修士論文
バイオインフォマティックスによる
イネ科モデル植物間
miRNA オルソログの
情報基盤構築
2013 年 3 月
三重大学大学院生物資源学研究科
生物圏生命科学専攻 陸圏生物生産学講座
分子遺伝育種学教育研究分野
後藤
聖弥
目次
項 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1 材料および方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・6 1. 植物材料 2. BLAST 解析によるB. distachyonオルソログ遺伝子の探索 3. BLAST 解析によるB. distachyonオルソログmiRNA の探索 4. B. distachyonにおけるmiRNA-標的遺伝子ペアの探索 5. Total RNA の抽出 6. RT-PCR を用いた miRNA 標的遺伝子の発現解析 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・9 1. イネとB. distachyon間の遺伝子オルソログ情報の構築 2. イネmiRNA に対するB. distachyonオルソログの探索 3. BdmiRNA標的遺伝子の推定 4. RT-PCR による発現解析 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・13 1. イネ-B. distachyon間の遺伝子およびmiRNAオルソログ情報の確立 2. BdmiRNA標的遺伝子の予測と発現解析 要約・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・21 引用文献 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・22 図表・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・321
緒言
高等植物において、葯や花粉、花糸などの雄性生殖器官の形成は、他器官と比べて複 雑な分化・発達機構を有する(Scott et al. 2002)。花粉の生成は、葯室の中で胞原細胞が 分裂し、側膜細胞層と胞子形成組織に分化することで開始される。側膜細胞層は内被、 中間層、タペート組織に分化する。内被は葯裂開に関与し、中間層はデンプンを含み、 栄養分貯蔵の役割を担う。タペート組織は、形成中の花粉粒に接しており、栄養、原料 供給に重要な働きを果たす。胞子形成組織細胞は、分裂を繰り返し花粉母細胞となり、 続いて減数分裂が起こり4 個の小胞子から成る花粉四分子を形成する。花粉四分子を隔 てるカロース壁は、タペート組織から分泌されるカロース分解活性を持つカレースによ る分解を受けて小胞子が分離・成熟し花粉粒となる。その後、不等分裂が起こり花粉管 細胞と雄原細胞に分化し、2 細胞性花粉となる。さらに雄原細胞が 2 個の精細胞に分裂 し、3 細胞性花粉(成熟花粉)となり葯の裂開と共に花粉が放出される(McCormick 2004, Zhang and Wilson 2009, Zhang et al. 2011)。近年、新たな遺伝子発現機構としてエピジェネティック制御が注目されている。エピ ジェネティック制御とは、DNA の塩基配列の違いによらない遺伝子発現の制御であり、 様々な生命現象に関わることが明らかになりつつある(Zhai et al. 2008)。エピジェネテ ィック制御は、プロモーター領域のメチル化やヒストン修飾により遺伝子抑制が起こる 転写型遺伝子サイレンシング(Transcriptional gene silencing; TGS)と転写された mRNA の分解により起こる転写後遺伝子サイレンシング(Post transcriptional gene silencing; PTGS)に分類される(Goldberg et al. 2007)。TGS の例として、シロイヌナズ ナにおいて開花時期遅延を引き起こす fwa は、野生型と比較してプロモーター反復配 列のメチル化が減少しているが、これはエピジェネティックな変化が原因である
2
(Soppe et al. 2000)。さらにその後の研究により、FWAは胚乳において母方由来のアレ ルからのみ特異的に発現するインプリンティング遺伝子であることが明らかとなった (Fujimoto et al. 2008)。また、ヒストン脱アセチル化酵素であるHDA6が、DNA メチ ル化酵素 MET1 と協調しながらヒストン修飾を通してヘテロクロマチン領域の抑制や 不活性化を制御することが、シロイヌナズナの hda 変異体を用いた研究により報告さ れた(To et al. 2011)。
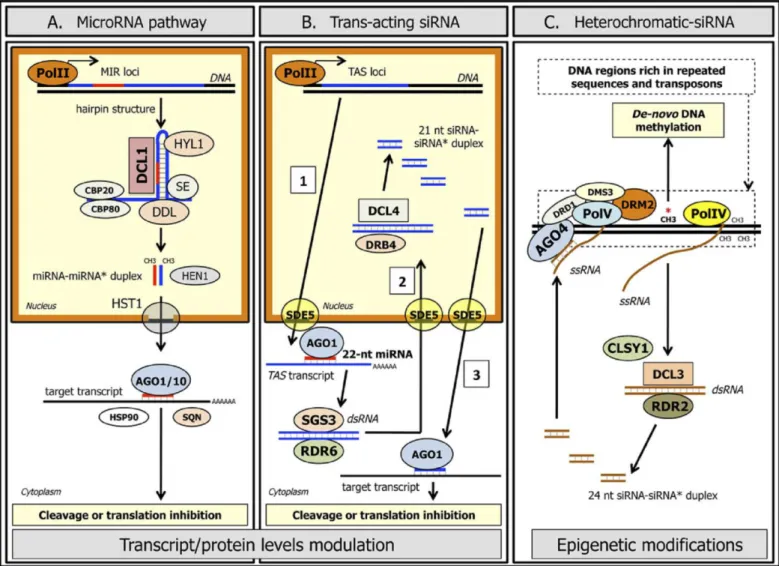
small RNA は、タンパク質をコードしない低分子 RNA(non-coding RNA)の 1 種であ るが、DNA 中のシトシンのメチル化や、ヘテロクロマチン形成に関与することが知ら れている一方で、相補な塩基配列を認識してmRNA を切断し、遺伝子の発現を抑制す ることも報告されており、TGS と PTGS の両者に関与する(Vaughn et al. 2007)。生成 経路の違いからsmall interfering RNA(siRNA)と micro RNA(miRNA)に分類される (Vaucheret et al. 2004)。siRNA に属するTrans-acting siRNA(ta-siRNA)は、TAS遺 伝子から転写された2 本鎖 RNA から生成される(図 1, Le Trionnaire et al. 2010)。ま た、ヘテロクロマチン型siRNA は、主にレトロトランスポゾン領域から生成される。 本研究で焦点を当てたmiRNA は、およそ 21-22nt 程の小さな一本鎖 RNA であり、動 植物の様々な種において配列相補的にmRNA を切断することにより、遺伝子発現を制 御している。miRNA の発現機構は、細胞核内で RNA ポリメラーゼの一種である RNA Polymerase II により miRNA 前駆体(pri-miRNA)が転写されて開始する。この pri-miRNA は、ステム・ループの二次構造を形成し、miRNA を含むステム部位から RNA 分解酵素 Dicer-like 1(DCL1)によって miRNA/miRNA*と呼ばれる二本鎖 RNA へとプロセシングされる。その後、核外輸送タンパク質であるHASTY により核から細 胞質に輸送され、miRNA/miRNA*二本鎖のうち、一方が成熟体 miRNA として RNA-induced silencing complex(RISC)に組み込まれ、もう一方(miRNA*)は分解され る。RISC に組み込まれた miRNA の配列情報を基に標的遺伝子 mRNA との結合部位
3
を認識し、エンドヌクレアーゼである Argonaute1(AGO1)タンパク質の働きにより mRNA を切断することで遺伝子発現を調節している(Bartel 2004, Vazquez et al. 2004, Park et al. 2005, Lu et al. 2005, Rajagopalan et al. 2006, Yang et al. 2006)。
植物におけるmiRNA は、シロイヌナズナの変異体を用いた研究により初めて同定さ れた。シロイヌナズナの様々な発達過程において機能する DCL1 と HEN1 の変異体 (caf-1、hen1-1)では、野生型に比べて miRNA 蓄積が減少し、花や葉の形態異常が引き 起こされたことから、DCL1とHEN1がmiRNA 生成に重要な役割を持つこと、miRNA が植物の成長過程に影響を与えることが明らかになった(Park et al. 2002, Reinhart et al. 2002)。植物の発達過程以外にも、非生物的ストレス応答に関与する miRNA も報告 されている。シロイヌナズナのmiR319c は、脱水や高塩濃度、低温、アブシジン酸処 理による非生物的ストレスにより発現量が増加することが報告されている(Reinhart et al. 2002)。またイネにおいては、低温ストレスによって発現が変動する 18 種の miRNA が同定されている。miR-1435、miR-1876、miR-1320、miR1884 はイネのみで発現し ており、種特異的な低温ストレス応答機構の存在が示唆されている(Unver and Budak 2009) 。 こ れ ま で の 研 究 に よ り 同 定 さ れ た miRNA は 、 miRBase(http://www.mirbase.org/)に情報が登録されており、シロイヌナズナやイネ、 トウモロコシ、ソルガムなど多くの種におけるmiRNA 情報が蓄積されている。
花粉は遺伝情報を次世代へと移す重要な因子であり、花粉発達過程に関与する遺伝子 も多く同定されている(Goldberg et al. 1993)。その一方で、miRNA によるエピジェネ ティック遺伝子制御機構の存在や機能に関する知見はごく限られている。この状況を打 開するために、当研究室において次世代シーケンサーによるsmall RNA の網羅的解析 が行われ、イネ雄性生殖器官発達過程におけるmiRNA による発現制御機構が明らかと なった(藤岡 2011)。雄性生殖器官発達を 4 ステージ(減数分裂/ 四分子期・小胞子期・2 細胞性花粉期・3 細胞性花粉期)に分類し、各ステージから次世代シーケンサーにより
4
small RNA を取得した。これらを分類することで、既知の miRNA やta-siRNA、ヘテ ロクロマチン型siRNA に加えて、新規 small RNA 候補の情報基盤が構築された。その 中でもmiRNA については、塩基配列を基準にした標的遺伝子の推定も行われており、 さらにタグカウントデータを基にした新規 miRNA の発現プロファイルとマイクロア レイによる花粉およびタペート細胞の標的遺伝子発現プロファイル(Hobo et al. 2008, Suwabe et al. 2008)の発現逆相関解析により、雄性生殖器官発達に関与する可能性のあ るmiRNA が選抜された。 本研究に用いたBrachypodium distachyonは、イネ科のモデルとしてヨーロッパを 中心 に注目され る植物で あ る。2001 年にイネ科のモデルとして提案された B. distachyon は、地中海沿岸や中東固有の一年草であり、コムギやオオムギなど農業的 に重要な穀物と同じイチゴツナギ亜科(Pooideae)に属する。30cm 程の植物サイズ、1-2 か月ほどのライフサイクル、2 倍体、300MB 程の単純なゲノム、生育が容易といった 特徴を持ち、2008 年にはイチゴツナギ亜科で初めて全塩基配列解読が完了した(The International Brachypodium Initiative 2010)。形質転換法についても、2001 年にパ ーティクルガン法(Draper et al. 2001)、2006 年にアグロバクテリウム法が確立されて いる(Vogel et al. 2006)。当研究室では、Alves et al.(2009)を改良したより短期間で可 能なB. distachyonの形質転換系を構築した(後藤 2010、Goto et al. 2012)。さらに、 遺伝資源コレクション、DNA マーカー、遺伝連鎖地図、BAC ライブラリー、物理地図、 変異体コレクション(http://brachypodium.pw.usda.gov, http://www.brachytag.org)、マ イ ク ロ ア レ イ 、 各 種 デ ー タ ベ ー ス (http://www.brachybase.org, http://www.phytozome.net, http://mips.helmholtz-muenchen.de/plant/index.jsp http://www.modelcrop.org)などの研究基盤整備が世界各地で進められており(The International Brachypodium Initiative 2010)、B. distachyonにおける研究環境は格 段に進歩してきている。B. distachyonは、イネやコムギ、オオムギのような主要な穀
5
類と近縁であることから(Bouchenak-Khelladi et al. 2008)、これら種で蓄積されてき た様々なデータをB. distachyonに用いることや、B. distachyonのデータをイネやム ギに適応することも可能である。実際に、コムギのさび病耐性遺伝子であるLr34/Yr18 やオオムギの光周期反応遺伝子Ppd-H1の同定にはB. distachyon由来のDNA マーカ ーが用いられた(Turner et al. 2005, Spielmeyer et al. 2008)。以上のことから、B. distachyon が次世代の単子葉モデルシステムとして有望視されているとともに、単子 葉植物の研究の幅が広がることが期待されている(Opanowicz et al. 2008)。 B. distachyonにおけるmiRNA の遺伝子発現機構はいくつか報告されているものの、 その数は限られておりまだ十分な知見は得られていない(Zhang et al. 2009)。そこで本 研究では、B. distachyonの雄性生殖器官発達機構におけるmiRNA 遺伝子発現機構の 情報基盤を構築することを目的とし、藤岡(2011)で報告されたイネ miRNA 情報を B. distachyonに移行するバイオインフォマティックス解析を行った。また、miRNA によ る発現制御が予想された標的遺伝子について、RT-PCR 発現解析による検証実験を行う とともに、イネマイクロアレイや各種データベースに蓄積されている情報と比較した。
6
材料および方法
1. 植物材料
本 研 究 に は 、International Brachypodium Initiative よ り 提 供 を 受 け た B. distachyon Bd21 を用いた。 種子は、ベンレート水和剤(住友化学)を用いて 1 日殺菌処理を行った後、バーミキュ ライトをいれたプラスティックポットに1 ポットあたり 3 粒、2 週間で 6 ポットの割合 で播種し、人工気象器(NKsystem)により 22℃・20 時間日長の条件で育成した。成熟 した種子は、乾燥させた後に4℃の休眠状態で保存した。 2. BLAST 解析によるB. distachyonオルソログ遺伝子の探索 イネ-B. distachyon間の遺伝子オルソログ情報を調査するため、イネマイクロアレイ (A rice 44K oligo microarray, Agilent Technologies)に搭載されている全遺伝子(プロー ブ)の BLAST 解析を行った。BLAST 解析は、National Center for Biotechnology Information(NCBI) よ り 提 供 さ れ て い る BLAST software(http://www.ncbi.nlm.nih.gov/Ftp/)を用い、HP XW9400 Workstation により 行った。BLAST 解析結果において、遺伝子については最も相同性が高いものをそれぞ れオルソログmiRNA および遺伝子とした。B. distachyonゲノムの塩基配列情報およ びB. distachyon遺伝子coding DNA sequence(CDS)情報は、B. distachyonデータベ ースBrachypodium.org(http://www.brachypodium.org/)から取得した。遺伝子 ID 表記 は 、 イ ネ 遺 伝 子 は The Rice Annotation Project Database(RAP-DB: http://rapdb.dna.affrc.go.jp/)の規定に、B. distachyon遺伝子はBrachypodium.org の 規定に統一した。
7
3. BLAST 解析によるB. distachyonオルソログmiRNA の探索
遺伝子と同様に、イネ-B. distachyon間のmiRNA オルソログ情報探索のため、イネ 雄性生殖器官発達への関与が推定された 191 種の新規 miRNA(藤岡 2011)の B. distachyonにおけるBLAST 解析を行った。BLAST 解析結果において、miRNA シー ケンスアライメント長が最も長く、ギャップあるいはミスマッチ数が少ないもの(最大 9b 以 下 ) を オ ル ソ ロ グ miRNA と 定 義 し た 。 オ ル ソ ロ グ miRNA は 、 miRase(http://www.mirbase.org/)に登録されている情報と比較した。
4. B. distachyonにおけるmiRNA-標的遺伝子ペアの探索
藤岡(2011)では、マイクロアレイを用いた遺伝子発現プロファイル(Hobo et al. 2008, Suwabe et al. 2008)と次世代シーケンサーを用いた small RNA の発現プロファイルを 網羅的に取得し、各発達過程のイネ雄性生殖器官における遺伝子およびsmall RNA の 発現データベースを構築している。また、それらデータベースをバイオインフォマティ ックス解析により融合し、新規miRNA 候補と予想標的遺伝子の発現逆相関解析により、 雄性生殖器官発達に関与するmiRNA を予測している。この情報を基に、B. distachyon の雄性生殖器官発達に関与が予想される miRNA とその標的遺伝子の情報基盤を構築 した。 miRNA と標的遺伝子では、一般的に 3 塩基までのギャップあるいはミスマッチが許 容されるため(藤岡 2011)、B. distachyonのmiRNA および遺伝子オルソログ情報を藤 岡(2011)のイネ miRNA-標的遺伝子ペアに対応させた後、配列アライメントを行い B. distachyon における miRNA-標的遺伝子ペアを推定した。配列アライメントには DNASIS Pro ver3.0(HITACHI)を用った。
8 5. Total RNA の抽出 サンプルには、葯前期、葯後期、雌しべ前期、雌しべ後期、葉、茎、根を用いた。発 達ステージの分類は、各ステージの花の形態(図 2)に基づき、減数分裂期・四分子期・ 小胞子期を前期、2 細胞性花粉期と 3 細胞性花粉期を後期とした。 各組織を約 0.1g ずつサンプリングし、直ちに液体窒素で凍結した。これら凍結サン プルを、Multi-Beads shocker(安井機器)を使用して 2,000rpm、10 秒間、3 回の破砕を 行った後、RNeasy Plant Mini kit(QIAGEN)を用いて各サンプルから Total RNA を抽 出した。
6. RT-PCR を用いた miRNA 標的遺伝子の発現解析
High Capacity RNA-to-cDNA™ Kit(Life Technologies)を用いて、Total RNA から cDNA を合成した。標的配列特異的プライマー(表 1)を設計し、鋳型 cDNA(50ng)に終 濃度として1×PCR Buffer、0.2mM dNTPs Mixture、0.5uM forward primer、0.5uM reverse primer、0.025U/µl TaKaRa Taq(TaKaRa)を加え、H2O で計 20µl に調整した
反応液で PCR を行った。PCR 条件は、96℃ 2 分の熱変性後、96℃ 30 秒、60℃ 30 秒、72℃ 1 分を 28 サイクル、72℃ 7 分の最終伸長とした。PCR 産物は、1%アガロ ースゲルを用いた電気泳動によって確認した。得られた遺伝子の発現パターンは、イネ マイクロアレイ(A rice 44K oligo microarray, Agilent Technologies)により遺伝子発現 プロファイルが構築されている RiceXPro(http://ricexpro.dna.affrc.go.jp/)の情報と比 較した。
9
結果
1. イネとB. distachyon間の遺伝子オルソログ情報の構築
イネ科植物のモデルであるイネとB. distachyonのゲノムには、それぞれ28,236 種、 25,532 種のタンパク質をコードする遺伝子が存在する(International Rice Genome Sequencing Project 2005, The International Brachypodium Initiative 2010)。これら 遺伝子について、2 種間の遺伝子オルソログ情報を確立するため、各種データベースと BLAST 解析により相同性検索を行った。
イネマイクロアレイに搭載されている 42,125 種の遺伝子(プローブ)について、B. distachyonに対するstand-alone BLAST 解析を行った。E-value が最も小さいものを オルソログ遺伝子として定義し、26,240 ペアのイネ-B. distachyon 遺伝子オルソログ 情報を獲得した(図 3)。また、ミシガン州立大学イネゲノムアノテーションデータベー ス(MSU)内のイネ科植物間遺伝子相同性データベース Ortholog Groups of Protein Sequences(OrthoMCL)より、13,369 ペアのイネ-B. distachyon間の遺伝子オルソログ 情報を獲得した(date not shown)。
上記2 通りの方法により獲得したイネ-B. distachyon間遺伝子オルソログ情報を統合 して解析した結果、MSU のみで得られた遺伝子オルソログ情報は 507 ペア、BLAST 解析のみで得られた遺伝子オルソログ情報は13,378 ペア、両データに共通する遺伝子 オルソログ情報は 12,862 ペアであった。MSU のみで得られた 507 ペアのうち、227 ペアはBLAST 解析においては 2 番目に高い相同性を示していたため、信頼性の高い情 報と判断してオルソログ遺伝子と定義した。これらの結果を総合すると、総計 26,467 ペアのイネ-B. distachyon 間遺伝子オルソログ情報が定義されたが、そのうち、B. distachyon遺伝子は18,535 種であり、B. distachyonで規定されている25,532 種の遺
10
伝子のうちのおよそ 72%(18,529/25,532)の遺伝子がイネとのオルソログ遺伝子である ことが予測された。
2. イネ miRNA に対するB. distachyonオルソログの探索
雄性生殖器官発達に関与する B. distachyonのmiRNA(以下 BdmiRNA)情報の基盤 を構築するために、イネmiRNA に対するB. distachyonのstand-alone BLAST 解析 を行った。 次世代シーケンサーを用いた網羅的解析により推定されたイネ雄性生殖器官発達に 関与する 191 種の新規 miRNA 情報(藤岡 2011)を基盤に、BLAST 解析による BdmiRNA の探索を行った。BLAST 解析における解析基準は、シーケンスアライメン ト長が最も長く、ギャップあるいはミスマッチ数が少ないもの(最大 9b 以下)をオルソ ログmiRNA とした。その結果、188 種の BdmiRNA を見出した(表 2)。その中で、両 種間で塩基配列が完全に一致するものは20 種(BdP20、BdP23、BdPT115、BdPT116、 BdPT32、BdPT43、BdPT48、BdPT49、BdPT5、BdPT51、BdPT59、BdPT77、BdPT85、 BdPT94、BdPT95、BdPT99、BdT18、BdT19、BdT21、BdT9)で、およそ 10%(20/191) は両種において塩基配列が完全に保存されていた。 獲得した188 種の BdmiRNA 配列を miRBase データベースの情報と比較したところ、 BdPT23 は bdi-miR529 と同一の塩基配列であった(図 4)。また、1 塩基のギャップあ るいはミスマッチを含むものは、BdPT116(bdi-miR166f)、BdPT48(bdi-miR172b)、 BdPT95(bdi-miR166a/b/c/d/e)、BdT9(bdi-miR156b/c/d)であった。 3.BdmiRNA 標的遺伝子の推定 藤岡(2011)では、網羅的解析により取得した新規 miRNA とそのターゲット遺伝子に ついて、塩基配列の相同性と発達ステージに沿った発現プロファイルを基にした逆相関
11
解析によりmiRNA の標的遺伝子を推定している。上記の BLAST 解析により取得した 遺伝子およびmiRNA のオルソログ情報を用いて、B. distachyonにおけるmiRNA と 標的遺伝子の探索を行った。イネでは191 種の miRNA に対して 628 種の標的遺伝子 が発見され、764 種の miRNA-標的遺伝子ペアが選抜されているが(藤岡 2011)、B. distachyonでは188 種の BdmiRNA に対して 357 種の標的遺伝子が見出され、470 種 のmiRNA-標的遺伝子ペアが推定された(表 3)。 miRNA と標的遺伝子の塩基配列比較 では、一般的に3 塩基までのギャップあるいはミスマッチを許容しているが(藤岡 2011)、 上記470 種のB. distachyon の miRNA-遺伝子ペアのうち、ギャップあるいはミスマ ッチ数が2 塩基のものが 5 ペア、3 塩基のものが 9 ペアであり、残りの miRNA-遺伝子 ペアは 4 塩基以上のギャップあるいはミスマッチ数であった。その内訳としては、 miRNA が 8 種、標的遺伝子が 11 種であった(表 4)。以上のことから、3 塩基までのギ ャップあるいはミスマッチ数を含むmiRNA-遺伝子ペア 13 種をB. distachyonとイネ で保存されているペアと定義した。 4. RT-PCR による発現解析 配列アライメントにより推定されたmiRNA-標的遺伝子ペアについて、それぞれの発 現に対する相関関係を調査した。BdmiRNA の発現プロファイルは、イネの結果を参照 した(表 5)。 上記で選抜された8 種の BdmiRNA のうち、BdPT32、BdPT36、BdPT44、BdT9、 BdT21 はイネにおいて減数分裂/四分子期と 2 細胞性花粉期/3 細胞性花粉期で発現パタ ーンが変動する。一方、BdPT5、BdPT48、BdPT71 は発現パターンの変動が見られな い、もしくは 2 細胞性花粉期-3 細胞性花粉期間で発現パターンが変動している。本研 究では、葯のサンプリングを前期、後期と分けたため、前者の5 種の標的遺伝子である
12
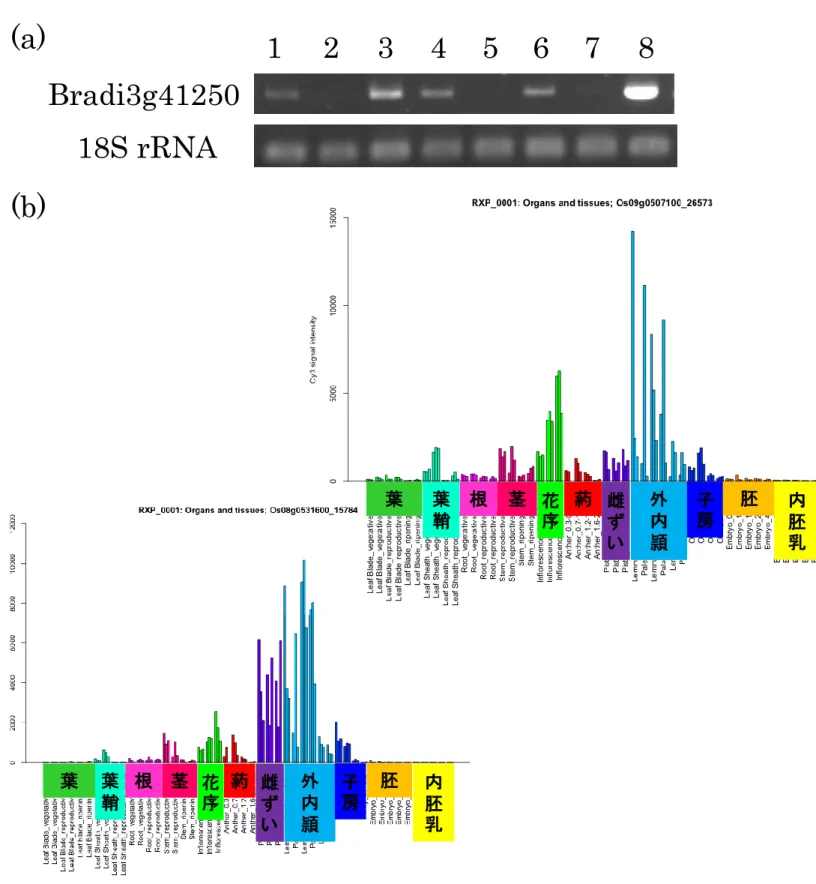
Bradi2g29330、Bradi2g16750 について、発達ステージ前後期の葯と雌しべ、栄養器 官である葉、茎、根における発現解析を行い、得られた発現プロファイルをイネマクロ アレイの発現データおよび RiceXPro に登録されている発現プロファイルと比較した (図 5)。
Bradi2g59110、Bradi3g03510、Bradi3g41250は、発達ステージ前期の葯、発達ス テージ前後期の雌しべ、茎で発現しており、3 種とも同様の発現パターンを示した(図 5A-C)。また葯においては、発達ステージの後期になると発現が減少した。これは、イ ネマイクロアレイにおける発現プロファイルと一致する。RiceXPro の発現データでは、 3 種の遺伝子とも茎での発現量が葉と根に比べて 2 倍以上高く、本解析結果とおおよそ 一致していた。 Bradi2g23350は、発達ステージ前後期の葯、茎、根で発現していた(図 5D)。発達後 期の葯では発現が減少しているが、イネの葯においては発達ステージに関係なく同程度 の 発 現量 を示 して おり本 解 析結 果と は異 なるが 、 栄養 器官 での 発現パ タ ーン は RiceXPro と一致していた。 Bradi2g29330は、発達ステージ前後期の葯特異的に発現しており、他の器官では発 現していなかった(図 5E)。発達ステージ後期の葯で発現が上昇することはイネマイク ロアレイでの発現プロファイルと一致しているが、イネでは全器官で発現が見られ、両 種間で異なる発現パターンを示した。 Bradi2g16750も葯特異的な発現パターンを示し(図 5F)、発達ステージ後期の葯で発 現が上昇していたが、イネの葯では発達ステージを通して同程度の発現量を示し、さら に雌しべや根での発現もみられ、異なる発現パターンを示した。 Bradi3g49160は、イネにおいて葯発達ステージが進むにつれて発現が上昇しており、 他器官では外頴や根、茎においても発現が見られたが、本解析では発現が見られなかっ た(図 5G)。
13
考察
1. イネ-B. distachyon間の遺伝子およびmiRNA オルソログ情報の確立
イネ科のモデルであるイネとB. distachyonは、それぞれ2004 年、2008 年に全ゲノ ム塩基配列が決定された(International Rice Genome Sequencing Project 2005, The International Brachypodium Initiative 2010)。これにより、詳細な物理地図や遺伝子 の同定、イネ科間のシンテニー関係など多くの知見が得られた。イネとB. distachyon の染色体はそれぞれ2n=24、2n=10 であるが、図のようなシンテニー関係が見られる。 また、タンパク質をコードする遺伝子はそれぞれ28,236 種、25,532 種存在するが、本 研究におけるBLAST 解析により 26,467 ペアの遺伝子がオルソログ関係であった。そ のうちイネ遺伝子は26,456 種、B. distachyonは18,529 種であり、オルソログ関係を 持つ遺伝子の割合はそれぞれで 93%(26,456/28,236)、72%(18,529/25,532)であった。 B. distachyonでの比率が低い原因として、B. distachyonはコムギ、イネからそれぞれ 3,200-3,900 万年前、4,000-5,400 万年前に分岐しているため、B. distachyonはイネよ りもコムギと近縁であることが挙げられる。イネ-B. distachyonで遺伝子オルソログと して対応付けが出来なかった遺伝子は、イネと分岐した後に独自に獲得した遺伝子であ るかもしれない。コムギは2012 年に全ゲノム塩基配列が解読されており(Brenchley et al. 2012)、三者間で比較を行うことでさらに詳細な遺伝子オルソログ情報を構築できる と思われる。 miRNA の 生 物 学 的 プ ロ セ ス は 様 々 な 生 物 種 に お い て 保 存 さ れ て お り (Jones-Rhoades et al. 2004, Guddeti et al. 2005, Kim et al. 2010, Xie et al. 2012)、植 物の miRNA は生物的/非生物的ストレス応答、情報伝達、遺伝子発現制御など多様な 機能を持つことが報告されている。シロイヌナズナで初めて同定されて以来、イネやソ
14
ル ガ ム 、 ト ウ モ ロ コ シ な ど 多 く の 植 物 種 の miRNA 情 報 が miRBase(http://www.mirbase.org/)に登録されいる(Unver et al. 2009, Nair et al. 2010, Baev et al. 2011)。2013 年現在、B. distachyonでは134 種の miRNA 情報が蓄 積されている。藤岡(2011)によりイネで推定された 191 種の新規 miRNA 配列に対する BLAST 解析では、188 種の BdmiRNA を獲得した。その中で、20 種(10%; 20/191)の miRNA はイネ-B. distachyon間で塩基配列が完全に一致していた。
BdPT23 は、miRBase に登録されている bdi-miR529 と同一の配列情報を持つ。 bdi-miR529 は、B. distachyonの実生small RNA ライブラリーから同定された(Zhang et al. 2009)。bdi-miR529 は miRNA*のタグカウント数が miRNA より 10 倍以上多い という特徴を持つ。これは、イネやEschscholzia californica、Physcomitrella patens における研究においても同様の結果が報告されている(Barakat et al. 2007)。本研究で 獲得した188 種の miRNA オルソログ情報には bdi-miR529 の miRNA*は含まれてい なかったが、miRNA*の塩基配列自身はゲノム内に存在しており、BdPT23 がとりうる ステム・ループ二次構造解析などのさらなる解析が必要である。
また、4 種の BdmiRNA が miRBase データベースとの比較により、既知 miRNA と 1 塩基のギャップあるいはミスマッチを含むことが分かった。miRNA は種を超えて保 存されており、数塩基の違いや、ゲノム上の領域の違いなどから多くのファミリーを形 成している。BdPT95 は bdi-miR166 a/b/c/d/e、BdT9 は bdi-miR156 b/c/d とそれぞれ 1 塩基の違いがあり、既知 miRNA と同一のファミリーの一部であることが示唆される。 BdPT116 と BdPT48 はそれぞれ bdi-miR166f、bdi-miR172b との 1 塩基の違いがあっ た。miR166 はシロイヌナズナやイネ、トウモロコシにおいて、クラス III ホメオドメ インーロイシンジッパー(HD-ZIP III)型転写因子をコードする遺伝子を標的とするこ とが報告されている(Reinhart et al. 2002, Jones-Rhoades et al. 2004, Juarez et al. 2004)。また、miR172 はシロイヌナズナやイネ、オオムギにおいてAPETALA 2 を標
15
的とすることが報告されている(Jones-Rhoades et al. 2004, Nair et al. 2010)。B. distachyonにおいても、bdi-miR166f はHD-ZIP III ファミリーが共通に持つSTART ドメイン、bdi-miR172b は APETALA 2 を標的とすることが報告されていており (Juarez et al. 2004)、これらの機構が種を超えて保存されていることが示されている。 そのため、BdPT116 と BdPT48 はこれら miRNA ファミリーの一部であり、他の植物 と同様の機能を有する可能性がある。 以上5 種の BdmiRNA は既知の miRNA ファミリーとの相同性が示されたが、本研 究で得られたBdmiRNA のうち、データベースに登録されていないものは 169 種ある。 これらは新規miRNA 候補となりうるため、発現解析などさらなる解析が必要である。 2. BdmiRNA 標的遺伝子の予測と発現解析
BLAST 解析により構築したB. distachyon の miRNA および遺伝子オルソログ情報 と764 種のイネ miRNA-標的遺伝子ペアとの比較解析により、470 種のB. distachyon miRNA-標的遺伝子ペアを推定した。このうち、ギャップあるいはミスマッチ数が 3 塩 基以内のものは13 ペア存在した。その内訳として、miRNA が 8 種、標的遺伝子が 11 種である。 これらB. distachyon miRNA-標的遺伝子ペアのうち、イネにおいて減数分裂/四分子 期と2 細胞性花粉期/3 細胞性花粉期(本研究の葯前期と後期の分類に相当)で発現パター ンが変動する5 種の miRNA(BdPT32、BdT9、BdT21、BdPT36、BdPT44)について、 それらの標的遺伝子(Bradi2g59110、Bradi3g03510、Bradi3g41250、Bradi2g23350、
Bradi3g49160、Bradi2g29330、Bradi2g16750)の発現解析を行った。BdPT32 は、
Bradi2g59110、Bradi3g03510、Bradi3g41250を予測標的遺伝子とし、これら標的遺 伝子は全てTranscription factor, SBP-box とアノテーション付けされている。SBP-box ファミリーには SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 遺伝子群
16
(SPL)が含まれ、花器官形成に働くAPETALA1プロモーターへの結合により雄性生殖 器官発達に関与することが明らかになっており、そのうちのいくつかはmiR156 の標的 配列を持つことがシロイヌナズナで明らかになっている(Kim et al. 2010, Xing et al. 2010)。また、miR156 を過剰発現させると葯が正常に発達せず、完全に不稔になる。
B. distachyonにおけるBradi2g59110、Bradi3g03510、Bradi3g41250の発現パター ンは、葯前期で発現し後期に減少する。この発現パターンはイネでの発現パターンと同 様である。以上のことから、イネと B. distachyon において SPL 遺伝子および miR156(BdPT32)の発現制御機構が保存されていることが推測され、花粉発達機構の共 通性を示す結果である。
BdT9 も、BdPT32 と同じく Bradi3g03510 を標的遺伝子とすることが予測された。 miRBase の情報と比較すると、BdT9 は bdi-miR156 b/c/d との相同性が高く、miR156 ファミリーに属することが予想される。bdi-miR156a/b/c は、Bradi3g03510を標的遺 伝子としていることが示されており(Vesselin et al. 2011)、本解析結果と一致している。 BdT9 も BdPT32 と同様にイネにおいて葯後期に発現が上昇しており、標的遺伝子と逆 相関の発現パターンを示すことから、SPL 遺伝子の発現制御機構に関与することが示 唆される。つまり、Bradi3g03510は複数のmiRNA による発現抑制を受けている可能 性がある。 シロイヌナズナにおいて、多くの SPL 遺伝子ファミリーの研究が行われている。 SPL10、SPL11、SPL2はアミノ酸配列の相同性が高く、重複した機能を持つことが示 唆される。SPL10の転写量を抑制すると、頂芽優勢の欠失や矮小化、花序形態の異常、 花器官の縮小化が見られる(Shikata et al. 2009)。また、SPL9とSPL10がmiR172 の 発現を活性化させることで、TARGET OF EAT(TOE)の発現を抑制し、栄養成長から生 殖成長への転換の促進を行っていることが報告されている(Wu et al. 2009)。本解析に おいて推定されたB. distachyon miRNA-標的遺伝子は 13 ペアであったが、そのうち
17
の7 ペアは Transcription factor, SBP-box をアノテーションにもつ遺伝子を標的とし ており、様々なSPL 遺伝子ファミリーがmiRNA 遺伝子制御機構に制御されているこ とが示唆される。
BdT21 は 、 Multicopper oxidase で あ る Bradi2g23350 を 標 的 と し て い る 。 Multicopper oxidase は、高等植物において多重遺伝子族を形成していることが知られ ている。生理的機能については明らかでないが、シロイヌナズナでは鉄代謝や細胞の創 傷治癒に関わることが示唆されている(McCaig et al. 2005)。また、植物における鉄は、 多くの酵素活性に必要であり、土壌から鉄を吸収している。本研究の発現解析では、茎、 根において強く発現していることから、Bradi2g23350は植物体と土壌間における鉄代 謝に関与していることが推測される。また葯において、発達前期から後期にかけて発現 量が減少しており、miRNA の発現プロファイルと逆相関を示すことから、miRNA に よる抑制が示唆される。しかし、この発現の減少はイネマイクロアレイでは見られず、 B. distachyon独自の機構である可能性も考えられる。
BdPT36 はBradi3g49160とBradi2g29330を予測標的遺伝子としている。Protein kinase, catalytic domain と ア ノ テ ー シ ョ ン 付 け さ れ る Bradi3g49160 は 、B.
distachyon において発現が見られなかった。イネにおいては、外頴や根、茎で発現が 見られたことから、外頴などの器官における発現解析を行う必要がある。 BdPT36 の 予 測 標 的 遺 伝 子 で あ る Bradi2g29330 は 、 酸 化 還 元 酵 素 で あ る Glutaredoxin とアノテーションされている。Glutaredoxin は、植物が非生物的ストレ スを受けた際に発生する活性酸素に対する防衛機構の役割を持つことが知られている。 本研究の発現解析では、発達ステージ前後期の葯特異的に発現しており、葯後期におけ る発現の上昇が見られたが、他の器官では発現していなかった。このことから、葯特異 的な活性酸素防衛機構に関与することが考えられる。イネでは全器官で発現が見られる ことから、イネとB. distachyon間では異なる役割を持つことが予想される。
18 BdPT44 は、Oxoglutarate である Bradi2g16750 を予測標的遺伝子としている。 Oxoglutarate は、細胞に対する酸素供給が不足した状態に陥った際に誘導される低酸 素誘導因子の媒体となる酵素である。Bradi2g16750 は、Bradi2g29330と同様に葯特 異的な発現パターンを示し、葯後期で発現が上昇することから、葯の発達における酸素 供給機構に関与するかもしれない。一方で、BdPT44 がイネにおいて葯後期に発現が上 昇しているため、Bradi2g16750はBdPT44 による遺伝子発現現抑制を受けていない可 能性がある。イネにおいても葯、雌しべ、根で発現が見られ、B. distachyonとの関連 性は見られなかったことから、イネとB. distachyon間ではBradi2g16750の異なる発 現機構が存在することが示唆され、B. distachyonにおけるBdPT44 の発現パターンを 詳細に調べる必要がある。 以上のように、標的遺伝子の発現解析により、B. distachyonにおける雄性生殖器官 の発達過程やその他器官のストレス応答、鉄代謝などの生理学的メカニズムに関わる miRNA-標的遺伝子ペアを推定することができた。その中でも、SPL遺伝子の発現制御 に関与する可能性のある miRNA が数多く見出されたが、SPL 遺伝子ファミリーの miRNA による発現抑制機構は種を超えて保存されている可能性が高いため、miRNA-標的遺伝子ペアのさらなる解析によりB. distachyonにおけるSPL遺伝子の知見が得 られることが期待される。 まとめ 本研究では、B. distachyonの雄性生殖器官におけるmiRNA の遺伝子発現機構の情 報基盤構築を目的として、イネの研究結果を基盤としたB. distachyonの遺伝子および miRNA オルソログ情報を構築し、それらを統合するバイオインフォマティックス解析 を行った。その結果、イネとB. distachyon間で26,467 ペアの遺伝子オルソログと 188 ペアのmiRNA オルソログを見出した。さらにその中から、miRNA と標的遺伝子の塩
19 基配列相同性の高い13 ペアの miRNA-標的遺伝子を獲得し、標的遺伝子の発現プロフ ァイルをRT-PCR により検証した。今後は、ノーザン解析や real-time PCR による発 現解析により本結果を検証し、形質転換による遺伝子機能の調査を行うことで、B. distachyon の雄性生殖器官の分化・発達における miRNA による遺伝子発現制御機構 の全容が解明されることが期待される。
20
要約
近年、植物の雄性生殖器官発達過程においてsmall RNA によるエピジェネティック 遺伝子発現機構が機能していることが明らかになりつつある。当研究室では、イネを材 料とし、次世代シーケンサーを用いた雄性生殖器官におけるmiRNA の網羅的な発現デ ータおよびマイクロアレイによる遺伝子発現プロファイルを構築している。これらデー タの機能解析には、単子葉モデル植物として注目されているBrachypodium distachyon を使用することは有用である。 本研究では、B. distachyonの雄性生殖器官におけるmiRNA の遺伝子発現機構の情 報基盤構築を目的として、イネの研究結果を基盤としたB. distachyonの遺伝子および miRNA オルソログ情報を構築し、それらを統合するバイオインフォマティックス解析 を行った。その結果、イネとB. distachyon間で26,467 ペアの遺伝子オルソログと 188 ペアのmiRNA オルソログを見出した。miRNA オルソログのいくつかは、データベー スに登録されている既知miRNA と同一のファミリーとの相同性が示唆されたが、未知 のmiRNA オルソログも多く存在した。これらは新規 miRNA 候補となりうるため、さ らなる解析が必要である。また、miRNA と標的遺伝子の塩基配列相同性の高い 13 ペ アの miRNA-標的遺伝子を獲得し、標的遺伝子の発現プロファイルを RT-PCR により 検証した。13 ペアのうち 7 ペアは雄性生殖器官発達に関与するSPL遺伝子をアノテー ションに持つ遺伝子を標的としており、様々なSPL遺伝子ファミリーがmiRNA 遺伝 子制御機構に制御されていることが示唆された。 今後は、ノーザン解析やreal-time PCR による発現解析により本結果を検証し、形質 転換による遺伝子機能の調査を行うことで、B. distachyon の雄性生殖器官の分化・発 達におけるmiRNA による遺伝子発現制御機構の全容が解明されることが期待される。21
謝辞
本研究の遂行ならびに本論文の作成にあたり、2年間に渡り終始暖かい激励とご指導、 ご鞭撻を賜りました諏訪部圭太先生に心から厚く御礼申し上げます。また、本研究をご 支援頂いた掛田克行先生、加賀谷安章先生に心より感謝申し上げます。 本研究においてバイオインフォマティックス技術から解析全般に渡って多大なるご 指導・ご協力頂いた藤岡智明博士、実験材料のサンプリングに際して貴重なアドバイス を頂いた鈴木(旧姓;増子)潤美さんに謹んで感謝申し上げます。 また、研究室の先輩として実験技術についてご指導頂いた松田智貴さん、お互いに切 磋琢磨し合った長坂香里さん、松田彩乃さん、渡辺香苗さんおよび日々の研究生活に際 して温かく支えてくださった分子遺伝育種学研究室の皆様に心より感謝申し上げます。 最後に本研究を行った2年間を含め大学生活6年の間、影ながら支援、援助を頂いた両 親および家族に感謝の意を表します。22
引用文献
Baev, V., Milev, I., Naydenov, M., Apostolova, E., Minkov, G., Minkov, I. and Yahubyan, G. (2011) Implementation of a de novo genome-wide computational approach for updating Brachypodium miRNAs. Genomics 97: 282-293.
Barakat, A., Wall, K., Leebens-Mack, J., Wang, Y.J., Carlson, J.E. and Depamphilis, C.W. (2007) Large-scale identification of microRNAs from a basal eudicot (Eschscholzia californica) and conservation in flowering plants. Plant Journal 51: 991-1003.
Bartel, D.P. (2004) MicroRNAs: genomics, biogenesis, mechanism and function. Cell 116: 281-297.
Bouchenak-Khelladi, Y., Salamin, N., Savolainen, V., Forest, F., Bank, M. Van Der, Chase, M.W. and Hodkinson, T.R. (2008) Large multi-gene phylogenetic trees of the grasses (Poaceae): progress towards complete tribal and generic level sampling. Molecular Phylogenetics and Evolution 47: 488-505.
Brenchley, R., Spannagl, M., Pfeifer, M., Barker, G.L.A., D’Amore, R., Allen, A.M., McKenzie, N., Kramer, M., Kerhornou, A., Bolser, D., Kay, S., Waite, D., Trick, M., Bancroft, I., Gu, Y., Huo, N., Luo, M.-C., Sehgal, S., Gill, B., Kianian, S., Anderson, O., Kersey, P., Dvorak, J., McCombie, W.R., Hall, A., Mayer, K.F.X.,
23
Edwards, K.J., Bevan, M.W. and Hall, N. (2012) Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 491: 705-710.
Christiansen, P., Andersen, C.H., Didion, T., Folling, M. and Nielsen, K.K. (2005) A rapid and efficient transformation protocol for the grass Brachypodium distachyon. Plant Cell Reports 23: 751-758.
Draper, J., Mur, L.A.J., Jenkins, G., Ghosh-Biswas, G.C., Bablak, P., Hasterok, R. and Routledge, A.P.M. (2001) Brachypodium distachyon A new model system for functional genomics in grasses. Plant Physiology 127: 1539-1555.
藤岡智明 (2011) 低分子 RNA を介したイネ雄性生殖器官形成・成熟におけるエピジェ ネティック制御機構の解析. 東北大学大学院生命科学研究科, 博士論文.
Fujimoto, R., Kinoshita, Y., Kawabe, A., Kinoshita, T., Takashima, K., Nordborg, M., Nasrallah, M.E., Shimizu, K.K., Kudoh, H. and Kakutani, T. (2008) Evolution and control of imprinted FWA genes in the genus Arabidopsis. PLoS Genetics 4: e1000048 1-11.
Goldberg, A.D., Allis, C.D. and Bernstein, E. (2007) Epigenetics: a landscape takes shape. Cell 128: 635-638.
Goldberg, R.B., Beals, T.P. and Sanders, P.M. (1993) Anther development: basic principles and practical applications. Plant Cell 5: 1217-1229.
24
後藤聖弥(2011) 次世代の単子葉モデル植物Brachypodium distachyonにおける形質転 換系の構築と効率化. 三重大学生物資源学部, 卒業論文.
Goto, S., Fujioka, T., Akimoto, T., Masuko-Suzuki, H., Watanabe, M., Suzuki, G., Sílvia C. Alves. and Suwabe, K. (2012) Improving the Agrobacterium-mediated transformation protocol for Brachypodium distachyon. Plant and Animal Genome XX: P0755.
Guddeti, S., Zhang, D.C., Li, A.L., Leseberg, C.H., Kang, H., Li, X.G., Zhai, W.X., Johns, M.A. and Mao, L. (2005) Molecular evolution of the rice miR395 gene family. Cell Research 15: 631-638.
Hobo, T., Suwabe, K., Aya, K., Suzuki, G., Yano, K., Ishimizu, T., Fujita, M., Kikuchi, S., Hamada, K., Miyano, M., Fujioka, T., Kaneko, F., Kazama, T., Mizuta, Y., Takahashi, H., Shiono, K., Nakazono, M., Tsutsumi, N., Nagamura, Y., Kurata, N., Watanabe, M. and Matsuoka, M. (2008) Various spatiotemporal expression profiles of anther-expressed genes in rice. Plant Cell Physiology 49: 1417-1428.
International Brachypodium Initiative (2010) Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 463: 763-768.
International Rice Genome Sequencing Project (2005) The map-based sequence of the rice genome. Nature 436: 793-800.
25
Jones-Rhoades, M.W. and Bartel, D.P. (2004) Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Molecular Cell 14: 787-799.
Juarez, M.T., Kui, J.S., Thomas, J., Heller, B.A. and Timmermans, M.C.P. (2004) MicroRNA-mediated repression of rolled leaf1 specifies maize leaf polarity. Nature 428: 84-88.
Kim, J.Y., Lee, H.J., Jung, H.J., Maruyama, K., Suzuki, N. and Kang, H. (2010) Overexpression of microRNA395c or 395e affects differently the seed germination of Arabidopsis thaliana under stress conditions. Planta 232: 1447-1454.
Le Trionnaire, G., Grant-Downton, R.T., Kourmpetli, S., Dickinson, H.G. and Twell, D. (2011) Small RNA activity and function in angiosperm gametophytes. Journal of Experimental Botany 62: 1601-1610.
Lu, C., Tej, S.S., Luo, S., Haudenschild, C.D., Meyers, B.C. and Green, P.J. (2005) Elucidation of the small RNA component of the transcriptome. Science 309: 1567-1569.
McCaig, B.C., Meagher, R.B. and Dean, J.F.D. (2005). Gene structure and molecular analysis of the laccase-like multicopper oxidase (LMCO) gene family in
26
McCormick, S. (2004) Control of male gametophyte development. Plant Cell 16: S142-S153.
Nair, S.K., Wang, N., Turuspekov, Y., Pourkheirandish, M., Sinsuwongwat, S., Chen, G., Sameri, M., Tagiri, A., Honda, I., Watanabe, Y., Kanamori, H., Wicker, T., Stein, N., Nagamura, Y., Matsumoto, T. and Komatsuda, T. (2010) Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage. Proceedings of the National Academy of Sciences, USA 107: 490-495.
Opanowicz, M., Vain, P., Draper, J., Parker, D. and Doonan, J.H. (2008)
Brachypodium distachyon: making hay with a wild grass. Trends in Plant Science 13: 172-177.
Park, M.Y., Wu, G., Gonzalez-Sulser, A., Vaucheret, H. and Poethig, R.S. (2005) Nuclear processing and export of microRNAs in Arabidopsis. Proceedings of the National Academy of Sciences, USA 102: 3691-3696.
Park, W., Li, J., Song, R., Messing, J. and Chen, X. (2002) CARPEL FACTORY, a dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in
Arabidopsis thaliana. Current Biology 12: 1484-1495.
27
evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes & Development 20: 3407-3425.
Reinhart, B.J., Weinstein, E.G., Rhoades, M.W., Bartel, B. and Bartel, D.P. (2002) MicroRNAs in plants. Genes and Development 16: 1616-1626.
Scott, R.J., Spielman, M. and Dickinson, H.G. (2004) Stamen structure and function. Plant Cell 16: 46-61.
Shikata, M., Koyama, T., Mitsuda, N. and Ohme-Takagi, M. (2009) Arabidopsis
SBP-box genes SPL10, SPL11 and SPL2 control morphological change in association with shoot maturation in the reproductive phase. Plant Cell Physiology 50: 2133-2145.
Sílvia C. Alves., Worland, B., Thole, V., Snape, J.W., Bevan, M.W. and Vain, P. (2009). A protocol for Agrobacterium-mediated transformation of Brachypodium distachyon community standard line Bd21. Nature Protocols 4: 638-649.
Soppe, W.J.J., Jacobsen, S.E., Alonso-Blanco, C., Jackson, J.P., Kakutani, T., Koornneef, M. and Peeters, A.J.P. (2000) The late flowering phenotype of fwa
mutants is caused by gain-of-function epigenetic alleles of a homeodomain gene. Molecular Cell 6: 791-802.
28
X., Appels, R. and Lagudah, E.S. (2008) Fine scale genetic and physical mapping using interstitial deletion mutants of Lr34/Yr18: a disease resistance locus effective against multiple pathogens in wheat. Theoretical and Applied Genetics 116: 481-490.
Suwabe, K., Suzuki, G., Takahashi, H., Shiono, K., Endo, M., Yano, K., Fujita, M., Masuko, H., Saito, H., Fujioka, T., Kaneko, F., Kazama, T., Mizuta, Y., Kawagishi-Kobayashi, M., Tsutsumi, N., Kurata, N., Nakazono, M. and Watanabe, M. (2008). Separated transcriptomes of male gametophyte and tapetum in rice: validity of a laser microdissection (LM) microarray. Plant Cell Physiology 49: 1407-1416.
To, T.K., Kim, J.-M., Matsui, A., Kurihara, Y., Morosawa, T., Ishida, J., Tanaka, M., Endo, T., Kakutani, T., Toyoda, T., Kimura, H., Yokoyama, S., Shinozaki, K. and Seki, M. (2011) Arabidopsis HDA6 regulates locus-directed heterochromatin silencing in cooperation with MET1. PLoS Genetics 7: e1002055 1-14.
Turner, A., Beales, J., Faure, S., Dunford, R.P. and Laurie, D.A. (2005) The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 310: 1031-1034.
Unver, T. and Budak, H. (2009) Conserved microRNAs and their targets in model grass species Brachypodium distachyon. Planta 230: 659-669.
29
Vain, P., Worland, B., Thole, V., McKenzie, N., Alves, S.C., Opanowicz, M., Fish, L.J., Bevan, M.W. and Snape, J.W. (2008) Agrobacterium-mediated transformation of the temperate grass Brachypodium distachyon (genotype Bd21) for T-DNA insertional mutagenesis. Plant Biotechnology Journal 6: 236-245.
Vaucheret, H., Vazquez, F., Crété, P. and Bartel, D.P. (2004) The action of
ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes and Development 18: 1187-1197.
Vaughn, M.W., Tanurdžić, M., Lippman, Z., Jiang, H., Carrasquillo, R., Rabinowicz, P.D., Dedhia, N., McCombie, W.R., Agier, N., Bulski, A., Colot, V., Doerge, R.W. and Martienssen, R.A. (2007) Epigenetic natural variation in Arabidopsis thaliana. PLoS Biology 5: e174 1617-1629.
Vazquez, F., Vaucheret, H., Rajagopalan, R., Lepers, C., Gasciolli, V., Mallory, A.C., Hilbert, J.L., Bartel, D.P. and Crété, P. (2004) Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Molecular Cell 16: 69-79.
Vogel, J. and Hill, T. (2008) High-efficiency Agrobacterium-mediated transformation of Brachypodium distachyon inbred line Bd21-3. Plant Cell Reports 27: 471-478.
30
Agrobacterium-mediated transformation and inbred line development in the model grass Brachypodium distachyon. Plant Cell, Tissue and Organ Culture 84: 199-211.
Wu, G., Park, M.Y., Conway, S.R., Wang, J-W., Weigel, D. and Scott, R.P. (2009) The sequential action of miR156 and miR172 regulates developmental timing in
Arabidopsis. Cell 138: 750-759.
Xie, K., Shen, J., Hou, X., Yao, J., Li, X., Xiao, J. and Xiong, L. (2012) Gradual increase of miR156 regulates temporal expression changes of numerous genes during leaf development in rice. Plant Physiology 158: 1382-1394.
Xing, S., Salinas, M., Höhmann, S., Berndtgen, R. and Huijser, P. (2010) miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell 22: 3935-3950.
Yang, J.H., Han, S.J., Yoon, E.K. and Lee, W.S. (2006) Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells. Nucleic Acids Research 34: 1892-1899.
Zhai, J., Liu, J., Liu, B., Li, P., Meyers, B.C., Chen, X. and Cao, X. (2008) Small RNA-directed epigenetic natural variation in Arabidopsis thaliana. PLoS Genetics 4: e1000056 1-11.
31
Zhang, J., Xu, Y., Huan, Q. and Chong, K. (2009) Deep sequencing of Brachypodium
small RNAs at the global genome level identifies microRNAs involved in cold stress response. BioMed Central Genomics 10: 449.
Zhang, D., Luo, X. and Zhu, L. (2011) Cytological analysis and genetic control of rice anther development. Journal of Genetics and Genomics 38: 379-390.
Zhang, D. and Wilson, Z.A. (2009) Stamen specification and anther development in rice. Chinese Science Bulletin 54: 2342-2353.
32
図表
図 2 雄性生殖器官発達ステージ
(a)葯前期 (b)葯後期 (c)雌しべ前期 (d)雌しべ後期
(a)
(b)
(c)
(d)
3細胞性花粉
2細胞性花粉
減数分裂期
・四分子期
小胞子期
葯の写真は東北大学院生命科学研究科植物生殖遺伝分野
鈴木(旧姓;増子)潤美氏より提供(unpublish data)。
図 3 イネ-B. distachyon
間遺伝子オルソログ情報
13,378
12,862
227
イネマイクロアレイのプローブをクエリーとしたB.
distachyon
に対するBLAST解析と、MSUデー
タベースにより獲得したイネ-B. distachyon
間遺
伝子オルソログ情報の相互比較。
MSU
BLAST解析結果
図 4 BdmiRNAとmiRBaseデータベースとの比較
BdPT23
bdi-miR529
10
20
30
40
50
60
AGAAGAGAGA GAGUACAGCC U
... ... ... ...
AGAAGAGAGA GAGUACAGCC U
... ... ... ...
BdPT48
bdi-172b
10
20
30
40
50
60
G
GGAAUCUUG AUGAUGCUGC AU
... ... ... ...
-
GGAAUCUUG AUGAUGCUGC AU
... ... ... ...
BdT9
bdi-156b
bdi-156c
bdi-156d
10
20
30
40
50
60
G
UGACAGAAG AGAGUGAGCA C
... ... ... ...
-
UGACAGAAG AGAGUGAGCA C
... ... ... ...
-
UGACAGAAG AGAGUGAGCA C
... ... ... ...
-
UGACAGAAG AGAGUGAGCA C
... ... ... ...
BdPT95
bdi-166a
bdi-166b
bdi-166c
bdi-166d
bdi-166e
10
20
30
40
50
60
U
UCGGACCAG GCUUCAUUCC C
-... ... ... ...
-
UCGGACCAG GCUUCAUUCC C
C... ... ... ...
-
UCGGACCAG GCUUCAUUCC C
C... ... ... ...
-
UCGGACCAG GCUUCAUUCC C
C... ... ... ...
-
UCGGACCAG GCUUCAUUCC C
C... ... ... ...
C
UCGGACCAG GCUUCAUUCC C
-... ... ... ...
BLAST解析により獲得したBdmiRNAと
miRBaseデータベースとの比較。
赤線はミスマッチを示す。
BdPT116
bdi-miR166f
10
20
30
40
50
60
U
U
UCGGACCA GGCUUCAUUC C
... ... ... ...
U
C
UCGGACCA GGCUUCAUUC C
... ... ... ...
Bradi2g59110
1 2 3 4 5 6 7 8
18S rRNA
RiceXProより引用したOs01g0922600
の発現プロファイル
Bradi2g59110
のRT-PCR
PCRは28サイクルで行った。各レーンのサンプルは、1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
図 5A BdPT32の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
Bradi3g03510
RiceXProより引用したOs06g0703500
の発現プロファイル
Bradi3g03510
のRT-PCR
PCRは28サイクルで行った。各レーンのサンプルは、1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
図 5B BdPT32の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
図 5C BdPT32の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
Bradi3g41250
RiceXProより引用したOs08g0531600
・
Os09g0507100
の発現プロ
ファイル
Bradi3g41250
のRT-PCR
PCRは28サイクルで行った。各レーンのサンプルは1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
図 5D BdT21の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
Bradi2g23350
RiceXProより引用したOs05g0458600
の発現プロファイル
Bradi2g23350
のRT-PCR
PCRは28サイクルで行った。各レーンのサンプルは1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
図 5E BdPT36の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
Bradi2g29330
RiceXProより引用したOs05g0353600
の発現プロファイル
Bradi2g29330
のRT-PCRの結果
PCRは28サイクルで行った。各レーンのサンプルは1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
図 5F BdPT44の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
Bradi2g16750
RiceXProより引用したOs05g0560900
の発現プロファイル
Bradi2g16750
のRT-PCRの結果
PCRは28サイクルで行った。各レーンのサンプルは、1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
1 2 3 4 5 6 7 8
18S rRNA
図 5G BdPT36の標的遺伝子とイネオルソログ遺伝子の発現プロファイル
Bradi3g49160
RiceXProより引用したOs02g0623600
の発現プロファイル
Bradi3g49160
のRT-PCRの結果
PCRは28サイクルで行った。各レーンのサンプルは、1. 葯前期 2. 葯後期
3. 雌しべ前期 4. 雌しべ後期 5. 葉 6. 茎 7. 根 8. ゲノムを示す。
(a)
(b)
(a)
(b)
葉
葉
根 茎
鞘
花
序
葯 雌
ず
い
外
内
頴
子
房
胚
内
胚
乳
primer
sequence(5'→3')
Bradi2g59110.1_F
TTCCAGCAGCAGCCATTTAC
Bradi2g59110.1_R
AGTTCCCCTGATCACCATTCAC
Bradi3g03510.1_F
AAAGCCAACAACTGCTGCAG
Bradi3g03510.1_R
ATGCATCGCAGGGTTTGAAG
Bradi3g41250.1_F
AATCCAAGACCGGAGCAAAG
Bradi3g41250.1_R
TTGAGCTGCTCGTTTTCCAC
Bradi2g23350.1_F
ACTACACTGCAAAGCTACGC
Bradi2g23350.1_R
GGGTAGAATGGGAAGTTGGAC
Bradi3g49160.1_F
AACTCGCAGAACAACTGGTG
Bradi3g49160.1_R
TGATATCATGGCAGCCGTTG
Bradi2g29330.1_F
TCGGCCAAGGACCATAATGG
Bradi2g29330.1_R
CGGTCTTGTCGCTGAACCTG
Bradi2g16750.1_F
AGATGTGCGCTGAGAAGGAT
Bradi2g16750.1_R
GCCGTCGGATTAGTATCTGC
表 1 発現解析に用いたプライマー
イネmiRNA イネmiRNA配列 BdmiRNA BdmiRNA配列 ミスマッチ P1 AAATAAGACGAACAATCAAATCTG BdP1 AAATAAGCCGAACAATCAAGTTAC 5 P10 TAGTTTAGATGTGGTTCCTTC BdP10 GCAGGAACCACATCTAAAATT 3 P11 TCGGAAAGGGTCAAATTGTCA BdP11 GTGGAAAGGGTCAAATTGGGA 4 P12 TCGGACGAGCAGCAGCCGCGA BdP12 CCGGACGAGCAGCCGCCGCGG 3 P13 TCGGCCTCGTGGATGGACCAGGA BdP13 TTCCGGTCGATCCACGAGGCCGC 4 P14 TCGGCCTCGTGGATGGACCAGGAG BdP14 CTTCCGGTCGATCCACGAGGCCGC 4 P15 TCTGGAGTTTTGACGAATTA BdP15 ACAGGAGTTTTGACGAATTC 3 P16 TGATCTCTTCAAGATAGTTGGC BdP16 TGATCTCTTCAAGATAGTTGGC 4 P17 TGGAAACATGTAGAAGATGA BdP17 GGGAATCATGTAGAAGATGA 2 P18 TGGAGAAGAGAATGGACACGG BdP18 TCGTGGCCATTCTCTTCTCCA 2 P19 TGGAGAAGAGAATGGACACGGA BdP19 GTCGTGGCCATTCTCTTCTCCA 3 P2 ATATACGGACACACATTTAAAGT BdP2 AAATTAAGTTATGTGTGTCCGTATAT 5 P20 TGGTGGTGACGGGTGACGGA BdP20 TGGTGGTGACGGGTGACGGA 0 P21 TGTGCAGTGGCTATACTCTCAA BdP21 GTGAGAGTATAGCCACTGCGCT 3 P22 TTATACAAGTAGAAACAAAAAT BdP22 GTATACATGTAGAAACAAAAAT 2 P23 TTCAATAAAGCTGTGGGAAA BdP23 TTCAATAAAGCTGTGGGAAA 0 P24 TTCGGAAAGGGTCAAATTGTCA BdP24 AGTGGAAAGGGTCAAATTGGGA 5 P25 TTGAGGAGCTCGAAGTAGGCAC BdP25 TTGAGGAGCTCGATGTAGCAGC 4 P26 TTGGAGAGAAGCTTGCATCTT BdP26 GATTTGCAAGCTTCTCTCCAT 4 P27 TTTCAAATTGTTTGACACCGT BdP27 CTTCAAATTGTTTGACATACGA 4 P28 TTTCACTTTGGACTAGGTATT BdP28 TTTCACTTTGGATTAGGTCAG 4 P29 TTTGAATCGCAGGGTAGAAAAA BdP29 CGATTTTACCCTGCGATTCAAG 5 P3 CGAGGCCTGCACGTTCACGT BdP3 ACGTGAACGTGCAGGCGGCG 2 P30 TTTTTTCGTTACGAGATGTAGA BdP30 TCCAACCATCTCGTAACGAAATGA 5 P31 TTTTTTGGAAGGCAGAGTATG BdP31 AGTTTTGGAAGGCCGAGTATG 3 P32 TTTTTTTCGTTACGAGATGTAGAA BdP32 TGCAAGAACTTACGAGATGTAGAT 9 P4 GAAGTCCTCGTGTTGCATCCC BdP4 GAAGTCCTCGTGTTGCATCCT 1 P5 GCTGTAGCACGGACTCCAAGACGC BdP5 GCTGTAGCACGTACTCCATGCTGT 5 P6 GGATGTGGGCAATGCTAGAATGAC BdP6 GAATGTGGGCAATGCTACAGAATGTT 5 P7 GGGGCTTGCATATTCTACCA BdP7 TGGTAGAATATGCAAAGGGT 5 P8 GTTGATTCTTTTTGGGACGGAGGG BdP8 CCCTCCGTCCCAAAATGAATAAAA 3 P9 TAACAAAGGACAACAGACTGA BdP9 TAACAAAGGACAACAGGAAGG 4 PT1 AAACATGTAAGAAAAAATCAACGG BdPT1 AAACATTTAAGAAAAAATTCAATGG 3 PT10 ACGTAAATTTTTTGAATAAGACGA BdPT10 TCGTCTTATTCAAAAAATTATGCG 3 PT100 TTGAACCTTGCATCGGCGAACT BdPT100 ATCGGAGGGATGCAAGGTTCAA 7 PT101 TTGAGGAGCTCGAAGTAGGCA BdPT101 TTGAGGAGCTCGATGTAGCAG 4 PT102 TTGATCTTTGCAGGCGGACCA BdPT102 TGGTTCGCCTGCAAAGATGGC 4 PT103 TTGATGACGACGTATATCGG BdPT103 TTGATGACGACGTAGTCCTC 5 PT104 TTGCAATAGATAATATGTGTT BdPT104 GACACATATTATGTATTGCAG 3 PT105 TTGCTCCGTTGGTTGGTGATC BdPT105 TTGCTCCGTTGGTTGGTGTTA 2 PT106 TTGCTTATCGTTCCGCTCAC BdPT106 TTGAGCTGAACGATAAGCAT 3 PT107 TTGGATTGATAGATAGATAGA BdPT107 TCTATCTATCTATCTATCCAA 1 PT108 TTGGCAGTAGAGGGCTGCAA BdPT108 TTGGCAGTAGAAGGCTGCAA 1 PT109 TTGGGTTATTTGAGGGCTAAG BdPT109 TTTTTTCCTCAAATAACCCAA 4 PT11 ACGTCTTATAACCTGAAACGGAGG BdPT11 CCTCTGTTTCAGATTATAAGACGT 2 PT110 TTGTAGCCAGCTGTAGCACGGA BdPT110 TTGTAGCCAGCTCTAGCAGAGG 4 PT112 TTTAACGAAGAGTTGCAGTGT BdPT112 AATCTGCAACTCTTCGCTAAA 3 PT113 TTTACATTGTGAAACGGATGAA BdPT113 TTCATCCGTTTCAAAATGTAAT 2 PT114 TTTAGTCCCGGTTGGTAACAT BdPT114 TTTAGTCCCGGTTGCTCGATCGT 5 PT115 TTTCCGATGCCTCCCATTCCTA BdPT115 TTTCCGATGCCTCCCATTCCTA 0 PT116 TTTCGGACCAGGCTTCATTCC BdPT116 TTTCGGACCAGGCTTCATTCC 0 PT117 TTTCGTGAGGCGTTAAAAACT BdPT117 TGGATGGAGGCGTTAAAAACT 5 PT118 TTTCTCTGCAATATCGTCCCT BdPT118 TTTCTCTGCAATATCACAGAT 5 PT119 TTTCTTGGAGAGATCAATGAG BdPT119 TTTCTTGGAGAGAACAATAGT 4 PT12 ACTAAAAAGTCAACGGTGTCAAAC BdPT12 GTTTTCACCGTTGACTTTTTAAT 3 PT120 TTTGATGAAATGTAGAAATGA BdPT120 TCCTTTCTACATTTGATCAAA 2 PT121 TTTGTAGCCAGCTGTAGCACGG BdPT121 CTTGTAGCCAGCTCTAGCAGAG 4 PT122 TTTGTAGCCAGCTGTAGCACGGA BdPT122 CTTGTAGCCAGCTCTAGCAGAGG 5 PT123 TTTGTAGCCAGCTGTAGCACGGAC BdPT123 CTTGTAGCCAGCTCTAGCAGAGGC 5 PT124 TTTGTTTGATTGCTGGAGCTA BdPT124 CAGCTCCAGCAAACAAACAAA 2 PT125 TTTTCACTTTGGACTAGGTAT BdPT125 TTTTCACTTTGGATTAGGTCA 3 PT126 TTTTCTCACATGTTTGACCAT BdPT126 TTTTCTCACATGTTTGAATGA 4