小脳・マシンインタフェースによる単一 Purkinje 細胞活動と 運動学習の因果関係直接評価法
片桐 和真
†田中 良幸
††平田 豊
†,††a)Cerebellum-Machine Interface to Evaluate Direct Causality between Cerebellar Purkinje Cell Single Unit Activity and Motor Learning
Kazuma KATAGIRI
†, Yoshiyuki TANAKA
††, and Yutaka HIRATA
†,††a)あらまし 身体運動の制御と学習の中核は小脳が担っている.小脳皮質神経回路を構成する細胞タイプのう
ち,Purkinje細胞は唯一小脳外へ軸索を伸ばす出力細胞である.これまでの研究により,Purkinje細胞の発火
頻度は,筋系を駆動するための運動指令をコードしているものと考えられている.したがって,運動の学習は
Purkinje細胞発火頻度パターンが変化することにより実現されるものと考えられている.こうした知見は,主に
動物を用いた行動実験とPurkinje細胞単一神経電位計測により得られたものである.しかし,このような実験 では,神経細胞活動変化と行動変化の間の相関関係を評価できるのみであり,単一の神経細胞活動が運動制御や 運動学習に果たす役割を直接評価することは困難である.本研究では,こうした従来のシステム神経生理学的手 法の限界を超え,小脳神経活動と運動学習の間の直接的因果関係を評価するためのスキームを提案する.提案ス キームは,前庭動眼反射(VOR)の運動学習を例に,金魚の眼球運動制御を担う前庭小脳Purkinje細胞の単一 神経活動計測を行い,ブレイン・マシンインタフェース(BMI)を介して直流モータを制御するものである.こ の時,直流モータの制御誤差をVOR運動学習を駆動する網膜像のスリップ信号として金魚に与える.この系に より,モータ制御の精度向上は,記録中のPurkinje細胞発火パターン変化に直接関連づけられる.この系を構 築し,動物実験を実施した結果,VOR運動学習にかかわると仮定されてきた記録中の単一Purkinje細胞の発 火パターンが徐々に変化し,直流モータの運動誤差が適応的に減少する事が示された.この結果は,単一の小脳
Purkinje細胞発火パターンの変化と運動学習の間の直接的因果関係を初めて示したものである.
キーワード 運動学習,前庭動眼反射,金魚,小脳,ブレイン・マシンインタフェース
1.
ま え が き小脳は身体運動の制御と学習の中核を担う.小脳 内情報処理メカニズムの理解は,
40
年以上に亘り神 経科学分野における中心的課題と捉えられてきた.1960
年代には,その神経回路構成が解剖学的に同定さ れ[1]
,小脳皮質神経回路は主に顆粒,Golgi
,籠,星 状,Purkinje
の各細胞により構成されることや,小脳 皮質神経回路は,これらの細胞により形成されるマイ†中部大学工学部,春日井市
College of Engineering, Chubu University, 1200 Matsumoto- cho, Kasugai-shi, 487–8501 Japan
††中部大学情報科学研究所,春日井市
Research Institute for Information Science, Chubu Univer- sity, 1200 Matsumoto-cho, Kasugai-shi, 487–8501 Japan a) E-mail: [email protected]
クロゾーンと呼ばれる基本神経回路の繰り返しによる 一様構造をもつこと等が明らかにされている
[2]
.ま た,こうした神経回路構造は魚類から霊長類まで保存 されていることから,小脳による運動制御と学習の基 本原理は,運動の種類や動物種を超えて共通と考えら れている[3]
.Purkinje
細胞は小脳皮質神経回路を構成する細胞 の中で,唯一小脳外へ軸索を伸ばす出力細胞である.これまでの研究により,例えば眼球運動にかかわる小 脳腹側傍片葉
Purkinje
細胞の発火頻度は,眼球筋肉 系を駆動するための運動指令をコードしていることが 示されている[4]
.こうした知見から,運動の学習はPurkinje
細胞発火頻度が変化することにより実現され るものと考えられている.またその基礎過程は,運動 誤差を減少させるように生じる,平行線維とPurkinje
細胞間シナプス伝達効率の可塑的変化であると考えら れている
[2]
.すなわち,Purkinje
細胞へ運動の誤差 情報を運ぶ登上線維入力と,感覚・運動情報を運ぶ平 行線維入力がほぼ同時に高まると平行線維–Purkinje
細胞間シナプスに長期減弱(LTD
)が生じ[5], [6]
,平 行線維入力のみが高まると,このシナプスに長期増 強(LTP
)が起こることが動物実験により示されてい る[7], [8]
.運動学習中の単一Purkinje
細胞活動計測 実験では,実際に運動学習の前後で,Purkinje
細胞集 団の平均的活動が,学習を促す方向に変化することが 示されている[9]
〜[11]
.こうした知見は,運動学習中の被験動物小脳単一
Purkinje
細胞から細胞外電位を計測し,学習前・中・後の細胞発火頻度変化を評価することにより得られ たものである.しかしながら,このようなシステム神 経科学における常套的実験手法では,神経細胞活動変 化と行動変化の間の相関関係を評価できるのみであ り,各細胞が運動学習による行動変化に果たした寄与 について,直接的な因果関係に言及することは不可能 である
[12]
.例えば,実験時に電極刺入等により計測 中の細胞が壊れた場合にも,行動に有意な変化が現れ ることはなく,このような破壊実験によっても単一の 神経細胞が運動制御や運動学習に果たす役割を直接 評価することは困難である.一方,運動学習中の単一Purkinje
細胞活動連続記録では,個々の細胞の発火頻 度が必ずしも学習を促すように変化するわけではなく,学習と無関係あるいは阻害する方向に変化する場合も あることが示されている
[10], [11]
.すなわち,個々のPurkinje
細胞においては,平行線維–Purkinje
細胞間 シナプスが運動誤差を減少させるように変化するとい う,上述の小脳における基本学習理論が一見当てはま らない場合がある.ただし,こうした現象は,各細胞 活動と運動出力が必ずしも対応していないために生じ ている可能性がある.細胞の中にはもともと正しい運 動指令を出力しているにもかかわらず,誤った運動誤 差情報を受け取り,修正しなくてもよい運動指令をシ ナプス可塑性により修正してしまっているものがある のかもしれない[13]
.その場合,ある細胞の出力がそ のまま唯一の運動指令として制御対象に与えられ,運 動出力と一対一の因果関係が担保される状況のもとで 正しい運動誤差情報が与えられれば,その細胞は常に 運動誤差を減少させるように発火頻度を変化させるは ずである.従来のシステム神経科学の手法では,これらの仮説
や小脳神経細胞活動と運動出力の間の因果関係を直接 確かめることは困難であった.そこで本研究では,ブ レイン・マシンインタフェース(
BMI
)を用い,この 問題を解決するためのスキームを提案する.また,動 物実験により提案法の妥当性を評価する.2.
方 法2. 1
概 要提案法は,眼球運動制御を担う前庭小脳
Purkinje
細 胞の単一神経活動計測を行い,BMI
を介して外部機器(直流モータ)を制御することにより,単一
Purkinje
細胞発火頻度変化と直流モータ運動誤差の直接的因果 関係を評価可能にするものである.ここでは,前庭動 眼反射(Vestibulo-ocular reflex
:VOR
)の運動学習 パラダイムを利用する.VOR
は頭部運動時にそれと は逆方向に同じスピードで眼球を回転させることによ り,視界のぶれ(網膜上の像のブレ)を抑える反射性 の眼球運動である.通常,眼球運動速度/
頭部運動速 度で定義されるVOR
ゲインはほぼ1
であるが,前庭(頭部運動)
–
視覚競合刺激を与え続けることにより,こ の値を大きくしたり(ゲインUp
),小さくしたり(ゲ インDown
)することができる[14]
.これは,前庭–
視覚競合刺激により生じる網膜像のブレを低減するよ うに誘発される運動学習である.このとき,数時間程 度の短期間に誘発されるVOR
ゲイン変化は,小脳で 実現されるものと考えられている[15], [16]
.提案法で は,まず,モータの目標軌道を被験体への頭部回転刺 激として与える.このときの前庭小脳単一Purkinje
細胞の神経スパイクを計測し,BMI
により直流モー タ駆動用指令信号に変換し,モータを駆動する.普段,この
Purkinje
細胞は眼球を動かすための運動指令を コードしていると考えられており[4], [17], [18]
,その 活動でモータを駆動しても,初めモータは目標軌道通 りに動かないと予想される.そこで,モータの目標軌 道と実現軌道の差(運動誤差)を実時間で計算し,被 験体に網膜像のブレとして与える.通常は登上線維を 介して誤差情報を受け取る同一領域内のPurkinje
細 胞集団が網膜像のブレを低減するように活動を変化さ せ,VOR
が適応的に変化するものと考えられている が,この系では,モータの実現軌道を適応的に変えら れるのは計測中の単一Purkinje
細胞のみである.こ こで,金魚はモータを制御する意思や直接的な動機は ないが,頭が動いた際,網膜像のブレが生じるため,これを低減するように
VOR
運動学習が誘発されるはずである.このとき,網膜像のブレはモータの運動誤 差と等しいため,結果として網膜像のブレが低減され れば,モータの運動誤差が減ることになる.こうした 系により,記録中の単一
Purkinje
細胞活動のみによ りモータの動きが決定されることから,モータ実現軌 道の適応的変化が生じれば,その原因は計測中の細胞 活動変化に帰着される.2. 2
実験系と実験方法被 験 体 に は ,和 金(
Carassius auratus
,体 長 約12 cm
)を用いる.金魚はこれまで,VOR
をはじめ とする眼球運動の制御や学習に関するシステム神経 科学実験によく用いられており,小脳を含めVOR
に かかわる神経回路が解剖・生理学的によく同定され,霊長類との高い類似性が報告されている
[19]
.実験系 は,金魚用水槽,水槽下部に設置されたターンテーブ ル,水槽上部に設置されたプラネタリウム(視覚刺激 装置),神経電位計測装置,BMI
,制御対象直流モー タ(maxon
社,RE-max24
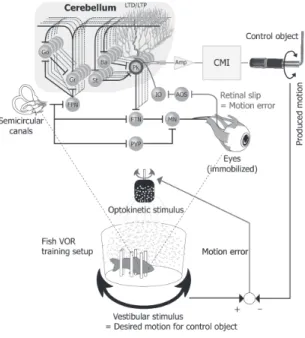
),並びに水質維持システ ムで構成される(図1
).図1
では,BMI
の代わりに,小脳・マシンインタフェース(
Cerebellum-Machine
図1 前庭動眼反射運動学習メカニズムを利用した小脳・
マシンインタフェース(CMI)による外部機器(直 流モータ)適応制御系
Fig. 1 Scheme of the adaptive direct current motor control by employing the vestibeloocular re- flex motor learning via cerebellar-machine in- terface (CMI).
Interface
)の意味を明確にするためCMI
と記してあ る.以後,本論文では,小脳・マシンインタフェースはCMI
,より一般的なブレイン・マシンインタフェース はBMI
と表記する.本実験系では,こうしたVOR
経 路内に含まれるPurkinje
細胞の中から一つを選択し,その神経活動で
CMI
を介して外部機器を制御する.金魚は,円筒形白色水槽の中央に左右半規管の中央 が一致するように固定される.水槽内の水は外部の循 環系に接続され,バイオフィルタ,温度調節器,エア レータにより水質・水温(
23
◦C
)が長時間一定に保た れるようになっている.水槽はターンテーブルの回転軸 に中心が一致するように設置され,ターンテーブルは サーボモータ(日本電装社,NMR-FFDB2B-801A
) により外部PC
からAD/DA
ボード(National In- struments
社,NI PCI-6024E
)を介して与えられる 指令(制御対象の目標軌道)に従うよう速度制御さ れる.本研究では,制御対象直流モータへの目標軌 道として,金魚VOR
のダイナミックレンジ内でこれ までの研究によく用いられている0.1 Hz
の正弦波を 用いた.水槽上部には白色LED
を光源にもつプラネ タリウムが設置されており,これにより水槽壁面に ランダムドットが投影され,サーボモータによりプラ ネタリウムを回転させることで,視運動性視覚刺激(
Optokinetic Stimulus
:OKS
)が金魚に与えられる.金魚に与えられた頭部回転刺激は三半規管(
Semi- circular canals
)で検出され,前庭神経核に送られる.前庭神経核内神経細胞の出力は苔状線維として小脳に 入力され,顆粒細胞(
Gr
)とGolgi
細胞(Go
)に投射 する.顆粒細胞の出力は平行線維となり,Goldgi
細胞,籠細胞(
Ba
),星状細胞(St
),Purkinje
細胞(Pk
) に接続する.このうちGolgi
細胞は顆粒細胞を抑制し,籠,星状細胞は
Purkinje
細胞を抑制する.Purkinje
細胞の出力が唯一小脳外に送られ,前庭神経核に投射 する.前庭神経核からの出力は外眼筋運動ニューロン(
MN
)を介して外眼筋に送られ,その結果,眼球が 頭部回転と逆方向に回転する.その際,眼球運動が頭 部運動を補償しきれなかったり,プラネタリウムの回 転によりOKS
が与えられたりすると網膜像のブレが 生じる.この網膜像のブレ情報は下オリーブ核(IO
) からPurkinje
細胞に登上線維によって送られる.こ れらの解剖・生理学的知見は主に哺乳類の研究で得ら れたものであるが,魚類でも基本回路構造は同一であ る[19]
.小脳
Purkinje
細胞神経電位は,2 mol/l
のNaCl
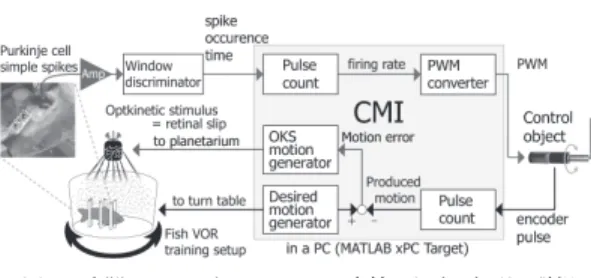
溶図2 小脳・マシンインタフェース内外における処理の詳細 Fig. 2 Detailed description of I/O to the CMI.
液を満たしたカスタムメードのガラス微小電極(先端 径約
2 µ m
,インピーダンス1.8
〜3.5 MΩ
)を用いて 計測する.水圧式3
軸マニピュレータ(Siskiyou
社,MX10R-H
)により位置を調整し,前庭小脳(vestibu- lolateral lobe
)Purkinje
細胞層[19]
をねらって電極を 刺入する.前庭刺激並びに眼球運動に反応するPurk- inje
細胞をisolate
し,細胞外神経電位を記録する.Purkinje
細胞は神経インパルスの発火頻度に情報を コードしていることから[4], [11], [17]
,神経電位をア ンプ(Neuralynx
社,ERP-27
)により増幅後,win- dow discriminator
(Cornerstone
社,WD-2
)により 神経インパルス発火時刻を検出し,カウンタボード(
National Instruments
社,NI PCI-6601
)を介してCMI
に入力する(図2
).本研究における
CMI
は,MATLAB xPC target
(
MathWorks
社)環境を利用して構築した.CMI
内 では,まず神経インパルス発火時刻データから発火頻 度F R
を算出する.この神経インパルス発火頻度を直 流モータ駆動用のパルス幅変調(PWM
)信号に変換 し,PWM
信号のデューティ比D [%]
を次式により算 出して,直流モータへ運動指令として与える.D = A ∗ F R − dc (1)
ここで,
A
は増幅の倍率であり,この値を変えること によりモータ実現軌道の振幅を調整する.dc
は発火頻 度の直流成分であり,発火頻度からこの値を差し引く ことにより,発火頻度が増した場合と減少した場合で モータの回転方向を変えられる.ここでは,F R
増加 時(D > 0
)にモータが反時計回り(CCW
),F R
減 少時(D < 0
)に時計回り(CW
)に回転するように 設定した.また,実験開始時には常にモータの回転速 度が目標軌道速度に満たないようA
の値を設定するこ とにより,Purkinje
細胞の発火頻度変調の振幅が増加 することにより運動誤差が減少するようにした.この ように算出されたPWM
信号のデューティ比により駆動される直流モータの実現軌道は,エンコーダの出力 をカウンタボード(同上)を介して
CMI
を構成するPC
に取り込み,PC
内で算出される.目標軌道とこ の実現軌道の差がモータの運動誤差(速度信号)とな る.この運動誤差信号をプラネタリウム駆動用のサー ボモータに入力することにより,運動誤差に相当するOKS
が金魚に与えられる.このとき,金魚の眼を後 述のように不動化しておくことにより,OKS
による 網膜像のブレはモータの運動誤差信号と一致し,運動 誤差が理想的に0
になると網膜像のブレもなくなるこ とになる.2. 3
被験体の実験前準備被験体の金魚には,実験
2
日以上前までに,麻酔下(
Sigma
社,MS-222
,1
:20 wt/vol
)において,水槽 に固定するためのヘッドポストを歯科用セメントによ り頭蓋骨(frontal bone
)に取り付ける.また同様に,小脳電位計測用に直径
5 mm
ほどの円形状開頭手術 をoccipital bone
に施す.開頭手術の際に取り除かれ た骨は,実験実施時まで魚を飼育槽で飼育するため,再び
“
蓋”
として穴を塞ぐために用いた.この蓋も歯 科用セメントにより固定並びに防水シールする.ま た,実験開始直前には,運動誤差情報を視覚刺激とし て網膜に正確に与えるために,眼禍と眼球のすき間にSuperglue
(Loctite
社,LGP-010
)を数滴入れ,金魚 の両眼を不動化する.実験時には,金魚を飼育水槽か ら実験用円筒形水槽に移し,2.2
の方法で固定する.その後,実験環境に慣らすために,明所において
30
分間以上安静状態に保つ.これらの実験手順は中部大 学動物実験委員会の承認を得て行った.2. 4
計測データ実験では,
CMI
内で計算される各信号のほか,Purk- inje
細胞の活動電位とその発火時刻,ターンテーブル(水槽)の回転速度(目標軌道),制御対象直流モータの 実現軌道,並びに運動誤差(
=
目標軌道−
実現軌道)をソフトウェア
Spike2
(CED
)を用い,A/D
ボードPower1401
(CED
)を介して,CMI
とは別のPC
内 に記録した.データ解析はMATLAB
上で行った.3.
結 果VOR
運動学習を担うと仮定されている前庭小脳Purkinje
細胞は,前庭刺激と視覚刺激に応答すること が知られている[18]
.金魚のvestibulolateral lobe
に 存在するPurkinje
細胞は,各々の刺激に対する応答 特性により,五つのタイプに分類されている[18]
.こ図3 CMIを利用した金魚前庭小脳単一Purkinje細胞 活動による直流モータ適応制御における計測データ 例.開始直後(A)と終了直前(B)の各々50秒間 Fig. 3 Example of recorded data from the adaptive direct current (dc) motor control by a single Purkinje cell in the vestibulolateral lobe of the goldfish cerebellum via the CMI. Initial (A) and last (B) 50 s.
れらのタイプは,
Purkinje
細胞が存在する小脳半球 と,その発火頻度が増加する前庭並びに視覚刺激の回 転方向との関係により分類される.本実験では,常に 小脳の左側からPurkinje
細胞を計測し,前庭刺激左 回転(CCW
)時と視覚刺激左回転時に発火頻度が増 加するH1E1
タイプ,前庭刺激左回転時と視覚刺激右 回転(CW
)時に発火頻度が増加するH1E2
タイプ,並びに前庭刺激左回転時に発火頻度が増加し,視覚刺 激には反応しない
H1
タイプを対象とした.このほか,視覚刺激のみに応じる
E1
,E2
というタイプの存在が 報告されているが[18]
,これらの細胞はモータ回転の 目標軌道情報を運ぶ前庭刺激に応じないことから,本 実験では計測対象外とした.提案スキームにおける,約
60
分間の単一Purkinje
細胞連続記録による直流モータ適応制御時の計測デー タ例を図3
に示す.ここでは,60
分のデータのうち,適応制御実験開始直後と終了直前の
50
秒間をそれぞ れA
,B
に示してある.上段からPurkinje
細胞の活 動電位,その発火頻度,目標軌道(点線)と実現軌道(実線)の重ね書き,最下段が運動誤差(目標軌道
–
実現軌道)である.A
,B
の比較より,制御開始直後(
A
)に比べ,約60
分後(B
)には運動誤差が減少し ているように見える.この運動誤差とPurkinje
細胞 発火頻度について,目標軌道の周期に合わせた周期平 均を求め,制御時間経過に伴う変化を求めた結果をそ れぞれ図4 A
,C
に示す.ここで,各周期平均波形を図4 小脳Purkinje細胞活動による直流モータ制御時の
運動誤差(A)とPurkinje細胞発火頻度(C),並 びに時計(CW)・反時計回り(CCW)別の平均誤 差(B)とPurkinje細胞平均発火頻度(D)の時間 変化例
Fig. 4 Changes in motion error and Purkinje cell fir- ing modulation during the adaptive dc mo- tor control by a vestibulolateral lobe Purkinje cell. A and C: motion error and Purkinje cell firing modulation, respectively. B and D: root mean square of the motion error and the Purk- inje cell firing rate evaluated separately during CW and CCW rotation of the dc motor.
求めるために用いた周期数は
10
,前後の周期平均波形 計算時の重複周期数は5
とした.また,これらの変化 を定量化するために,各周期平均結果をCW
回転区 間とCCW
回転区間に分け,運動誤差については次式を用いて平均運動誤差を求めた.
E
CW= 2 T
T /2 0
E
2( t ) dt (2)
E
CCW= 2 T
T T /2
E
2( t ) dt (3)
ここで,
E
は運動誤差の周期平均波形,T
は1
周期の 時間10
秒である.発火頻度については各区間の平均 値をとり平均発火頻度を求めた.これらの結果をそれ ぞれ同図B
,D
に示す.図4 A
では,学習時間の経過 とともに濃い黒の領域が徐々に薄くなる傾向にあり,運動誤差が減少していく様子が分かる(特に
CCW
区 間).また同図C
からは,A
の運動誤差の減少と平行 して発火頻度の振幅が増加していることが分かる.こ れらの変化をCW
とCCW
区間に分けて平均誤差と して定量的に評価した同図B
,C
からは,CCW
区間 の平均誤差が減少傾向にあり,発火頻度振幅もこれに 平行して増加傾向にあることが分かる.一方,CW
区 間の平均誤差及び発火頻度振幅は,はっきりとした増 加・減少傾向を示していない.10
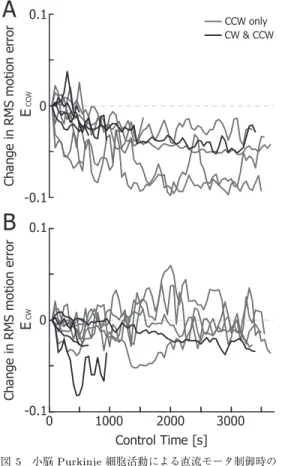
分以上の連続記録に成功した13
個のPurkinje
細胞のうち,CW
・CCW
いずれかまたは両区間にお ける運動誤差を減少させた10
個について,平均運動誤 差を重ね書きしたものを図5 A
(CCW
)とB
(CW
) に示す.ここで,縦軸の値は実験開始直後の平均運動 誤差値を0
とし,平均運動誤差の変化量を表している.この図から,
CW
・CCW
両区間の運動誤差を減少さ せたPurkinje
細胞は4
個(黒線),CCW
の運動誤差 のみを減少させたものは6
個(灰線),CW
のみの誤 差を減少させたものは0
個であった.その際,いずれ の場合も,誤差の減少は単調ではなく,小さな増減を 繰り返しながら減少していくものであった.ここで,本実験では,モータの
CW
回転はPurkinje
細胞発火 頻度が直流成分(式(1)
,dc
)よりも小さくなること により生じ,かつこの減少量が大きくなるほど運動誤 差が減少するように設定してある(詳しくは実験方法 参照).そのため,Purkinje
細胞の発火頻度振幅が大 きくなり0[spk/s]
に達した場合,CW
区間の誤差は それ以上減らすことができなくなる(発火頻度は負の 値をとれないため).そこで,CCW
区間のみの運動 誤差を減少させたPurkinje
細胞の発火頻度をこの点 に着目して評価したところ,いずれの細胞においても 適応制御実験中,発火頻度が0 [spk/s]
にクリップされ図5 小脳Purkinje細胞活動による直流モータ制御時の
反時計回り(CCW(A))と時計回り(CW(B))
別の平均運動誤差時間変化の重ね書き.平均運動 誤差を減少させた全て(10個)のPurkinje細胞 によるデータを使用.黒色はCW,CCW区間と も運動誤差を減少させたPurkinje細胞によるデー タ,灰色はCCW区間のみの運動誤差を減少させた Purkinje細胞によるデータ.
Fig. 5 Changes in root-mean-square motion errors evaluated separately during CCW (A) and CW (B) rotation of the dc motor as in Fig. 4 (B). Data from all the Purkinje cells (10 cells) that reduced motion error are super- imposed. Black lines are from Purkinje cells that reduced the root-mean-square motion er- ror during both CCW and CW rotations, and gray lines are from those that reduced the er- ror during only CCW period.
ていないことを確認した.
4.
む す び本研究では,小脳・マシンインタフェース(
CMI
) を利用し,運動学習と小脳Purkinje
細胞活動間の直 接的因果関係を評価するためのスキームを提案した.提案スキームは,運動学習のモデルシステムとして古
くから研究されている前庭動眼反射(
VOR
)の運動 学習メカニズムの枠組みの中で,単一Purkinje
細胞 神経活動による直流モータ適応制御を評価するもので ある.実験動物には金魚を用い,VOR
運動学習の座 と考えられてきた前庭小脳Purkinje
細胞からガラス 微小電極により細胞外電位を記録し,学習機能をもた ない単純なCMI
を介して直流モータを駆動した.そ の際,モータの運動誤差情報を,VOR
運動学習を誘 発する誤差信号である網膜像のブレとして金魚に与え ることにより,運動誤差とPurkinje
細胞活動の因果 関係を評価した.こうした系により,計測中の単一小 脳Purkinje
細胞活動と制御対象運動出力との間に一 対一の因果関係が担保される.実際にこの系を構築し て動物実験を実施した結果,以下の三つのことが示さ れた:1
)小脳は単一のPurkinje
細胞出力を運動指令 として制御対象(直流モータ)の運動誤差を適応的に 減少させることができる(記録したPurkinje
細胞13
個中10
個).その際,2
)運動誤差は単調には減少せ ず,増減を繰り返しながら減少していく(誤差を減少 させたPurkinje
細胞10
個中10
個).また,3
)運動 誤差の減少は制御対象直流モータのCCW
とCW
回 転両側で見られる場合と(10
個中4
個),CCW
回転 時のみで生じる場合があり(10
個中6
個),CW
側の みの誤差が減少することは極めてまれであった(10
個 中0
個).ここで,モータのCCW
,CW
回転はそれ ぞれPurkinje
細胞発火頻度が増加,減少する前庭–
視 覚競合刺激の方向に割り当てられており,また,全て のPurkinje
細胞は小脳左側から記録されたことから,CCW
回転が各Purkinje
細胞存在位置と同側となる.これらの結果は,
VOR
運動学習の枠組みの中で,前 庭小脳Purkinje
細胞活動変化と運動誤差減少の間の 直接的因果関係を初めて実証したものである.また,小脳
Purkinje
細胞が,その活動と運動出力の間に一 対一の因果関係が成り立っている場合にも,運動誤差 を単調に減少させることなく,中には誤差を減少させ ないものも存在する(13
個中3
個)ことを明示したも のである.また,上記3
)の結果は,頭部運動方向に 選択的なVOR
運動学習[20], [21]
が成立するための小 脳内メカニズムを提供するものと考えられる.本実験で計測対象とした
H1E1
とH1E2
タイプのPurkinje
細胞では,前庭(H
),視覚(E
)各刺激に対 するPurkinje
細胞の反応がそれぞれ異なる様式で加 法的に出力される.そのため,計測中のPurkinje
細 胞のタイプによって,与える目標軌道と視覚刺激の方向の組合せを変える必要があった.本実験では,発火 頻度振幅が増すと直流モータの運動誤差が減少するよ うに
Purkinje
細胞発火パターンとモータ回転の関係 を設定した.そのため,H1E2
タイプのPurkinje
細胞 を用いた実験では,発火頻度振幅が増す方向に変化す るよう前庭刺激と視覚刺激を逆方向に与えるVOR
ゲ インUp
パラダイムを適用し,H1E1
タイプでは,前 庭刺激と視覚刺激を同方向に与えるゲインDown
パラ ダイムを用いた.VOR
運動学習では,ゲインUp
と ゲインDown
学習において異なる学習・記憶保持特性 が示されており[22]
,小脳内での実現メカニズムの差 異も示唆されている[23]
.今後は,各タイプの細胞数 を増やすことにより,ゲインUp
とDown
のメカニズ ムの差異にも言及できるものと考えられる.謝辞 本研究の一部は文部科学省科学研究費補助金 基盤研究(
C
)並びに大幸財団の補助を受けた.文 献
[1] J.C. Eccles, M. Ito, and J. Szent´agothai, The cerebel- lum as a neuronal machine, Springer-Verlag, Berlin;
New York, 1967.
[2] M. Ito, The cerebellum and neural control, Raven Press, New York, 1984.
[3] 川人光男,脳の計算理論,産業図書,1996.
[4] M. Shidara, K. Kawano, H. Gomi, and M. Kawato,
“Inverse-dynamics model eye movement control by Purkinje cells in the cerebellum,” Nature, vol.365, no.6441, pp.50–52, 1993.
[5] M. Ito, “Long-term depression,” Annual Review of Neuroscience, vol.12, no.1, pp.85–102, 1989.
[6] M. Ito, “Cerebellar long-term depression: character- ization, signal transduction, and functional roles,”
Physiological reviews, vol.81, no.3, pp.1143–1195, 2001.
[7] M. Sakurai, “Synaptic modification of parallel fibre- Purkinje cell transmission in in vitro guinea-pig cere- bellar slices,” J. Physiology, vol.394, no.1, p.463, 1987.
[8] T. Hirano, “Depression and potentiation of the synaptic transmission between a granule cell and a Purkinje cell in rat cerebellar culture,” Neuroscience Letters, vol.119, no.2, pp.141–144, 1990.
[9] E. Watanabe, “Neuronal events correlated with long- term adaptation of the horizontal vestibulo-ocular reflex in the primate flocculus,” Brain Research, vol.297, no.1, pp.169–174, 1984.
[10] M. Kahlon and S.G. Lisberger, “Changes in the re- sponses of Purkinje cells in the floccular complex of monkeys after motor learning in smooth pursuit eye movements,” J. Neurophysiology, vol.84, no.6, pp.2945–2960, 2000.
[11] Y. Hirata and S. Highstein, “Acute adaptation of the
vestibuloocular reflex: signal processing by floccular and ventral parafloccular Purkinje cells,” J. Neuro- physiology, vol.85, no.5, pp.2267–2288, 2001.
[12] 川人光男,脳の情報を読み解くBMIが開く未来,朝日新 聞出版,2010.
[13] 平 田 豊 ,“記 憶 の シ ス テ ム:小 脳 ,” Clinical Neuro- science, vol.29, no.2, pp.167–171, 2011.
[14] 平田 豊,“動的視覚安定化の脳内情報処理,”日本神経回 路学会誌,vol.11, no.4, pp.176–192, 2004.
[15] C.D. Kassardjian, Y.F. Tan, J.Y.J. Chung, R.
Heskin, M.J. Peterson, and D.M. Broussard, “The site of a motor memory shifts with consolidation,” J.
Neuroscience, vol.25, no.35, pp.7979–7985, 2005.
[16] M. Anzai, H. Kitazawa, and S. Nagao, “Effects of reversible pharmacological shutdown of cerebel- lar flocculus on the memory of long-term horizontal vestibulo-ocular reflex adaptation in monkeys,” Neu- roscience research, vol.68, no.3, pp.191–198, 2010.
[17] Y. Hirata, A. Yoshikawa, P.M. Blazquez, and S.M.
Highstein, “Evaluation of the inverse dynamic model in cerebellum during visual-vestibular interactions at different VOR gains in squirrel monkeys,” Neurocom- puting, vol.65, pp.709–717, 2005.
[18] A.M. Pastor, R.R. Cruz, and R. Baker, “Character- ization of Purkinje cells in the goldfish cerebellum during eye movement and adaptive modification of the vestibulo-ocular reflex,” Progress in Brain Re- search, vol.114, pp.359–381, 1997.
[19] H. Straka, J.C. Beck, A.M. Pastor, and R.
Baker, “Morphology and physiology of the cerebellar vestibulolateral lobe pathways linked to oculomotor function in the goldfish,” J. Neurophysiology, vol.96, no.4, pp.1963–1980, 2006.
[20] 吉川明昌,平田 豊,“前庭動眼反射の方向選択的運動学習 とゲイン増減メカニズムの差異,”信学論(D),vol.J92-D, no.1, pp.176–185, Jan. 2009.
[21] Y. Hirata, J. Lockard, and S. Highstein, “Capacity of vertical VOR adaptation in squirrel monkey,” J.
Neurophysiology, vol.88, no.6, pp.3194–3207, 2002.
[22] Y. Kuki, Y. Hirata, P. Blazquez, S. Heiney, and S.
Highstein, “Memory retention of vestibuloocular re- flex motor learning in squirrel monkeys,” Neurore- port, vol.15, no.6, pp.1007–1011, 2004.
[23] E.S. Boyden, A. Katoh, and J.L. Raymond,
“Cerebellum-dependent learning: the role of multi- ple plasticity mechanisms,” Annual review of neuro- science, vol.27, pp.581–609, 2004.
(平成23年5月18日受付,11月12日再受付)
片桐 和真
平21中部大・工・情報卒.平23同大大 学院修士課程了.眼球運動の運動学習に関 する研究に従事.平20年度本学会東海支 部研究発表会優秀賞,平22年度電気関係 学会東海支部連合大会IEEE学生奨励賞各 受賞.
田中 良幸 (正員)
平16中部大・工・電子卒.平18同大大 学院修士課程了.現在,同大学術情報セン ター兼情報科学研究所勤務.眼球運動の運 動学習に関する生体・生理工学・計算論的 研究に従事.日本ロボット学会,日本神経 回路学会各会員.
平田 豊 (正員)
平2豊橋技科大・工・電気・電子卒.平 4同大大学院修士課程了.平7同大学院博 士課程システム情報工学専攻了.博士(工 学).同年宇宙開発事業団招聘研究員(新 技術事業団科学技術特別研究員).平9ワ シントン大・医・Research Associate(平 10より科学技術振興事業団若手長期在外研究員).平12中部 大学工学部電子工学科講師,助教授を経て,平16同大情報工 学科教授.眼球運動の運動学習に関する生体・生理工学・計算 論的研究,瞳孔の制御機構とその自律神経モニタへの応用に従 事.平7年度本会論文賞,猪瀬賞各受賞.日本視覚学会,日本 神経回路学会,IEEE,北米神経科学会各会員.