大腸菌はモデル生物であるにも拘わらず、全酵素のうち 30-40% はその機 能が証明されていない。そこで我々は大腸菌機能未知酵素 YhhY の活性を 同定することを目指した。まず、試験管内反応の結果、YhhY はメチオニン、
ヒスチジン、フェニルアラニンをアセチル化することを明らかにした。次に、
yhhY遺伝子過剰発現株のメタボローム解析を行い、YhhY は生体内でもアミ ノ酸のアセチル化を行っていることが強く示唆された。このように、我々は大 腸菌機能未知酵素 YhhY の試験管内外での働きを明らかにした。
大腸菌機能未知酵素 YhhY による 特異的なアミノ酸のアセチル化
Amino acid acetylation by orphan enzyme YhhY in Escherichia coli
井内 仁志
慶應義塾大学大学院政策・メディア研究科後期博士課程 Hitoshi Iuchi
Doctoral Program, Graduate School of Media and Governance, Keio University
冨田 勝
慶應義塾大学環境情報学部教授 Masaru Tomita
Professor, Faculty of Environment and Information Studies, Keio University
マルタン ロベール
東北大学高度教養教育・学生支援機構准教授 Martin Robert
Associate Professor, Natural Sciences Section, Institute for Excellence in Higher Education (IEHE), Tohoku University
While the model organism Escherichia coli (E. coli) is one of the best characterized organisms, the function of only 30-40% of E.coli enzymes has been experimentally validated. Toward this objective, we screened in vitro the purified YhhY protein, (a predicted acetyl transferase) for activity on amino acids. Using Acetyl-CoA as donor, we detected acetylated products mainly with methionine, histidine and phenylalanine. Next, to confirm its activity in vivo, we performed metabolome analysis of E. coli overexpressing the yhhY gene using capillary electrophoresis time-of-flight mass spectrometry. Upon overexpression the same three acetylated-amino acids accumulated in the cell supporting the in vitro results. In this work we therefore revealed that the YhhY protein could display acetylating activity on different amino acids both in vitro and in vivo.
[研究論文]
Abstract:
大腸菌、YhhY、アミノ酸アセチル化、メタボロミクス、キャピラリー電気 泳動飛行時間型質量分析装置
escherichia coli, YhhY, amino acid acetylation, metabolomics, capillary electrophoresis-time-of-flight mass spectrometry
Keywords:
1 序論
大腸菌はこれまで最もよく研究されてきたモデル生物の一つである。しか しながら、大腸菌でさえ、全酵素のうち 30-40% はその機能が実験的に証明 されていない[1, 2, 3]。ゆえに、いくつかのアセチル化アミノ酸のような “orphan metabolite” (みなしご代謝物) 、つまり、それらを作り出す酵素がわかって いない代謝物も大量に存在する。これらの分子が代謝マップ上にギャップを 生み、代謝システムを理解する上で一つの障害になっている[4]。この問題へ の解法としてアミノ酸配列からタンパクの機能を高速に予測するアルゴリズ ムが数多く開発されてきた[3, 5]。これらのアルゴリズムを用いてタンパクの 機能を情報学的に予測することで、生化学的に重要な知見は数多く得られて きた[3]。しかしながら、代謝システムを深く理解するにはこのようなトップ ダウン的なアプローチだけではなく、一つ一つの酵素の機能を実験的に証明 するボトムアップ的なアプローチも欠かせない。なぜならば、アミノ酸配列 の相同性から得られる情報はあくまでも機能の予測であり、情報学的解析に よって機能を同定することはできないためである。例えば 1 残基のセリンか らアスパラギン酸への置換によって活性が変わることは数多く報告されてい るが、この活性の変化をアミノ酸配列の情報から見出すことはいかなるアル ゴリズムを用いても非常に難しい[6]。それにも拘わらず上記で述べたように 実験的に証明されたタンパク質はわずかであり、タンパク質の実験的な証明 は不足している。このことは現在の生物学における一つのボトルネックであ る [4]。そこで我々は大腸菌機能未知酵素の活性を同定することを目指した。
ここで、大腸菌抽出液はアセチル CoA 依存的にフェニルアラニン、ヒスチジ ン、アラニンをアセチル化することが報告されている[7]。このことは 40 年以 上前に発表されたにも拘わらず、我々が調べた範囲では未だにこれらのアミ ノ酸をアセチル化する酵素は同定されていない。そこで我々は古典的なアセ
チル化反応測定法と近年注目を集めているメタボローム解析を併用すること で、大腸菌の機能未知酵素を同定することを目指した。そのために、いくつ かの機能未知酵素の活性について質量分析機を使ってスクリーニングしたと ころ、YhhY がいくつかのアミノ酸をアセチル化することが示された (未発表 データ)。本研究ではこの YhhY のより詳細な生化学的機能を明らかにするこ とを目指した。
2 対象と手法
2.1 菌株、YhhY タンパクの発現と精製
タンパク精製用の 6 x His タグ付きyhhY遺伝子過剰発現株は ASKA ライ ブラリーから取得した[8]。培養には LB 培地(蒸留水 1 L あたり 10 g トリプ トン、5 g 酵母抽出液、10 g 塩化ナトリウム)を用いた。タンパクの発現と精 製には Overnight Express Autoinduction System (Novagen)を用いた。yhhY遺 伝子発現ベクターを持った大腸菌をクロラムフェニコール 25 μg/mL 含んだ LB 培地を用いて 37℃で一晩前培養した。2 μLの培養液を新鮮な 2 mL LB 培地に植菌し、40 μLの OnEx Solution 1 と 100 μLの OnEx Solution2 を加 えて 37℃で 16 時間振盪培養した。その後、10,000 rpm、24℃で 5 分間遠心し、
上清を捨てた。ペレットはタンパク精製まで -20℃で保存した。細胞破砕溶 液(50 mM HEPES、300 mM 塩化ナトリウム、1 mM Tween20、 10 mL あた りコンピートミニ EDTA(Roche)を 1 錠) でペレット中の細胞を破砕した。タ ンパク精製には MgneHis Ni-Particles(Promega 社)を用いた。細胞破砕液に 30 μLのビーズを加えた後、洗浄液 1 (50 mM HEPES、300 mM 塩化ナトリ ウム、10 mM Tween20、80 mM イミダゾール)と洗浄液 2 (50 mM HEPES、
300 mM 塩化ナトリウム、10 mM Tween 20、80 mM イミダゾール、80 mM ジチオトレイトール)でそれぞれ 2 回ずつ洗浄した。精製したタンパクは保存 溶液(50 mM HEPES、300 mM 塩化ナトリウム、10 mM Tween 20、50% グ リセロール)中で、-20℃で使用するまで保存した。
2.2 試験管内反応
YhhY と 20 種のアミノ酸との試験管内反応は全て 0.2 M トリス塩酸緩衝液
中で行った。保存していたビーズは反応直前に洗浄液 3 (50 mM トリシン、20 mM 塩化カリウム、5 mM 塩化マグネシウム、0.2 M 塩化マンガン、0.1 mM 硫 化鉄、0.1 mM 硫酸亜鉛 ) で 2 回洗浄した。20 mM アセチル CoA、各 10 mM のアミノ酸(アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、
グルタミン、グルタミン酸、グリシン、ヒスチジン、ロイシン、イソロイシン、
リシン、メチオニン、フェニルアラニン、プロリン、セリン、トレオニン、ト リプトファン、チロシン、バリン)と 5 μL の YhhY を精製したビーズを加え て、37℃で振盪させながら反応させた。なお、この反応は各アミノ酸毎に独立 に反応させた。反応速度定量と阻害剤添加実験は全て 0.2 M トリス塩酸緩衝 液中で行った。阻害剤添加実験時は 500 μM、1000 μM、2000 μM、2500 μM、
3000 μM ヨードアセトアミド、または 50 μM、100 μM、150 μMエチルマレイ ミドを加えて反応させた。反応開始から、15 分、30 分、45 分、1 時間反応さ せた。その後、30 μL 染色溶液 (6.4 M グアニジン塩酸塩、10 mM エルマン試 薬 ) を加えて、反応を停止させ、かつ溶液中の遊離 CoA を染色した。染色液 を加えた直後に OD440 nmをスペクトロメーターで測定した。
2.3 菌株、メタボローム解析
菌株は大腸菌 K-12 BW15113 株をコントロールとして、ASKA clone から
取得したyhhY 遺伝子過剰発現株を用いた[8]。サンプルの調製は大橋らの方
法に従った [9]。前培養は 2 mL の LB 培地、37℃、好気条件で行った。定常 期に入ったら、50 mL の LB 培地に OD600 が 0.05 になるように植菌した。
OD600 がおよそ 0.5 になった 50 mL の大腸菌培養液を 0.45 μm ポアサイズ のフィルター(ミリポア社)に通した。予め 37℃の 20 mL の蒸留水を 2 回フ ィルターに通した後、フィルターを 4℃の 2 μM 2-モルホリノエタンスルホン 酸、2 μM10- カンファースルホン酸、2 μM メチオニンスルフォンを加えた 4 mL メタノールに浸した。その後、音波処理によって細胞をフィルターから解 離させた。さらに 4 mL メタノール、1.6 mL 蒸留水を加えて、ボルテックス した。メタノール層を 5-kDa cutoff filter (ミリポア社)に入れて遠心(2,300 g で 5 分、4℃)し、高分子を除去した。メタノールは遠心濃縮機で蒸発させ た。メタボローム解析はキャピラリー電気泳動 - 飛行時間型質量分析装置 (ア
ジレント社)を用いて行った。機器の設定は平山らの方法をもとに、いくつか の点を改変した [10]。サンプルは 50 mbar で 5 秒注入した。窒素ガスの流量は 7 psig にした。
3 結果
3.1 YhhY による試験管内でのアミノ酸のアセチル化
YhhY のアセチル基転移反応の基質を同定するための実験スキームを図 1 に示した。まず、yhhY遺伝子過剰発現ベクターを持った大腸菌を培養し、
YhhY を精製した。次に、精製した YhhY と 20 種のタンパクを構成するアミ ノ酸、およびアセチル基の供給源としてアセチル CoA を試験管内に加えて反 応させた。このとき、アセチル CoA からアミノ酸にアセチル基転移反応が起 これば、副産物として遊離 CoA がつくられる。次に、チオール基と反応して
OD412nmに最大吸収波長を持つ 2-ニトロ-5-メルカプト安息香酸を生成するエ
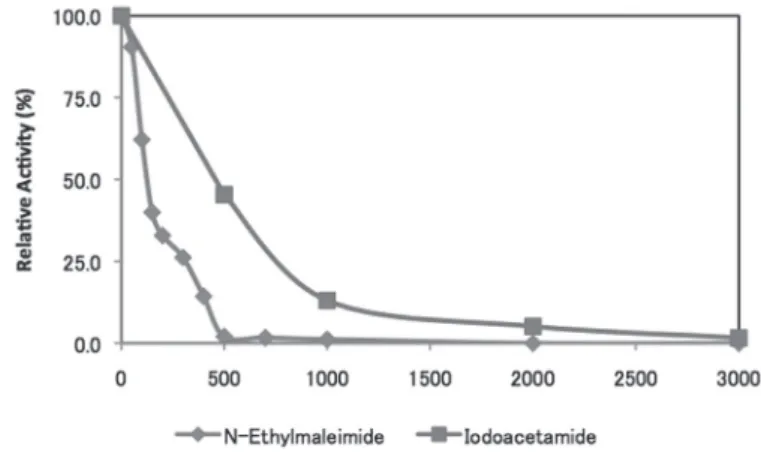
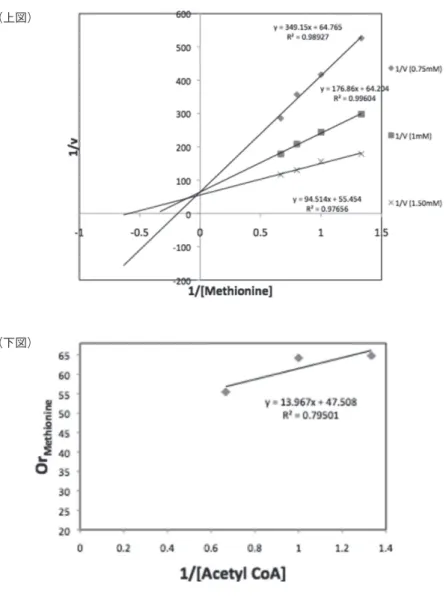
ルマン試薬を反応液に加えた。最後に反応で精製された 2-ニトロ-5-メルカプ ト安息香酸の量を測定するため、スペクトロメーターで OD440nmを測定し、遊 離 CoA を定量した。その結果、YhhY とメチオニン、ヒスチジン、フェニル アラニンを反応させた時に遊離 CoA が生成された(図 2)。かつ、生成された CoA の量は加えたアミノ酸によって異なっていた。次に、図 1 の実験系に競 合的にアセチル化を阻害するチオール反応試薬であるエチルマレイミドやヨ ードアセトアミドを加えたところ、YhhY の活性が落ちることが明らかになっ た(図 3)。さらに、反応速度を求めるために反応液の時系列サンプリングを 行い、ラインウィーバー = バークプロットとオーディネーションプロットを描 画した(図 4)。
3.2 YhhY の細胞内での活性
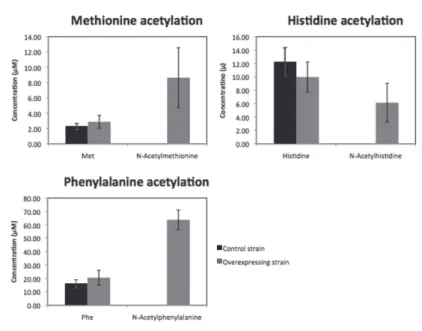
次に YhhY の細胞内での活性を明らかにするために、yhhY遺伝子過剰発現 ベクターを持った大腸菌株のメタボローム解析を行った。その結果、試験管 内反応でアセチル化された 3 種のアミノ酸はyhhY遺伝子過剰発現株からのみ 検出され、対照群からは検出されなかった (図 5)。また、メチオニン、ヒスチ ジン、フェニルアラニンについて、2 群間に有意差は無かった。
図 1 試験管内反応実験系
試験管内反応の模式図である。まず、大腸菌から YhhY を精製した。次に YhhY、ア セチル CoA とアミノ酸を試験管内で反応させた。さらに、エルマン試薬で遊離 CoA を染色し、OD412nmを測定した。
図 2 YhhY による特異的なアミノ酸のアセチル化
大腸菌から精製した YhhY、アセチル CoA と 20 種のアミノ酸を一晩反応させた ときの、アセチル基転移反応の副産物である CoA をエルマン試薬によって定量した (n=1)。縦軸は CoA の濃度 (mM)。
図 3 エチルマレイミドとヨードアセトアミドによる YhhY の阻害 大腸菌から精製した YhhY、アセチル CoA とメチオニンを試験管内で反応させてい る反応系にチオール反応試薬であるエチルマレイミド、あるいはヨードアセトアミド を加えた時の、チオール反応試薬を加えていない時に対する活性の相対値。横軸はチ オール反応試薬の濃度 (uM)、縦軸は活性相対値。菱形のプロットはエチルマレイミド、
四角形のプロットはヨードアセトアミド。
4 議論
4.1 YhhY による試験管内でのアミノ酸のアセチル化
大腸菌機能未知酵素 YhhY はアミノ酸配列の相同性からアミノ酸に対する アセチル基転移酵素であることが予測されていたが、どのアミノ酸をアセチ ル化するのかはわかっていなかった[5]。そこで我々は図 1 に示した実験系を 構築した。そして、実際に 20 種のアミノ酸に対してスクリーニングを行った ところ YhhY がメチオニン、ヒスチジン、フェニルアラニンをアセチル化す ることを明らかにした (図 2)。さらに、同じ反応時間で副産物として生成され た CoA の量が異なったことから、YhhY はそれぞれの基質に対して異なる反 応速度をとることが示唆された。次に、よく知られたチオール基に結合する 試薬であるエチルマレイミドとヨードアセトアミドによって YhhY のアセチ ル基転移反応は阻害されることが明らかになった (図 3)。さらにチオール基 に結合する試薬を反応系に加える事で YhhY の活性が落ちたことから、YhhY は反応の過程でフリーのチオール基を必要とする事が示唆された(図 3)。
図 4 YhhY に関するラインウィーバー = バークプロットとオーディネーション プロット
ラインウィーバー = バークプロット(上図)とオーディネーションプロット (下図)。
ラインウィーバー = バークプロットの横軸はメチオニン濃度の逆数 (1/mM)、縦軸は 反応速度の逆数 (sec/mmol)。菱形のプロットはアセチル CoA が 0.75 mM、四角形の プロットは 1 mM、バツ印のプロットは 1.5 mM。オーディネーションプロットの横軸 はアセチル CoA 濃度の逆数。縦軸はメチオニンのオーディネート値。
(上図)
(下図)
図 5 yhhY遺伝子過剰発現株細胞内でのアセチル化アミノ酸の蓄積 コントロール株とyhhY遺伝子過剰発現株におけるメチオニン、ヒスチジン、フェニ ルアラニンととアセチルメチオニン、アセチルヒスチジン、アセチルフェニルアラニ ンの蓄積量 (n=3)。エラーバーは標準偏差。色が濃いバーはコントロール株、色が薄 いバーは過剰発現株。
4.2 YhhY の反応速度と反応様式
YhhY のアセチル CoA とメチオニンに対する基質親和性を求めるためのラ インウィーバー = バークプロットとオーディネーションプロットを図 4 に示し た。また、それぞれの Km と Vmax を表 1 に示した。ラインウィーバー = バ ークプロットにおいて、基質濃度を変えた時にそれぞれの近似直線が平行に ならなかったことから、YhhY によるアセチル化反応は三元共有結合複合体 をとると考えられる。即ち、YhhY はアセチル CoA 及びメチオニンと結合し、
3 つの分子が複合体を形成した状態をとる。その後、CoA とアセチル化され たアミノ酸を産物として放出する。YhhY が三元共有結合複合体をとるとい うことは反応に遊離 CoA が必要であり、このことはエチルマレイミドやヨー ドアセトアミドのようなチオール反応試薬によって反応が阻害されることと
矛盾しない。この反応様式の模式図を図 6 に示した。
4.3 細胞内での YhhY の働き
ここまでで YhhY の試験管内での活性を示してきた。最後に YhhY の細胞 内での働きを明らかにするためにメタボローム解析を行った。その結果、試 験管内反応で検出されたアセチルメチオニン、アセチルヒスチジン、アセチ ルフェニルアラニンはyhhY遺伝子過剰発現ベクターを持った大腸菌からのみ 検出され、yhhY遺伝子過剰発現ベクターを持たない対照群からは検出されな かった。このことは試験管内反応の結果と一致しており、細胞内でも YhhY が 3 種のアミノ酸のアセチル化反応を担っている可能性を強く示唆している。
また、基質となるメチオニンとヒスチジン、フェニルアラニンの濃度につい て 2 群に差は無かった。これは YhhY がこれらのアミノ酸の蓄積量に与える 影響よりも、YhhY 以外の酵素が関わる反応が与える影響の方が大きかった ことが原因であると考えられる。
表 1 YhhY のメチオニンを基質とした時の反応速度 KmMet (mM) KmAcetyl CoA (mM) Vmax (uM/min)
0.0047 0.29
0.021
図 6 YhhY の反応様式
YhhY によるアセチル化反応はその過程で三元共有結合複合体をとる。第一段階と して YhhY とアセチル CoA およびメチオニンなどのアミノ酸が複合体を形成する。
第二段階としてアセチル CoA からアミノ酸へアセチル基転移反応が起きる。第三段 階としてこの反応によって生成された分子が解離する。
これまで一般的に、アミノ酸やタンパク質のアセチル化は主に真核生物で 行われているもので、原核生物においてアセチル化は非常に限定的であると されていた。しかし、近年の研究で、大腸菌やサルモネラ菌で 100 以上のア セチル化されたタンパクが見つかっている[11]。このことは原核生物において も、タンパクの翻訳後修飾が生命システムの中で何かしらの役割を担ってい る可能性を示している。本研究では YhhY がタンパクをアセチル化するかど うかは確認していないが、今後は基質にペプチドやタンパクを用いた実験を 行う予定である。
5 結論
我々は古典的な CoA アッセイ法とメタボローム解析を併用して大腸菌機能 未知酵素 YhhY の機能を同定する事を目指した。その結果、YhhY は試験管 内でメチオニン、ヒスチジン、フェニルアラニンを異なる反応速度でアセチ ル化すること、この反応はその過程で三元共有結合複合体をとることを明ら かにした。加えて、YhhY のアセチル CoA とメチオニンに対する基質親和性 を求めた。さらに、メタボローム解析の結果から、YhhY は試験管内でもア ミノ酸のアセチル化を行っていることが強く示唆された。このように我々は YhhY の試験管内と細胞内での働きを明らかにした。このことは代謝システム のより深い理解に寄与すると考えられる。
引用文献
[1] Pouliot, Y. and Karp, P.D.,“A Survey of Orphan Enzyme Activities.” BMC Bioinformatics, 8, 2007, p.244.
[2] Chen, L. and Vitkup, D.,“Distribution of Orphan Metabolic Activities.” Trends in Biotechnology, 25 (8), 2007, pp.343–348.
[3] Yamada, T., Waller, A.S., Raes, J., Zelezniak, A., Perchat, N., Perret, A., Salanoubat, M., Patil, K.R., Weissenbach, J. and Bork, P., 2012. “Prediction and Identification of Sequences Coding for Orphan Enzymes Using Genomic and Metagenomic Neighbours.” Molecular Systems Biology, 8 (581), 2012, p.581.
[4] Arita, M., “The Metabolic World of Escherichia Coli Is Not Small.” Proceedings of the National Academy of Sciences of the United States of America, 101 (6), 2004, pp.1543–1547.
[5] Lespinet, O. and Labedan, B.,“ORENZA: A Web Resource for Studying ORphan
ENZyme Activities.” BMC Bioinformatics, 7, 2006, p.436.
[6] Scott, D.B., T a Blanpied, T.A., Swanson, G.T., Zhang, C. and Ehlers, M.D., “An NMDA Receptor ER Retention Signal Regulated by Phosphorylation and Alterna- tive Splicing.” The Journal of Neuroscience : The Official Journal of the Society for Neu- roscience, 21 (9), 2001, pp.3063–3072.
[7] Krishna, R.V., Krishnaswamy, P.R. and Rao, D.R., “Enzymic Synthesis of N-Acetyl- L-Phenylalanine in Escherichia Coli K12.” The Biochemical Journal, 124(5), 1971, pp.905–913.
[8] Kitagawa, M., Ara, T., Arifuzzaman, M., Ioka-Nakamichi,T., Inamoto, E., Toyonaga, H. and Mori, H.,“Complete Set of ORF Clones of Escherichia Coli ASKA Library (a Complete Set of E. Coli K-12 ORF Archive): Unique Resources for Biological Research.” DNA Research : An International Journal for Rapid Publication of Reports on Genes and Genomes, 12(5), 2005, pp.291–299.
[9] Ohashi, Y., Hirayama, A., Ishikawa, T., Nakamura, S., Shimizu, K., Ueno, Y., Tomi- ta, M. and Soga, T., 2008. “Depiction of Metabolome Changes in Histidine-Starved Escherichia Coli by CE-TOFMS.” Molecular bioSystems, 4(2), 2008, pp.135–147.
[10] Hirayama, A., Kami, K., Sugimoto, M., Sugawara, M., Toki, N., Onozuka, H., Kinoshita,T. et al.,“Quantitative Metabolome Profiling of Colon and Stomach Can- cer Microenvironment by Capillary Electrophoresis Time-of-Flight Mass Spec- trometry.” Cancer Research, 69 (11), 2009, pp.4918-4925.
[11] Hu, L.I., Lima, B.P., Wolfe, A.J.,“Bacterial protein acetylation: the dawning of a new age.” Mol. Microbiol., 77(1), 2010, pp.15–21.
〔受付日 2015. 2. 26〕
〔採録日 2015. 7. 9〕