December 1991
Jpn J Anim Reprod Vol 37 (5)
SHIMAMURA
AWARD
LECTURE
FOR 1991—
Studies
on Relationship
of Biological
Rhythms
to Reproductive
Functions
in Animals
Noboru MURAKAMI
Department of Veterinary Physiology, Faculty of Agriculture, University of Miyazaki, Miyazaki 889-21
Summary. Biochemical, neural, endocrine and behavioral rhythms play a central role in many different aspects of the reproductive process in animals. These rhythms provide the basis for temporal organization of the reproductive functions of individual animals. In this paper, I introduce our results about the relationship of biological clock to reproductive function, analysis of mechanisms in biological clock, and manipulation of circadian rhythms. Main contents are as follows:
1) The ovulation is induced between 14 and 16 hr after stimulation dissociating entirely with solar
hours in light estrous rat. The effectiveness of the cervical stimulation for induction of ovulation in
light estrous rats is related to not only the duration of light estrus but also the time after transfer to
LL. 2) Both nocturnal and diurnal prolactin surges are controlled
by a circadian system. 3)
Pseudopregnancy
is induced by a single injection of progesterone
at a special time. 4) The time of
parturition
and of pre-partum
progesterone
decrease is closely associated with an endogenous
circadian system. 5) Food restriction is more potent zeitgaber than light in circadian adrenocortical
rhythms. 6) The circadian rhythms of blinded rat pups is entrained by the nursing dam. 7) Social
cues have a profound effect on the synchronization
of the free-running
period of adrenocortical
rhythm. 8) Carbachol, an acetylcholine agonist, shortens the free-running period of circadian activity
rhythm. 9) Melatonin accelerates the re-entrainment
of the circadian rhythm to shifted LD cycle. 10)
Light signals increase the acetylcholine concentration in suprachiasmatic nucleus, but not the other
control site. 11) The cAMP levels in rat suprachiasmatic
nucleus show an endogenous circadian
rhythm. 12) Long term cultured neuron from rat suprachiasmatic
nucleus retain the capacity for
circadian oscillation of vasopressin release. 13) Inhibitors of protein
synthesis phase shift the
circadian clock in chick pineal cells.
KEY WORDS: CIRCADIAN RHYTHM, OVULATION, PSEUDOPREGNANCY, PARTURITION, SUPRACHIASMATIC NUCLEUS, PINEAL
Jpn J Anim Reprod
37, 1P-11P, 1991
1991年度島村賞受賞講演論文
生殖機構 と生体 リズム機構の共役 に関する基礎的研究
村 上
昇
宮 崎 大 学 農 学 部 獣 医 学 科 家 畜 生 理 学 講 座 〒889-21宮 崎 市 学 園 木花 台西1-1は じめ に
あ ら ゆ る生 物 に は 自己 固有 の計 時機構(い わ ゆ る生 物 時 計)が 備 わ っ て い る.こ れ は 生物 が進 化 の 過 程 で 地 球 の 自転 や 公 転 に伴 う外 部 環 境 の周 期 的現 象 に 適 応 し,生 体 機 構 の 恒 常 性 を維 持 す る た め に獲 得 した 防御 機 構 の 一 つ と考 え ら れ る.時 計 の局 在 部 位 は動 物 種 に よ って異 な るが,軟 体 動 物 の 目,は 虫 類 や鳥 類 の松 果 体,お よび ほ 乳 類 で の 視 交 叉 上 核 が 挙 げ られ る.こ れ らの 時計 は通 常, 外 部 環 境 の 周 期 的 変 動 に 同調 して種 々 の生 理 機 能 の 日内 リズ ムや 年 内 リズ ム を駆 動 して い る.こ の 中 に は生 化 学 的,内 分 泌 学 的,神 経 学 的 そ して行 動 学 的 に様 々 な リズ ム が 認 め られ,そ の一 部 は生 殖 機 構 と強 く連 結 し,個 々 の 動 物 の 性 周 期 や繁 殖 周 期 な どを支 配 して い る.こ の た め,生 殖 機 構 を理 解 す る上 で,一 日を単 位 と した,ま た 一 年 を単 位 とす る時 間軸 を取 り入 れ た機構 解 析 が必 要 に な る.一 方 で は,生 物 時 計 の人 為 的操 作 で リズ ム を変 化 させ る こ と に よ り生 殖 機 構 な ど を調 節 で きる可 能性 が存 在 す る.本 研 究 は こ の様 な観 点 か らラ ッ トを用 い て そ の 生 殖 内 分 泌 機構 を,最 も基 礎 とな る概 日 リズ ム機構 の面 か ら解 析 す る と 同時 に計 時 機構 の基 礎 解 析 か らそ の応 用 へ と発 展 させ る こ と を 目 的 と して来 た もの で,こ れ まで に得 た著 者 らの 知 見 を紹 介 した い.ラ ッ トの 生 殖 機 構 と概 日 リズ ム機 構 との共 役
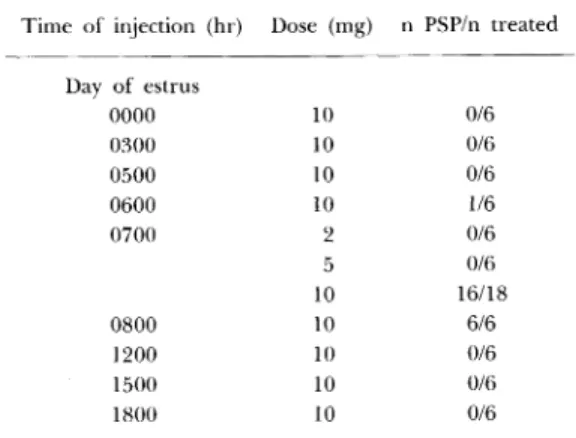
a)性 周 期 お よ び排 卵 機 構 と概 日リ ズ ム との共 役 通 常,24時 間 の 規 則 正 しい 明暗 交 代 条 件 下 で は ラ ッ ト は4日 また は5日 の 規 則 正 しい性 周 期 を繰 り返 す.こ れ は24時 間 周 期 の 明 暗 条 件 に 日内 リズ ムが 同調 して い る た め で,排 卵 前LHサ ー ジ とそ れ に続 く排 卵 は極 め て 限局 した 時 間 帯 に 起 こ る.し か し明 暗 交 代 の な い連 続 照 明 下 へ 移 す と,リ ズムの周期 は24時 間 よ り著 しく長 くな り, 行 動 な どの リズ ム は 不 明 瞭 に な る.こ の 様 な過 程 で,規 則 的 な性 周期 は 乱 れ,つ い に は連 続 発 情 と な る.そ の 結 果 自然 排 卵 は停 止 し,そ の か わ りに 兎 や 猫 に見 られ る交 尾 排 卵 機構 が 具 備 さ れ る.Fig.1は こ の よ うな 連 続 発 情 ラ ッ トの交 尾 刺 激 とそ れ に 伴 う排 卵 の 時 間 的 関 係 を示 し た もの で あ る.一 日の どの 時 刻 に 交 尾 刺 激 を加 えて も刺 み 激 す る 時刻 に 関係 な く刺 激14-16時 間 後 に一 様 に排 卵 す る こ とが判 明 した.さ らに,子 宮 頸 部刺 激 後 の 種 々 の 時 間 にLH-RH抗 血 清 の投 与 に よ る排 卵 阻止 実 験 を行 った 結 果,こ の交 尾 排 卵 は,刺 激後 か な り早 期 に神 経 反 射 的 にLH-RHの 放 出 を 招 い て 起 こ る も の と推 察 さ れ た (Table1).次 に,こ の よ う な 自然 排 卵 機 構 か ら交 尾 排Table 1. The incidence of ovulation after a single injection of
1 ml of anti-LHRI-1 or anti-BSA at various time after cervical stimulation in the light estrous rats
Fig. 1. Incidence of ovulation after cervical stimulation
in light-estrous rats. The stimulation was applied at arbitrary solar hr (0 hr) to a group of animals having
14 to 16 days of continuous vaginal estrus. Each circle
represents an animal tested.
Fig. 2. Effectiveness of cervical stimulation for induction of
ovulation in light estrous rats. Ovulated (open circle) and not ovulated rats (closed circle) were plotted according to their career prior to the test. Line A represents the time of establishment of light estrus, so that duration of continuous estrus of each rat is induced by a distance (b) between line A and each circle. The area enclosed by lines A, B and the abscissa indicates the condition where light estrous rat did not
卵 機 構 へ の 移 行 が どの よ う に して 起 こ る のか を調 べ た も の がFig.2で あ る.そ の 結 果,連 続 発 情 日数 が 少 な く て も長 期 間 連 続 照 明 下 にい た場 合 や,連 続 照 明 下 日数 が 少 な くて も連 続 発 情 日数 が 長 い場 合 に こ の交 尾 排 卵 機構 が 獲 得 さ れ る こ と が 判 明 し た(Murakami et al.,1978). 性 周 期 の 回 帰 は こ の 様 に 光 に 同 調 し た 計 時 機 構 と性 中 枢 な ど の 上 位 中 枢 の カ ッ プ リ ン グ に よ り起 こ る も の で あ る が,生 体 か ら 切 り放 し た 卵 巣 で も黄 体 の 下 垂 体 性 刺 激 ホ ル モ ン に 対 す る 性 周 期 依 存 性 変 化 が 存 在 す る (Murakami et al.,1982; Takahashi and Murakami
1982)こ とか ら,(光)一 時 計一 視 床 下 部 一 下 垂 体 一 卵 巣 軸 の ダ イナ ミ ック な統 制 機構 に よ り制 御 さ れ て い る と 言 え る. b)黄 体 相 と概 日リ ズ ム と の共 役 ラ ッ トは交 尾 刺 激 に よ り偽 妊 娠 あ る い は妊 娠 が 誘 起 さ れ2-3週 間 の 黄 体 相 が 成 立 す る.こ の 間黄 体 か ら旺 盛 なprogesteroneの 分 泌 が 起 こ るが,こ の分 泌 を刺 激 して い る もの は下 垂 体 か ら分 泌 され る プ ロ ラ クチ ン(PRL) で あ る.こ のPRLの 分 泌 パ タ ー ン は非 常 に 特 徴 的 で, 性 周 期 中 に は見 られ な い一 日2回 の サ ー ジ を形 成 す る. そ して こ の2回 のサ ー ジ いず れ もが毎 日の 明 暗 条件 に依 存 して い る.例 え ば,Fig.3に 異 な った 明 暗 条 件 下 で の PRLサ ー ジ を示 す.明 暗 条 件 が 異 な って も常 に暗 期 後 半 と明期 後 半 の2回,即 ち 明暗 条 件 に 同調 して 出現 して お り,こ のPRLサ ー ジ は 光 を同 調 因子 と した 概 日 リズ ム 機構 に 強 く共 役 して い る と言 え る(Murakami et al, 1986).ま た,こ の 黄 体 相 の 成 立(PRLサ ー ジ の 形 成) や 黄 体 相 の 維 持(PRLサ ー ジ の 維 持)機 構 に は 末 梢 血 中 のprogesteroneの 上 位 中 枢 へ の フ ィ ー ドバ ッ ク 情 報 が 密 接 に 関 係 し,そ の 情 報 伝 達 と 反 応 に も 概 日 リ ズ ム 機 構 と の 共 役 が 推 察 さ れ て い る(Murakami et al.,1979; Takahashi et al.,1980). ラ ッ ト黄 体 相 の 成 立 は 交 尾 刺 激 の み で な く発 情 期 に 大 量 のprogesteroneの 単 一 投 与 で も起こ す こ と が 出 来 る. Table2は そ の 効 果 と 時 間 の 関 係 を 示 す が,5時 点 灯19 時 消 灯 の 明 暗 条 件 下 飼 育 で はprogesteroneを 発 情 期 の 午 前7-8時 に 投 与 す る と 最 も 効 率 良 く偽 妊 娠 が 誘 起 で き る(Murakamietal.,1980).こ のprogesterone投 与 に よ る偽 妊 娠 成 立 はprogesterone投 与 直 後 の 一 過 的 な PRLの 分 泌 とそ の 後 形 成 さ れ るPRLサ ー ジ に よ る もの と 推 測 され る(Murakami et al.,1980).す な わ ち,以 上 の結 果 は ラ ッ ト黄 体 相 の成 立 機構 お よ び維 持 機 構 に 時 間軸 を基 礎 と した上 位 中枢 の概 日 リズ ム機 構 が 関与 して い る こ と を意 味 して い る.
Fig. 3. Effect of photo period on the
secre-tion of prolactin levels in the pseudopreg-nant rats. Samples were obtained by de-capitation (A) or tail tip incision method (B, C). Lighting regimen is 14L: 10D (A, B: lights on 0500-1900, C: lights on 1700-0700)
Table 2. Effect of a single injection of progesterone on the
c)分 娩 と概 日 リズ ムの 共 役 Fig.4はSprague-Dawley(SD)ラ ッ ト及 びWistar ラ ッ トの 分 娩 時 刻 と明 暗 条 件 の 関係 を調 べ た もの で あ る, SDラ ッ トの 分 娩 は5時 点 灯19時 消 灯 の 明 暗 条 件 下 で は 妊 娠22日(精 子 確 認 を1日 と して)の 明期 に起 こ っ た. 妊 娠6日 以 後 に明 暗 条 件 を変 化 した状 態 に置 く と,分 娩 時 刻 は移 動 し,新 た な明 期 に起 こ っ た.す な わ ち,分 娩 が 単 に受 精 卵 の 着 床 か ら の 日数 に よ っ て決 ま る の で は な く光 条 件 を 同調 因子 とす る概 日 リズ ム機 構 の支 配 下 に も あ る こ とが 判 る(Murakami et al.,1987).こ の こ とは 恒 常 暗 下 に置 い た時,日 内 リズ ム が 自由継 続 し,個 々 の ラ ッ トの 周 期 の違 い か ら分 娩 時刻 に 違 い が 出 る こ と,ま たFig.5に 示 す 様 に 生 物 時 計 の 局 在 部 位 で あ る 視 交 叉 上 核 を破 壊 す る と分 娩 時刻 が 明期 の み で な く暗期 に も起 こ る こ と か ら も支 持 さ れ る(Murakami et al.,1987). 以 上 の様 に ラ ッ トの排 卵 機 構 か ら分 娩機 構 に 至 る まで これ らが 外 部 環 境,特 に明 暗 条 件 を 同調 因子 と した リズ ム 機 構 に密 接 に カ ップ リ ン グ して い る こ とは 明 らか で あ る.光 条 件 に よ る 自然 排 卵 か ら交 尾 排 卵 へ の変 化 や分 娩 時 刻 を操 作 で きる こ とは見 方 を変 え る と,光 の条 件 と言 う リズ ム の 同調 因子 を変 化 させ る こ とに よ り,リ ズ ム機 構 に連 結 した繁 殖 機構 を操 作 出 来 る 可 能性 を示 した こ と に な る.す な わ ち,繁 殖 の機構 解 明 や応 用 に お い て リズ ム機 構 の 解 析 や応 用 が 一 つ の ア プ ロ ー チ とな る わ け で あ る.そ こ で以 下 に,リ ズ ム機 構 を操 作 す る幾 つ か の 方法 を紹 介 す る.
生体時計の操作
a)同 調 因子 に よる操 作 本 来,個 々 の 動 物 が 有 す る生 物 時 計 の リズ ムの 周 期 は 24時 間 よ り多 少 ず れ た 各 自固 有 の 周 期 を持 って い る.そ の た め外 部 環 境 に 周 期 的 因 子 が 存 在 しな けれ ば,位 相 は 毎 日ず れ て 行 き 自由 継 続 リズ ム を示 す.し か し明 暗 な ど の24時 間 周 期 の 因 子 が 作 用 す る と時 計 は その 周 期 にセ ッ ト(同 調)さ れ る.こ の 様 な 因 子(同 調 因子)に は光, 温 度 や 音 な どが あ げ られ る.著 者 らは,制 限 給 餌 が 光 より 強 力 な 同 調 因 子 で あ る こ と(Murakami and
Taka-Fig. 4. Incidence of time of parturition in Sprague-Dawley (a, b, c) and Wistar (d) rats kept in a lighting regimen of 14L: 10D; lights on 0500-1900 (a, d) or 1700-0700 (b). In (c), rats were kept in constant darkness. Half circle and complete
circle symbols represent the time of parturition in 1 and 2 rats, respectively. Time of birth was determined at 3-hr intervals.
Fig. 5. Time of parturition in rats with a complete
lesion of suprachiasmatic nucleus (closed circle), a partial lesion of SCN or intact (open circle) (B). The lesion of suprachiasmatic nucleus was performed on
day 13 of pregnancy, and circadian rhythm of water intake was investigated on day 19 of pregnancy to determine whether the rhythm was abolished by
hashi,1982;Takahashi and Murakami,1983; Murakami et aL,1986),母 親(Takahashi et al.,1982,1984;Sasaki et aL,1984)や 群 飼 育 に よ る 社 会 的 因 子(Takahashi and Murakami,1982,1983)も 同 調 因 子 に な り得 る こ と を 見 い だ し た.Fig.6は 明 暗 交 代 条 件 下 で 制 限 給 餌 を 行 っ た 時 の 副 腎皮 質 ホ ル モ ン リズ ム を示 した もの で あ る。 自由 給 餌 の場 合,暗 期 開 始 直前 に ピー クが あ るが,毎 日2時 間 の 制 限給 餌 を明期 に行 う と,2週 間 後 に は給 餌 直前 に ピー ク を形 成 して い る.即 ち,餌 の 来 る時刻 を予 知 して い る こ とに な り,こ の ホ ル モ ン リズ ム が 計 時 機 構 に よ り 制御 され て い る と言 え る.こ の 制 限 給 餌 に よ る副 腎 皮 質 ホ ル モ ンの 同 調 性 を利 用 し,分 娩 時 刻 を意 図 す る時 刻 に 持 っ て くる こ と も一部 の 動 物 で可 能 で あ る(Murakami θ∫謡,未 発 表).Fig.7は ラ ッ ト乳 児 の リズ ム が 母 親 の リ ズ ム に 同調 さ れ る こ と を示 す 結 果 で あ る.生 後 直 ち に眼 球 を摘 出 し,半 分 の 子 供 は産 み の 親 に半 分 の 子 供 は産 み の親 と逆 転 した リ ズ ム を持 つ 雌 ラ ッ トに保 育 させ る と, 子 供 はす べ て 保 育 さ れ た母 親 の リズ ム を受 け継 いで 離 乳 後 も継 続 した.こ の こ とは,育 母 が保 育 を通 して 子供 の リズ ム をセ ッ トす る こ と を示 して い る.Fig.8は 盲 目 ラ ッ トを個 別 ケ ー ジ で 一 匹ず つ飼 育 した時 と,同 じケ ー ジ 内 に6匹 を ま とめ て群 飼 育 した と きの成 長 過 程 で の 自由 継 続 リズ ム の頂 点位 相 の ず れ を示 した もの で あ る.個 別 ケ ー ジ の ラ ッ トで は ラ ッ ト間 の 固 々 の リズ ム の 周期 の ず れ が は っ き り分 か る が,群 飼 育 に よ り,お 互 い の 周 期 が
Fig. 6. Effect of restriction of feeding time on the
circadian adrenocortical rhythm. The rats were kept under 12L: 12D (lights on 0600-1800). The time of food presentation was restricted between 08 hr and 10 hr (middle), and between 08 hr and 16 hr (right) without change of lighting condition. In rats shown in left panel, food were given ad libitum.
Fig. 7. Two days pattern of blood corticoster-one levels in blinded pups raised by original or foster mothers. Two left panels show the blood corticosterone rhythms in blinded pups born under LD or DL and raised by original
mother. Two right panels show the blood corticosterone level in blinded pups born under DL (LD) and raised by foster mother which was kept under LD (DL), respectively.
Fig. 8. Free-running patterns of the circadian
adrenocor-tical rhythm in blinded rats housed either singly (right) or group (left). Circadian peak time is shown on the ordinate and the number of weeks after blinding is shown on the
abscissa. Each circle represents the acrophase of the rhythm in each animal at the examination week.
同調 して行 っ た こ とに よ りば らつ きが 少 な くな って い る. す な わ ち,社 会 的 な 因子 が お互 い の リズ ム の 周期 を同調 した と解 釈 さ れ る. b)薬 物 に よ る操 作 著 者 ら は 自 作 の 行 動 計 を 使 用 し て(Yokawa et al., 1986)種 々 の 薬 物 の 行 動 リ ズ ム に 及 ぼ す 効 果 を 調 べ た が, ア セ チ ル コ リ ン 系 薬 物(Murakami et al.,1986,1987; Furukawa et al.,1987)と メ ラ トニ ン(Murakami et al.,
1983)に 最 も効 果 の あ る こ と を見 い だ した.Fig.9は ア セ チ ル コ リ ンの作 動 薬 で あ る カ ル バ コー ル を 自由 継 続 リ ズ ム を示 す ラ ッ トの視 交 叉 上核 の 近傍 に投 与 した 時 の リ ズ ム の周 期 に及 ぼ す効 果 を示 して い る.い ず れ も周 期 が 投 与 前 に比 べ て著 し く短縮 した.一 方,メ ラ トニ ンは急 な 光 条件 へ の変 化 に対 し リズ ム を速 や か に 再 同調 させ た. こ の再 同調 促 進 効 果 は 行動 リズ ム の み で な く副 腎 皮 質 ホ ル モ ン リズ ム で も見 られ た.そ の結 果 を ま とめ てTable 3に 示 した.明 暗 条 件 逆転 後4日 で メ ラ トニ ンの再 同調 促 進 効 果 が顕 著 な こ とが わ か る.こ の メ ラ トニ ンの再 同 調 促 進 効 果 を利 用 した 東 西飛 行 で,時 差 ボ ケ の解 消 効 果 が す で に 臨床 的 に認 め られ て い る. 以 上 は,同 調 因子 や 薬物 を利 用 して リズ ム の位 相 や周 期 を 変化 させ る例 を示 した が,こ れ らは 単 に リズ ム学 で は な く生 物 時 計 が基 礎 とな る種 々 の生 理機 構 を間接 的 に 制御 す る分 野 に大 い に 貢 献 で きる と考 え られ る.メ ラ ト ニ ンや制 限給 餌 の応 用 が す で に繁 殖 機構 や 時差 ボ ケ解 消 へ の 一端 に応 用 され て い る .こ う して 見 る と,時 計 機 構 の 本 質 の解 明 が さ ら に重 要 な 課 題 とな って 来 る.
生物時計機構の解析
a)1n vivoで の 解 析 ほ 乳動 物 の 生 物 時 計 は視 床 下 部 に 存 在 す る約8千 の神 経 細 胞 集 団 か らな る視 交 叉 上 核 に存 在 す る.そ の 時 計機 構 を解析 す る 上 で 一 つ の 方 法 は 時 計 の 構 成 要 素 を見 つ け る こ とで あ る.視 交 叉 上 核 中 の 幾 らか の 物 質 を検 索 した 結 果,ホ ル モ ン等 の 情 報 伝 達 物 質 で あ る サ イ ク リ ッ ク AMP(cAMP)の 濃 度 に 日 内 変 動 が あ り,光 に 同 調 す る と 同 時 に 光 情 報 が な く て も時 計 自 身 の リズ ム を 反 映 す る こ と を 見 い だ し た(Murakami et al.,1983).Fig,10 は そ の 結 果 を示 す が,明 期 後 半 に ピ ー クが あ り,明 暗 条 件 を逆 転 した 条 件 下 で もや は りピ ー ク は 明期 後 半 に現 れ た.ま た 盲 目 に し た り恒 常 明 下(LL)で は ピ ー ク の 移 動 が み られ,こ の リズ ムが 内 因 性 の もの で あ る こ とが 分 か る.こ の リズ ムが 視 交 叉 上 核 特 有 の もので あ る こ と は Fig.11に 示 す 様 に腹 内 側 核 な ど のcAMPの 日内 リズ ムTable 3. The incidence of reentrainment of circadian adrenocortical rhythm after reverse of LD cycle
Fig. 9. Free running rhythm of locomotor activity in
rats implanted with paraffin pellet (A), or pellet containing carbachol (B, C). Arrows indicate the day of implantation. Locomotor activity was recorded
が,視 交 叉 上 核 の リズ ム と全 く逆 の位 相 を持 って い る こ と か ら推 察 され る.そ の 後,こ のcAMPの リ ズ ム は 単 に時 計 か らの 出力 に す ぎな い の で は?と の 議 論 もあ った が,最 近,視 交 叉 上 核 の 活 動 電 位 の リズ ムがcAMPに よ っ て 転 移 す る こ と に よ り,cAMPが 時 計 機 構 の構 成 要 素 の 一 つ で あ る と考 え られ て い る. 次 に ア セ チ ル コ リ ンが 光情 報伝 達 に関 与 して い る こ と を 見 い だ し た(Murakami et al,1984).Fig.12は 光 照 射 後 に視 交 叉 上核 中 の ア セ チ ル コ リ ン含 量 が 増 加 す る こ と を示 した もの で あ る.こ の 場 合 も他 の 神 経 核 で は変 化 が な い こ とか ら視 交 叉 上核 中 の アセ チ ル コ リ ン と光 情 報 に密 接 な 関係 が あ る こ とが 分 か る.さ ら に,上 述 した よ う に,ア セ チ ル コ リ ン作 動 薬 で あ る カル バ コ ー ル の投 与 は リズ ム の周 期 を短 縮 した.併 せ て 考 えれ ば,ア セ チ ル コ リ ン系 が視 交 叉 上核 の 光 情 報 の 伝 達 や 発 信 に関 与 して い る可 能性 は 高 い. 通 常,時 計 機 構 の 解 析 は時 計 か ら発 せ ら れ た リズ ム を 末 端 の行 動 や ホ ル モ ンの リズ ム を指 標 に行 わ れ て い る. しか し末端 で の 発 現 リズ ム に は多 くの 修 飾 が 加 わ っ て い
Fig. 10. Twenty four hour pattern of cAMP
con-centrations of suprachiasmatic nucleus under va-rious lighting conditions. Intact rats were kept under LD (A), DL (B), LL (D) for 4, 2 or 1 weeks,
respectively. C: rats were blinded and kept under LD for 5 weeks. Each circle and vertical bar represents the mean ±S.E.M. of 6 rats. Asterisks
indicate the significant difference between the time of day (*P•ƒ0.01, **P•ƒ0.05)
Fig. 11. Twenty four hour pattern of cAMP concen-trations of ventromedial hypothalamus under va-rious lighting conditions. A, B, C, D of the figure correspond to those of Fig 10.
Fig. 12. Effect of light on the acetylcholine
concen-tration of the suprachiasmatic nucleus. The light was given 3 hr after darkness in intact (open circle) and blinded (black circle) rats. Acetylcholine
con-centration increased significantly 30 and 60 min after light on.
る こ とを考 慮 しな け れ ば な らな い.例 え ば,副 腎皮 質 ホ ル モ ンの リズ ム の発 現 は視 床 下 部 や 下 垂体 の ホ ル モ ンに よ り支 配 され て い る こ と以 外 に,甲 状 腺 ホ ル モ ンや そ の
他 の 物 質 が 複 雑 に 関 与 し て い る(Murakami and Taka-hashi,1983; Murakami et al., 1984; Murakami, 1984).
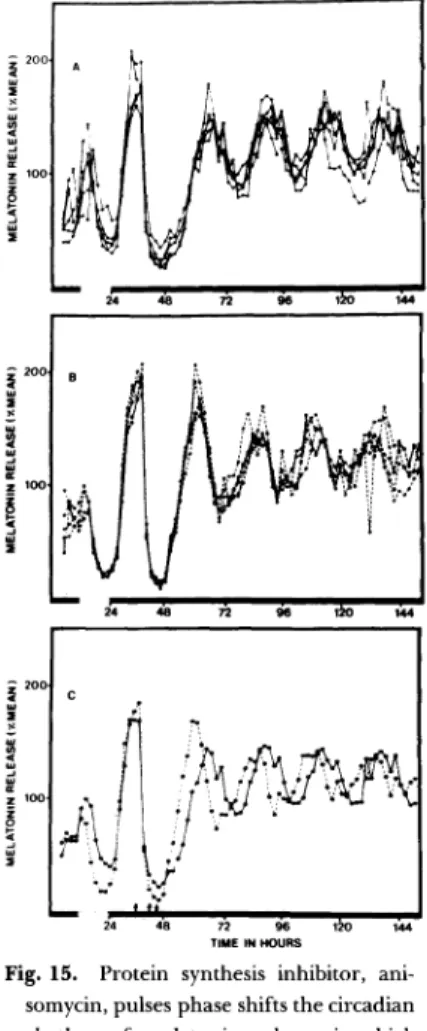
最 近,著 者 らは 光 を急 に変 化 した 時,時 計 自身 の 新 た な 光 へ の 再 同 調 は 速 や か に起 こ るが,行 動 リズ ム な ど末 端 の リズ ム の再 同調 に は か な りの時 間 を要 す る こ と を見 い だ した(Takamureβ 彦鳳,1991),時 計 機 構 解 析 を 末 端 の リズ ム を指 標 に す る場 合 注 意 を要 す る こ と を意 味 して い る.そ こ で,次 に時 計 の存 在 す る細 胞 で機 構 を解 析 す る試 み を検 討 した. b)In vitroで の 解 析 著 者 らは ほ乳 動 物 の視 交 叉 上核 の 時 計発 信 機 構 を細 胞 レベ ル で解 明 す る た め に,そ の神 経 細 胞 の培 養 を行 い, そ の 時計 自体 の リズ ム をバ ゾ プ レ ッ シ ン放 出 リズ ム で 捉 え る こ と に 成 功 し た(Murakami et aL,1991),Fig,13 は 視 交 叉 上核 神 経 細 胞 の 培 養 写 真 で あ る.神 経細 胞 は ラ ミニ ンで 被 覆 した 培 養 皿 上 で,無 血 清 培 養 され る と,神 経 回 路 網 状 の構 造 を呈 し,そ こか ら分 泌 され た バ ゾ プ レ ッ シ ン はFig.14に 示 す 様 に,概 日 リズ ム を示 した.こ の 培 養 系 で,今 後 細胞 レベ ル で の 時 計 機構 解 析 が 進 む こ とが 期待 され る. 始 め に記 した 様 に,ほ 乳 動 物 の 生 物 時 計 が 視 交 叉 上 核 に局 在 す るの に 対 し鳥 類 で は一 部 の 例 外 を除 い て 松 果 体 に存 在 す る と考 え られ て い る.さ ら に鳥 類 松 果 体 細 胞 は 光 受 容 体(photoreceptor)を 持 ち,光 情 報 は 細 胞 中 の 時 計 発 信 機 構 へ 伝 達 され そ こか ら発 す る リズ ム は メ ラ ト ニ ン分 泌 と して 現 れ る(Takahashi et al,1989),即 ち, 計 時 機 構 を解 析 す る上 で 一 つ の細 胞 内 に,光 入 力 機 構一 時 計 機 構一 出力 機 構 の す べ て を備 え た好 個 な材 料 と な る. そ こで まず,時 計 の発 信 や光 入 力 機 構 に蛋 白質 の 合 成 が 関 与 して い る か 否 か を検 討 した.Fig.15は ひ よ この 松 果 体 細 胞 の メ ラ トニ ン 分 泌 リ ズ ム を示 す(Murakami 1990,1991).暗 期 に ピ ー ク を持 ち,以 後 恒 常 暗(DD) 下 で も自 由継 続 リズ ムが 示 さ れ る.こ の状 態 で蛋 白合 成 阻害 薬anisomycinを パ ル ス 投 与 す る と リ ズ ム の 後 退 が 起 こ っ た.こ の 蛋 白合 成 阻害 薬 の効 果 は リズ ムの 位 相 に 特 異 的で,あ る時 間の 投 与 は リズ ム の前 進 を起 こ し,ま た あ る特 定 時 間 の投 与 は リ ズ ムの 後 退 を起 こ した.Fig, 16は そ の結 果 を ま とめ た位 相 反 応 曲 線 で あ る.こ の結 果 は 時計 の発 信 機 構 に新 た な蛋 白質 の合 成 が 必 要 で あ る こ とを意 味 して い る.さ ら に興 味 深 い こ と に,光 パ ル ス の効 果 と逆 転 した 関係 に あ り,光 に よ る リズ ム機 構 へ の
Fig. 13. Photomicrographs of cultured neurons pre-pared from SCN regions in 1-day-old rats. Cells were plated in 96 well dishes pre-coated with laminin. This picture was taken on day 11 of culture.
Fig. 14. Two individual samples showing circadian oscillation of vasopressin release from cultured SCN neurons. Perfusion was started after 21 days in culture. Flow rate was 0.1 ml/hr and samples were collected at 3 hr intervals.
効 果 が あ る蛋 白 を合 成 した結 果 で あ る こ とを暗 示 して い る.こ の可 能 性 は,光 に よ る リズ ム の移 動 が 光 と同 時 に 蛋 白 合 成 阻 害 薬 の 投 与 に よ り阻 止 さ れ た(Fig.17)こ とか ら証 明 され る.35S-メ チ オニ ンの蛋 白合 成 へ の取 り込 み を調 べ る と,光 照 射 に よ り,76Kダ ル ト ン と80K ダ ル ト ン蛋 白合 成 が 促 進 さ れ た こ とが 判 明 した. 以 上,概 日 リズ ム を 中心 に,そ の基 礎 か ら応 用,ま た 繁 殖 機 構 との 関係 に つ い て 著 者 らの研 究 を紹 介 した.繁 殖 機 構 も計 時機構 も共 に,進 化 の 過 程 で 生 命,種 の 維 持 に不 可 欠 な もの で あ る.最 近 で は分 子 生 物 学 的 技 術 の進 歩 と相 ま って 多 くの 問 題 が 解 明 され て き たが,未 解 明 な 部 分 も まだ まだ 多 い.今 回 の 研 究 の 一 部 で もが 将 来 の発 展 の役 に立 て れ ば幸 い で あ る.
謝
辞
本 研 究 に対 し島 村 賞 の 栄 誉 を与 えて 下 さ っ た 日本 家 畜 繁 殖 学 会 に 深 く感 謝 申 し上 げ ます.ま た推 薦 の労 を取 っ て下 さ った 江 藤 禎 一,高 橋 迫 雄 教 授 に深 謝 致 します.本 研 究 は東 京 大 学 家 畜 生 理 学 講 座,東 京 神 経 科 学 総 合 研 究 所,及 び宮 崎 大 学 の 多 くの 先 生 方 や卒 業 生 の御 協 力 と御Fig. 15. Protein synthesis inhibitor, ani-somycin, pulses phase shifts the circadian rhythm of melatonin release in chick
pineal cells. A: individual record of 4 cultures exposed to a 6-hr anisomycin beginning at CT 3. B: individual control culture. C ensemble averages of the 2
groups shown in A and B.
Fig. 16. Comparison of the phase response curves for 6 hour pulses of anisomycin, 6 hour pulses of light, 4 hr pulses of light or dark.
Fig. 17. Effects of anisomycin on the light-induced phase
shift in chick pineal cells. A: Six hr pulses of anisomycin together with light were started at CT18 (L: light pulse only, A: anisomycin pulse only, L+A: light and anisomy pulses). Anisomycin blocked the phase advance effect induced by light. B: Six hr pulse of anisomycin was started 3 hr before light pulses. Anisomycin was given between CT 15 and CT21. Light pulse was given between CT18 and CT21. L: light only, A: anisomycin only, A-L: light pulses followed on aisomycin. Dimetylanisomycin
援 助 の 上 にで き あが っ た もので,深 甚 の謝 意 を 表 し ます. 特 に,研 究 へ の 興 味 を与 え,最 初 に こ の道 へ の 紹介 を下 さ った 元 宮 崎 大 広 江 一 正 教 授,研 究 へ の 強烈 な情 熱 とイ ンパ ク トと を与 えて 下 さ っ た東 大 の高 橋 迫 雄 教 授,鈴 木 善祐,本 間 運 隆 両 東 大 名 誉 教 授,生 体 リズ ム研 究 の御 指 導 を頂 い た 国 立 神 経 セ ンタ ー の高 橋 清 久 部 長 そ して現 在, 御 指 導 並 び に共 同研 究 を して頂 い て い る宮 崎 大 学 の江 藤 禎 一教 授 に この 場 を借 りて,深 謝 の意 を表 しま す.最 後 に,勇 気 と支 持 を与 え続 け て くれ る妻 の和 代 と家 族 に感 謝 し ます 。
文
献
Furukawa T, MurakamiN, Takahashi K, Etoh,T(1987)
Effect of implantation of carbachol pellet near the
suprachiasmatic nucleus on the free-running period of
rat locomotor activity rhythm. Jpn J Physi 37: 321-326.
Murakami N, Takahashi M, Suzuki Y (1978) Conditions forestablishment of reflex ovulation in light estrous rats.
Endocrinol Jpn 25 (3) : 299-303.
Murakami N, Takahashi M, Suzuki Y (1979) Indispensable
role of peripheral progesterone level for the occurr-ence of prolactin surges in pseudopregnant rats. Biol
Reprod 21: 263-268.
Murakami N, Takahashi M, Suzuki Y (1980) Induction of
pseudopregnancy and prolactin surges by a single
injection of progesterone. Biol Reprod 22: 253-258.
Murakami N, Takahashi M, Suzuki Y, Homma K (1982)Responsiveness of dispersed rat luteal cells to luteiniz-ing hormone and prolactin during the estrous cycle and early pseudopregnancy. Endocrinology 111: 500-508.
Murakami N, Takahashi, K (1982) Circadian
adrenocortic-al rhythm in the adrenalectomized rat with or without autotransplanted adrenal gland. Neuroendocrinology 34:
157-162.
Murakami N, Hayafuzi C, Sasaki Y, Yamazaki J, Takahashi K (1983) Melatonin accerelate the reentrainment of circadian adrenocortical rhythm in inverted
illumina-tion cycle. Neuroendocrinology 36: 385-391.
Murakami N, Takahashi K (1983) Enhancement of
corti-costerone release by repeated injections of ACTH in
the dexamethasone pretreated rat. Acta Endocrinol
(Copenhagen) 102: 583-588.Murakami N, Takahashi K (1983) Circadian rhythm of
297-304.
adenosine 3', 5'-monophosphate content in sup-rachiasmatic nucleus (SCN) and ventromedial hypothalamus (VMH) in the rat. Brain Res 276:
Murakami N, Kawashima K, Takahashi K (1984) Effect of
light on the acetylcholine concentration of sup-rachiasmatic nucleus in the rat. Brain Res 311: 358-360.
Murakami N, Takahashi K (1984) Thyroid hormone
maintains circadian rhythm of blood corticosterone levels in the rat: by restoring the synthesis and release of ACTH after thyroidectomy. Acta Endocrinol
(Copenhagen) 107: 519-524.
Murakami N (1984) Factors other than ACTH for
man-ifestation of circadian rhythm in blood corticosterone levels in the rat. Ibn I Vet Sci 46: 437-442.
Murakami N, Kuroda H, Etoh T (1986) Restricted feeding
regimen affects the diurnal prolactin surge but not
nocturnal surges in pseudopregnant rat.Jpn J Vet Sci
48: 29-34.Murakami N, Furukawa T, Etoh T, Takahashi K (1986)
Effect of continuous supply of light or carbachol to the
SCN on the free-runnig rhythm. Jpn J Physiol 36:
411-416.Murakami N, Abe T, Yokoyama M, Katume J, Kuroda H,
Etoh T (1987) Effect of photoperiod, Injection of
pentobarbiton sodium or lesion of suprachiasmatic
nucleus on pre-partum decrease of blood progesterone
concentrations or time of birth in the rat.] Reprod Ferti
79: 325-333.Murakami N, Furukawa T, Etoh T, Takahashi K (1987) Effect of cholinergic agonist and continuous light on the period of free-running rhythm of rats. In: Compa-rative Aspect of Circadian Rhythm (Hiroshige T, Honma K eds.), Hokkaido Univ. Press, Sapporo. pp
135-144.
Murakami N (1990) Involvement of protein synthesis in the
circadian clock and light entrainment in chick pineal
cells. Jpn J Anim Reprod 36: 53-61 (in Japanese) .
Murakami N (1991) Existence of clock related protein inchick pineal cells. Horumon to Rinsyo 39 (6) : 11-16 (in
Japanese) .
Murakami N, Takamure M, Takahashi K, Utunomiya K,
Kuroda H, Etoh T (1991) Long-term cultured neurons from rat suprachiasmatic nucleus retain the capacity for circadian oscillation of vasopressin release. Brain
Res 545: 347-350.
Sasaki Y, Murakami N, Takahashi K (1984) Critical period
for the entrainment of the circadian rhythm in blinded pups by dam. Physiol Behav 33: 105-109.
Takahashi JS, Murakami N, Nikaido SS, Pratt BL,
Robert-son LM (1989) The avian pineal, a vertebrate model system of the circadian oscillator: Cellular regulation of circadian rhythm by light, secondary messengers and macromolecular synthesis. Rec Prog Hor Res 45:279-348.
Takahashi K, Hayafuji C, Murakami N (1982) Foster
mother rat entrains circadian adrenocortical rhythm inblinded pups. Am J Physiol 243: E443-449.
Takahashi K, Murakami N, Hayafuji C, Takahashi Y (1982)
Entrainment of the circadian adrenocortical rhythm in rats. In: Biological Rhythms and Biological Clock (Nakagawa H ed.) , Kyouritu Press, Tokyo. pp
213-226 (in Japanese) .
Takahashi K, Murakami N (1982) Entraining agents for the
circadian adrenocortical rhythm in rats. In: Vertebrate
Circadian System (Ashoff J, Daan S, Groo G eds.) ,
Takahashi K, Murakami N, Hayafuji C (1984) Further
Springer-Verlag Berlin-Heiderberg pp 194-202.evidence that circadian rhythm of blinded rat pups is
entrained by the nursing dam. Am J Physiol 246:
359-363.
Takahashi K, Murakami N (1990) Circadian Rhythms. In: Review of Neuroscience (Itoh M, Narahashi H eds.) ,
Igakusyoin, Tokyo. pp 175-195 (in Japanese) .
Takahashi K, Okawa M, Murakami N (1990) Entrainingmechanism of the clock in mammals from the clinical
viewpoint. Adv in Neurol Sci 34: 892-900 (in Japanese) .
Takahashi K, Inoue ST, Honma K, Murakami N (1991)Discussion for Cliobiology. Clinical Neurosci 9: 82-94
(in Japanese) .
Takahashi M, Murakami N, Naito H, Suzuki Y (1980) Blockade of pseudopregnancy in the rat by treatment with anti-progesterone serum. Biol Reprod 22: 423-429. Takahashi M, Murakami N, Naito, H, Suziki Y (1981)
Central regulation in occurrence of luteal phase in rats.
Folia Endoc 57: 131-136 (in Japanese) .
Takamure M, Murakami N, Takahashi K, Kuroda H, Etoh
T (1991) Reentrainment of the circadian clock itself, but not measurable rhythms to a new light-dark cycle in the rat. Physiol Behav 50 (2) : 443-449.
Yokawa T, Murakami N, Furukawa T, Etoh T (1986) Effect of constant light on the circadian rhythm in locomotor activity in rats. Exp Anim 35: 185-188.