耐酸性通性嫌気性細菌の代謝工学的アプローチ
によるコハク酸生産能改良に向けた研究
Study of metabolic engineering approach to improve
succinate productivity in aciduric facultative anaerobe
田島 義教
Yoshinori TAJIMA
目次 1章 緒論 1-1. 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1-4[page] 1-2.コハク酸発酵法に関するこれまでの知見・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・4-6 1-3.コハク酸生産能を向上させる為の代謝工学的アプローチに関するこれまでの知見・・・・・・・7-9 1-4.従来のコハク酸発酵法の課題と解決に向けた施策・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・10-11 1-5.酸性嫌気条件下におけるコハク酸発酵法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・12 1-6.Enterobacter aerogenes に関するこれまでの知見・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・13 1-7.研究の意義と目的・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・14-16 2 章 耐酸性通性嫌気性細菌 E. aerogenes のコハク酸生産菌としての能力評価 2-1.緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・17-18 2-2.実験材料と方法 2-2-1.菌株・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・19 2-2-2.培地と培養条件・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・19 2-2-3.発酵槽を用いた酸性好気条件下と酸性嫌気条件下での培養試験・・・・・・・・・・・・・・・・・19 2-2-4.E. coli と E. aerogenes の乾燥菌体重量(Dried Cell Weight; DCW)算出式・・・・・・・・・・・・19 2-2-5.グルコース濃度、OD、有機酸、アルコールの定量・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20 2-3 結果
2-3-1.E. aerogenes の酸性(pH5.0)好気条件下における培養特性・・・・・・・・・・・・・・・・・・・・・・・24 2-3-2.E. aerogenes の酸性(pH5.0)嫌気条件下における培養特性・・・・・・・・・・・・・・・・・・・25-26 2-3-3.E. aerogenes AJ116037 株の酸性(pH5.0)嫌気条件下における代謝産物の同定・・・・27-28 2-4 考察
2-4-2. E. aerogenes の細胞形態がグルコース消費速度に及ぼす影響・・・・・・・・・・・・・・・・・29-30 2-4-3.E. aerogenes AJ116037 株の酸性(pH5.0)嫌気条件下における代謝反応・・・・・・・・・・30-31 2-5.結言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・32 3 章 E. aerogenes の主要な NADH 酸化経路の遮断と異種炭酸固定反応の導入によるコハク酸生 産能の改良 3-1.緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・33-36 3-2.実験材料と方法 3-2-1.菌株とプラスミド・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 3-2-2.培地と培養条件・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 3-2-3.E. aerogenes からのゲノム DNA 抽出法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37-38 3-2-4.マイクロチューブを用いたコハク酸発酵・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・38 3-2-5.本章で使用したプライマー・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・38 3-2-6.E. aerogenes の形質転換法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・38-39 3-2-7.λ-red recombination system による adhE 遺伝子破壊株の構築・・・・・・・・・・・・・・・・・・・39-40 3-2-8.λ-red recombination system による budA 遺伝子破壊株の構築・・・・・・・・・・・・・・・・・・・・・40 3-2-9.A. succinogenes PCK 発現用プラスミド pSTV28-pck の構築・・・・・・・・・・・・・・・・・・・・・40-41 3-2-10.C. glutamicum PYC 発現用プラスミド pSTV28-pyc の構築・・・・・・・・・・・・・・・・・・・・・41-42 3-2-11.可溶性画分タンパク質の抽出方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 3-2-12.SDS-PAGE による PYC と PCK の発現解析・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 3-2-13.各種酵素活性(LDH、ACDH、AR)の測定・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・43 3-2-14.E. aerogenesの DCW 算出式・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・43 3-2-15.グルコース濃度、OD、有機酸、アルコールの定量・・・・・・・・・・・・・・・・・・・・・・・・・・・・・43 3-3.結果
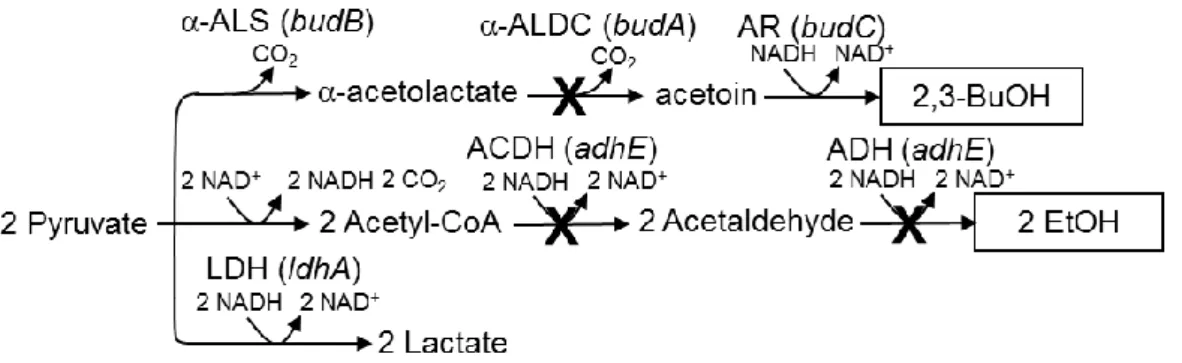
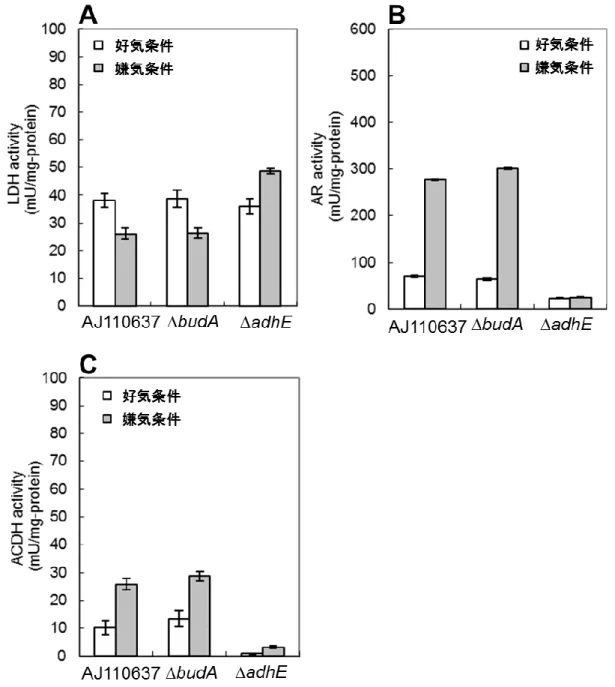
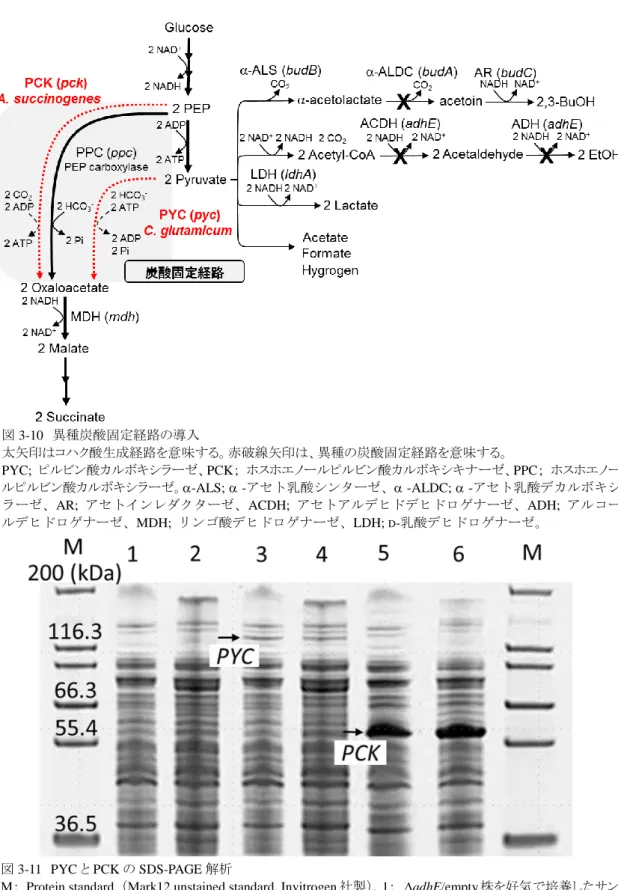
3-3-1.エタノール、及び 2,3-ブタンジオール生成経路の遮断がグルコース資化能に及ぼす影 響・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・49-50 3-3-2.budA、及びadhE 株における LDH、AR、ACDH 活性測定・・・・・・・・・・・・・・・・・・・51-53 3-3-3.エタノール、及び 2,3-ブタンジオール生成経路の遮断によるコハク酸生産能の改良・54-55 3-3-4.C. glutamicum 由来 PYC と A. succinogenes 由来 PCK の発現解析・・・・・・・・・・・・・・・56-57 3-3-5.C. glutamicum 由来 PYC と A. succinogenes 由来 PCK の導入によるコハク酸生産能の改 良・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・58-59 3-3-6.C. glutamicum 由来 PYC と A. succinogenes 由来 PCK の導入とエタノール生成経路の遮 断の組み合わせによるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・60-61 3-4.考察
3-4-1.エタノールと 2,3‐ブタンジオール生成経路の遮断によるコハク酸生産能の改良・・・・・・・62 3-4-2.adhE 株における AR の総活性値・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・62 3-4-3.異種炭酸固定経路の導入によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・63 3-4-4. C. glutamicum 由来 PYC と A. succinogenes 由来 PCK の導入とエタノール生成経路の遮断 の組み合わせによるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・64 3-4-5. adhE/PYC 株とadhE/PCK 株のコハク酸収率の差異・・・・・・・・・・・・・・・・・・・・・・・・・64-65 3-5.結言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・66 4 章 E. aerogenes のピルビン酸由来副生経路の遮断と、PYC と PCK の共発現によるコハク酸生 産能の改良 4-1.緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・67-69 4-2.実験材料と方法 4-2-1.菌株とプラスミド・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70 4-2-2.培地と培養条件・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70
4-2-3.マイクロチューブを用いたコハク酸発酵・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70 4-2-4.発酵槽を用いた pH 制御下でのコハク酸発酵・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70 4-2-5.E. aerogenes からのゲノム DNA 抽出法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70-71 4-2-6.本章で使用したプライマー・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・71 4-2-7.E. aerogenes の形質転換法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・71 4-2-8.λ-red recombination system による遺伝子破壊株の構築・・・・・・・・・・・・・・・・・・・・・・・・・・71 4-2-9.薬剤耐性遺伝子除去用プラスミド pRSF-Para-IX の構築・・・・・・・・・・・・・・・・・・・・・・・・71-72 4-2-10.pRSF-Para-IX を用いた薬剤耐性遺伝子の除去・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・72 4-2-11.A. succinogenes PCK と C. glutamicum PYC 共発現プラスミド用 pSTV28-pck+pyc の構 築・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・72 4-2-12.E. aerogenes の DCW 算出式・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・72-73 4-2-13.グルコース濃度、OD、有機酸、アルコールの定量・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・73 4-3.結果 4-3-1.乳酸生成経路の遮断によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・78-79 4-3-2.酢酸生成経路の遮断によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・80-81 4-3-3.2,3-ブタンジオール生成経路の遮断によるコハク酸生産能の改良・・・・・・・・・・・・・・・82-84 4-3-4.PCK と PYC の共発現によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・85-86 4-3-5.培養 pH がコハク酸生産能に及ぼす影響・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・87-89 4-4.考察 4-4-1.ピルビン酸由来副生経路の遮断によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・90 4-4-2.PYC と PCK の共発現によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・90-91 4-4-3.培養 pH の低下がコハク酸生産能に及ぼす影響・・・・・・・・・・・・・・・・・・・・・・・・・・・・・91-93 4-5.結言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・94
5 章 E. aerogenes のコハク酸生成経路における ATP 獲得量の向上によるコハク酸生産能の改良 5-1.緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・95-96 5-2.実験材料と方法 5-2-1.菌株とプラスミド・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97 5-2-2.培地と培養条件・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97 5-2-3.マイクロチューブを用いたコハク酸発酵・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97 5-2-4.発酵槽を用いた pH 制御下でのコハク酸発酵・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97 5-2-5.E. aerogenes からのゲノム DNA 抽出法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97 5-2-6.本章で使用したプライマー・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・97-98 5-2-7.E. aerogenes の形質転換法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・98 5-2-8.λ-red recombination system による遺伝子破壊株の構築・・・・・・・・・・・・・・・・・・・・・・・・・・98 5-2-9.pRSF-Para-IX を用いた薬剤耐性遺伝子の除去・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・98 5-2-10.A. succinogene PCK の染色体固定・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・98 5-2-11.frdABCD オペロンのプロモーター置換株の構築・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・98 5-2-12.SDS-PAGE 解析・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・98-99 5-2-13.ES08ptsG 株と ES08ptsI 株の M9 グルコース培地での生育速度・・・・・・・・・・・・・・・・99 5-2-14.E. aerogenes の DCW 算出式・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・99 5-2-15.グルコース濃度、OD、有機酸の各種定量・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・99 5-3.結果
5-3-1.コハク酸生成経路における ATP 収支を向上させる為の代謝工学的アプローチ・・・104-107 5-3-2.ピルビン酸オキシダーゼの不活化によるコハク酸生産能の改良・・・・・・・・・・・・・・・108-109 5-3-3.A. succinogenes PCK 発現カセットの染色体固定と SDS-PAGE による発現解析・・・・・・110 5-3-4.A. succinogenes PCK の発現量がコハク酸生産能に及ぼす影響・・・・・・・・・・・・・・・・・・111 5-3-5.フマル酸レダクターゼの発現強化によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・112
5-3-6.ピルビン酸ギ酸リアーゼの不活化によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・113 5-3-7.グルコース PTS の遮断がグルコース資化能に及ぼす影響・・・・・・・・・・・・・・・・・・・114-115 5-3-8.グルコース PTS の遮断によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・・116 5-3-9.ES08ptsG 株における弱酸性(pH5.7)嫌気条件下のコハク酸発酵・・・・・・・・・・・・117-119 5-4.考察 5-4-1.理論 ATP 収支の向上によるコハク酸生産能の改良・・・・・・・・・・・・・・・・・・・・・・・・・・・・・120 5-4-2.E. aerogenes の嫌気条件下でのグルコース取り込み機構・・・・・・・・・・・・・・・・・・・・120-121 5-4-3.ES08ptsG 株を用いた弱酸性(pH5.7)嫌気条件下のコハク酸発酵・・・・・・・・・・・・・・・・121 5-4-4.コハク酸発酵の展望・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・121-122 5-5.結言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・122 6 章 結論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・123-125 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・126-133 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・134
1
1 章 緒論
1-1 緒言 現在、日本におけるプラスチックの生産量は、年間 1,400 万トンに至る。これらプラスチックの殆 どは石油を原料に製造され、使用後に埋め立て廃棄される製品循環を辿る。このように、日本を初 めとする先進国の殆どが石油資源の消費に基づく経済活動「石油消費社会」を営んでいる。一方、 プラスチックを取り巻く環境は、石油価格の上昇や製造時の環境汚染に対する懸念、更には、プラ スチックを消費した後に廃棄する埋立て処理場の残余容量の急減が指摘されており、社会的な問 題となっている。 この問題に対する有望な解決策の一つとして、バイオマスから完全生分解性プラスチックを生産 する方法が考えられる[1,2]。糖やスターチを含む植物バイオマスは、光合成の働きにより再生利用 が可能である。また、得られた完全生分解プラスチックは、土壌中で微生物の働きにより、二酸化 炭素と水に完全分解され、再び植物に利用される。このように、資源を循環利用することが持続可 能な社会の実現に肝要である。現在、最も実用化が進んでいる完全生分解プラスチックとして、ポリ乳酸(Polylactic acid; PLA) とポリブチレンサクシネート(Polybutylene succinate; PBS)が挙げられる(表1-1)。PLAは、引張強 度に優れるものの、伸び率は低い熱可塑性高分子である[1,2]。その為、硬材として自動車部品、 繊維、ボトルなどに利用されている。近年では、PLAの融点が170 ℃程度で扱い易い理由から、 3Dプリンターの樹脂などに用途が拡大されている。これに対して、PBSは伸び率が高く、包装用資 材やコンポストバッグなど、PLAとは異なる用途で利用される[1,2]。2010年、バンクーバーで開催さ れた冬期オリンピックでは、選手村の食堂で使用するスプーンやフォークに三菱化学社製のPBS (GS Pla®)が採用され、廃棄物処理量の低減化に成功した。このように物性が異なるPLAとPBSは、 異なる用途展開が期待され、2010年代には、生分解性プラスチックのプラスチック製品市場に占め る割合は、約10%にまで増加すると予測されている。
2
このようにPLAとPBSは、代表的な完全生分解性プラスチックである一方、両者の原料製法は異 なる。現在、PLAの原料であるL-乳酸は、再生可能な植物バイオマスからも発酵法で製造が可能 である。2002年から、Nature Works LLC(米)は、PLA(IngeoTM)の商業生産を開始し、年間14万ト
ンのPLAを製造販売している。このように、植物バイオマスを原料に製造されたPLAは、完全生分 解性プラスチックの先駆者として、資源循環型の製品サイクルを実現したと言える。 それに対して、PBSの原料であるコハク酸は、石油資源から化学合成法でのみ製造される。コハ ク酸は、ナフサに含まれるn-ブタンやベンゼン等を原料として、気層酸化により得られた無水マレイ ン酸から化学合成される。現在、年間20~30万トンの無水マレイン酸がコハク酸に変換され、PBS や基幹化学物質(γ-ブチロラクトン、テトラハイドロフラン、1,4-ブタンジオール)の原料として利用さ れている[3,4]。2014年、コハク酸から合成される製品群の世界市場は年間約1兆5000億円にのぼ り、PBSの普及に牽引される形で、その市場規模は益々拡大すると見込まれる[5]。 このようにコハク酸は、PBS を含む化学基幹物質として、その用途は多岐に渡り、市場規模も極め て大きいと言える[5]。コハク酸を発酵法で製造する技術開発は、完全生分解性プラスチックの普 及に大きく貢献するだけでなく、基幹化学物質の一つをバイオマスから製造する点で脱石化社会 に む け た 重 要 な 取 り 組 み の 一 つ と 言 え る 。 こ の よ う な 背 景 か ら 、ア メ リ カ の エ ネ ル ギ ー 省
(Department of Energy; DOE)は、2004 年にコハク酸を再生可能な植物バイオマスから製造すべき 12 基 幹 化 学 物 質 の 一 つ に 認 定し た [4] 。 2000 年 以 降 、Myriant 、 BioAmber/ 三 井 化 学 、 BASF/Corbion-Purac, DSM/Roquette などの化学メーカーが主体となり、コハク酸発酵の研究開発 が精力的に行われているが、依然として本格的な商業生産は行われていない[5]。 このような社会的な要請に対して、本研究では従来のコハク酸発酵法よりも経済的で且つ低環 表 1-1 PLA と PBS における構造式、原料、原料製法、物性及び用途 石油化学合成法 発酵法
3 境負荷型な発酵法として、新たに酸性嫌気条件下におけるコハク酸発酵法を提案するに至った。 この発酵を実現するには、従来のコハク酸生産菌開発の延長線上ではなく、新たに耐酸性通性嫌 気性細菌をプラットフォーム菌に選定し、代謝工学的アプローチにより、該細菌のコハク酸生産能 を改良させることが必要である。 本章では、本研究に着手するに至った社会的な背景と動機を整理する。次に、コハク酸発酵法 に関する基礎的な知見を纏めると共に、これまで検討されてきたコハク酸生産菌を整理することで、 現行のコハク酸発酵法の課題を抽出する。また、この課題について化学的な論拠に基づいた解決 法を模索し、新たなアプローチとして酸性嫌気条件下におけるコハク酸発酵法に関して、その具現 性を評価した。次に、この発酵法を行う上で、プラットフォーム菌に求める形質について整理をおこ うと共に、これまでに実施されたコハク酸生産能を向上する為の代謝工学的アプローチを纏めた。 最後に、Enterobacter aerogenes の知見を整理する過程で、コハク酸生産のプラットフォーム菌とし ての可能性を検討した。以上を纏めたうえで、本研究の課題を整理すると共に、本研究の目的と意 義を明らかにした。
4 1-2.コハク酸発酵法に関するこれまでの知見 コハク酸は、TCAサイクルの中間代謝物であり、好気と嫌気の両環境下で発酵生産が可能であ る。通常、腸内細菌の殆どは、好気条件下でコハク酸を細胞外に排出しない。一方、コハク酸から フマル酸への反応を触媒するコハク酸デヒドロゲナーゼ(Succinate dehydrogenase;SDH)を欠失し た菌株では、グルコースを資化時に、コハク酸を主要な生成物として培地中に蓄積する(図1-1) [7,8]。この時、つまり、コハク酸生成経路として酸化的にTCAサイクルを利用すると、二酸化炭素を 生成する反応を経由する為、グルコースからの重量収率は68%となる。それに対して、嫌気条件下 では、多くの菌種がコハク酸を細胞外に排出する。実際、E. coliの野生株では、グルコースを消費 し、重量収率で10%程度のコハク酸を培地中に蓄積した[9]。嫌気条件下でコハク酸が生成される 機構は、酵素反応的に説明が可能である。オキサロ酢酸からスクシニルCoAまでの全ての反応が 可逆反応であるのに対して、スクシニルCoAからα-ケトグルタル酸の反応が不可逆である(図1-1)。 その為、細胞内のスクシニルCoAとコハク酸濃度が上昇し、排出担体を有するコハク酸のみが選択 的に細胞外へ排出される[10,11]。この時、つまり、還元的にTCAサイクルを利用すると、二酸化炭 素の固定反応を経由する為、グルコースからの重量収率も112%となり、好気条件と比較して1.65 倍高い値となる。重量収率の観点から両培養条件を比較すると、嫌気条件下の方がコハク酸発酵 に好ましい環境と言える。実際、コハク酸生産菌の殆どが嫌気条件下でコハク酸を生産するように 開発が進められている [12-14]。 1990年以降、様々な細菌種を用いて、コハク酸生産菌の開発が行われてきた。表1-2に、これま でに報告された代表的なコハク酸生産菌の属種と発酵成績を纏めた。 表1-2に示すように、コハク酸生産菌は、2つのグループに大別でき、一つは、Actinobacillus succinogenes, Anaerobiospirillum succiniciproducens, Mannheimia succiniciproducensなどの元来
コハク酸を生産するルーメン細菌群とEscherichia coliやCorynebacterium glutamicumなどの従来ア ミノ酸発酵などの生産菌として利用されてきた腸内細菌群である[12-16]。前者は、牛などの反芻動 物のルーメンからコハク酸を生産する細菌として単離されてきた。ルーメン内では、牧草サイレージ
5 が糖化された後に、これら細菌群の働きによりコハク酸などの有機酸に変換される。代表的なルー メン細菌である、Actinobacillus succinogenesでは、グルコース重量の50%以上をコハク酸に変換で きる [12,17]。コハク酸発酵開発の初期では、これらルーメン細菌を利用した、効率的なコハク酸発 酵プロセスの検討が多数報告された[18,19]。一方、遺伝子組換え技術が確立されていないルーメ ン細菌群では、更にコハク酸生産能を改善する取り組みは困難であり、次第に研究目的が遺伝子 組換え技術の確立に移行した [20-22]。
一方、遺伝子組換え技術が既に確立されているE. coliやC. glutamicumでは、コハク酸生産能を 向上させる遺伝形質が次々に報告され、ルーメン細菌群を上回る発酵成績が報告されている [15,16]。特に、E. coli KJ134株は、酢酸、エタノール、乳酸、ギ酸の生成経路に関わる10個の遺 伝子を破壊した多重欠損株で、グルコースからコハク酸を71.6 g/L蓄積した。この時の重量収率は、 100%であり、理論収率(112%)に漸近した値である[16]。 このように、代謝工学的アプローチを用いて、コハク酸生産能を改良することは可能である。その 為、新たなプラットフォーム菌を選定する際に、該細菌の遺伝子組換え技術の成熟度は、重要な 選択基準の一つに成り得る。
6 菌株名 炭素源 通気条件 培養 pH 蓄積 (g/L) 収率 % 引用文献 Actinobacillus succinogene グルコース 嫌気 7.0 105.8 82 [12] Anaerobiospirillum succiniciproducens グルコース 嫌気 6.4 83.0 88 [13] Manheimia succiniciproducens グルコース 嫌気 6.5 52.4 76 [14] Corynebacterium glutamicum グルコース 嫌気 6.7 146.0 90 [15] Escherichia coli グルコース 嫌気 6.7 71.6 100 [16] 図 1-1 好気と嫌気条件下におけるコハク酸生成経路の比較 左図、好気条件下では、二酸化炭素の生成を伴いながらコハク酸が生成される。右図、嫌気条件下では、二酸化 炭素を固定しながらコハク酸が生成される。SDH; コハク酸デヒドロゲナーゼ。 表 1-2 各種コハク酸生産菌の培養条件(炭素源、通気条件、培養 pH)と発酵成績(蓄積、収率)の比較
7 1-3.コハク酸生産能を向上させる為の代謝工学的アプローチに関するこれまでの知見 E. coliやC. glutamicumでは、コハク酸生産能が向上した遺伝子改変株が多数報告されている [15,16]。これらの報告を整理すると、嫌気条件下でコハク酸生産能を向上させる代謝工学的アプ ローチは、2つの基本的戦略に基づいている。それは、①コハク酸生成経路と競合するNADH酸化 経路の遮断と②コハク酸生成経路の律速反応である炭酸固定経路の強化である。まず、①につい て詳細な説明を行う。一般的に、好気条件下では、呼吸鎖の働きによりNADHが再酸化され、生命 活動に必要なATPが生産される(図1-2)。一方、酸素が存在しない嫌気条件下では、乳酸デヒドロ ゲナーゼ(Lactate dehydrogenase: LDH)やエタノールデヒドロゲナーゼ(Alcohol dehydrogenase: ADH)、及びリンゴ酸デヒドロゲナーゼ(Malate dehydrogenase: MDH)が呼吸鎖の役割を果たし、 NADHの再酸化を行う(図1-2)。つまり、嫌気条件下では、乳酸やエタノール、更にはコハク酸を生 産する過程でNADHの酸化を行い、細胞内の酸化還元バランスの維持を行っている(図1-2)。 コハク酸生産能を向上させるには、NADHをコハク酸生成経路にあるMDHで、いかに効率良く再 酸化が出来るかが重要である。その為には、競合するNADH酸化経路を遮断する必要がある。例 えば、E. coliでは、消費したグルコース重量の約50%が乳酸に変換される[9]。コハク酸を主要な生 成物にする為には、LDHの反応経路を遮断する方法が定石と言える。 次に、②コハク酸生成経路の律速反応である炭酸固定経路の強化について説明する。E. coliで は、LDHの反応経路を遮断したのみでは、コハク酸が主要な生成物として蓄積されない。なぜなら、 炭 酸 固 定 反 応 で あ る ホ ス ホ エ ノ ー ル ピ ル ビ ン 酸 カ ル ボ キ シ ラ ー ゼ ( Phosphoenolpyruvate carboxylase: PPC)がコハク酸生成時の律速反応となり、オキサロ酢酸の供給量を制御している為 である(図1-3)。この代謝制御は、PPCがリンゴ酸などのC4カルボン酸によりアロステリックなフィー ドバック阻害を受ける事に起因している[23,24]。 この課題を解決する方法として、これまでに外来生物種からC4カルボン酸の阻害が解除された PPCを導入する試みが報告されている。例えば、Sorghum vulgare由来のフィードバック阻害解除型 PPCをE. coliに発現させた結果、グルコースからのコハク酸蓄積が3倍増加した[51]。また、PPCとは、
8
異 な る 炭 酸 固 定 経 路 を 利 用 す る 方 法 も 提 案 さ れ て い る 。 Corynebacterium glutamicum や Lactococcus lactisなどの細菌種では、ピルビン酸カルボキシラーゼ(Pyruvate carboxylase: PYC)
がピルビン酸からオキサロ酢酸の反応を触媒する(図1-3)。Sánchez等は、E. coli のADHとLDHを 不活化した株にLactococcus lactis由来PYCを導入する方法で、18.7 (g/L)のグルコースから15.6 (g/L)のコハク酸を生産する菌株の構築に成功した[46]。これは、重量収率で83.4%のコハク酸を 生産した事を意味し、①と②の戦略を組み合わせた良例である。 このように、①の戦略を実行するには、その細菌の嫌気条件下における主要なNADH酸化経路 を同定し、その反応を触媒する酵素を特定する必要がある。また、②の戦略を実行するには、既存 の炭酸固定反応を触媒する酵素の性質を見極める必要がある。ゲノム配列情報が開示された細 菌を用いることで、これらの反応を推定することが可能となり、確度の高い代謝工学的アプローチ が実行できる。
9 図 1-3 E. coli とコハク酸生産菌における炭酸固定反応 左図、E. coli では PPC を介して、炭酸固定反応が行われる。PPC は、リンゴ酸などの生成物によるアロステリックなフィ ードバック阻害をうける。右図、コハク酸生産菌では、フィードバック阻害が解除された異種由来の PPC*や PYC を炭 酸固定反応として用いる。PPC; ホスホエノールピルビン酸カルボキシラーゼ、PYC;ピルビン酸カルボキシラーゼ。 図 1-2 好気と嫌気条件下における酸 化還元バランス 左図、好気条件下では、呼吸鎖により NADH が酸化される。右図、嫌気条件 下では、乳酸、エタノール、コハク酸生 成経路で NADH が酸化される。NDH; NADH デヒドロゲナーゼ、CYA/CYD; シトクロムデヒドロゲナーゼ、PPC; ホ スホエノールピルビン酸カルボキシラ ーゼ、PYC; ピルビン酸カルボキシラ ーゼ、LDH; 乳酸デヒドロゲナーゼ、 ADH; アルコールデヒドロゲナーゼ、 MDH; リンゴ酸デヒドロゲナーゼ。
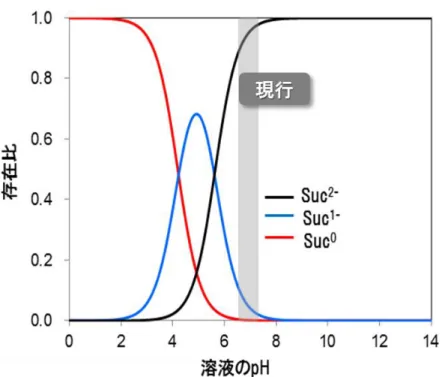
10 1-4.従来のコハク酸発酵法の課題と解決に向けた施策 様々な細菌種から、高力価なコハク酸生産菌が報告される一方で、これらの菌株の培養方法に は共通の特徴がある。表1-2が示すように、全ての菌種が中性嫌気条件下で、コハク酸発酵を行っ ている。コハク酸発酵では、コハク酸が酸性物質である為、培養pHを維持する目的でアルカリ(中 和剤)を培地に添加する必要がある。アルカリの添加は、培地の浸透圧を上げ、コハク酸生産能を 低下させる。その為、多くの研究では1価のアルカリよりも2価のアルカリであるMg(OH)2やCa(OH)2 が用いられる。取り分け、安価な中和剤としてCa(OH)2が多用される[25,26]。 図1-4に示したコハク酸の解離曲線が示すように、中性付近では殆ど全てのコハク酸が、2つの プロトンが解離した2価のアニオン(Suc2-)として存在する為、1 molのコハク酸を中和するには、理 論上、1 molのCa(OH)2が必要となる。このように、中性条件下でのコハク酸発酵では、多量のアル カリが消費される計算になる。 一方、コハク酸とアルカリが共存する発酵ブロスからコハク酸を単離する工程では、硫酸などの 強酸を添加し、培地pHを2付近まで下げ、溶解度の低いフリー体(Suc0 ;電荷的に中性)のコハク酸 結晶を取り上げる方法が知られている[27,28]。仮に、Ca(OH)2を中和剤として選択した場合、培地 中のカルシウムイオンは、硫酸と難溶性の硫酸カルシウム(石膏)を形成する。硫酸カルシウムは、 水に対する飽和溶解度が約3 g/Lと極めて低く、pHを下げていく過程で殆どのカルシウムイオンが 石膏という形で結晶として取り上げられる。その後、コハク酸イオンを含む発酵ブロスを最終的に pH2.0付近まで下げる方法でフリー体のコハク酸結晶のみを取り上げることが可能である。 この一連のプロセスでは、最終的に、コハク酸と石膏が生産される(図1-5)。発酵工程で得られ る石膏は、着色などの問題から産業価値が低く、その殆どが産業廃棄物として処理されることが想 定される。仮に、pH7.0でコハク酸発酵を行うと、コハク酸重量の1.15倍の石膏を廃棄する計算にな る。この一例が示すように、中性付近のコハク酸発酵製造プロセスでは多量のアルカリと酸を使用 し、結果として、著量の副生塩を製造するプロセスが想定される。
11 図 1-4 各 pH におけるコハク酸のイオン化状態
コハク酸の pKa を pKa1 4.16 と pKa2 5.6 として解離曲線を作成した。
グレイの領域は、これまでに報告されている細菌を用いたコハク酸発酵の pH 領域(pH6.4~7.0)を意味する。
図 1-5 コハク酸発酵製造法の概要
発酵法では、培養 pH を維持する目的でアルカリを添加し、コハク酸を単離する工程では酸を使用する。その 結果、多量の副生塩が生産される。
12 1-5.酸性嫌気条件下におけるコハク酸発酵法 コハク酸の解離曲線が示すように、使用する副原料(アルカリと酸)と生産される副生塩の重量は、 培養pHに依存的と言える。仮に、培養pHを7.0から5.0まで下げると、コハク酸の殆どが1価の酸とし て存在する(図1-4)。この時、培養pHを5.0に維持する為に必要なアルカリ量は、pH7.0の場合と比 較して半分となる。投入したアルカリ量が半減化出来れば、副生塩もまた原理的に半減の重量とな る。その為、酸性条件下でのコハク酸発酵は、副原料の削減による製造コストの減尐のみならず、 副生塩の生成を抑制した無駄の尐ない環境低負荷型の生産プロセスと言える。一方、これまでに 報告されたコハク酸生産菌の殆どが、pH6.0以下の環境ではコハク酸生産能が顕著に低下、もしく は消失することが知られている[25,26]。その為、酸性嫌気条件下でのコハク酸発酵を実現するに は、同環境化で効率的にコハク酸を生産できる菌株の開発が肝要であり、新たな挑戦と言える。ル ーメン細菌群の例のように、酸性嫌気条件下でコハク酸を生産する菌種を使用する選択肢もある が、そのような菌種の報告例はこれまでにない。そこで、遺伝子組換え技術が可能な菌種の中から、 耐酸性が高い菌種を選択し、代謝工学的アプローチでコハク酸生産能を向上させる戦略が現実 的である。実際、耐酸性能が高い乳酸桿菌Lactobacillus plantarumからコハク酸生産菌を開発す る取り組みも報告されているが、コハク酸生産能は低く、更なる改善が必要である[29]。
13 1-6. Enterobacter aerogenes に関するこれまでの知見
乳酸菌以外にも、耐酸性が高い細菌として Pantoea spp.、Klebsiella spp.、Enterobacter spp.等が 知られている。中でも、Enterobacter aerogenes は、γ-プロテオバクテリアに属する通性嫌気性細菌 で、生育可能な pH 領域が E. coli よりも幅広く、pH4.0 の酸性環境下でも生育が可能である。また、 酸性嫌気条件下でグリセロールやグルコースなどの炭素源を資化し、2,3-ブタンジオール、エタノ ール、水素などを生産する形質を有する[30-33]。その為、バイオ燃料の生産菌として、これまで幅 広く研究が進められてきた。一方、嫌気的な環境下で多様な代謝産物を生産するにもかかわらず、 その代謝反応を触媒する酵素、制御機構などの知見は乏しい。実際、E. aerogenes を用いたバイ オ燃料生産菌の開発では、変異株(非遺伝子組換え体)の解析が主流であり、代謝工学的なアプ ローチは、殆ど実施されていない。2012 年に E. aerogenes の全ゲノム配列が開示されると、遺伝子 欠損技術やオミックス解析など、遺伝学的な解析手法が徐々に報告されるようになったが、依然と して、E. coli コハク酸生産菌のような高度な遺伝子組換え体の報告例はない[34]。
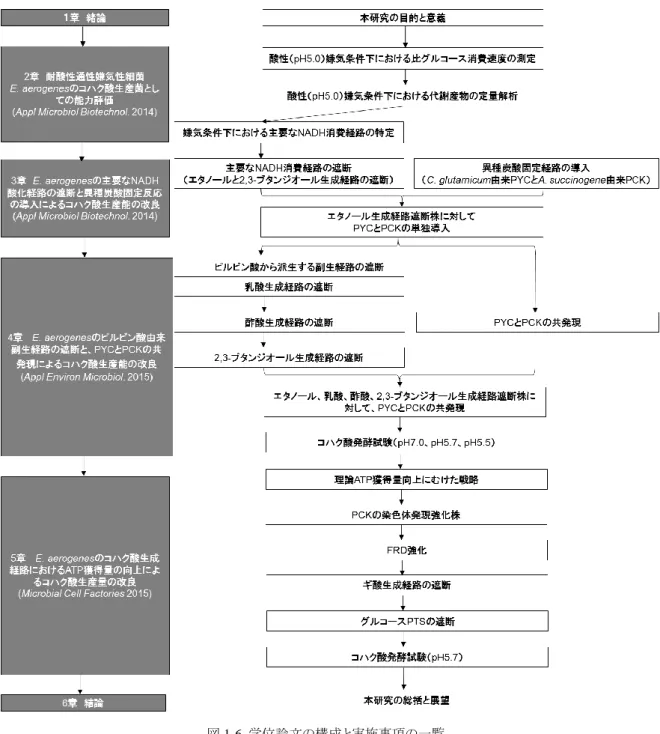
14 1-7.研究の意義と目的 PBS の原料であるコハク酸を、植物バイオマスを用いて発酵法で生産する技術開発は、脱石化 社会の取り組みの一つであり、資源循環型社会の実現に繋がる社会的な要請と言える。一方、現 状の細菌から構築されたコハク酸生産菌の殆どが、中性条件下でしかコハク酸生産能を示さない。 その為、現状のコハク酸発酵は、中性付近で行うプロセスが想定され、多量の副原料を使用し、副 生塩を廃棄する課題が顕在する。この課題に対して、本章ではコハク酸発酵における培養 pH を下 げる方法、つまり酸性嫌気条件下でコハク酸発酵を行うことで、副原料の使用量と副生塩の生成 量を削減できることを示した。一方、これまでに報告されたコハク酸生産菌の殆どが、pH6.0 以下の 環境ではコハク酸生産能が顕著に低下、もしくは消失することが知られており、現状のコハク酸生 産菌の改良では、酸性(pH<6.0)嫌気条件下でのコハク酸発酵は困難と言える。そこで、本研究 では酸性(pH<6.0)嫌気条件下でのコハク酸発酵を実現する為に、新たに耐酸性通性嫌気性細 菌である E. aerogenes をプラットフォーム菌に選定し、そのコハク酸生産能を向上させる為の代謝 工学的アプローチを明らかにすることを目的とした。 以下に本研究の構成ならびに目的を纏めた(図1-6)。 1章 「緒論」では、コハク酸発酵におけるこれまでの知見、及び課題を明らかにし、本研究の目的 と意義を示した。 2章 「耐酸性通性嫌気性細菌E. aerogenesのコハク酸生産菌としての能力評価」では、酸性嫌気 条件下でコハク酸発酵を行う為のプラットフォーム菌としてE. aerogenesを候補とし、その耐酸性を 評価することを目的とした。E. aerogenesの酸性(pH5.0)嫌気条件下における比グルコース消費速 度を指標とし、既存の代表的なコハク酸生産菌であるE. coliと比較する方法で、耐酸性能を評価し た。また、E. aerogenesの同環境下における主要なNADH酸化経路を特定する目的で、培養液中 の代謝産物の定量分析を行った。 3章 「E. aerogenesの主要なNADH酸化経路の遮断と異種由来炭酸固定反応の導入によるコハク
15 酸生産能の改良」では、NADH酸化経路の遮断によるNADH供給量の増加、もしくは炭酸固定反 応の導入によるオキサロ酢酸供給量の増加がE. aerogenesのコハク酸生産能の向上に与える影響 を明らかにすることを目的とした。2章で特定した主要なNADH酸化経路であるエタノールと2,3-ブ タンジオール生成経路の遮断、更にはC. glutamicum由来PYCと、A. succinogenes由来PCKの導 入を行い、コハク酸生産能の改良を目指した。 4章 「E. aerogenesのピルビン酸由来副生経路の遮断と、PYCとPCKの共発現によるコハク酸生産 能の改良」では、ピルビン酸から副生する生成物の代謝経路の遮断と、PYCとPCKの共発現がコ ハク酸生産能の向上に与える影響を明らかにすることを目的とした。3章で得られた菌株に対して、 更 に 乳 酸 、 酢 酸 、 2,3- ブ タ ン ジ オ ー ル 生 成 経 路 の 遮 断 と 、 C. glutamicum 由 来 PYC と A. succinogenes由来PCKの共発現を実施し、コハク酸生産能に与える影響を評価した。また、本章で 得られた菌株の酸性環境下におけるコハク酸生産能を評価する目的で、pHを酸性側に制御した 条件でコハク酸培養試験を実施した。 5章 「E. aerogenesのコハク酸生成経路におけるATP獲得量の向上によるコハク酸生産量の改良」 では、コハク酸生成経路で得られるATP量を向上させる方法で、コハク酸生産能の改善を目指した。 4章で得られた菌株に対して、A. succinogene由来PCKの導入とグルコースPTSの遮断を行い、コハ ク酸生産能に及ぼす影響を評価した。また、改変された菌株の酸性(pH5.7)嫌気条件下における コハク酸発酵試験を行い、得られた発酵成績を既報と比較する方法で、技術的進捗を示した。 6章 「結論」では、各章で得られた結果を纏め、本研究を総括し、社会的意義や展望を示した。
16
17
2 章 耐酸性通性嫌気性細菌 E. aerogenes のコハク酸生産菌としての能力評価
2-1. 緒言 これまでに報告されているコハク酸生産菌の殆どが、中性環境下でのみコハク酸を生産する(表 1-2)。なぜなら、これら細菌群の生育可能な pH 領域は、中性付近に限定されており、酸性環境下 では生育出来ない形質であることに起因している。その為、酸性嫌気条件下でのコハク酸発酵を 実現するには、同環境下で効率的にグルコースを資化できる菌種を選択し、新たなコハク酸生産 菌を作出する方法が考えられる。 同環境下でコハク酸発酵を行う為のプラットフォーム菌の選定基準を二つ挙げる。一つは、コハ ク酸生産能を向上させる為の遺伝子改変技術が適用できる菌種。もう一つは、高い耐酸性能を保 持した菌種である。本検討では、両基準を満たす菌種として、E. aerogenes を候補とした。以下に 二つの選択基準に対する論拠を説明する。最初に、該細菌の遺伝子改変技術に関しては、2012 年に Shin 等によって、E. aerogenes KCTC2190 株の全ゲノム配列が決定され、代謝経路の全体像 が推定出来るようになった[34]。また、遺伝子改変技術に関しても、同年、Jung 等によって、λ-red recombination system を利用した遺伝子破壊株の構築法が確立された[35]。また、遺伝子の発現 強化に関しては、E. coli で汎用的に使用されている発現プラスミド(pMW 系、pSTV 系、pUC 系) をシャトルベクターとして利用できる。このように、E. coli で確立された遺伝子改変技術とツールの 一部が E. aerogenes にも適用でき、コハク酸生産能を改善する為の一連の遺伝子改変技術の基 盤は、整備されている。 次に、E. aerogenes の耐酸性について議論する。「耐酸性が高い」という表現には、注意を払う必 要がある。耐酸性が高いという言葉は、低 pH での生存能力(survivability)が高い意味と、生育下 限 pH が低い意味(growth ability)の両方で用いられるが、両者が示す表現型は異なる。例えば、 E. coli は、pH2.0 の環境下で 30 分間暴露しても生存率は高く、生存能力の観点から、E. coli は高18 ず、菌体増殖は出来ない。このように、生育下限 pH の観点からは、耐酸性が高い菌種とは言い難 い。コハク酸発酵などの物質生産においては、糖が資化される速度が重要であり、この点を考慮す ると、我々が求める形質は後者に近いが、同義では無い。本検討で求められる耐酸性は、「酸性嫌 気条件下で早い基質消費速度を有する(assimilality)」と定義する。とりわけ、発酵原料として工業 的に用いられるグルコースの消費速度が、同環境下で高いことが重要である。
E. aerogenes の耐酸性を示す知見として、E. aerogenes IV-2 株は、pH5.0 かつ嫌気条件下でグル コースを資化し、2,3-ブタンジオールを生産することが知られている[31]。このように定性的に、本 菌種の耐酸性能が高いことを示す結果は既に報告されている。一方で、これまでに E. aerogenes の同環境下での比グルコース消費速度を数値化し、他の菌種と比較した事例はない。本章では、 基準株を含む 2 種類の E. aerogenes AJ110637 株と ATCC13048 株の酸性(pH5.0)嫌気条件下の 比グルコース消費速度(g/g[DCW]/h;乾燥重量あたり1グラムの菌体が 1 時間に消費するグルコー ス重量グラム)を E. coli MG1655 株(基準株)と比較定量する方法で、その耐酸性を評価する事を 目的とした。これらの結果を基に、E. aerogenes が酸性嫌気条件下でコハク酸発酵を実施する為に 適したプラットフォーム菌であるかの判断を行う。 また、培養終了後の培養液中の代謝産物を定量分析する方法で、E. aerognese の同環境下にお ける代謝反応の解明を目指した。ゲノム配列の情報から、E. aerognese には乳酸、エタノール、2,3-ブタンジオールの生成経路が存在することが明らかである。これらの生成経路には、NADH の酸 化反応が含まれており、嫌気条件下での酸化還元バランスの均衡化に重要な役割を果たしている。 効率的にコハク酸を生産させる為には、コハク酸生成経路で NADH の酸化を行う必要があり、乳 酸、エタノール、2,3-ブタンジオールの生成経路は、NADH を酸化するという観点で、コハク酸生 成経路と競合する関係にある。本章では、E. aerogenes の主要な代謝産物を同定する方法で、コ ハク酸生産能を向上させる為に、優先的に遮断すべき NADH 酸化経路の特定を目指した。
19
2-2.
実験材料と方法2-2-1. 菌株
本章で使用した菌株を表 2-1 に記載した。
2-2-2. 培地と培養条件
E. coli と E. aerogenes の培養は、汎用的に Luria-Bertani (LB)培地を用いた。必要に応じて寒
天 20 g/L、抗生物質(抗生物質濃度 Km;50 mg/L、Cm;40 mg/L)を添加して使用した。特に本文 中に記載が無い場合は、37 ℃で培養を実施した。嫌気培養時には角型ジャー標準型(三菱ガス 化学社製)に AnaeroPack-A04(三菱ガス化学社製)を 1 個使用した。培地組成を表 2-2 に示す。 2-2-3. 発酵槽を用いた酸性好気条件下と酸性嫌気条件下における培養試験 容量 1L の発酵槽を用いて、酸性好気条件下と酸性嫌気条件下の培養を実施した。酸性好気 条件下の培養手順を補足資料 1、酸性嫌気条件下の培養手順を補足資料 2 に記載した。
2-2-4. E. coli と E. aerogenes の乾燥菌体重量(Dried Cell Weight; DCW)算出式
E. coliとE. aerogenesの乾燥菌体重量(DCW)は以下の手順に従って算出した。OD600値は、 Spectrophotometer U-2001(日立社製)を用いてA600の吸光度を測定した値から、ブランク値を引 いたものを使用した。微量天秤にて1.5 mL容量のマイクロチューブの風袋重量を予め測定し、 OD600値が1, 5, 10, 20, 50, 100の菌体溶液1 mLをマイクロチューブに分注した。その後、4 ℃、 15,000 rpm(21,600 g)で1分間遠心した。上清を捨て、0.1 N 塩酸を1 mL分注し、3回洗浄を行っ た。上清を捨てた菌体ペレットを、真空乾燥機にて一晩、60 ℃で乾燥した。微量天秤にて乾燥重 量を測定。再度、乾燥機にて一晩乾燥させ、乾燥重量に変化が無い事を確認した。得られた乾燥 重量から風袋重量を差し引いた値をDCW値とした。本方法により得られたOD600値とDCWの換算 式を以下に示す。E. coli;g[DCW]/L=0.287×OD600 E. aerogenes;g[DCW]/L=0.291×OD600
20 2-2-5. グルコース濃度、OD、有機酸、アルコールの定量
グルコース濃度は、AS-310 Biotec Analyser (サクラ精機社製)を用いて酵素法(酵素電極)にて 定量分析した。OD600値は、 U-2001 spectrometer (日立社製)を用いて分析した。 有機酸は HPLC(High Performance Liquid Chromatography)により定量分析を行った。有機酸分析の詳細な 条 件 を以 下 に 示 す [9]。分析機器;ELITE LaChorum (島津社製)、<分 離条件>カラム ; Shim-pack SCR-102H 二本直列接続、ガードカラム SCR-102H、移動相; 5 mM p-トルエンス ルホン酸、流量 ; 0.8 ml/min、温度 ; 50 ℃<検出条件>緩衝液; 5 mM p-トルエンスルホ ン酸 および 100 μM EDTA を含む 20 mM Bis-Tris 水溶液、流量 ; 0.8 ml/min、検出器; CDD-10AD polarity + response SLOW temperature 53 ℃(カラム温度+3℃) scale1×24 μS /cm。エタノール, 2,3-ブタンジオール, アセトインの分析は GC(Gas Chromatography)により定量 した。分析機器;GC4000 (GL-Science社製)、検出方法;flame ionization detector、 分離カラム; A TC-BOND Q capillary (GL-Science社製).
21 菌株 注釈 薬剤耐性 引用・入手先 Strains E. coli strain MG1655 基準株 ― ATCC1 E. aerogenes strains ATCC13048 基準株 ― ATCC1
AJ110637 野生株 ( FERM BP-10955) ― AIST2 [33]
LB 培地 最終濃度 Bacto-tryptone 10 g/L Bacto-yeast extract 5 g/L NaCl 5 g/L SOC 培地 最終濃度 Bacto-tryptone 20 g/L グルコース 10 g/L Bacto-yeast extract 5 g/L NaCl 5 g/L MS 培地 最終濃度 グルコース 40 g/L MgSO4・7H2O 1 g/L Bacto-yeast extract 2 g/L (NH4)2SO4 1 g/L KH2PO4 1 g/L MnSO4・5H2O 10 mg/L FeSO4・7H2O 10 mg/L ビオチン 1 mg/L 1)
American Type Culture Collection
2) 独立行政法人 産業技術総合研究所 特許生物寄託センター 表 2-1 本章で使用した菌株一覧 表 2-2 本章で使用した培地一覧 120 ℃、20 分間 オートクレイブで殺菌後、使用した。 120 ℃、20 分間 オートクレイブで殺菌、放冷後、終濃度が 10 g/L になるように別殺菌したグルコースを添加した。 ステリカップ(SCGPU05ER)を使用し濾過滅菌を行った後、使用した。
22 1. グリセロールストックから菌体を、LB プレートに均一に塗布し、37 ℃、好気条件下で 16 時間前 培養を行った。 2. LB プレート上の菌体をかき取り、10 mL の MS 培地に懸濁した。 3. OD600を測定後、OD600が 40 になるように MS 培地で希釈した。 4. OD600 40 の培養液 10 mL を 290 mL の MS 培地に稙菌した。この時、植菌後の MS 培地の OD600 は、約 1.3 となり、DCW 換算で約 0.4 g[DCW]/L となるように調整した。 5. 稙菌後の MS 培地 300 mL を全量 1 L 発酵槽に移液し、BMZ-P 培養装置(ABLE Biott 社製) にて培養を行った。培養管理方法として、培地 pH は 5.0 になるにようにアンモニアガスにて制御し、 培地温度は 34℃で制御を行った。また、300 mL/min の空気を 0.22μm フィルターを通して通気し、 培地の溶存酸素濃度が 5%以上になるよう撹拌制御を行った(各種設定値; 培養 pH 5.0 ± 0.05、温度 34.0 ± 0.2 ℃、撹拌 700 rpm)。 6. 約 2 時間ごとに 2 mL の培養液をサンプリングし、OD600と培地中のグルコース濃度を測定し た。 補足資料 1 酸性好気条件下における培養試験の操作手順 酸性好気条件下における培養試験の操作手順
23 1.グリセロールストックから菌体を、LB プレートに均一に塗布し、37 ℃、好気条件下で 16 時間前 培養を行った。 2. 菌体が生育した LB プレートを、AnaeroPack-A04 を用いて、37 ℃にて 16 時間、嫌気条件下に 馴致させた。 3.LB プレート上の菌体をかき取り、10 mL の MS 培地に懸濁した。 4. OD600を測定後、OD600が 10 になるように MS 培地で希釈した。 5. OD600 10 の培養液を 290 mL の MS 培地に稙菌した。この時、培地の OD600は、約 0.3 となり、 DCW 換算で約 0.1 g[DCW]/L となるように調整した。 6. 稙菌後の MS 培地 300 mL を全量 1 L 発酵槽に移液し、BMZ-P 培養装置にて培養を行った。 培養管理方法として、培地 pH は 5.0 になるにようにアンモニアガスにて制御し、培地温度は 34℃ で制御した。培養中は、嫌気条件にする為、300 mL/min の高純度窒素ガスを 0.22μm フィルター を通して通気した(各種設定値; 培養 pH 5.0 ± 0.05、温度 34.0 ± 0.2 ℃、撹拌 700 rpm)。 7. 約 2 時間ごとに 2 mL の培養液をサンプリングし、OD600と培地中のグルコース濃度を測定した 酸性嫌気条件下における培養試験の操作手順 補足資料 2 酸性嫌気条件下における培養試験の操作手順
24 2-3. 結果
2-3-1. E. aerogenes の酸性(pH5.0)好気条件下における培養特性
代表的な発酵原料の一つであるグルコースを用いて、E. aerogenes と E. coli の酸性(pH5.0)好 気条件下における比グルコース消費速度(乾燥重量 1 グラムの菌体あたり、1 時間に消費するグル コース重量グラム; g/g[DCW]/h)を測定した。本検討では腸内細菌群の培養に汎用的に用いられ る MS 培地を使用し、1 L 発酵槽にて培養試験を行った。培養条件は、温度 34 ℃、培養 pH は 5.0 制御にて実施した。
E. aerogenes からは AJ110637 株(野生株)と ATCC13048 株(基準株)を選択し、E. coli からは
MG1655 株(基準株)を選択した。LB 培地で一晩前培養した菌体を 0.4 g[DCW]/L になるように 300 mL MS 培地に稙菌し、培養を行った。継時的に培養液をサンプリングし、菌体量と培地のグル コース濃度を測定した。結果を図 2-3 に纏めた。
図 2-3 A に示すように、E. aerogenes の方が、E. coli よりも速い生育速度が観察された。実際、培 養 8 時間目の E. aerogenes ATCC13048 株と AJ116037 株の菌体量は、それぞれ 8.10 と 8.07 (g[DCW]/L)に対して、E. coli MG1655 株は、E. aerogenes の約 1/3 に相当する、2.87(g[DCW]/L) であった。また、図 2-3 B に示すように、E. aerogenes ATCC13048 株と AJ116037 株は約 8 時間 の培養で、初期濃度にあたるグルコース約 40 g/L を完全に消費したのに対して、E. coli MG1655 株は 12 時間の培養で消費したグルコース量は、僅か 12.0 g/L であった。これらの値から、酸性 (pH5.0)好気条件下の比グルコース消費速度(g/g[DCW]/h)を算出すると、降順に E. aerogenes AJ116037 株、E. aerogenes ATCC13048 株、E. coli MG1655 となり、それぞれ 1.43、1.42、0.69 g/g[DCW]/h の 値 を 示 し た 。 以 上 の 結 果 よ り 、 酸 性 ( pH5.0 ) 好 気 条 件 下 で は 、 E. aerogenes AJ116037 株と ATCC13048 株が、E. coli MG1655 株よりも早い速度で増殖をし、2 倍以上高い比グ ルコース消費速度を示した。
25 2-3-2. E. aerogenes の酸性(pH5.0)嫌気条件下における培養特性 酸性(pH5.0)嫌気条件下における比グルコース消費速度を算出する目的で、1 L 容量の発酵槽 を用いて培養試験を実施した。嫌気条件の環境にする為、培養を通して、高純度窒素ガス(< 99%)を通気した。LB 培地で 16 時間培養した菌体を、Anaeropack A-04 を用いて、嫌気条件下で 16 時間馴致させた後、菌体を 0.1 g[DCW]/L になるように 300 mL MS 培地に稙菌した。継時的に、 サンプリングを行い、菌体量(g[DCW]/L)と培地のグルコース濃度を測定した。結果を図 2-4 に纏 めた。 図 2-4 A に示すように、E. coli MG1655 株は、菌体量が 0.5 g[DCW]/L 以上増加しなかったのに 対して、E. aerogenes AJ116037 株と ATCC13048 株では、培養終了まで菌体量の増加が観察され た。最終的に、23 時間の培養で、E. coli MG1655 株、E. aerogenes ATCC13048 株、E. aerogenes AJ116037 株の菌体量は、それぞれ 0.53、1.71、2.11 g[DCW]/L となり、E. aerogenes AJ116037 株 が最も高い菌体量を示した。また、図 2-4 B に示すように、グルコース消費量も E. aerogenes AJ116037 株が最も高い値を示した。実際、23 時間の培養で、E. coli MG1655 株と E. aerogenes ATCC13048 株が、それぞれ 7.6 g/L と 28.8 g/L のグルコースを消費したのに対して、E. aerogenes AJ116037 株は 37.8 g/L のグルコースを消費した。
これらの測定値から酸性(pH5.0)嫌気条件下の比グルコース消費速度(g/g[DCW]/h)を計算す ると、降順に E. aerogenes AJ116037 株、E. aerogenes ATCC13048 株、E. coli MG1655 となり、それ ぞれ 1.41、1.27、0.76 g/g[DCW]/h の値を示した。以上の結果より、酸性(pH5.0)嫌気培養条件下 では、E. aerogenes AJ116037 株が、最も高い比グルコース消費速度を示した。以上の結果から、E. aerogenes AJ116037 株をコハク酸生産菌の新たなプラットフォーム菌に選定した。
26
図 2-3 酸性(pH5.0)好気条件下における菌体量と培地グルコース濃度の継時的変化 左図 A; 菌体量(g/[DCW]/L)、右図 B; 培地中のグルコース濃度(g/L)。
●;E. coli MG1655、 ▲;E. aerogenes ATCC13048、■; E. aerogenes AJ116037。
図 2-4 酸性(pH5.0)嫌気条件下における菌体量と培地グルコース濃度の継時的変化 左図 A; 菌体量(g[DCW]/L)、右図 B; 培地中のグルコース濃度(g/L)。
27
2-3-3. E. aerogenes AJ116037 株の酸性(pH5.0)嫌気条件下における代謝産物の同定
E. aerogenes は、嫌気条件下で水素、エタノール、2,3-ブタンジオールを生成物として培地中に
蓄積する[30-33]。本章では、これらの代謝産物の定量測定を行い、酸性(pH5.0)嫌気条件下にお ける E. aerogenes AJ116037 株の主要な NADH 酸化経路の特定を目指した。酸性(pH5.0)嫌気条 件下で培養後の培養液を遠心し、フィルター濾過により除菌した上清液を用いて各種代謝産物の 分析を行った。有機酸の定量は HPLC、アルコールの定量は GC により解析した。代謝産物の定量 解析の結果を表 2-3 に纏めた。
酸性(pH5.0)嫌気条件下では、E. coli MG1655 株は、乳酸を主要な生成物として培地中に蓄積 した。培地中の乳酸濃度は、3.5 g/L となり、消費されたグルコース重量の 46.0%が乳酸に変換さ れた。以上の結果から、E. coli MG1655 株では乳酸生成経路が主要な NADH 酸化経路として機 能していることが示された。一方、E. aerogenes AJ116037 株は、エタノールと 2,3-ブタンジオール が主要な生成物となり、それぞれ 10.7 g/L と 9.8 g/L の濃度が検出された。エタノールと 2,3-ブタン ジオールの両生成物は、消費したグルコース重量あたり、それぞれ 28.3%と 25.9%の収率で変換さ れた結果を示した。また、AJ116037 株では、ATCC13048 株と比較すると、乳酸と酢酸の生成量が 低い特徴が観察された。ATCC13048 株が、それぞれ 6.7 g/L と 3.9 g/L の乳酸と酢酸を生産するの に対して、AJ116037 株では、約半分に相当する 3.5 g/L と 1.9 g/L であった。以上の結果から、E. aerogenes AJ116037 株では、エタノールと 2,3-ブタンジオール生成経路が、主要な NADH 酸化経
路として機能していることが分かった。また、E. aerogenes AJ116037 株の培養液には、1.4 g/L のコ ハク酸が検出されており、同株が酸性(pH5.0)嫌気条件下でコハク酸生産能を有する結果を得 た。
28 表 2-3 各菌株の各培養条件におけるグルコース消費量と生成物濃度 菌株名 培養条件 グルコース 消費量 g/L 最終生成物 g/L ピルビン酸 リンゴ酸 ギ酸 コハク酸 乳酸 酢酸 EtOH1 2,3-BuOH2 アセトイン E. coli MG1655 好気3 12.0 ± 0.63 0.1 ± 0.14 ND5 ND ND ND 0.9 ± 0.1 ND ND ND 嫌気6 7.6 ± 1.0 0.2 ± 0.1 < 0.1 1.3 ± 0.1 0.4 ± 0.1 3.5 ± 0.4 1.5 ± 0.2 0.7 ± 0.1 ND ND E. aerogenes ATCC13048 好気 43.4 ± 2.1 ND ND 0.1 ± 0.1 ND ND 0.2 ± 0.1 ND 0.2 ± 0.1 0.6 ± 0.1 嫌気 28.8 ± 1.2 0.5 ± 0.1 < 0.1 0.9 ± 0.3 1.4 ± 0.1 6.7 ± 0.6 3.9 ± 0.3 7.2 ± 0.5 6.8 ± 0.5 < 0.1 E. aerogenes AJ110637 好気 43.4 ± 1.4 ND ND 0.2 ± 0.1 ND ND ND ND 0.8 ± 0.1 1.8 ± 0.2 嫌気 37.8 ± 0.6 0.4 ± 0.1 < 0.1 0.9 ± 0.3 1.4 ± 0.1 3.5 ± 0.2 1.9 ± 0.2 10.7 ± 0.6 9.8 ± 0.5 0.1 ± 0.1 1) EtOH はエタノールを意味する。 2) 2,3-BuOH は 2,3-ブタンジオールを意味する。 3)
E. coli MG1655 株は 12 時間、E. aerogenes ATCC13048 株と E. aerogenes AJ110637 株は 8 時間培養後の培養液を分析した。
4)各値は n=3 の平均値を意味し、±以後の値は標準偏差値を意味する。
5)
ND は未検出を意味する。 6)
29 2-4 考察
2-4-1. E. aerogenes AJ110637 株の酸性(pH5.0)嫌気条件下における比グルコース消費速度
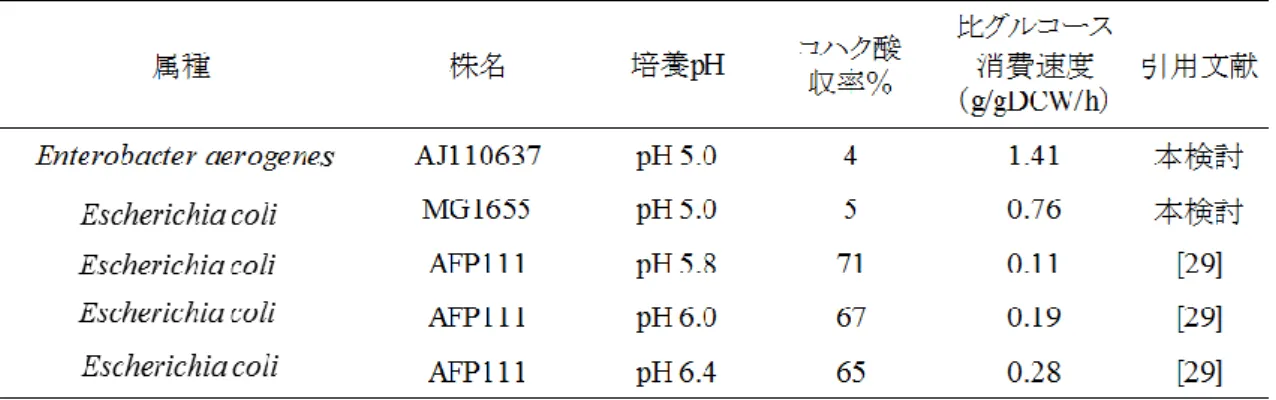
Lu 等は、E. coli AFP111 株を用いて、pH がコハク酸生産能に及ぼす影響を検討した[29]。表 2-4 に示すように、E. coli AFP111 株は、グルコースから収率 60%以上でコハク酸を生産する能力 を有する。この検討の中で、E. coli AFP111 株は、pH 6.4 で最も高い比グルコース消費速度が観察 され、その値は 0.28 g/gDCW/h であった。この結果と本検討で得られた E. coli MG1655 株の値 (pH5.0 0.76 g/gDCW/h)を比較すると、コハク酸生産能の向上により比グルコース消費速度は顕著 に低下したと推定できる。また、E. coli AFP111 株は pH の低下に依存して、比グルコース消費速度 も低下した。E. coli AFP111 株の pH5.8 における比グルコース消費速度は、0.11 g/g[DCW]/h となり、 pH6.0 から僅か pH を 0.2 下げただけで、比グルコース消費速度は 40%以上低下した。この傾向を 加味すると、E. coli AFP111 株のコハク酸生産能が維持できる限界 pH は、5.8 付近であると推察さ れる。一方、E. aerogenes AJ110637 株は、pH 5.0 の環境下で E. coli MG1655 株よりも 1.9 倍高い 比グルコース消費速度が観察された。恐らく、E. coli と同じように E. aerogenes においてもコハク酸 生産能の向上に比例して、グルコース消費速度が低下すると推定されるが、どれくらい低下するか は、AJ110637 株から実際にコハク酸生産菌を作出し、確かめる以外の手段はない。いずれにして も、E. aereogens は、これまでにコハク酸生産菌として開発された細菌の中では、酸性嫌気条件下 で高い比グルコース消費速度を有していることが示された。
![図 2-4 酸性(pH5.0)嫌気条件下における菌体量と培地グルコース濃度の継時的変化 左図 A; 菌体量(g[DCW]/L)、右図 B; 培地中のグルコース濃度(g/L)。](https://thumb-ap.123doks.com/thumbv2/123deta/5825872.541737/33.892.146.742.597.940/おける菌体培地グルコース濃度継時左図菌体量右図培地グルコース.webp)