目 次 --- 2
I 背 景 --- --- --3
II 研 究 方 法 --- ---12
1. 使 用 系 統 --- ---12
2. RNA 干 渉 法 (RNA interference) --- ---13
3. Gal4-UAS system--- ---13
4. 電 気 生 理 学 的 解 析 --- ---14
4-1. 細 胞 内 電 位 記 録 法 --- ---14
4-2. Paired pulse stimulation (PPS) --- ----16
4-3. 高 頻 度 反 復 刺 激 に よ る シ ナ プ ス プ ー ル サ イ ズ の 測 定 ---16 4-4. 電 気 生 理 学 的 手 法 に お け る 解 析 方 法 --- ---17 5. 形 態 観 察 --- ---18 III 実 験 結 果 --- ---19 1. P 因 子 挿 入 に よ る cenG1A 変 異 体 に お け る 電 気 生 理 学 的 解 析 ---19 2. RNA 干 渉 法 に よ り 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 ---21 2-1. 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 に お け る 電 気 生 理 学 的 解 析 ---21 2-2. 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 に お け る シ ナ プ ス 形 態 解 析 ---25 2-3. 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 に お け る 放 出 確 率 の 解 析 ---28 2-4. 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 に お け る シ ナ プ ス 小 胞 数 の 解 析 ---31 IV 考 察 --- ---33 1. 神 経 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 ---33 2. 筋 肉 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 ---35
3. negative regulator と し て の CenG1A 機 能 --- ---36
V 謝 辞 --- ---38
4
シ ナ プ ス 前 細 胞 に 存 在 す る 神 経 伝 達 物 質 を 含 ん だ シ ナ プ ス 小 胞 は 、 放 出 可 能 プ ー ル (RRP: Readily releasable pool)と 、 貯 蔵 プ ー ル (RP: reserve pool)の 主 に 二 種 類 の プ ー ル に 分 か れ て い る 。 RRP の 小 胞 は シ ナ プ ス 前 膜 に 付 着 し た 状 態 で 存 在 し 、 シ ナ プ ス 前 膜 の 脱 分 極 に よ る Ca2 +の 流 入 に よ り 、 直 ち に シ ナ プ ス 前 膜 の 放 出 活 性 体 (active zone)に 融 合 す る こ と が で き る 。 こ の シ ナ プ ス 小 胞 の 前 膜 へ の 付 着 と 融 合 の メ カ ニ ズ ム と し て SNARE 仮 説 が 提 唱 さ れ て い る 。 SNARE 仮 説 と は 、 小 胞 の 膜 へ の 付 着 が 、小 胞 膜 上 に 存 在 す る v-SNARE、シ ナ プ ス 前 膜 側 に 存 在 す る t-SNARE と 細 胞 質 タ ン パ ク 質 で あ る SNAPs や NSF 等 と の 相 互 結 合 に よ る も の だ と す る と い う 仮 説 で あ る 。 他 に も 、 シ ナ プ ト タ グ ミ ン 等 の 多 く の タ ン パ ク 質 が 関 与 し 、 標 的 膜 へ の 付 着 と 融 合 を 達 成 し て い る と 考 え ら れ て い る (Cowan et al, 2001)。一 方 、RP の 小 胞 は 、 ア ク チ ン で 構 成 さ れ た 細 胞 骨 格 に 結 合 し て お り 、 シ ナ プ ス 前 膜 の 脱 分 極 に よ る 細 胞 内 へ の Ca2 +の 流 入 後 直 ち に は 放 出 さ れ ず 、 RP か ら RRP へ と 移 動 す る 。 こ の 二 種 の プ ー ル (RP と RRP)は 更 に 分 類 す る こ と が で き 、 放 出 さ れ る 能 力 を 持 つ が 前 膜 に 付 着 し て い な い‘ non-RRP’で あ る 小 胞 の プ ー ル を 、リ サ イ ク リ ン グ プ ー ル と 呼 び 、小 胞 は RP、 リ サ イ ク リ ン グ プ ー ル 、 RRP の 3 つ の プ ー ル に 分 け ら れ る と 考 え ら れ て い る 場 合 も あ る (図 2A)(Zucker and Regehr, 2002)。
図 2. シナプス小胞プール
5
シ ナ プ ス 伝 達 で は 、神 経 伝 達 物 質 が 放 出 さ れ る と 、細 胞 膜 と 融 合 し た 小 胞 膜 が 、 エ ン ド サ イ ト ー シ ス に よ り 再 び 取 り 込 ま れ 、 神 経 伝 達 物 質 を つ め ら れ て 再 利 用 さ れ る リ サ イ ク リ ン グ と い う 現 象 が お き て い る 。シ ナ プ ス 小 胞 リ サ イ ク リ ン グ に は 、 2 つ の 反 応 の 速 い 経 路 と 1 つ の 遅 い 経 路 の 計 3 つ の 経 路 が あ る 。1 つ め は 、active zone に 小 胞 が 結 合 し た ま ま 神 経 伝 達 物 質 を 最 充 填 す る kiss and stay。 2 つ め は 、 局 所 的 な リ サ イ ク リ ン グ で ク ラ ス リ ン 非 依 存 的 に 行 わ れ る Kiss and run。 3 つ め は 、 ク ラ ス リ ン 依 存 的 な エ ン ド サ イ ト ー シ ス に よ る endosomal recycling で あ る (図 3)。 こ の 3 つ め の 経 路 で は 、ク ラ ス リ ン と い う タ ン パ ク 質 が 重 要 な 役 割 を 担 っ て い る 。ク ラ ス リ ン は 膜 の 細 胞 質 に 面 し た 側 に か ご 状 の 網 目 構 造 を 組 み 上 げ 、 細 胞 膜 を 出 芽 さ せ る 。 出 芽 し た 小 胞 の 根 元 を 囲 む よ う に ダ イ ナ ミ ン タ ン パ ク が 並 び 、 小 胞 を 膜 か ら く び り 切 る こ と に よ り 、 小 胞 は 前 膜 か ら エ ン ド サ イ ト ー シ ス さ れ 細 胞 内 に 回 収 さ れ る 。 こ れ ら の 経 路 に よ り シ ナ プ ス 小 胞 は 再 び プ ー ル に 充 填 さ れ る 。 こ の シ ナ プ ス 小 胞 リ サ イ ク リ ン グ の 過 程 に も 多 く の 分 子 が 関 与 し て お り 、 た と え ば 、 低 分 子 量 G タ ン パ ク で あ る Rab5 や Rab11 な ど が 関 与 し て い る こ と が 報 告 さ れ て い る (Maxfield and McGraw, 2004)。Rab5 は endosome の 形 成 と 機 能 に 関 与 し て お り (Lawe et al., 2002)、Rab11 は 、エ ン ド サ イ ト ー シ ス に よ り 取 り 込 ま れ た 分 子 の リ サ イ ク リ ン グ を 制 御 し て い る と い う 報 告 が あ る (Chen et al., 1998; Ren et al., 1998)。こ れ ら 多 く の 因 子 が 働 き 、 シ ナ プ ス 小 胞 放 出 過 程 が 適 切 に 行 わ れ る こ と で 、 シ ナ プ ス 伝 達 が 達 成 さ れ て い る 。
6 図 3. シナプス小胞リサイクリング経路
Active zone に結合したまま伝達物質を充填される(Kiss-and-stay)。クラスリン非依存的で局所的なエンド
7 シ ョ ウ ジ ョ ウ バ エ に つ い て モ デ ル 生 物 と し て の シ ョ ウ ジ ョ ウ バ エ シ ョ ウ ジ ョ ウ バ エ は 、 ラ イ フ サ イ ク ル が 短 く 扱 い や す い た め 、 古 く か ら モ デ ル 生 物 と し て 多 く の 研 究 で 使 わ れ て き た 。 そ の た め 、 数 々 の 遺 伝 学 的 、 分 子 生 物 学 的 手 法 が 確 立 さ れ て お り 、 分 子 レ ベ ル に お い て も 個 体 レ ベ ル に お い て も 解 析 が 可 能 で あ る 。 こ れ ら の 理 由 か ら 優 れ た モ デ ル 生 物 と な っ て お り 様 々 な 研 究 に 用 い ら れ て い る (Keshishian et al., 1996; Kazama et al., 2003; Marqués and Zhang, 2006; Uytterhoeven et al., 2011; Müller et al., 2011)。 シ ョ ウ ジ ョ ウ バ エ で 用 い る 遺 伝 学 的 手 法 の 代 表 例 と し て GAL4 エ ン ハ ン サ ー ト ラ ッ プ 法 が あ る (Brand and Perrimon, 1993)。 こ れ は 、 ゲ ノ ム 上 に 存 在 す る 組 織 特 異 的 な エ ン ハ ン サ ー 活 性 を 利 用 し て 、 酵 母 由 来 の 転 写 因 子 GAL4 を 発 現 さ せ る 手 法 で あ る 。 エ ン ハ ン サ ー と は 遺 伝 子 の 前 後 や イ ン ト ロ ン の 中 な ど に 位 置 し 、付 近 の DNA 領 域 の 転 写 活 性 を 調 節 す る DNA 領 域 の こ と を 示 す 。 シ ョ ウ ジ ョ ウ バ エ の ゲ ノ ム に は 少 な く と も 数 千 以 上 の エ ン ハ ン サ ー が 存 在 す る こ と が わ か っ て い る 。 ま た 、 シ ョ ウ ジ ョ ウ バ エ は ト ラ ン ス ポ ゾ ン の 一 種 で あ る P 因 子 を も っ て い る 。P 因 子 は P 因 子 転 移 酵 素 に よ り ゲ ノ ム 内 を ジ ャ ン プ す る こ と が で き る 。 酵 母 由 来 の 転 写 調 節 因 子 で あ る gal4 遺 伝 子 を こ の P 因 子 に 組 み 込 む こ と に よ っ て 、 ゲ ノ ム 内 の さ ま ざ ま な 位 置 に gal4 遺 伝 子 が 挿 入 さ れ た エ ン ハ ン サ ー ト ラ ッ プ 系 統 が 作 成 で き る 。 そ れ ぞ れ の エ ン ハ ン サ ー ト ラ ッ プ 系 統 で は 、P 因 子 挿 入 位 置 の 近 く に あ る エ ン ハ ン サ ー の 調 節 を 受 け て GAL4 タ ン パ ク 質 が 発 現 す る 。 こ の GAL4 タ ン パ ク 質 は 、 UAS (upstream activator sequence) と 呼 ば れ る DNA 配 列 に 連 な る 遺 伝 子 の 発 現 を 誘 導 す る 。 任 意 の 遺 伝 子 を UAS の 下 流 に 組 み 込 ん だ 系 統 と 、GAL4 エ ン ハ ン サ ー ト ラ ッ プ 系 統 を 掛 け 合 わ せ る こ と に よ り 次 の 世 代 の 個 体 は GAL4 タ ン パ ク 質 が 発 現 し て い る 組 織 で の み 、 UAS 下 流 の 任 意 の 遺 伝 子 が 発 現 す る 個 体 と な る 。 様 々 な GAL4 エ ン ハ ン サ ー 系 統 、 UAS 系 統 が ゲ ノ ム プ ロ ジ ェ ク ト に よ り 作 成 さ れ 、 ス ト ッ ク セ ン タ ー に 保 存 さ れ て い る
8 シ ョ ウ ジ ョ ウ バ エ 幼 虫 神 経 筋 接 合 部 位 シ ョ ウ ジ ョ ウ バ エ 幼 虫 の 神 経 筋 接 合 部 位 は 、 A2-A7 の 各 体 節 に 片 側 あ た り 30 個 の 筋 肉 細 胞 が 規 則 正 し く な ら び 、 腹 部 神 経 節 か ら 伸 び た 運 動 ニ ュ ー ロ ン が ど の 筋 肉 細 胞 に シ ナ プ ス を 形 成 し て い る か が 明 ら か に な っ て い る (keshishian et al., 1996) (図 4)。 そ の た め 、 シ ナ プ ス の 応 答 と 形 態 を 単 一 シ ナ プ ス レ ベ ル で 解 析 す る こ と が 可 能 で あ り 、 シ ナ プ ス 形 成 、 機 能 調 節 の 分 子 メ カ ニ ズ ム の 解 明 に 適 し た 実 験 系 で あ る と 考 え ら れ て い る 。 シ ョ ウ ジ ョ ウ バ エ 神 経 筋 接 合 部 位 に お け る 情 報 伝 達 も 、 神 経 終 末 膜 の 脱 分 極 に よ り 、 シ ナ プ ス 小 胞 の エ キ ソ サ イ ト ー シ ス が お き 、 受 容 体 が 活 性 化 さ れ る こ と に よ っ て 行 わ れ て い る 。 シ ョ ウ ジ ョ ウ バ エ 神 経 筋 シ ナ プ ス で は 、 シ ナ プ ス 間 隙 に 放 出 さ れ る 神 経 伝 達 物 質 は グ ル タ ミ ン 酸 で あ る 。 シ ョ ウ ジ ョ ウ バ エ 幼 虫 の 筋 肉 細 胞 に は 、2 種 の グ ル タ ミ ン 酸 受 容 体 が 存 在 す る 。細 胞 内 電 位 記 録 法 を 用 い 、筋 肉 細 胞 の 静 止 電 位 か ら 、 受 容 体 へ の 伝 達 物 質 結 合 に よ っ て 生 じ た 筋 肉 細 胞 膜 電 位 変 化 を 記 録 す る こ と に よ っ て 、 シ ナ プ ス 機 能 に つ い て 解 析 を 行 う こ と が 可 能 で あ る 。 ま た 、 シ ョ ウ ジ ョ ウ バ エ の 神 経 筋 シ ナ プ ス の シ ナ プ ス 前 細 胞 に は 、 タ ー ミ ナ ル に 約 84,000 の 量 子 (小 胞 )を 含 ん で お り 、 全 シ ナ プ ス 小 胞 の 約 80% が 貯 蔵 プ ー ル に (Reserve pool: RP)、14~ 19% が リ サ イ ク リ ン グ プ ー ル (Recycling pool)、約 0.4% が 即 時 放 出 可 能 プ ー ル (readily releasable pool: RRP)に 貯 蔵 さ れ て い る と い わ れ て い る (Rizzoli and Betz, 2005)(図 2B)。
図 4. ショウジョウバエ神経筋接合部位
ショウジョウバエ幼虫神経筋接合部位は、半体節に約 35 のモーターニューロンが 30 個の筋肉細胞に投 射している。ISN: intersegmental nerve, SN: segmental nerve, vm: ventral midline, dm: dorsal midline.
9
シ ョ ウ ジ ョ ウ バ エ 神 経 筋 接 合 部 位 を 用 い た 研 究
神 経 筋 接 合 部 位 を モ デ ル 系 と し て 、 様 々 な 因 子 の シ ナ プ ス で の 機 能 が 研 究 さ れ て い る 。 例 え ば 、 Phoshoinositide 3-kinase (PI3K)が 神 経 筋 シ ナ プ ス 伝 達 の negative regultor で あ る 可 能 性 を 示 唆 す る 研 究 や (Howlett et al., 2008)、細 胞 接 着 因 子 で あ る Fasciclin II (FasII)の 神 経 筋 シ ナ プ ス 発 達 や 可 塑 性 に お け る 機 能 な ど が 研 究 さ れ て い る (Davis et al., 1997; Sánchez-Soriano and Prokop, 2005 )。最 近 の 研 究 で は 、FasII が 足 場 タ ン パ ク 質 で あ る Discs large (Dlg)と 共 に 機 能 し 、 活 動 依 存 的 な シ ナ プ ス 発 達 を 調 節 し て い る と い う こ と も 明 ら か に な っ た (Beumer et al.,2002; Kazama et al., 2007; Morimoto et al., 2010)。ま た 、シ ナ プ ス 後 細 胞 か ら 、前 細 胞 に 働 き か け 伝 達 物 質 放 出 過 程 を 調 節 す る 逆 行 性 シ グ ナ ル (Retrograde signal) が 存 在 す る こ と が 明 ら か に な っ て お り (Davis and Goodman, 2002; Davis, 2006)、bone morphogenetic protein (BMP)と い う シ グ ナ ル 分 子 が 同 定 さ れ て い る (Aberle et al., 2002; Marqués et al., 2002; Marqués and Zhang, 2006)。 こ の よ う に 、 神 経 筋 接 合 部 位 を 用 い た 研 究 で 多 く の 知 見 が 得 ら れ て い る 。
Centaurin に つ い て
10
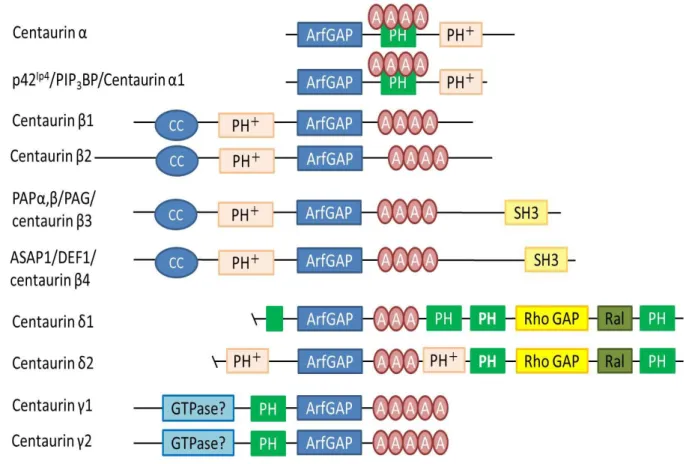
告 も あ る (Wassink et al., 2005) 。し か し 、Centaurin 遺 伝 子 の シ ナ プ ス に お け る 機 能 、 自 閉 症 と の 関 連 は ま だ 明 ら か に な っ て い な い 。Centaurin が ArfGAP や PH な ど の ド メ イ ン を も つ こ と か ら 、 私 は Centaurin family が 小 胞 の リ サ イ ク リ ン グ に 関 係 し て い る 可 能 性 が あ る と 考 え た 。 シ ョ ウ ジ ョ ウ バ エ に お い て centaurin 遺 伝 子 は 、 遺 伝 子 が 2 種 (β, γ)、 1 つ の 遺 伝 子 か ら 産 生 さ れ る isoform も 3 つ (A, B, C)の み で 哺 乳 類 と 比 較 す る と 圧 倒 的 に 分 子 の 数 が 少 な い 。 本 研 究 で は 、 シ ョ ウ ジ ョ ウ バ エ の 2 つ の centaurin 遺 伝 子 の う ち 、 centaurin gamma 1A (cenG1A) に 着 目 し た 。 cenG1A 遺 伝 子 は 、 シ ョ ウ ジ ョ ウ バ エ 幼 虫 の 筋 肉 細 胞 に も 神 経 細 胞 に も 存 在 し 、 発 生 段 階 に お い て 運 動 ニ ュ ー ロ ン が 筋 肉 細 胞 に 投 射 し て い く 過 程 で 、 運 動 ニ ュ ー ロ ン の 投 射 に 依 存 し て 筋 肉 細 胞 内 で 発 現 量 が 増 加 し て い た 遺 伝 子 で あ る こ と が わ か っ て い る (Flybase, Fukui et al., 2012)。そ の た め 、CenG1A は 神 経 筋 シ ナ プ ス に お い て も 何 ら か の 機 能 を も つ こ と が 期 待 で き る 分 子 で あ る 。
図 5. Centaurin family
12 II 研 究 方 法 1, 使 用 系 統 全 て の 実 験 に お い て キ イ ロ シ ョ ウ ジ ョ ウ バ エ (Drosophila melanogaster)の 25℃ で 飼 育 し た 3 齢 幼 虫 を 実 験 動 物 と し て 用 い た 。 cenG1A 遺 伝 子 の シ ナ プ ス 機 能 へ の 影 響 を 調 べ る た め に 、P 因 子 挿 入 に よ り cenG1A 遺 伝 子 に 変 異 が お き て い る 個 体 (20232, 12957)(図 6)と 、Gal4-UAS system (Brand and Perrimon, 1993) を 用 い て 、組 織 特 異 的 に cenG1A 遺 伝 子 の 発 現 を 調 節 し た 個 体 (図 6)を 使 用 し た 。 Gal4-UAS system と は 、 Gal4 系 統 と UAS(upstream activating sequence) 系 統 を か け あ わ せ る こ と で 、 任 意 の 分 子 を 特 定 の 組 織 に 発 現 さ せ る こ と が で き る 実 験 系 で あ る (後 述 )。
本 研 究 で は 、 以 下 の 系 統 の 個 体 を 実 験 に 使 用 し た 。
・ Yellow White (yw) cenG1A 遺 伝 子 の 変 異 体 の 遺 伝 的 バ ッ ク グ ラ ウ ン ド に な っ て い る 個 体 。 対 照 個 体 と し て 用 い た 。
・ 20232, 12957 P 因 子 挿 入 に よ り cenG1A 遺 伝 子 に 変 異 が 起 き て い る 個 体 (Bellen et al., 2004)。
・ elav-Gal4 全 神 経 細 胞 特 異 的 に GAL4 タ ン パ ク 質 を 発 現 し て い る GAL4 系 統 。 ・ 24B-Gal4 全 筋 肉 細 胞 特 異 的 に GAL4 タ ン パ ク 質 を 発 現 し て い る GAL4 系 統 。 ・ UAS-31811R-2(X)RNAi UAS 配 列 の 下 流 に cenG1A 遺 伝 子 に 対 す る RNA 干 渉 を
13 2, RNA 干 渉 (RNAi: RNA interference)
RNAi と は 、遺 伝 子 を 不 活 性 化 す る こ と が で き る 手 法 で あ る 。細 胞 ま た は 生 物 に 不 活 性 化 し た い 遺 伝 子 と 塩 基 配 列 が 一 致 す る 二 本 鎖 RNA 分 子 を 導 入 す る 。二 本 鎖 RNA は Dicer と い う 二 本 鎖 合 成 酵 素 に よ っ て 21bp 程 度 に 小 さ く 切 断 さ れ る 。こ の 断 片 が RISK と い う タ ン パ ク 複 合 体 と 結 合 し 、 標 的 遺 伝 子 か ら つ く ら れ る mRNA と ハ イ ブ リ ッ ト 形 成 し 、 分 解 、 ま た は 翻 訳 を 阻 害 す る 。 分 解 に よ っ て 生 じ た 短 い 断 片 RNA は 新 た な 二 本 鎖 RNA 分 子 作 り に 使 わ れ 、 標 的 遺 伝 子 の mRNA は 絶 え ず 分 解 さ れ る (Alberts et al., 2002)。こ の よ う な メ カ ニ ズ ム に よ り 、RNAi を 用 い る こ と で 標 的 遺 伝 子 を 不 活 性 化 す る こ と が で き る 。シ ョ ウ ジ ョ ウ バ エ に は 2 つ の Dicer (Dicer-1, Dicer-2)が 存 在 す る 。 シ ョ ウ ジ ョ ウ バ エ RNAi 機 構 に お い て は , 細 胞 内 に 取 り 込 ま れ た 外 来 二 本 鎖 RNA が Dicer-2 お よ び そ の 結 合 因 子 R2D2 依 存 的 に 小 分 子 RNA( small interfering RNA; siRNA) に 切 断 さ れ , 一 本 鎖 siRNA が RISC 中 核 因 子 AGO2 に 取 り 込 ま れ る こ と で RISC が 形 成 さ れ る 。 様 々 な 遺 伝 子 に 対 し て RNA 干 渉 を 起 こ す 配 列 を も つ UAS 系 統 が 作 成 さ れ 、stock center に 保 存 さ れ て い る (Drosophila Genetic Resource Center (DGRC) 京 都 )。 ま た 、 こ れ ら の 系 統 に Dicer を 共 発 現 さ せ る こ と で 、 RNA 干 渉 が 促 進 さ れ る (Dietzl et al., 2007)。 本 研 究 で は 、 centaurin 遺 伝 子 に 対 す る RNAi 配 列 を も つ UAS-31811R-2(X)RNAi と い う 系 統 を 使 用 し た 。
3, Gal4-UAS system
14 図 7. GAL4-UAS system
Gal4 タンパク質が発現している場所でのみ、UAS の下流の任意の配列の発現を調節できる実験系。本 研究では、神経細胞特異的に Gal4 タンパクを発現している elav-Gal4、筋肉細胞特異的に Gal4 タンパク を発現している 24B-Gal4。cenG1A 遺伝子に対する RNAi 配列をもつ UAS-31811R-2(X)RNAi。
15
2μm 以 下 、抵 抗 15MΩ 以 上 )を 挿 入 し て 行 っ た (図 8 下 図 )。1 個 体 か ら は 1 体 節 の み を 使 用 し た 。 増 幅 器 は Multiclamp700A (Axson instruments Inc., CA, USA)、 AD 変 換 機 は Digidata1322A (Axson instruments Inc., CA, USA)を 使 用 し た 。
記 録 測 定 時 の 外 液 は カ ル シ ウ ム 濃 度 を 二 種 類 (CaCl2 1.5mM,CaCl2 0.375mM) に 変
化 さ せ た 。刺 激 電 極 か ら 吸 い 込 ん だ 神 経 束 に 2- 4mV, 0.2Hz の 刺 激 を 10 回 与 え て 、 刺 激 に よ る 神 経 伝 達 物 質 放 出 に 伴 う 膜 電 位 の 変 化 (Excitatory junctional potentials: EJPs)を ガ ラ ス 記 録 電 極 に よ り 記 録 し た 。 静 止 膜 電 位 か ら ピ ー ク 電 位 ま で の 差 異 を EJP の 大 き さ (EJPs amplitude)と し 、10 回 の 刺 激 に よ る 応 答 を 1 個 体 の EJPs amplitude と し た 。 さ ら に 、 個 体 ご と に 得 ら れ た EJPs amplitude を 平 均 し 、 そ の 系 統 の EJPs amplitude の 値 と し て 使 用 し た 。ま た 、0.375mM の CaCl2を 含 む HL3 溶 液 下 で 刺 激
16 4-2, Paired Pulse Stimulation (PPS)
Paired Pulse Stimulation (PPS)は 、数 十 ミ リ 秒 の 時 間 間 隔 で 二 回 刺 激 を 行 い 、シ ナ プ ス 伝 達 を 誘 起 さ せ る 手 法 で あ る 。 一 般 に 、 一 回 目 の 刺 激 に よ る 応 答 に 対 し 、 二 回 目 の 刺 激 に よ る 応 答 が 増 大 す る 。こ れ を Paired pulse facilitation (PPF)と い い 、残 存 Ca2 +に 起 因 す る 現 象 で あ る と い わ れ て い る (Zucker and Regehr, 2002)。 PPS に お
17
図 9. 高頻度刺激による放出可能シナプス小胞数の測定
50Hz の高頻度刺激を 30sec 行ったときの EJPs 記録。刺激開始から 刺激終了時にかけて EJPs amplitude は徐々に減衰してくる。Scale bar は 5000ms, 10mV。
4-4, 電 気 生 理 学 的 手 法 に お け る 解 析 方 法
EJPs amplitude, mEJPs amplitude の 算 出 は 全 て Mini analysis (Synaptosoft Inc., NJ USA)を 使 用 し た 。膜 電 位 が -55mV~ -76mV の デ ー タ を 使 用 し た 。PPS、高 頻 度 刺 激 に お け る amplitude は 、 Clamp fit 9.0 (Axson instruments Inc., CA, USA ) を 用 い て 静 止 膜 電 位 か ら 、最 大 の ポ イ ン ト を 手 動 で 測 定 し た (図 10)。全 て の デ ー タ は 、one-way analysis of variance (ANOVA)に よ り 統 計 処 理 を 行 っ た 。
図 10. EJP における amplitude の測定
18 5, 形 態 学 的 解 析 CenG1A 発 現 を 抑 制 し た 際 に シ ナ プ ス 形 態 に 与 え る 影 響 を 解 析 す る た め に 、抗 体 染 色 を 用 い て 、生 理 学 実 験 に 用 い た 6 番 筋 肉 を 含 む 、6,7 番 筋 肉 の シ ナ プ ス の 形 態 観 察 を 行 っ た 。 一 次 抗 体 と し て 、
goat anti-horse radish peroxides (HRP), mouse anti-GluRIIA, mouse anti-nc82 二 次 抗 体 と し て
donkey anti-mouse Alexa488, donkey anti-goot Alexa594 (Molecular Probes, Engene, OR, USA, ) を 使 用 し た 。 観 察 は 共 焦 点 レ ー ザ ー 顕 微 鏡 (Olympus FV1000D IX81 confocal laser scanning microscope, Olympus, Tokyo, Japan) を 用 い て 行 い 、 解 析 は IPLab software (Scanalytics, Fairfax, VA, USA) を 用 い て 行 っ た 。
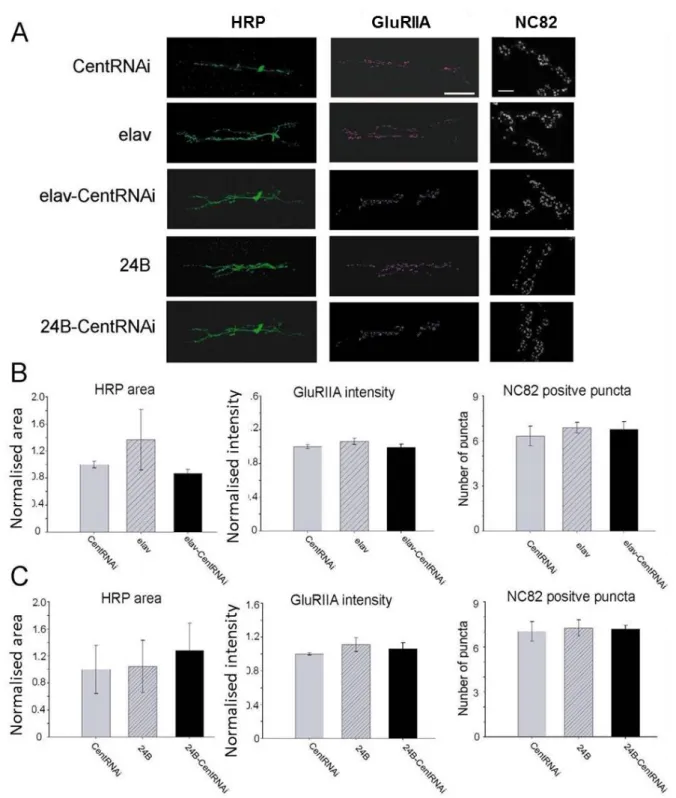
シ ナ プ ス 前 細 胞 の マ ー カ ー と し て 使 用 し た 抗 HRP 抗 体 を 用 い る こ と に よ り 染 色 さ れ た 神 経 細 胞 (図 11A 上 図 )の 面 積 を そ の 神 経 が 投 射 し て い る 筋 肉 細 胞 の 面 積 で 割 り 付 け た 値 を 神 経 の 面 積 と し て 使 用 し た 。ま た 、GluRIIA 抗 体 を 用 い る こ と に よ り 染 色 さ れ た 6,7 番 筋 肉 の シ ナ プ ス の GluRIIA(図 11A 下 図 )の 蛍 光 強 度 を GluRIIA 量 と し て 用 い た 。HRP に 対 す る 抗 体 で 染 色 さ れ た エ リ ア の 面 積 、GluRIIA の 蛍 光 強 度 の 値 は 、そ れ ぞ れ CentRNAi の 個 体 か ら 得 ら れ た 値 で ノ ー マ ラ イ ズ し 、そ れ ぞ れ の 系 統 の 値 と し て 使 用 し た 。さ ら に active zone の マ ー カ ー で あ る nc82 を 用 い る こ と に よ り 染 色 さ れ た puncta の 数 は 、6.7 番 筋 肉 に 投 射 し て い る シ ナ プ ス か ら ラ ン ダ ム に 4 つ の bouton (図 11B)を 選 択 し 、そ の bouton 内 に 観 察 で き る nc82 positive puncta を 測 定 、 bouton 一 つ 分 の puncta 数 の 平 均 を そ の 個 体 の puncta 数 と し て 用 い た 。
図 11. 抗体染色による形態観察
A,上図, HRP に対する抗体によって染色された 6,7 番筋シナプスの神経細胞。下図, GuRIIA 抗体によ って染色された 6,7 番筋シナプスの GluRIIA 受容体。Scale bar は 50μm。B, NC82 によって染色され た active zone。円は 1 つの bouton を示す。1 サンプルからランダムに 4 つの bouton を選択して puncta を数えた。Scale bar は 10μm。
HRP HRP
GluRIIA
20
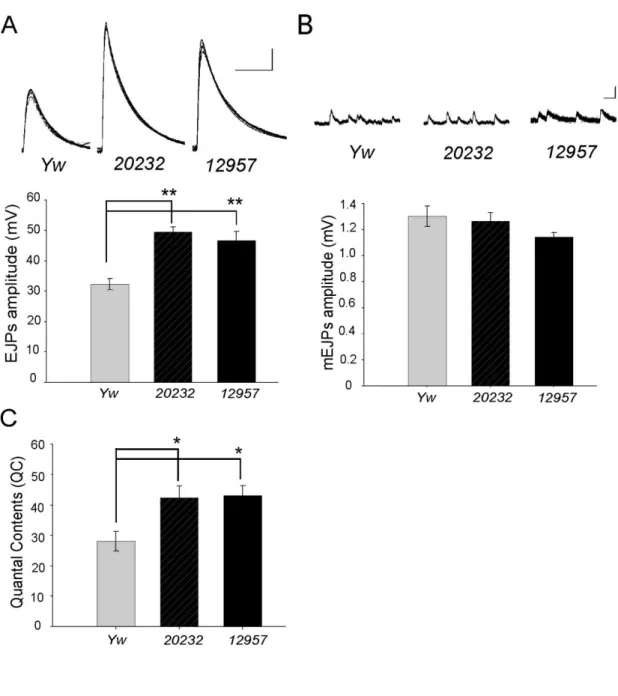
図 12. P 因子挿入による cenG1A 変異体における電気生理学的解析
21
C, Quantal content の平均値のグラフを示す。二種の cenG1A 変異体(中央:20232、右:12957)は対照個体 (左:Yw)と比較して QC が有意に増大している (yw, n=11; 20232, n=9; 12957, n=7. *P<0.01, ANOVA)。Bars は平均値±標 準 誤 差 。
2, RNA 干 渉 法 に よ り 組 織 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体
cenG1A 遺 伝 子 の 変 異 体 に お け る 電 気 生 理 学 的 解 析 の 結 果 か ら 、 CenG1A が 神 経 筋 シ ナ プ ス 伝 達 機 構 に お い て 機 能 を も つ 可 能 性 が 高 い こ と が 示 唆 さ れ た 。 シ ョ ウ ジ ョ ウ バ エ 神 経 筋 シ ナ プ ス に お い て CenG1A は 、 シ ナ プ ス 前 、 後 細 胞 の 両 方 に 発 現 し て い る こ と が わ か っ て い る (Flybase, Fukui et al., 2012)。そ の た め 、シ ナ プ ス 前 細 胞 に 発 現 し て い る cenG1A 遺 伝 子 、 シ ナ プ ス 後 細 胞 に 発 現 し て い る cenG1A 遺 伝 子 そ れ ぞ れ の シ ナ プ ス 伝 達 に お け る 機 能 を 解 明 す る た め に 、 RNAi 干 渉 法 (RNAi 法 )を 用 い て cenG1A 遺 伝 子 の 発 現 を 組 織 特 異 的 に 抑 制 し た 個 体 で シ ナ プ ス 機 能 解 析 を 行 っ た 。組 織 特 異 的 に cenG1A 遺 伝 子 の 発 現 を 抑 制 す る た め に Gal4-UAS system を 用 い 、 全 神 経 細 胞 で cenG1A 遺 伝 子 の 発 現 を 抑 制 し た 個 体 (31811R-2(X)RNAi×elav-Gal4 (elav-CentRNAi))、全 筋 肉 細 胞 で cenG1A 遺 伝 子 の 発 現 を 抑 制 し た 個 体 (31811R-2(X)RNAi×24B-Gal4 (24B-CentRNAi))、 そ れ ぞ れ に 対 す る 対 照 個 体 (31811R-2(X)RNAi×yw (CentRNAi) 、 31811R-2(X)RNAi×elav (elav) 、 31811R-2(X)RNAi×24B (24B))を 用 い 、 そ れ ぞ れ に つ い て 電 気 生 理 学 的 解 析 と 形 態 学 的 解 析 を 行 っ た 。
2-1, 組 織 特 異 的 に CenG1A発 現 を 抑 制 し た 個 体 の 電 気 生 理 学 的 解 析
22
23
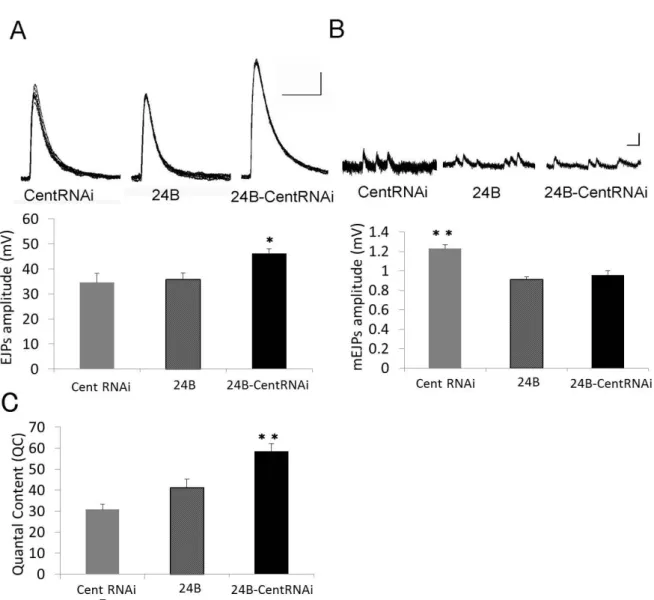
図13. 神経細胞特異的に CenG1A 発現を抑制した個体における電気生理学的解析
24
図14. 筋肉細胞特異的に CenG1A 発現を抑制した個体における電気生理学的解析
26
図15. 組織特異的に CenG1A 発現を抑制した個体における形態学的解析
27
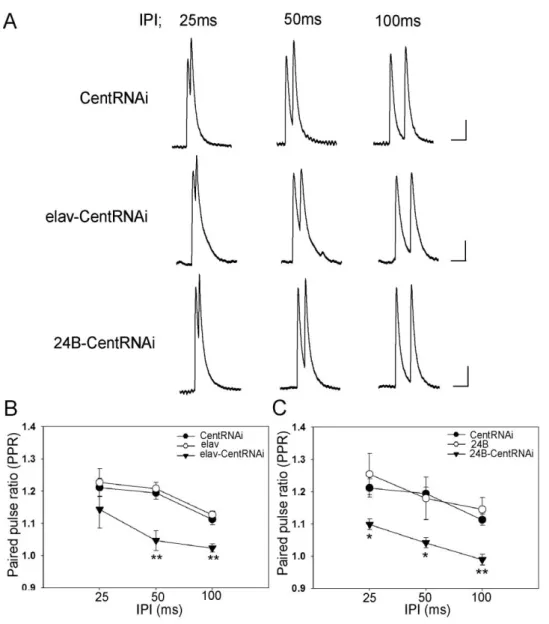
28 2-3, 組 織 特 異 的 に CenG1A発 現 を 抑 制 し た 個 体 に お け る 放 出 確 率 の 解 析 こ れ ま で の 結 果 か ら 、 神 経 細 胞 、 ま た は 筋 肉 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 ど ち ら に お い て も 、 神 経 伝 達 物 質 放 出 量 の 増 加 が お き て い た が 、 シ ナ プ ス の 形 態 に は 影 響 が み ら れ な い こ と が わ か っ た 。 伝 達 物 質 放 出 量 が 増 加 す る 要 因 と し て 、 一 般 に 、 シ ナ プ ス サ イ ト 数 の 増 加 、 放 出 確 率 の 増 加 、 シ ナ プ ス 小 胞 数 の 増 加 な ど が 考 え ら れ る 。 上 述 の nc82 に よ る 抗 体 染 色 に お い て 、 active zone 数 に 差 異 が 見 ら れ な か っ た こ と か ら 、CenG1A 発 現 を 抑 制 し た 個 体 で サ イ ト 数 が 変 化 し て い る 可 能 性 は 低 い 。 そ こ で 残 る 神 経 伝 達 物 質 放 出 量 増 加 の 要 因 で あ る 放 出 確 率 と シ ナ プ ス 小 胞 数 に つ い て 解 析 を お こ な う こ と に し た 。
ま ず 、 放 出 確 率 を 調 べ る た め に 、 Paired pulse stimulation (PPS)を 行 っ た 。 PPS に お い て 一 回 目 の 刺 激 に 対 す る 応 答 と 二 回 目 の 刺 激 に 対 す る 応 答 の 相 対 比 に よ り 算 出 さ れ る Paired pulse ratio (PPR)は Ca2 +依 存 的 な 放 出 確 率 の 指 標 と し て 用 い ら れ て
29 間 隔 50, 100ms の 際 に は 、残 存 Ca2 +
量 の 違 い か ら 、こ の Ca2 +依 存 的 な 何 ら か の 要 因
30
図16. 組織特異的に CenG1A 発現を抑制した個体における放出確率
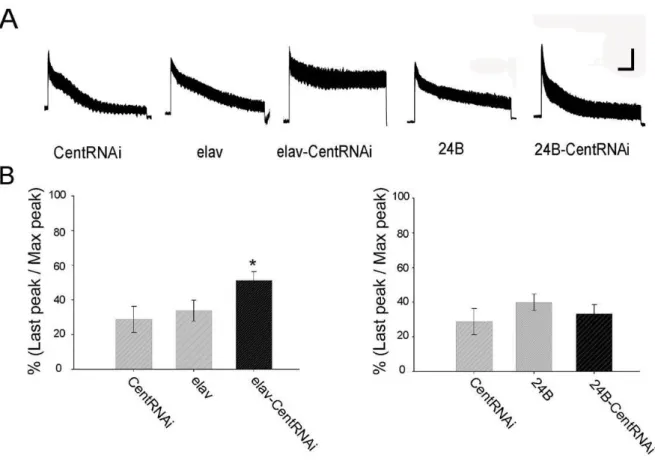
31 2-4, 組 織 特 異 的 に CenG1A発 現 を 抑 制 し た 個 体 に お け る シ ナ プ ス 小 胞 数 の 解 析 神 経 伝 達 物 質 放 出 量 増 加 の 要 因 の う ち 、PPS に よ り 放 出 確 率 が 増 加 し て い る 可 能 性 が 高 い 事 が 確 認 で き た 。 次 に 残 る 要 因 で あ る 、 シ ナ プ ス 小 胞 数 に つ い て 解 析 を お こ な っ た 。 シ ナ プ ス 小 胞 は RP と RRP の 2 つ の プ ー ル に わ け ら れ 、 リ サ イ ク リ ン グ さ れ て い る 。高 頻 度 刺 激 下 で は 、RRP の シ ナ プ ス 小 胞 と 、RPR か ら RP へ と 移 行 し た シ ナ プ ス 小 胞 が 放 出 さ れ る 。RRP と RP の シ ナ プ ス 小 胞 を 放 出 さ せ る た め に 50Hz の 高 頻 度 刺 激 を 行 い 、 応 答 を 測 定 す る こ と で 、 放 出 可 能 シ ナ プ ス 数 に つ い て 解 析 を お こ な っ た 。図 17A の よ う に 50Hz の 刺 激 を 30sec 行 う と 、EJP amplitude が 徐 々 に 減 衰 し て く る 。 こ の 減 衰 は シ ナ プ ス 小 胞 の 枯 渇 を 示 し て い る 。 シ ナ プ ス 小 胞 数 の 増 加 、 ま た は リ サ イ ク リ ン グ 速 度 の 増 加 、 あ る い は こ の 両 方 が お き て い る 個 体 で は 、 こ の 減 衰 度 合 が 弱 く 、 こ れ ら が 減 少 し て い る 個 体 で は 、 減 衰 度 合 が 強 い こ と が 予 想 で き る 。そ こ で 実 験 で は 、刺 激 開 始 時 に 得 ら れ た 最 大 の EJP amplitude か ら 、30sec 後 刺 激 終 了 時 の EJP amplitude が ど れ だ け 減 衰 し た か 、そ の 減 衰 度 合 を % で 算 出 し 、 放 出 可 能 シ ナ プ ス 小 胞 数 の 指 標 と し て 使 用 し た 。
32
図17. 組織特異的に CenG1A 発現を抑制した個体におけるシナプス小胞数の解析
33 IV, 考 察
34
細 胞 で CenG1A を 抑 制 す る こ と に よ っ て 、 高 頻 度 刺 激 に よ る EJP の 減 衰 が 抑 制 さ れ 、放 出 可 能 シ ナ プ ス 小 胞 数 が 増 加 し て い る 可 能 性 が 高 い と い う 結 果 が 得 ら れ た 。 こ の 結 果 か ら 、CenG1A が シ ナ プ ス 小 胞 リ サ イ ク リ ン グ に 関 与 し て い る 可 能 性 が 考 え ら れ る 。
ま た 、elav-CentRNAi で は mEJP amplitude が 対 照 個 体 と 比 較 し て 減 少 し て い る こ と が わ か っ た 。mEJP の 減 少 は 、シ ナ プ ス 後 細 胞 の 感 受 性 、ま た は シ ナ プ ス 小 胞 サ イ ズ の 減 少 に 起 因 す る と 考 え ら れ る が 、 シ ナ プ ス 後 細 胞 の GluRIIA 量 に は 変 化 が み ら れ な か っ た 。そ の た め 、こ の 個 体 で み ら れ た mEJP の 減 少 は 、シ ナ プ ス 後 細 胞 の 感 受 性 の 変 化 に よ る も の で は な く 、 シ ナ プ ス 小 胞 サ イ ズ の 減 少 、 つ ま り シ ナ プ ス 小 胞 内 に 充 填 さ れ て い る 神 経 伝 達 物 質 量 の 減 少 に よ る も の だ と 考 え ら れ る 。 こ の 1 つ の 小 胞 内 の 神 経 伝 達 物 質 量 の 減 少 は CenG1A の 抑 制 に よ り 、 リ サ イ ク リ ン グ が 活 発 に な る こ と に よ っ て 、 充 分 な 量 の 神 経 伝 達 物 質 が 充 填 さ れ ず 、 未 成 熟 な シ ナ プ ス 小 胞 が 放 出 さ れ て い る た め に 生 じ た 可 能 性 が 考 え ら れ る 。
35
2, 筋 肉 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体
36
図18. 組 織 特 異 的 に CenG1A を 抑 制 し た 個 体 に お け る 結 果 の ま と め
左 図 , シ ナ プ ス 前 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 で は 、放 出 確 率 の 増 加 と シ ナ プ ス 小 胞 数 の 増 加 が 生 じ て い る 可 能 性 が 高 い 。 右 図 , シ ナ プ ス 後 細 胞 特 異 的 に CenG1A 発 現 を 抑 制 し た 個 体 で は 、 放 出 確 率 の 増 加 が 生 じ て い る 可 能 性 が 高 い 。
3, negative regulator と し て の CenG1A 機 能
37 図19. CenG1A 機能モデル
39 VI, 引 用 文 献
Aberle, H., Haghighi, A.P., Fetter, R.D., McCabe, B.D., Magalhaes, T.R., and Goodman, C.S., (2002) Wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron, 33, 545 –558.
Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., and Walter, P., (2002) Molecular Biology of The Cell, Garland Science; 4th edition.
Ashery, U., Koch, H., Scheuss, V., Brose, N., and Rettig, J., (1999) A presynaptic role for the ADP ribosylation factor (ARF) -specific GDP/GTP exchange factor msec7 -1. Proc. Natl. Acad. Sci. USA, 96, 1094-1099.
Bellen, H.J., Levis, R.W., Liao, G., He, Y., Carlson, J.W., Tsang, G., Evans -Holm, M., Hiesinger, P.R., Schulze, K.L., Rubin, G.M., Hoskins, R.A., Spradling, A.C. (2004). The BDGP gene disruption project: single transposon insertions associated with 40% of Drosophila genes. Genetics, 167(2), 761 --781.
Beumer, K., Matthies, H.J., Bradshaw, A., and Broadie, K., (2002) Integrins regulate DLG/FAS2 via a CaM kinase II-dependent pathway to mediate synapse elaboration and stabilization during postembryonic development. Development, 129, 3381 –3391.
Brand, A.H., and Perrimon, N., (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Developmen t, 118, 401–415.
Busche, M.A., Eichhoff, G., Adelsberger, H., Abramowski, D., Wiederhold, K.H., Haass, C., Staufenbiel, M., Konnerth, A., and Garaschuk, O., (2008) Clusters of hyperactive neurons near amyloid plaques in a mouse model of Alzheimer’s diseas e. Science, 321, 1686-1689.
Chan, C.B., and Ye, K., (2011) What we have learnt about PIKE from the knockout mice. Int. J. Biochem. Mol. Biol., 2, 228 -239.
40
Cowan, W.M., Südhof, T.C., and Stevens, C.F., (2001) Synapses. The Johns Hopkins University Press; 1st edition.
Davis, G.W., (2006) Home ostatic control of neural activity: from phenomenology to molecular design. Annu. Rev. Neurosci., 29, 307 –323.
Davis, G.W., Schuster, C.M., and Goodman, C.S., (1997) Genetic analysis of the mechanisms controlling target selection: target -derived Fasciclin II regulates the pattern of synapse formation. Neuron, 19, 561 –573
Davis, G.W., and Goodman, C.S., (2002) Genetic analysis of synaptic development and plasticity: homeostatic regulation of synaptic efficacy. Curr Opin Neurobiol., 8, 149–156.
Dietzl, G., Chen, D., Schnorrer, F., Su, K.C., Barinova, Y., Fellner, M., Gasser, B., Kinsey, K., Oppel, S., Scheiblauer, S., Couto, A., Marra, V., Keleman, K., and Dickson, B.J., A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. (2007) Nature. 448, 151 -6.
Fitzsimonds, R.M., and Poo, M.M., (1998) Retrograde signaling in the development and modification of synapses. Physiol. Rev., 78, 143 –170.
Fukui, A., Inaki, M., Tonoe, G., Hamatani, H., Homma, M., Morimoto, T., Aburatani, H., and Nose, A., (2012) Lola regulates glutamate receptor expression at the Drosophila neuromuscular junction. Biology Open, 1, 362 -75.
Gibson, J.R., Bartley, A.F., Hays, S.A., and Huber, K.M., (2008) Imbalance of neocortical excitation and inhibition and alte red UP states reflect network hyperexcitability in the mouse model of fragile X syndrome. J. Neurophysiol., 100, 2615-2626.
Howlett, E., Lin, C.C., Lavery, W., and Stern, M., (2008) A PI3 -kinase-mediated negative feedback regulates neuronal excitability. PLoS Genet., 4, e1000277.
41
Jaworski, J., (2007) ARF6 in the nervous system. Eur. J. Cel l Biol., 86, 513-24.
Kazama, H., Morimoto-Tanifuji, T., and Nose, A., (2003) Postsynaptic activation of calcium/calmodulin-dependent protein kinase II promotes coordinated pre - and postsynaptic maturation of Drosophila neuromuscular junctions. Neuroscien ce, 117, 615-625.
Kazama, H., Nose, A., and Morimoto -Tanifuji, T., (2007) Synaptic components necessary for retrograde signaling triggered by calcium/calmodulin -dependent protein kinase II during synaptogenesis. Neuroscience. 145, 1007 -15.
Keshishian, H., Broadie, K., Chiba, A., and Bate, M., (1996) The drosophila neuromuscular junction: a model system for studying synaptic development and function. Annu. Rev. Neurosci., 19, 545 -575.
Klassen, M.P., Wu, Y.E., Maeder, C.I., Nakae, I., Cueva, J.G., Lehrman, E.K., Tada, M., Gengyo-Ando, K., Wang, G.J., Goodman, M., Mitani, S., Kontani, K., Katada, T. and Shen, K. (2010) An Arf-like small G protein, ARL-8, promotes the axonal transport of presynaptic cargoes by suppressing vesicle aggregation. Neuron, 66, 710 -723.
Kobayashi, H., and Fukuda, M., (2012) Rab35 regulates Arf6 activity through centaurin-β2 (ACAP2) during neurite outgrowth. J. Cell Sci., 125, 2235 -2243.
Krauss, M., Kinuta, M., Wenk, M.R., De Camilli, P., Takei, K. and Haucke, V. (2003) ARF6 stimulates clathrin/AP -2 recruitment to synaptic membranes by activating phosphatidylinositol phosphate kinase type I. J. Cell Biol., 162, 113 -24.
Lawe, D.C., Chawla, A., Merithew, E., Dumas, J., Carrington, W., Fogarty, K., Lifshitz, L., Tuft, R., Lambright, D. , and Corvera, S., (2002) Sequential roles for phosphatidylinositol 3-phosphate and Rab5 in tethering and fusion of early endosomes via their interaction with EEA1. J Biol Chem. 277, 8611 -7.
42
Macleod, G. T., Hegström-Wojtowicz, M., Charlton, M. P., and Atwood H. L., (2002) Fast Calcium Signals in Drosophila Motor Neuron Terminals.. J Neurophysiol 88 , 2659-2663.
Manabe, T., Wyllie, D.J., Perkel, D.J., and Nicoll, R.A., (1993) Modulation of synaptic transmission and long-term potentiation: effects on paired pulse facilitation and EPSC variance in the CA1 region of the hippocampus. J. Neurophysiol., 70 , 1451-1459.
Marqués, G., Bao, H., Haerry, T.E., Shimell, M.J., Duchek, P., Zhang, B., and O’Connor, M.B., (2002) The Drosophila BMP type II receptor wishful thinking regulates neuromuscular synapse morphology and function. Neuron, 33, 529 –543.
Marqués, G., and Zhang, B., (2006) Retrograde signaling that regulates synaptic development and function at the Drosophila neuromuscular junction. Int. Rev. Neurobiol., 75, 267-285.
Maxfield, F.R., and McGraw. T.E., (2004) Endocytic recycling. Nat Rev Mol Cell Bio l. 2, 121-32.
McNaughton, B.L., (1982) Long-term synaptic enhancement and short -term potentiation in rat fascia dentata act through different mechanisms. J. Physiol., 324, 249 -262.
Moore, C.D., Thacker, E.E., Larimore, J., Gaston, D., Underwood, A., Kear ns, B., Patterson, S.I., Jackson, T., Chapleau, C., Pozzo -Miller, L., and Theibert, A., (2007) The neuronal Arf GAP centaurin alpha1 modulates dendritic differentiation. J. Cell Sci., 120, 2683-2693.
Morimoto, T., Nobechi, M., Komatsu, A., Miyakawa, H., a nd Nose, A., (2010) Subunit-specific and homeostatic regulation of glutamate receptor localization by CaMKII in Drosophila neuromuscular junctions. Neuroscience, 165, 1284 –1292.
Müller, M., Pym, E.C., Tong, A., and Davis, G.W., (2011) Rab3 -GAP controls the progression of synaptic homeostasis at a late stage of vesicle release. Neuron, 69, 749-762.
43
Ren, M., Xu, G., Zeng, J., De Lemos -Chiarandini, C., Adesnik, M., and Sabatini, D.D., (1998) Hydrolysis of GTP on rab11 is required for the direct delivery of transferrin from the pericentriolar recycling compartment to the cell surface but not from sorting endosomes. Proc Natl Acad Sci U S A. 95, 6187 -92.
Rizzoli, S.O., & Betz, W.J., (2005) Synaptic vesicle pools. Nat. Rev. Neurosci., 6, 57 -69.
Rizzoli, S.O., and Betz, W.J., (2002) Effects of 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one on synaptic vesicle cycling at the frog neuromuscular junction. J. Neurosci., 22, 10680 -10689.
Sánchez-Soriano, N., and Prokop, A., (2005) The influence of pioneer neurons on a growing motor nerve in Drosophila requires the neural cell adhesion molecule homolog FasciclinII. J Neurosci., 25, 78–87.
Südhof, T.C., (2004) The synaptic vesicle cycle. Annu Rev Neurosci., 27, 509 -47.
Tanaka, K., Imajoh-Ohmi, S., Sawada, T., Shirai, R., Hashimoto, Y., Iwasaki, S., Kaibuchi, K., Kanaho, Y., Shirai, T., Terada, Y., Kimura, K., Nagata, S. and Fukui, Y. (1997). A target of phosphatidylinositol 3,4,5 -trisphosphate with a zinc finger motif similar to that of the ADP -ribosylation-factor GTPase-activating protein and two pleckstrin homology domains. Eur. J. Biochem., 245, 512 -519.
Tao, H.W., and Poo, M., (2001) Retrograde signaling at central synapses. Proc. Natl. Acad. Sci. USA, 98, 11009 –11015.
Uytterhoeven, V., Kuenen, S., Kasprowicz, J., Miskiewicz, K. and Verstreken, P. (2011) Loss of skywalker reveals synaptic endosomes as sorting stations f or synaptic vesicle proteins. Cell, 145, 117 -132.
44
Wucherpfennig, T., Wilsch-Bräuninger, M. and González -Gaitán, M. (2003) Role of Drosophila Rab5 during endosomal trafficking at the synapse and evoked neurotransmitter release. J. Cell Biol., 161, 609 -624.
Yamaguchi, K., Takada, M., Fujimori, K., Tsuchimoto, Y., Kushima, Y., Sanada, M., Fujiwara, T., and Akagawa, K. (1997). Enhancement of synaptic transmission by HPC -1 antibody in the cultured hippocampal neur on. Neuroreport 10, 3641 -4.
Yamamoto-Furusho, J.K., Barnich, N., Xavier, R., Hisamatsu, T. and Podolsky, D.K. (2006) Centaurin beta1 down -regulates nucleotide-binding oligomerization domains 1 - and 2-dependent NF-kappaB activation. J. Biol. Chem., 281, 36 060-36070.
Zhang, Y.Q., Bailey, A.M., Matthies, H.J., Renden, R.B., Smith, M.A., Speese, S.D., Rubin, G.M., and Broadie, K. (2001). Drosophila fragile X -related gene regulates the MAP1B homolog Futsch to control synaptic structure and function. Cell, 107, 591–603.
Zucker, R.S., and Regehr, W.G., (2002) Short -term synaptic plasticity. Annu Rev Physiol. 64, 355-405.