金 沢大学十全医学会雑誌 第84巻 第5‑6号 419‑426 (1975)
大 腸 菌 にお ける デ オ キ シ リボ シル転 移 反 応 につ いて
‑ チ ミ ジ ン フ ォ ス フ ォ リ ラ ー ゼ及 び プ リ ン ヌ ク レオ シ ドフ ォ ス フ ォ リ ラ ー ゼ に よ る デ オ キ シ リボ シル転 移 反 応 一
金沢大学 医学部第二生化学 講座 (主任 :久野 滋教授 )
林 道 子
(昭和50年5月13日受付)
動物お よび微生物 にお いて種 々のnucleosideの 加 リン酸分解 あ るいはnucleosideと核 酸 塩 基 相 互 間 のpentose転移反応 はnucleoside代 謝上重要 な 役割 を果 してい る. この加 リン酸分解反応 はnucleo sidephosphorylaseによ って
Ⅹ‑deoxyribose+Pi一一Ⅹ+deoxyribose‑1‑ phosphate
の如 く可逆的 に触媒 され る.一般 的 には反応 は生体 内 で右側 に進行 し主 と してCatabolism に 関 与 す る と 考 え られ るが ,充分 な deoxyribose‑卜phosphate 供給下 で はnucleoside合成 を行 うことがで きる.
deoxynucleosideのphosphorylaseと して thymi dinephosphorylase(tpp)とpurinenucleoside phosphorylase(pup)の2種 が知 られ て い る1)〜3‑
これに対 して deoxynucleosideの pentose転移反 応 す なわち Transdeoxyribosylation(TDR反 応 )
は何 れの生物 内で も重要 な反応 と して存 在 す るが , こ れ を触媒 す る酵素,trams‑N‑deoxyribosylaseは現 在迄 の ところ乳酸菌頬 でのみ見 出 され4)‑6).他 の 生 物 には独立 した静索 と して存在 して いない .従 って他 の 生物 にお けるpurineお よびpyrimidine間 の TDR 反応 は tpp と pupの両酵素 の共 同作 用 によ る も の と考 え られて きた7)8).す なわち
Thymidin。+PiLPE Thymine+d。。Xyrib。S。‑ 卜phosphate
Adenine+deoxyribose‑1‑phosphatep‑u‑P deoxyadenosine+Pi
の連続 した反応 によ って転移反応 が行 われ る. この反 応 は両酵素 の性質 か ら考 えて Z.nuitroで当然起 り得
419
る反応 で はあ るが ,必 ず しもZ'nvivoで同 じ反 応 形 式 で起 る ことは意味 しない .上記 の反応 が効 果的 に起 るためには中間体 の deoxyribose‑1‑phosphateが 充分量産生 され ることが必要 であ り,事実 Z'nvitro で の TDR反応 はinvivoでの反応 に比 して着 る し く遅 い ことが知 られてい る8). しか しなが ら大 腸 菌 で は上記 の2酵素 の何 れかが欠 除 した場合 TDR反 応 が起 らない ことか らinvivoの反応 も両酵素 が 関 与 して い ることは間違 いな く9),invivoで は効 果 的 に TDR反 応 を行 うために両酵素 の物理 的 配 置 等 に未 知 の機構 が存在 して い る もの と推定 され る.本研究 で は大腸菌か ら分離精製 した tppお よび pupの 性 質 とinvitroでの TDR反応 の条件 お よ び特 性 を 明 らか にす る目的 で行 われ た .その結果 inz)itroにお いて見 られ るTDR反応 は上記 の2反 応 の 単 な る連 鎖 によ る もので はな く,恐 らく両酵素 は弱 い相互作用 によ って complexを形成 し, このcomplexに よ って TDR反応 が行 われて い る もの と推定 された .
方 法
酵素源 と して大腸菌Y70122を用 いた10).本 菌 はth‑
ymine要求性 で tppと pupを constitutiveに産 生 す る.菌 はnutrientbrothで培養L stationary
phaseに遷 した菌 を集 めて酵素抽 出 を行 った . 使用 した薬 品 はすべて市販 の もの を用 いた .Contr‑ olled・PoreGlass,CPG‑10‑240‑(120/240)は ELEC TRO‑NUCLEONICS,lNC.よ り購 入 した .
酵素反応 の測定
tpp活性 の測定 はRazzelお よび Khoranall) の 方法 によ った .す なわち16mMの thymidine,0.05M
succinate‑0.05Msodium arsenate(pH6.0)と酵 Transdeoxyribosylation Activity in Escherichia coli. Trans、eoxyribosylation catalyzed by thymidinephosphorylaseand pJurine nucleoside phosphorylase.Michiko Hayashi,DepartmentofBiochemistry(Ⅱ)(Director:Prof.S.Kuno)School of Medicine, Kanazawa University.
420
素 を含 む反応 液 (0.20ml) を10分 間37oCで 反 応 さ せ た後 ,1.5mlの0.3NNaOHを加 え,300mfJの吸 光度 を 測定 した .吸光度1.0の増 加 が0.471〟Moleのthymirle
生成 に相 当す る. 1〟Moleの thymineを1分 間 に 生 成 す る酵素量 を1unitと した .
pup活性 の測定 は Tsuboiお よ び Hadson12‑ の 方 法 の変法H)を用 いた .す なわ ち5mMdeoxyguano sine,0.1Msodiumacetate‑0.05Msodium arsenate (pH6.0)と酵素 を含 む反応 液 (0.25ml) を37oC10
分 間反応 させ た後 .0.5mlの20%Na2C03を加 え ,更 に
0.75mlの6倍 稀釈 の PhenoIReagentを加 え て370 C20分 間静 置 した後,660m〟で の吸光 度 を 測 定 した . 吸光度1.0の増加 が guanine0.278〟Mole生成 に相 当 す る.1〟Moleの guanineを1分 間 に産 生 す る酵 素 量 をlunitと した .
TDR活性 はMunch‑Petersenの方 法13)に 多 少 の 変更 を加 え た‖).す なわ ち4mMの thymidine,lmM
の adenine.10〟Mの リン酸,0.016Macetatebuffer (pH6.5)お よび酵素 液 を含 む0.50mlの反 応 液 を370 C30分 間反応 させ た後, 10N過塩 素 酸 0.025mlを 加 えて反応 を止 め , この間生 じた deoxyribose リ ン 酸 お よびdeoxyadenosineをBurtonの法15一に よ り測定 した .tppに よ るthymidine分 解 を 補 正 す るため adenineを除 いた反 応 液 を同様 に処理 し, こ の値 を上 記 の値 か ら差引いた もの を TDR活 性 と し た .1分 間 に1m〟Moleの deoxyadenosineを 生 成 す る酵素 量 を1 unit と した .
蛋 白量 はLowry等 の方 法16)で Bovine serum albumineを standardと して測定 した .
結 果
辞表 の精製
6gの菌 を Taketo及 びKunoll)の方 法 に従 っ て
osmoticshockを行 い .上 澄 液 (shocked fluid) 30mlを得 た . これ を図1に示 した方 法 で 精 製 を 行 っ た .す なわ ちshockedfluidを兼 溜水 に対 して12時 間透析 した後 ,遠心 して上 清 と沈激 に分 け ると.tpp
の大 部分 は上 清 に,pupの大 部分 は沈激 に集 った .
tpp:上 記 の上 清 を80%飽和 の硫安 で沈澱 し,そ の 沈毅 を約1mlの0.05MTris‑ClpH7.5に溶 解 L Sep・
hadexG‑200(2.6×50cm)カ ラ ム ク ロ マ トグ ラ フ ィーを行 った .tppはpupよ り僅 か に遅 れて流 出す るので その活性 部 を集 めて80%飽 和 の 硫 安 で 沈 毅 さ せ ,少量 の0.05MTris‑CIpH7.5に溶解 した後 .同緩 衝液 に対 して透析 を行 った .遠心後 上 清 を あ らか じめ
0.2MKCト0.05MTris・ClpH7.5で平 衝化 した CPG‑
10カ ラム (1.6×30cm)に流 し.次 に同 緩 衝 液 で 溶 出 した .活性 部分 を集 めて透析 した後,0.2MKC卜0.02 M glycylglycine pH8.0で平 衝化 したCPG‑10カ
ラムに流 し.同緩 衝液 を250ml流 した後 .0.02MglyG・ ylglycinepH8.0で溶 出 した .混在 す るpupは0.2 MKClを含 む緩 衝 液 で溶 出 され,tppは後者 の緩 衝液 で溶 出 され る.
pup:shockedfluidを透析 して得 られ た 沈 激 を
0.05MTris・ClpH7.5に溶解 し,遠心 に よ り不 溶物 を 除去 した後,DEAE‑celluloseカ ラム (2.6×30cm)
に吸着 させ た . カ ラムはあ らか じめ0.05M Tris‑CI pH7.5で平衝化 .辞素 を吸着 させ た後,KClを 含 む 緩 衝 液 の Iineargradientで溶 出 した (0.05MTris‑Cl
pH7.5‑0.6MKClin0.05MTris‑CIpH7.5各500 ml).pupを含 む フ ラク ショ ンを合 して 透 析 を 行 っ た 後 ,同一 条件 で再 び ク ロマ トグラフ ィ‑を行 った .宿 性 を含 む フラク シ ョンを合 した後80%飽和 の硫安 で塩 析 し,少 量 の0.05MTris・CIpH7.5に溶解 し,同緩 衝 液 で透析 を行 った .この酵素 液 をあ らか じめ0.2MKC1
‑0.02Mglycylglycine pH8.0で平 衝化 した CPG‑
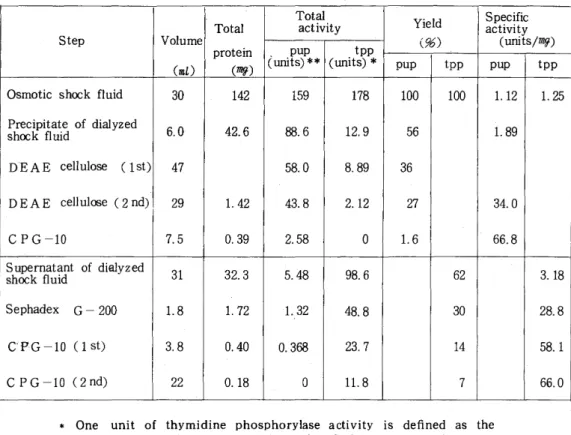
'lloヵ ラム (1.6×30cm)に流 し, 同 緩 衝 液 で 溶 出 し た .表1に両酵 素 の 精 製 結 果 を示 す .何 れの辞素 も
shockednuidか ら約50倍 に精製 され た . しか しデ ィ ス ク電 気泳動 で は酵素 活性 と一 致 す る主 bandの 他 に2,3の淡 い bandが認 め られ た .
Fig.1.Purificationschemeofthymidine phosphorylaseand purinenucleoside phosphorylase.
E.COli
osmotidtShock Shockfluid
diaLyzeLdagainstdistiLLcLdwater SUpqnatant
(tppfr.)
I80'/・ sat・(NH▲)2SO'
ppt StplhadexG・200 Coturnnchrα¶itOgraPhy
∫ 80'/・sNat・(NH4)2SO'
PPt CP6‑l10
Columnchrornatography
(1st) CPG・I10
Colurnnchrornatography (2nd)
tIpp
PrcLCipitale (puplr.) DEA∫EICeLLutose CoturnnchromatograPhy
Hst) DEAlE・ceHuLosc・ CotumnchroTnatOgraPhy
(2nd)
∫ 8 0 I / ・ s
at・(NH̀)2SOAPPt CP6‑↓10
CoLurnnchronatography P
∫
uP柿
大腸 菌におけるデオキ リボ シル転移反応
TableI Purification ofpurine nucleoside phosphorylase and thymidihe phosphorylase
421
Step Vo(l7umenL) prTotot(alei呼 )n (̲ pup1mitSToa)c**ttaivilt(yunittspp)*pupYi(%)eldtpp Sacpt(eiuncviiftiiytcs′mg) pup tpp Osmoticsh∝kfluid 30 142 159 178 1
00 100 1.12 1,25 Precipitateofdialyzed
sD'DEAE celh∝kfEAE cluidelllululo岱e(2nd)se (lst) 6.47209 42.1.462 8548.8.3.608 8.2.12.81992 3566 314..890
27
CPG‑10 7,5 0.39 2.58
0
1.6 66.8Supematantofdi alyzed
sh∝kfluid 31 32.3 5.48 98.6 62
3.18 SephadeX G‑200 1.8 1.72 1.J32
48.8 30 28.8 CP.G‑10(1st) 3.8
0.40 0.368 23.7 14 58.1 CPG‑10(2nd) 22 0.18
0
ll,87 66.0
*One unit ofthymidine phosphorylase activity
is de負ned as the amount required to produce 1 FLmOle ofthymille
Per min.
〜 + One unit of purine nucleoside phosphorylase activity
is defined as the amountrequired to produce 1FLmOle ofguanin
e per min.
tppおよ び pupの カラム クロマ ト
グラフ ィーにお ける挙動 :
図2にtppおよびpupの SephadexG‑20 0ク ロマ トの挙動 を示す .pupはtppよ りも多少早 く溶 出 され,TDR活性 は両酵素 の 中 間 部 に認 め られ る が ,両酵素 の完全
分離 はで きなか った.
図3はDEAE・cellu
loseクロマ トの挙動 であ るが . SephadexG‑200の場合 と
逆 に tppが pupよ り も 少 し早 く溶 出 されて くる
.緩 衝液 あ るいは塩濃度 を種 々変えて溶出条件 を検討 したが .両酵素 の完全 な分離
は不可能であ った.図4および図5にCPG‑10カ
ラムにおけ る挙 動 を 示 す.0.02M glycylglyci
nepH7.0あ る い はpH8.0 で溶出す るとtppおよび pupの
何 れ も略 々 void volumeの部 に溶 出 され る.これに
対 して0.2MKC1‑
0.02MglycylglycinepH7.0で は何 れ の 酵 素 も カ ラ ムに吸着 されて溶出 されない
.また0.2MKCト0.02M glycylglycine
pH8.0で溶 出 した場合 pupは溶 出 さ れて くる ものの カラム容積 の約2倍 の溶
出液 が必要で あ り,tppはpH7.0の場合 と同様溶
出す ることはで き ない.従 って0.2MKC1‑0.0
2MglycylglycinepH8.0 で tppとpup
を完全 に分離 す る こ と が 可 能 で あ る.この高塩濃度
での吸着 はCPG‑10の非 特 異 的 吸 着 を妨 げ るpo
lyethyleneglycolを溶 出液 に添加 す ることによ って完全 に防 ぐことがで き
る. しか し塩濃 度 の上昇 によって吸着 が増大 す る理 由 は不明 で あ る .
tppおよび pupによるthymidine,adenine のtppTDRお よび反応 :pup存在下 間
で thymidineとadenine よ り de
柿
20
0 400
a
0.02MGLycyLgLyc
lne 訳
化 妃l
u Lu
Js
山]0∈rfLLJdd十OSU!uJsa)o∈rruddtu!∈\sato∈rfuddISN0 0LOS0 0LOS00LOSu!uJ s小一OuJrrUdndU盲 JSa
]0∈r f
∈ d d
tm0l 6b.04'/。0P0olyeth
yleneglyCOL
i n
O・02MGlycyLgLycin .U]u lS小
】0∈rT
udndOOLOS51
0 15 20 0.04'/'PoLyeth
yLeneglycoL
inO・2MKCL・0・02Mー < in0,02MG
LycyLgLyclne
GLy
cyLgLycllrv
5 10 15 20
]uJStt.un ⊂﹀
tO [正二
⊂l
Cl Fig.2. Gel filtration pattern o
f shock nuid through Sephadex G‑200.To a
column (2.6
×50cm)equilibrated with 0.05
M Tris・CIpH 7.5,2mlofshock 角uid (contain
ing 22 units oftpp and 6.8unitsofpup)was
applied and eluted with 0.05 M Tris‑ClpH 7
.5.Activity was measured as described i
n ■'METHODS"
‑ ‑‑ .protein:・・‑‑・‑1・‑.top;and
,Pup.
Fig.3. Chromatography of thy
midine phos‑
0・5 phorylase and purinenucleosid
e phosphory‑ 1ase on DEAE cellulose column.
Forexperi・ mentaldetails,see
..RESULTS". Columnsize
(2.6×30cm)
‑ ‑‑,protein;一日‑‑一一‑・,
tpp; .pup u
Z∈
ls。 ]仝ヒrfu dndOO10SuTヒ JSa]0ErrudndFig.4.Chromatographyofthym idinephosp・horylaseandpurinenucleosidephospho、rylaseonCPG‑10atpH7.0.The0.50unitsoftppand0.65unitsofpupinavolumeof0.2
mlwereappliedonacolu mnofCPG‑10
(0.5×30cm),andelutedwitha)0.02Mglyc・ylglycinepH7.0,b)0.04%polyethyleneglycolin0.02MglycylglycinepH7.0,C)15mlof0.2MKClin0.02MglycylglycinepH7.0followedby0.02MglycylglycinepH7.0,andd)15mlof0.04%polyethyleneglycolin0.2MKC卜0.02MglycylglycinepH7.0followedby0.04%polyethyleneglycolin0.02Mgトycylgly cinepH7.0.,tpp;一 一
大腸 菌 におけるデオキ リボシル転移反応
Fig5. Chromatography ofthymidine phosp・
horylaseandpurinenucleoside phosphorylase on CPG‑10atpH8.0.Methodswere same as described in Fig4,except being used the buffer,pH 8.0.
U!∈\so]CUJrfuddI
b
Ol04 '/.PoLyetrvLery gLycoL
in0.02MGLvCVtdLv亡inさ
Ⅳ︼8HP utuJsa]0ErfuJdndOOLOS
5 10 ml
u!uJJsむ︼0∈rTtDddI OSSN00LOSu!∈\salCuJr(ヒddl
0.2MKCLin
o・02MGLycytgLycin㌻一一一 ・0・02MGtycylgLycine
10 15 20 25ml
OIQ4'ILEp.IyF̲try̲Le̲ワe̲gLycoL 0.04'/'PoLyethyLenegLycoL in0.2MKCL・0.02M
GlycylgLyclne inO・02MGtycyLglycLn
e
10 15 20
U!uJs山︼0ErruJdつd
OOtOS tJ!u
lSa
)0∈rhdndOOLOSFig6.Dependency oftransdeoxyribosylation on inorganic phosphate.
Theexperimentcarriedoutasdescribedin..M ETHODS",excepttheconcentrationofinorgani cphosphate.
+ ・ ‑ M J.
activity in the presence ofadenine;〇一一‑○,activity in the absence ofadenine;
△‑ △,the difference (transdeoxyribosylati‑ On activity).
u!uJJs心lOuJrfu
10
20 30 40 50
phosphate(JJM) Fig 423
7.PH optima ofthymidine phosphoryla‑
se.p
urine nucleoside phosphorylase and tra‑ nsd
eoxyribosylation. Activity was measured asdescribedin‑'METHODS"
exceptbeing used the indicated buf
fer. a)Purinenucleosideph
osphorylase
● ‑●
,0.1M acetatebuffer;
△‑ △,0.1M imida
zolebuffer. b)Thymidinephospho
rylase
き●
,0.05M succinatebuffer;
△‑ △,0.05M imidazolebuf fer. C)Transdeoxyribosylation
activity
● ‑● .
0.016M acetatebuffer;
Ⅹ‑ X,0.016M Tris‑maleat
ebuffer; A‑ A.0.016M TrisIClbu
f fer.
4 5 p6 7 8
H
TableⅡKm value for inorganic phosph
ate
Enzyme 也
Thymidinephosphoryl