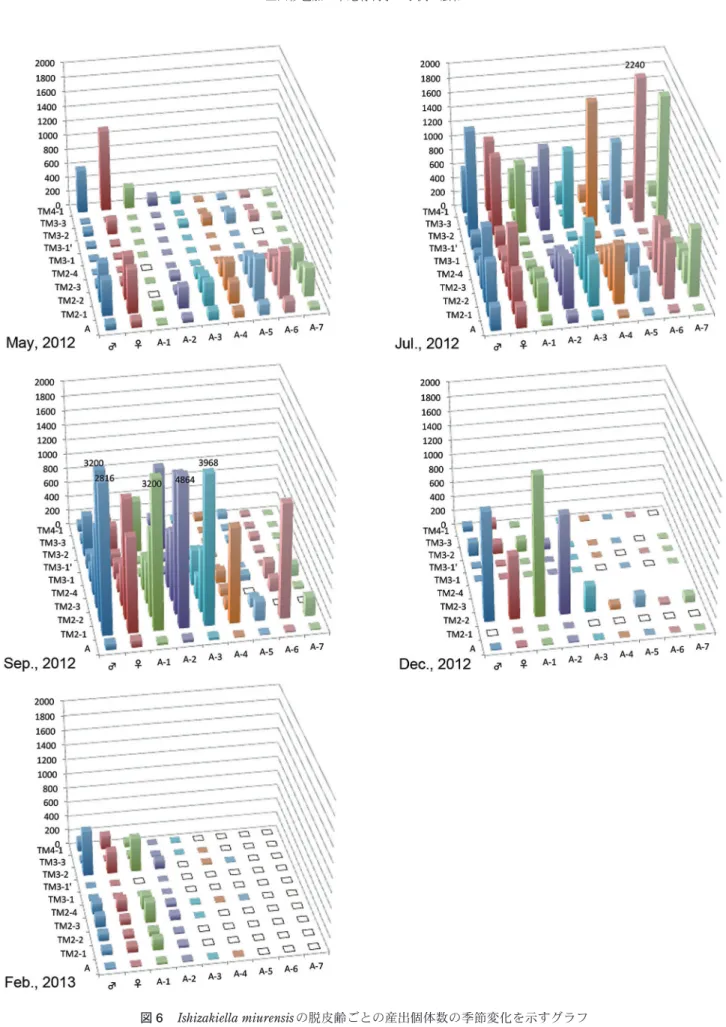
I. miurensis の個体群動態を明らかにするため,I. miu-rensis が年間を通して議論に十分な個体数が産出した 10 地点 (図 6) を選定した。面積10cm×20cm,深さ 1cm (200cm3) あたりに換算した各地点の脱皮齢ごとの産出 個体数 (成体のみ雄と雌を識別した) の季節変化を図 6 に,模式化したライフサイクルを図7 に示す。個体数の 増減,齢構成は地点によって差があるものの,全体とし て以下のような特性があると言える。2012年 5月は, A-1齢の個体数が少なく (左から 3 列目の緑色で示され た脱皮齢),成体とA-2齢以下の間にギャップが存在す る。成体の個体数は,幼体と比較すると多く,越冬した 個体であると考えられる。また,A-2齢からA-7齢の個体 数が増え始めており,越冬個体によって,繁殖がすでに 開始されている可能性がある。7月は,全体に個体数が 多くなり,A-1齢の個体数も増えたため,A-1齢のギャッ プがなくなった。成体の増加とA-1齢のギャップの消失 は,5月により若い脱皮齢だった個体が成長したためと 説明できる。この時の成体は,越冬個体と,春に新しく 成長した個体が混合する集団であるといえる。また,若 い脱皮齢の著しい増加は,I. miurensis の繁殖がこの時 期に盛んであることを示す。春から初夏に成長した個体 が成熟し,繁殖期を迎えていると考えられる。地点 TM4-1のみ 5月,7月は他の地点と異なった。すなわ ち,成体が最も多く,より若い幼体に向けて少なくなっ た。地点TM4-1は,調査地点の中で,最も上流にあり, 泥質で,干出した干潟の水たまりである。地点TM4-1 の齢構成は,堆積環境の違いが影響している可能性があ る。9月になると,A-7齢からA-5齢の産出個体数が減 り,A-4齢以降の脱皮齢が多くなる。これは, 7月のよ り若い脱皮齢の個体の成長と,繁殖のピークが過ぎた事 を示すと考えられる。12月になると全体に個体数が減 少し,A-3齢より若い個体は産出しない地点が増加す る。さらに,2月になると,A-3齢より若い個体はほぼ 無産出になる。12月,2月の幼体の著しい減少は,冬季 は繁殖が行われない時期であることを示唆する。さら に,2月に成体個体およびA-1齢,A-2齢が産出してい ることから,I. miurensis は成体と A-1齢,A-2齢で越冬 すると考えられる。このように,一年を通してみると, の生活の場が安定するため,個体数が多くなると考えら れる。また,産出個体数が多いラインTM2では,岸側 で個体数が多く,川側へ行くほど個体数が少なくなると いう分布を示した。これは,岸側ほど河川 (波) の営力 を受けにくく,反対に岸から離れるほど河川 (波) の営 力を受けやすい,あるいは受ける時間が長いためと説明 できる。 ラインTM3では,一番岸側のTM3-1とTM3-1’では個 体数が少なく,岸から20mの地点TM3-2の個体数が一 番 多 か っ た。 地 点TM3-1とTM3-1’は堆積物表面がし まった状態にあり,デトリタスは少ない (図2D) ため個 体数が少ないと推測できる。一方,地点TM3-2はライ ンTM2の地点と同様,小さな水たまりや澪筋によって 堆積物表面に,水がひたひたとある状態で,デトリタス に覆われているため個体数が多いと推測できる。また, ラインTM3はライン TM2と比べると干潟の幅が短い。 すなわち,堤防から約40mの地点TM3-3は,同じく40 mの地点TM2-3と比べて,汀線までの距離が短く,さ らに,地点TM3-4’は試料採取時の汀線のすぐ陸側であ る。このため,地点TM3-3,TM3-4’は,波の影響を受 けやすく,個体数が少ないと説明できる。ラインTM3 においても,堆積物表面に水がひたひたとした状態で, デトリタスが多いため,水の流れ (波) の営力を受けに く い 所 に 多 く 生 息 す る と 考 え ら れ る。2012年 7月に TM3-2で個体数が最大になったのはデトリタスが多く 分布していたためである可能性が高い。 ラインTM4は他の調査ラインと比較すると,泥粒子 が多くぬかるんでいた。実際に試料を篩で洗った際も, 63 μmの篩上に残る砂の量が最も少なかった。全体に干 出しており,地点TM4-1のみに水たまりがあった。水 たまりのない地点TM4-2,TM4-3,TM4-4,TM3-1’に産 出した個体数が少ない事から,I. miurensis には,小さ な水たまりや澪筋があり,堆積物表面に水がひたひたと する程度の水分が必要であると考えられる。この事は, 小櫃川河口干潟での産出報告(中尾・塚越,2008)とも 一致する。ただし,I. miurensis の産出個体数が少ない 地 点TM3-3, TM3-4’, TM4-2, TM4-3, TM4-4は,泥が多 い地点とも言える。これらの地点での産出個体数の少な さは,底質の違い(含泥率の高さ)によるものなのか, 水たまりの有無によるものかまでは本研究のデータでは 判断することはできなかった。詳細な議論のためには, ライン上での底質粒度,水分量,有機物量といった物理 的環境の違いを今後は明らかにしなければならない。
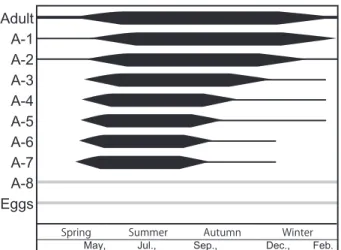
夏の繁殖期の終わりに生まれた個体であるか,あるいは 冬季にも僅かながら繁殖が行われていることを示唆す る。いずれにせよ,冬季に背甲の中に卵と若い脱皮齢が 観察されるにもかかわらず,背甲の外では若い齢が観察 されなくなる事から,I. miurensis は気温の低い期間は 成長を止め雌の背甲内にとどまり,春先に雌の背甲から 出て,水温の上昇に伴って成長を開始し,繁殖すると考 えられる。夏から秋にかけての個体群は,数世代が混合 した群集といえる。 小櫃川河口干潟のI. miurensis では,一年中,雌の背 越冬した個体(成体あるいはA-1齢,A-2齢)が産み出 した世代と,春に成長を開始して成熟した個体が,夏季 に新しい世代を作っていると捉えることができた。しか しながら,夏季にいくつの世代が産まれるのかまでは今 回の結果からは明らかにできなかった。 一方,冬季に若い幼体個体は見られなかったが,雌の 背甲の中には,12月,2月にも,卵とA-8齢,A-7齢が 僅かながら確認された。成体,A-1齢,A-2齢のみなら ず,卵と初期幼体が,雌の背甲の中で越冬する可能性が 考えられる (図 7 に灰色で示す)。これら初期幼体は,
図5 Ishizakiella miurensis の成体から A-7 齢までの SEM 画像
スケールは100 μmを示す.Female, carapace in left lateral view (SUM-CO-2412), point TM1-2; male, carapace in left lateral view (SUM-CO-2401); point TM1-2; A-1, carapace in left lateral view (SUM-CO-2413), point TM2-1; A-2, carapace in left lateral view (SUM-CO-2414), point TM2-1; A-3, carapace in left lateral view (SUM-CO-2415), point TM2-1; A-4, carapace in left lateral view (SUM-CO-2416); point TM2-1; A-5, carapace in left lateral view (SUM-CO-2417), point TM2-1; A-6, carapace in left lateral view (SUM-CO-2418), point TM2-1; A-7, carapace in left lateral view (SUM-CO-2419), point TM2-1.
図6 Ishizakiella miurensis の脱皮齢ごとの産出個体数の季節変化を示すグラフ
x軸に脱皮齢,y軸に地点,z軸に個体数を示す.個体数は,堆積物表面 10cm×20cm,深さ 1cm (堆積200cm3) あたりの個体数に換算し,成体のみ雌雄を 個別に示す.個体数の変化が見やすいように,z軸の最大値を 2000個体とし,2000個体を超えた場合は個体数を数値で示した.底面の空白は試料が採取 出来なかったこと,白抜きの四角は産出個体数が0 であることを示す.
は,種間関係が生活史に与える影響に関する研究の好例 になると考える。 5 .まとめ 多摩川河口干潟の貝形虫相が調査され,Ishizakiella miurensis と Nipponocythere sp. のみが生息し,さらに I. miurensis がほぼ独占する種多様性が著しく低い群集で あることが初めて明らかにされた。I. miurensis の独占 は,貧酸素状態によってもたらされた可能性がある。ま たI. miurensis の分布は,デトリタスの分布に規制され る可能性がある。遺骸では,東京湾を挟んだ小櫃川河口 干潟に生息する種が多摩川河口干潟にも産出するため, 調査域を広げることで,成体の報告された2種以外が生 息している可能性が残された。さらに,I. miurensis の 生活史が明らかにされ,冬季には成長を止め,成体ある いはA-1齢,A-2齢,背甲の中の卵とA-7齢,A-8齢で越 冬することが初めて示された。 謝辞 本研究を進めるにあたり,日本大学文理学部地球科学科の 小坂和夫教授には,研究の機会をあたえて頂き,研究を進め るにあたり,様々な助言を頂いた。また,毎回の調査にもご 協力頂いた。日本大学文理学部地球システム科学科卒業生, 和田絵里香氏と表野竜馬氏には野外調査にご協力頂いた。静 岡大学理学部塚越哲教授には原稿を読んで頂き,有益なコメ ントを頂くと共に,分類に関するご助言を頂いた。また,文 献収集にもご協力頂いた。国立科学博物館の蛭田眞平博士に は,淡水生貝形虫類の同定に関するご助言を頂いた。国土交 通省京浜河川事務所田園調布出張所の皆様には,多摩川河口 干潟での調査の許可を頂いた。その他多くの方々のご協力に 感謝し,ここに御礼を申し上げる。 甲の中に卵が観察され,若い脱皮齢が春に増加し,夏に 発生後期の幼体が群集の中で優占するようになり,冬季 は個体数が減少し,雌の成体個体が個体群の中で優占す ることが報告されている(Tsukagoshi, 1994)。小櫃川 河口干潟ではI. miurensis が多く産出する場所でも,他 の種が同所的に生息している(中尾・塚越,2008)。一 方,多摩川河口干潟ではI. miurensis が独占状態である ことが本研究によって明らかにされた。Horne(1983) は,海域と汽水域の貝形虫の年間の生活史を総括し,そ れらの生活史には,水温(とそれに関連する食物量)の 他に,多種との競争が影響を与えると述べている。東京 湾西岸と東岸に生息する I. miurensisの生活史の比較 安藤晴夫・川井利雄 2007.東京湾内における底生生物生息 状況の解析結果について,東京都環境科学研究所年報 2007: 77-84.(http://www.tokyokankyo.jp/kankyoken_ contents/report-news/2007/Dojyo_Mizu-4.pdf)
Bodergat, A. and Ikeya, N. 1988. Distribution of Recent Ostra-coda in Ise and Mikawa Bays, Pacific coast of central Ja-pan. In Hanai, T., Ikeya, N. and Ishizaki, K. (eds.) Evolutionary Biology on Ostracoda. Proceedings of the Ninth International Symposium on Ostracoda, pp.413-428, Kodansha, Tokyo.
Frydl, P. M. 1982. Holocene ostracods in the southern Boso Peninsula. The University Museum, the University of Tokyo, Bulletin, 20 : 61-140. 堀越彩香・青木 茂・岡本 研 2012.東京湾多摩川河口干 潟におけるムロミスナウミナナフシの分布と生息環境特 性,日本ベントス学会,66:71-81. 堀越英之・中尾有利子・遠藤邦彦 2009.埼玉県三郷市の沖 積層から産出した貝形虫化石群集を指標とする古環境変 引用文献 遷.日本大学文理学部自然科学研究所研究紀要,44: 149-157.
Horne, D. J. 1983. Life-cycle of podocopid Ostracoda –a review (with particular reference to marine and brackish-water species). In Maddocks, R. F. (ed.) Applications of Ostraco-da: Proceedings of the Eighth International Symposium on Ostracoda, pp.581-590, Department of Geosciences, Uni-versity of Houston, Houston.
Ikeya, N. and Hanai, T. 1982. Ecology of Recent ostracods in the Hamana-ko region, the Pacific coast of Japan. The Uni-versity Museum, UniUni-versity of Tokyo, Bulletin, 20 : 15-59. 池谷仙之・長谷川浩志・鹿島徹子 1987.3.介形虫化石の
分析―川崎市街地 (神奈川県) の完新世介形虫群集―. 松島義章・川崎市都市地質研究会 (編),川崎市内沖積 層の総合研究,pp.51-64.川崎市博物館資料収集委員 会,川崎.
Ikeya, N. and Itoh, H. 1991. Recent Ostracoda from the Sendai Bay region, Pacific coast of northeastern Japan. Report of Spring Summer Autumn Winter
Adult A-1 A-2 A-3 A-4 A-5 A-6 A-7 A-8 Eggs
May, Jul., Sep., Dec., Feb. 図7 Ishizakiella miurensis の生活史の模式図 図はHorne (1983) を参考に作成した.成体から A-7齢までの線の太さ は,個体数の多さを相対的に示す.A-8齢と卵は,雌の背甲内に確認さ れたものを参考にし,灰色で示す.ただし,背甲の中の個体は計数して いないため,灰色で示された線は雌の背甲内の卵とA-8齢の有無のみを 示す.
け る 底 生 動 物 群 集 の ハ ビ タ ッ ト 分 析. 日 緑 工 誌,29 (1):62-67.
中尾有利子・中西利典・木村克己 2008.中川低地南部の沖 積層から産出した貝形虫化石.日本大学文理学部自然科 学研究所研究紀要,43:277-286.
Nakao, Y. and Tsukagoshi, A. 2002. Brackish-water Ostracoda (Crustacea) from the Obitsu River Estuary, central Japan. Species Diversity, 7 (1) : 67-115. 中尾有利子・塚越 哲 2008.干潟域に生息する貝形虫群集 の季節変化と経年変化.日本大学文理学部自然科学研究 所研究紀要,43:249-275. 中村泰男・東 博紀・牧 秀明 2010.貧酸素水塊の形成機 構と生物への影響評価に関する研究(特別研究)平成 19∼21年度.国立環境研究所特別研究報告 SR-93-2010, 国立環境研究所,35pp. 西栄二郎・田中克彦 2006.多摩川河口川崎市側の干潟にお ける底生動物相.神奈川自然誌資料,27:77-80. Ozawa, H. 2009. Middle Pleistocene ostracods from the
Na-ganuma Formation in the Sagami Group, Kanagawa Pre-fecture, central Japan: palaeo-biogeographical significance of the bay fauna in Northwest Pacific margin. Paleontologi-cal Research, 13, 231-244.
田中源吾・金子 稔・大石雅子・青木貴子 2012.千葉県の 完新統沼層より産出した介形虫化石群, 群馬県立自然史 博物館研究報告, 16:111-118.
Tsukagoshi, A. 1994. Natural history of the brackish-water os-tracode genus Ishizakiella from East Asia: Evidence for heterochrony. Journal of Crustacean Biology, 14 (2) : 295-313.
八木 宏・江連伸明・井瀬 肇・Pokavanich, T.・灘岡和夫・ 諸星一信・古土井健・有路隆一・森重輝政・小林 聡 2007.東京湾羽田周辺水域における貧酸素水塊と懸濁物 質の時空間変動特性.海岸工学論文集,54:1036-1040. Yajima, M. 1978. Quaternary Ostracoda from Kisarazu near
Tokyo. Transactions and Proceedings of the Palaeontologi-cal Society of Japan, New Series, 112 : 371-409.
Yajima, M. 1982. Late Pleistocene Ostracoda from the Boso Peninsula, Central Japan. The University Museum, the Uni-versity of Tokyo, Bulletin, 20 : 141-227.
Yasuhara, M. and Irizuki T. 2001. Recent Ostracoda from the Northeastern part of Osaka Bay, Southwestern Japan. Journal of Geosciences, Osaka City University, 44 : 57-75. Faculty of Science, Shizuoka University, 25 : 93-145.
入月俊明・後燈明あすみ・河野重範・吉岡 薫・野村律夫 2009.兵庫県相生市沖の播磨灘北部における現生貝形虫 群集と環境要因との関連.島根大学地球資源環境学研究 報告,28:1-9. 入月俊明・納谷友規・山口正秋・水野清秀 2011.中期更新 世(MIS 11およびMIS 9)における古東京湾湾奥の環境 変遷―埼玉県菖蒲コアの下総層群貝形虫化石の群集解 析―.地質学雑誌,117 (1):35-52.
Irizuki, T., Seto, K. and Nomura, R. 2008: The impact of fish farming and bank construction on Ostracoda in Uranouchi Bay on the Pacific coast of southwest Japan-Faunal chang-es between 1954 and 2002/2005. Paleontological Rchang-esearch, 12 : 283-302.
Irizuki, T., Takata, H., and Ishida, K. 2006. Recent Ostracoda from Urauchi Bay, Kamikoshiki-jima Island, Kagoshima Prefecture, southwestern Japan. LAGUNA, 13 : 13-28. Ishizaki, K. 1968. Ostracodes from Uranouchi Bay, Kochi
Pre-fecture, Japan. The Science Reports of the Tohoku University, Sendai, Second Series (Geology), 40 (1) : 1-45.
Ishizaki, K. 1969. Ostracoda from Shinjiko and Nakanoumi, Shimane Prefecture, western Honshu, Japan. The Science Reports of the Tohoku University, Sendai, Second Series (Geology), 41 (2) : 197-224.
Ishizaki, K. 1971. Ostracoda from Aomori Bay, Aomori Prefec-ture, Northeast Honshu, Japan. The Science Reports of the Tohoku University, Sendai, Second Series (Geology), 43 (1) : 59-97.
石井裕一・和波一夫・木瀬晴美・安藤晴夫 2012.都市河川 河口域の底生動物と底質環境との関係.東京都環境科学 研究所年報2012 : 9-16.
Kamiya, T., Ozawa, H. and Obata, M. 2001. Quaternary and Re-cent marine Ostracoda in Hokuriku district, the Japan Sea coast. In Ikeya, N. (ed.), Field Excursion Guidebook; 14th International Symposium of Ostracoda, pp.73-106, Organis-ing Committee of 14th ISO, Shizuoka.
北 園 芳 人・ 鈴 木 敦 巳・ 林 泰 弘・ 喜 津 木 郁 人・ 滝 川 清 2001.有明海と八代海の干潟における底質特性と底生動 物の豊かさに関する研究.海岸工学論文集,48:1131-1135. 小林達明・野田泰一・鈴木奈津子・稲田陽介・清水良憲・桑 原 茜・高橋輝昌 2003.東京湾の自然渚·人工渚にお