東アジア産マツムシソウ属(マツムシソウ科)の系 統分類学的研究
著者 須山 知香
著者別表示 Suyama Chika
雑誌名 植物地理・分類研究
巻 54
号 2
ページ 105‑126
発行年 2006‑10‑31
URL http://hdl.handle.net/2297/00050274
はじめに
マツムシソウと言えば,一般には高原や高山のお 花畑に群生する様子が思い浮ぶ。ところが,愛知県 東部の丘陵地で出会った 舌状花の無い奇妙なマツ ムシソウ は,標高わずか数十メートルの場所に生 育していた。この様な形態的特徴を示す集団を調べ てみると,周伊勢湾地域の低地やそれに続く丘陵地 に限って分布していたことから,これを新変種ミカ ワ マ ツ ム シ ソ ウS c a b i o s a j a p o n i c a M i q . v a r . breviligulaSuyama & K. Uedaと命名・記載した
(Suyama and Ueda 2005)。このミカワマツムシ ソウを調べた際に,日本のマツムシソウ属植物は,
1950年代以前には分類学的な見解が研究者により 大きく異なっていたにも関わらず,近年は一貫して 日本に一種のみ の 分類に関する問題は無い グ ループであると見なされ,生物学的な分類群の再検 討がなされないままであることを知った。
日本のマツムシソウ属植物は北海道から九州にか けて分布し,その生育地は海岸沿いの岩場,低山お よび丘陵地帯の開けた土地,山地・高原の草地から 亜高山帯の岩場や砂礫地と様々な環境で見られる
(Fig. 1 ; Table 1)。しかし,本植物は明るく開け た場所を好むため,森林が良く発達する日本におけ る本植物の生育地は互いに離れており,非連続的で ある。この分布の特性から,私は「日本のマツムシ ソウには種内分類群間,あるいは地域集団ごとに遺 伝的変異が蓄積されているのでは」という直感を抱 いた。さらには,沿海州,中国東北部,朝鮮半島,
台湾等の近隣地域には日本のマツムシソウ属植物の 近縁種が存在していることから,日本列島の植物相
の成立過程を推測する材料としても本植物群の分類 学的再検討および系統地理学的研究に興味を覚えた。
そこで私は,先ず日本においてマツムシソウ属植 物が認識されてきた歴史をあらため,そこから浮か び上がった諸問題を念頭に,各分類群について実際 の分布および形態的・生態的な形質の再検討等を行 った。さらに分子系統解析により本植物群の類縁関 係を明らかにし,日本および東アジアにおけるマツ ムシソウ属植物の新たな分類学的見解を持つことが できた。本報では日本産マツムシソウ属植物の分類 史,分子系統解析による系統地理学的考察,主に東 アジア産マツムシソウ属植物で観察・発見した新た な形態学的・生態学的情報の報告と,これらを総合 的に評価した結果としての日本産マツムシソウ属植 物の新たな分類体系を提案する。
日本産マツムシソウ属の分類史
マツムシソウ科植物はヨーロッパ,アジア,アフ リカに分布し,世界で10属約320種が知られる一 回繁殖型,あるいは多回繁殖型の草本または低灌木 植物である(Verlaque 1985)。日本にはマツムシ ソウ属Scabiosa L.とナベナ属Dipsacus L. の2 属が分布する。このうちマツムシソウ属はヨーロッ パ,アジア,アフリカに約90種(Verlaque 1985)
が知られている。
日本におけるマツムシソウ属植物の認識の歴史を 遡るため,本草学時代の記述を調べた。明代に李時 珍(1596)の記した「本草綱目」には,従来マツ ムシソウ属の掲載は無いとされてきたが,今回確認 した1930年商務印書館の影印版にある鬱金香(第
920―1192 金沢市角間町 金沢大学大学院自然科学研究科植物自然史研究室
Chika Suyama : Phylogeotaxonomy ofScabiosa(Dipsacaceae)in East Asia
Laboratory for Plant Natural History, Graduate School of Natural Science and Technology, Kanazawa University, Kakuma-machi, Kanazawa 920―1192, Japan
!The Society for the Study of Phytogeography and Taxonomy 2006
2006 年度植物地理・分類学会奨励賞受賞記念論文
須山知香:東アジア産マツムシソウ属(マツムシソウ科)の系統分類学 的研究
105
14巻草部)という名称は,現在チューリップある いはサフランを指すとして一般的に解釈されている が,その図版(第2冊図巻上)にはマツムシソウ 属植物(恐らくS. graminifolia)の特徴を持つ絵 が掲載されている。金陵胡承竜刊本(1596)の本 草綱目附図上巻にある鬱金香の図版では植物の描写 がより簡略であるため,種類の特定は困難である。
この本草綱目は1607年に日本へ渡来したとされて いる。以降,日本の本草学者はこれに記された植物 と日本の植物とを対比させながら,花譜(貝原1694), 大和本草(貝原1709),本草図譜(岩崎1830―1844), 重訂本草綱目啓蒙(小野原著1847;確認は刈米考
註1933),頭註國譯本草綱目(李原著;白井監修・
校註1929―1934)等を編んだが,これらにはマツ
ムシソウ属植物に関する記述は見あたらない。国内 でのマツムシソウの記録は,和漢三才図会(寺島
1713;確認は谷川編集1980)に「玉毬花,松虫草」
として,また花彙(島田・小野1765;確認は奥山
解説1977)に「玉毬花・マツムシサウ・リンボウ
ギク(輪鉾菊)」,救荒本草啓蒙(小野1842)に「山 蘿蔔・マツムシサウ・リンボウギク」,日本産物志
(伊藤1873)に「リンバウキク・マツムシサウ」と
しての記述が見られる。
国外の文献に日本のマツムシソウが登場するのは,
ツュンベリーC. P. Thunbergが1784年に著した F l o r a J a p o n i c aで マ ツ ム シ ソ ウ をS c a b i o s a
maritima L. と記したのが最初である。このS.
maritimaは南フランスのモンペリエから記載され
た植物でヨーロッパ・アフリカに分布するが,現在 マツムシソウとは別節(セイヨウマツムシソウ節:
sect. Cyrtostemma)に分類されている植物である。
その後,日本のフロラに関する初期の文献では,
Amoenitatum Exoticarum Politico-physico- medicarum(Kämpfer 1712)およびFlorae Japoni- cae familiae naturalis(Siebold and Zuccarini 1845―1846)には日本のマツムシソウについての記 述は無い。その後,シーボルトP. F. Sieboldが長 崎の出島で栽培されていた植物を持ち帰ったものを タイプとして,1867年 に は ミ ク エ ルF. A. W.
Miquelが日本固有種S.japonica Miq.を記載した。
その中でマツムシソウは 日本の尾張,上野,美濃 に多く自生し,シーボルトはその和名をRinboogik
(リンボウギク)とした とある。Franchet and Savatier(1875)のEnumeratio Plantarum in Ja- ponia sponte crescentiumでも日本のマツムシソ ウはS. japonica Miq. とされ,日本の文献として
‘Kwa-wi’(花彙),‘Sò Mokou Zousseiz’(草木図説)
を 引 用 し て こ れ の 和 名 を‘Matsu-mousisò
(Tanaka); Rin boo guikou(Sieb.)’と記した。
20世紀に入ると,日本の研究者達がマツムシソ ウを,モンゴル北部,中国東北部,朝鮮半島北部に 分布するとされる幾つかの種と比較して,これらと 同一(Tatewaki 1928, 1940 ; Kitagawa 1939)又 は こ れ ら と 変 種 関 係 に あ る(中 井1909 ; Hara 1937;原1940 a, 1940 b ; Hurusawa 1949, 1951)
と し て 扱 っ た。ま た,マ ツ ム シ ソ ウ の 学 名 に Scabiosa fischeriDCをあてることもあった(Tate- waki 1928 ; Hara 1937)。S. fischeriはde Can-
dolleが1830年にダフリアで採集された種子から
の栽培個体をタイプとして記載したものである。但 し,この学名は現在一般的には,先に発表されたS.
comosa Fisch.のシノニムとして扱われている。に もかかわらず,ロシアの一部の植物学者達は,慣習 として長年この分類群の学名にS. comosaではな くS. fischeriを用いてきた。この経緯の詳細説明 はBobrov(1957)にある。筆者が2001年に調査 を行ったロシア科学アカデミー極東支部植物標本庫 VLAでは,実際にラベルおよびカバーで双方の学 名が混同して用いられていた。1900年代から40 年代までの間,ロシアの研究者がこれらの学名を恣 意的に使用していたことに加え,日本では恐らくロ シア産の標本を検分する機会が不十分であったこと により,S. comosaとS. fischeriは異なる分類群 として認識されていた。例 え ば,原(1940 a)は,
ダフリア産のマツムシソウ属植物は,S. fischeri とは異なるがS. comosaと同一であるという記述 をしており,S. comosaとS. fischeriを別分類群 と認識していることが判る。
この様に,1950年代までには日本産のマツムシ ソウの学名はScabiosa fischeri var. japonica(中 井1909 ; Hara 1937), S. comosa var. japonica
(Tatewaki 1940), S. tschiliensis var. japonica
(Hurusawa 1949),S.japonica(中井1943)の4 通りが提唱されることとなった。また,Hurusawa
(1951)はS. japonicaに2亜種を設定して,日本 のマツムシソウをS.japonicassp. japponica var.
japonicaとした。そして中井(1943)が記載した
S. mansenensis, S. zuikoensisやS. tschiliensis といった大陸産のマツムシソウをマツムシソウの亜 種S. japonica ssp. tschiliensisの下の変種にした うえで,エゾマツムシソウをこれと同亜種に含めて S.japonicassp.tschiliensisvar.acutilobaとした。
1960年代から近年まで,日本の植物相に関する 主な文献(大井1965;北村他1979;北村1981;
奥山1983 ; Yamazaki 1993)では,日本に分布す るマツムシソウ属植物を日本固有の独立種マツムシ ソウScabiosa japonica 1種であるとしてきた。そ してその種内には,形態・生態の差異による変種・
106
㪋㪌㪥
㪊㪌㪥 㪈㪋㪇㪥
Scabiosa jezoensis S. japonica var. japonica S. japonica var. alpina S. japonica var. littoralis S. japonica var. breviligula
A
C D E
B
【エゾマツムシソウ節sectionPrismakena Bobr.】
エゾマツムシソウ Scabiosa jezoensisNakai(Fig. 1 A)
【マツムシソウ節sectionScabiosa】
マツムシソウ S.japonicaMiq. var.japonica(Fig. 1 B)
タカネマツムシソウ S.japonicavar.alpina(Takeda)Takeda(Fig. 1 C)
ソナレマツムシソウ S.japonicavar.littoralis(Nakai)Konta & S. Matsuda(Fig. 1 D)
ミカワマツムシソウ S.japonicavar.breviligulaSuyama & K. Ueda(Fig. 1 E)
Table. 1. Taxonomy and distribution map ofScabiosain Japan.日本産マツムシソウ属植物の分類と分布
A : Habit ofS. jezoensis ; B : S. japonica var.japonica; C : S. japonicavar. alpina; D : S.japonica var.
littoralis; E : S.japonicavar.breviligula.
Fig. 1. Taxonomy and distribution map ofScabiosain Japan.日本産マツムシソウ属植物の分類と分布.
A : Habit ofS.jezoensis; B : S.japonica var.japonica; C : S.japonicavar. alpina; D : S.japonica var.
littoralis; E : S.japonicavar.breviligula.
107
品種が以下のように認められてきた(Fig. 1 ; Table 1):基本品種のマツムシソウS. japonicavar.
japonica f. japonicaは北海道〜九州の主に高原や 開けた山地に生育する。静岡東部および関東地方の 海岸域に見られる矮小個体は記載時以降品種ソナレ マツムシソウvar.japonicaf.littolarisとされてき たが,近年これを変種とする見解が示されている(大 場2003 ; Konta and Matsumoto 2006;植田・須
山2006)。本州・四国の亜高山帯砂礫地には変種(ま
たは品種)のタカネマツムシソウvar. alpinaが生 育する。北海道及び青森北部に生育するエゾマツム シソウS. jezoensis Nakaiは近年までマツムシソ ウの変種var.acutilobaとされてきた。また,本種 の花の色は通常薄紫色であるが,白花の個体が区別 さ れ て シ ロ バ ナ マ ツ ム シ ソ ウv a r . j a p o n i c a f . albiflora(Honda)Haraやシロバナタカネマツム シソウvar.alpina f.albaSugim. として扱われる こともある。そして前述のように,東海地方の低地
・丘陵地から変種ミカワマツムシソウvar. bre- viligulaが記載された(Suyama and Ueda 2005)
(Fig. 1)。
分類の問題点
タカネマツムシソウは一般に 高い所に生えてい るマツムシソウ として捉えられてきた。このこと は,マツムシソウ属植物の分類に関する文献中にお いて,「高山上に見るものはマツムシソウの高山生 一品と見なし,これにタカネマツムシソウの名を与 える」(武田1932),「タカネマツムシソウは丈の低 い高山型」(大井・北川1983),「タカネマツムシソ ウはマツムシソウの高山型で草丈は低いが,花は逆 に大きくなる傾向がある。低地のマツムシソウから 連続的に変異する」(井上1997),等の記述として も示されている。高山植物全般について,「標高の 低いところに生える植物と比べて草丈が低く,大型 の花を付け,葉が肥厚したり密な毛で覆われる等の 特徴を持つことが多いがこれは,厳しい高山の環境 に適応した結果である」との解説を多くの普及書等 で目にする(田辺1961,等)ことから,タカネマ ツムシソウについての前述の解釈は,自然に受け入 れられてきたのであろう。その分類学的位置につい てはマツムシソウの変種である(Scabiosa japonica var. alpina)とする見解(原1940 a ; Hara 1952 ; Hurusawa 1951;大井1965;北村他1979;北村 1981 ; Yamazaki 1993)と,品種(S.japonicavar.
japonicaf.alpina)とする見解(Tatewaki 1940;
中井1943;清水1982)がある。そして国内の普及
書や地域植物誌等を見ても本植物の分類ランクは 様々に扱われている。
また,エゾマツムシソウは近年マツムシソウの変 種とされてきたが,かつてはその分類上の位置付け に大きな議論があった。これをマツムシソウとは異 なる分類群として扱ったのは,Hara(1937)が最 初であった。Hara(1937)は,北海道(日高,胆 振)及び本州北部(陸奥)のマツムシソウ属植物は 大陸と同じタイプであって,北海道から九州に広く 分布するマツムシソウとは異なると認識し,これに Scabiosa fischeri DC(トウマツムシソウ,チョウ センマツムシソウ)の学名を用いた(この学名は前 述の通りS. comosaのシノニムである)。その際S.
fischeriの分布域を上記の他,朝鮮半島(Korea), 中国(China, Manchuria),沿海州(Ussuri),ア ムール地方(Amur)およびダフリア地方(Dahuria)
としている。そして日本に広く分布するマツムシソ ウをこれの変種とした。しかし,原はその後1940 年にダフリア産のS. fischeriの標本を検分し直し た際にその形態は日本産のマツムシソウとは全く異 な る と の 見 解 に 達 し , 日 本 産 の も の を 固 有 種S.
japonicaと し た。そ し て,ダ フ リ ア に 産 す るS.
fischeriは形態がS. japonicaとは大きく異なるが,
それ以外の大陸産マツムシソウはS.japonicaと比 べて葉の先端が尖ることの外に大きな形態的差異は 無いとして,これらを日本産マツムシソウの変種S.
japonicavar.acutiloba Hara(トウマツムシソウ)
として記載した(原1940a, 1940b)。北海道及び本 州北部のマツムシソウ属植物もこのvar. acutiloba に含まれ,タイプには北海道日高郡様似町産の標本 が用いられた。一方,中井(1943)は日本,朝鮮 半島および中国東北部のマツムシソウ属植物の比較 研究を行った際に,北海道に生育するものを独立種 S.jezoensis Nakai(エゾマツムシソウ)とした。
その後Hurusawa(1951)は,エゾマツムシソウ
はS.japonicaよりむしろ中国東北部の五台山から 記載されたS. tschiliensis Grüningとの関係が深 いという見解を示した。その結果,S. japonicaに 2亜種を設定し,日本のマツムシソウをS.japonica ssp. japponicaとする一方で,S. tschiliensis, S.
mansenensis Nakai, S. zuikoensis Nakaiといっ た大陸産のマツムシソウをマツムシソウの亜種ssp.
tschiliensisに纏め,エゾマツムシソウをこれの変 種ssp. tschiliensis var. acutilobaとした。この際,
var. acutilobaの生物学的範疇を,原(1940b)が 記載した北海道,本州北部,朝鮮半島,中国,沿海 州及びアムール地方の広域ではなく,タイプのみに 限った(Hurusawa 1951)。しかしこのHurusawa
(1951)はシノニムリストを挙げたのみの論文であ り,どの様な形質の評価に基づいてこの学名の組み 変えが行われたのかは記されていない。いずれにせ 108
よ,これら中井(1943),Hurusawa(1951)の見 解は広く支持されることはなかった。
以上,日本産マツムシソウ属分類史の詳細を述べ たが,概観するとエゾマツムシソウおよびタカネマ ツムシソウは分類学的定義やランク評価の見解に相 違があったものの,60年代以降に新たな議論がな されてこなかったこと,そして東アジアに分布する 近縁種と日本産種との類縁関係は具体的には未検証 であったことが判明した。
分子系統解析で見る日本産マツムシソウ属植物の系統 近年は手法の発達と簡易化により,伝統的な比較 形態学的手法による類縁性の考察に加えて,遺伝子 解析による生物の系統推定が盛んに行われている。
形態学的手法のみでの系統推定は,例えば比較形質 に収斂・退行が起こっている際には時として誤った 評価を出す恐れがある。しかし,経時変化による中 立的な塩基置換を反映した分子データを用いた系統 解析は,より客観的な系統推定を可能とし,また,
進化速度の速い塩基置換データを得ることにより比 較形質に乏しい近縁な分類群間・個体群間の系統推 定を行うことが出来る。そこで本研究では,日本産 マツムシソウ属植物の種内分類群および大陸産近縁 種の分子系統解析を行った。
1)マツムシソウ科内の系統解析
日本産マツムシソウ属植物の系統地理を考えるた めには,現在の日本に生育するマツムシソウ属植物 の近縁種を調べ出す必要がある。そのためにはまず,
科全体の系統解析を行うことによって東アジアに分 布する近縁種間全体の系統関係を把握し,解析を行 う際に使用する外群にはどの分類群が最適であるか を調べることが必要である。
材料と方法:マツムシソウ科は,これまでのマツ ムシソウ目での系統関係解析の結果から,モリナ科 Morinaceae,トリプロステギア科Triplostegiaceae,
オミナエシ科Valerianaceaeと近縁であることが 示されている(Backlund 1996 ; Bell et al. 2001 ; Donoghue et al. 2003 ; Bell and Donoghue 2003 ;
Bell 2004)。また,モリナ科はマツムシソウ科に,
トリプロステギア科はオミナエシ科に含むとする見 解もある(Cronquist 1988)。モリナ科は中国南部 の標高の高い地域からアジア大陸の中央山脈に沿っ て黒海近辺まで,大陸の東西にまたがって分布し,
Acanthocalyx,Cryptothladia,Morinaの3属が認 められている(Cannon and Cannon 1984)。今回 の解析には,これら各属から1種を用いた。トリ プロステギア科は中国南部から東南アジアにかけて 分布し,Triplostegia1属に2種が認められている
(Backlund and Nilsson 1997)。この両種を解析に 用 い た。オ ミ ナ エ シ 科 は 世 界 に 約8属(Eriksen 1989)約350種(Bell 2004)が知られる,マツム シソウ科とほぼ同規模の大きさの分類群である。こ のオミナエシ科の分子系統解析(Bell 2004, Hi- dalgo et al. 2004)の結果から,マツムシソウクレー ドにより近縁であったPatrinia,Fedia,Nardostachys, Valerianellaの4属から各1種を解析に 用 い た。
マツムシソウ科のサンプルはマツムシソウ属,ナベ ナ 属,Pterocephalus, Bassecoia, Cephalaria, Knautia, Succisa, Succisellaの8属から1〜数種 を用いて,合計4科16属59種のデータを準備し て解析に適したサンプルを選定した。
マツムシソウ属では果実形態により幾つかの節が 認められている。近年マツムシソウ科のモノグラフ を 記 し たVerlaque(1986 a, 1986 b)で は 節 の 認 識の変遷を纏めたうえでsect. Asterothrix Font Quer, sect. Cyrtostemma Mert. & Koch, sect.
Trochocephalus Mert. & Koch, sect. Scabiosa L.
(マツムシソウ節)を採用している。本研究ではこ れらに加えて 東 ア ジ ア 固 有 のsect. Prismakena
Bobr. (エゾマツムシソウ節)を認め,地中海固有
の2種からなるsect. Asterothrix以外の全ての節 が含まれる様にサンプルを準備した。
解析に用いた領域はITS(ITS 1+ITS 2)及びtrnL
intronで,本研究で新たに塩基配列決定を行った
他,国際塩基配列データベースに登録されているデ ータ(Caputo et al. 2004)を合わせて使用した(須 山他2005 ; Suyama and Ueda 2005 b ; Suyama and Ueda未発表)。
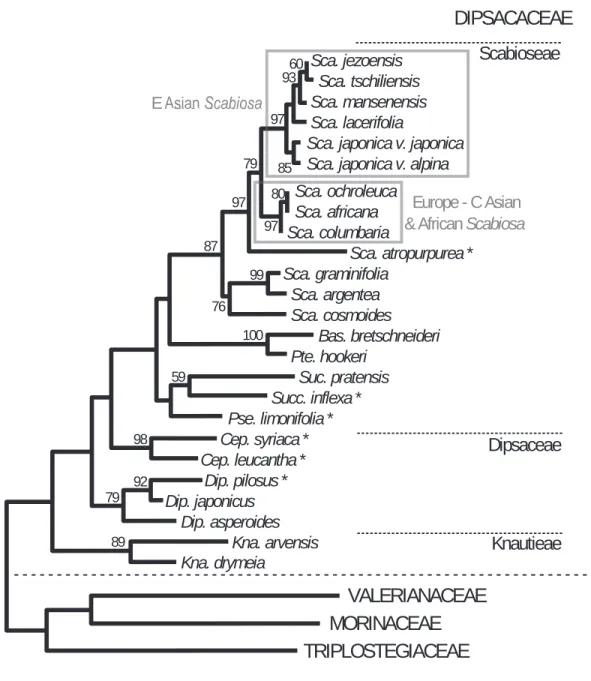
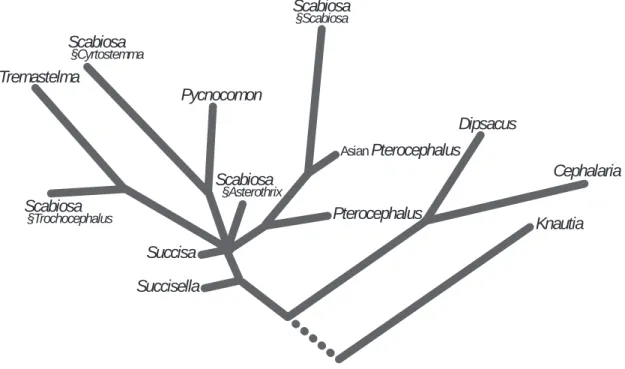
科内系統解析の結果:トリプロステギア科,モリ ナ科,オミナエシ科の3科を外群としたマツムシ ソウ 科 の 系 統 樹 は,Knautia属2種,ナ ベ ナ 属3 種,Cephalaria属2種,Pterocephalus hookeriと Bassecoia bretschneideriが,それぞれ単系統とな っ た(Fig. 2)。ま た,Succisa属 とSuccisella属 は そ の 基 部 にPseudoscabiosa limonifoliaを 併 せ 持つクレードを形成した。これらの系統関係は,従 来の形態学及び細胞分類学的体系(Verlaque 1986 a, 1986 b : Fig. 3)と概ね一致する結果である。
また,マツムシソウ属クレードの中では,エゾマ ツムシソウ節,Trochocephalus節がそれぞれ単系 統となった。マツムシソウ節は全体が一つのクレー ドに纏まり,ヨーロッパ〜中央アジアに分布するS.
columbaria,S.ochroleucaと南アフリカに分布す るS. africanaが単系統となる一方で,マツムシソ ウはエゾマツムシソウ節の姉妹群となった。マツム シソウが分類されるマツムシソウ節は約30種から なり,アジア,ヨーロッパ,アフリカの広域に分布 109
Sca. jezoensis Sca. tschiliensis Sca. mansenensis Sca. lacerifolia
Sca. japonica v. japonica Sca. japonica v. alpina Sca. ochroleuca
Sca. africana Sca. columbaria
Sca. atropurpurea * Sca. graminifolia
Sca. argentea Sca. cosmoides Bas. bretschneideri Pte. hookeri
Suc. pratensis Succ. inflexa * Pse. limonifolia * Cep. syriaca * Cep. leucantha * Dip. pilosus * Dip. japonicus Dip. asperoides Kna. arvensis Kna. drymeia
0.01 substitutions/site bootstrap shown >50%
Scabioseae DIPSACACEAE
VALERIANACEAE TRIPLOSTEGIACEAE
MORINACEAE
Dipsaceae
Knautieae
60
97 93
79 85 80 97 97
99 76 87
100
59
98
79 92
89
E Asian Scabiosa
Europe - C Asian
& African Scabiosa
Fig. 2. Phylogeny of Dipsacaceae infered from nDNA(ITS)and cpDNA(trnL intron), Maximum likelihood tree
(Suyama and Ueda 2005 b, Suyama and Ueda in prep.).核ITS領域および葉緑体trnLイントロン領域を用いた 最尤法によるマツムシソウ科分子系統樹.図中で省略した属名を以下に示す.図中*は国際塩基配列データーベース DDBJ/EMBL/GenBankから得た配列. 登録番号は以下の通り. Abbreviations of genus name in figure : Sca. = Scabiosa, Bas. = Bassecoia, Pte. =Pterocephalus, Suc. =Succisa, Succ. = Succisella, Pse. =Pseudoscabiosa, Cep. =Cephalaria, Dip. =Dipsacus, Kna. =Knautia. * : the sequences from. DDBJ/EMBL/GenBank. Accession numbers are below: Sca. atropurpurea: AY290017, AY290006; Suc. inflexa: AJ426555, AJ426556, AJ427392;Pse. limonifolia: AJ426535, AJ426536, AJ427383; Cep. leucantha: AJ426523, AJ426524, AJ 427376 ; Cep.syriaca: AJ 426525, AJ 426526, AJ 427377 ; Dip.pilosus: AY 290016, AY 290005.
110
Succisella Succisa Scabiosa
§Trochocephalus
Scabiosa
§Asterothrix
Scabiosa
§Cyrtostemma
Pycnocomon
Pterocephalus
Asian
Pterocephalus Scabiosa
§Scabiosa
Dipsacus
Cephalaria Knautia Tremastelma
§Scabiosa §Cyrtostemma
§Prismakena §Asterothrix
§Trochocephalus
Fig. 3. Phylogenetic cladogram of genera in Dipsacaceae based on morphology and cytotaxonomy(Verlaque 1986 b, plate 33, modified). Verlaque 1986 bによる形態および染色体数に基づいたマツムシソウ科における属の系統図
(Verlaque 1986 b図33から作成).
Fig. 4. Distribution map of section inScabiosa.マツムシソウ属の各節の分布域.
111
するマツムシソウ属の中では規模が大きな一群であ る(Verlaque 1986 a)。今 回 の 結 果 か ら,東 ア ジ ア産マツムシソウ属植物はマツムシソウ節から派生 したグループであると考えられる。これは,東アジ ア固有のエゾマツムシソウ節と広域分布群であるマ ツムシソウ節以外の各節がヨーロッパから中近東地 域のみの分布であるというマツムシソウ属各節の分 布に照らし合わせても,矛盾の無い考え方であろう
(Fig. 4)。
以上のことから,日本産マツムシソウ属植物の系 統地理を考察するための系統樹では,マツムシソウ 節を外群とした東アジア産マツムシソウ属の系統樹 を作成するのが適切であると判断した。
2)解析に適した領域の探索
材料と方法:様々な遺伝子および非遺伝子領域を 用いた分子系統解析による分類群間の比較研究の蓄 積により,領域ごとの進化速度の違いや特性などが 明らかになってきた(Soltis et al. 1998)。その中 で,種子植物の種間レベルの系統解析には,核DNA のInternal Transcribed Spacer reagion : ITSや,
葉緑体DNAのtrnL intron,trnL-trnF intergenic region等が頻繁に使用されている(Soltis et al.
1998)。しかし,本研究で行う種内の変種・品種間,
さらには分類群の地域変異という低次分類群間の系 統解析には,より進化速度が速く,低次分類群間の 系統解析に有効であるような突然変異を得ることが 必要である。そこで,この様な領域を効率良く検出 することを狙いとしてPCR-SSCP法により遺伝的 変異に富んだ領域の探索を行った(Suyama and
Ueda 2007印刷中)。解析には遺伝子多型の解析用
に開発された葉緑体ユニバーサルプライマーセット
(Nishizawa and Watano 2000)により増幅される 葉緑体遺伝子のイントロン及び遺伝子間の13領域
(各領 域 長 は 約250―410 bp,各 領 域 の 合 計 は 約4 Kbp)を用いた。
結果:petB intron(petB/1),trnG(UCC)intron
(UCC/1), petD-rpoA intergenic, trnV(UAC)- trnM(CAU)intergenic の領域で,5分類群(マ ツムシソウ,タカネマツムシソウ,ソナレマツムシ ソウ,ミカワマツムシソウ,エゾマツムシソウ)12 サンプル間にバンドパターンの変異が多く見られた。
ま た , こ れ ら の 領 域 で 朝 鮮 半 島 産 のS c a b i o s a mansenensis f.pinnata,および中国東北部産のS.
tschiliensisを比較した所,日本産のどの分類群と も異なるバンドパターンを示した(Suyama and
Ueda 2007印刷中)。このことから,前述の領域は
分子系統解析に適していると判断した。
3)東アジア産マツムシソウ属植物の分子系統解析 材料と方法:東アジアおよび日本におけるマツム シソウ属植物の分子系統解析には,2)により検出 した遺伝的変異に富むcpDNA上のイントロンおよ び遺伝子間領域とnDNAのITSをダイレクトシー ケンス法により遺伝子配列を決定し,1)により選 定された分類群(マツムシソウ節)を外群として解 析を行った(Suyama and Ueda未発表)。
解析試料のサンプリングは,日本のものについて は可能な限り各分類群の分布全域をカバーするよう に行った。また,各採集地ではPCR-SSCP法によ り5個体の試料を比較し,採集地内の個体間には 遺伝的変異が無いことを確認した。
結果:東アジア産マツムシソウ属植物32サンプ ルによる系統解析の結果作成された日本産マツムシ ソウ属植物の系統樹では,エゾマツムシソウ節に分 類 さ れ る エ ゾ マ ツ ム シ ソ ウ お よ び 大 陸 産 の Scabiosa tschiliensis,S.mansenensisと,日本産 マツムシソウ(広義)は,大きく2つのクレード に分かれた(Fig. 5)。日本産マツムシソウ(広義)
の分岐を支持するブートストラップ値は100% と いう高い値となる一方で,エゾマツムシソウ節の分 岐を支持するブートストラップ値は低く,50% 未 満(36.4%)となった。これは,両者の中間的位置 に当たる台湾固有種のニイタカマツムシ ソ ウS.
lacerifoliaとの相関関係によるものと思われる。ニ イタカマツムシソウの2サンプルを含まないデー タセットで試算すると,マツムシソウ節分岐のブー トストラップ値は82% となった。ニイタカマツム シソウは今回の解析ではマツムシソウ(広義)の姉 妹群となったが,そのブートストラップ値は55%
と低い値であり,この関係は弱いものである。マツ ムシソウ(広義)の中で,四国産と本州中部白山産 のタカネマツムシソウはその他の分類群が形成する マツムシソウ種群のクレードの基部に位置する側系 統であり,マツムシソウ種群のクレードよりも早く に分岐したことが示された。また,最尤法の系統樹 では従来マツムシソウS.japonica var.japonica f.
japonica(狭義のマツムシソウ)と同定されてきた 佐渡島産のサンプルが南アルプス産のタカネマツム シソウと同一クレードとなった。ソナレマツムシソ ウ2サンプルとミカワマツムシソウ2サンプルは,
それぞれ単系統となった。マツムシソウ(狭義)は,
本州・四国・九州の10サンプルを解析に用いたが,
地域間の系統性は見出されなかった。
112
0.01 substitutions/site bootstrap shown >50%
ᣣᧄ↥
§Scabiosa
บḧ↥࿕⒳
䊙䉿䊛䉲䉸䉡ᗲ⍮
䊙䉿䊛䉲䉸䉡㐳㊁䋱 䊙䉿䊛䉲䉸䉡ㄭ⇰
䊙䉿䊛䉲䉸䉡ਛ࿖
㩷 㩷 㩷 㩷 䇭䇭䊙䉿䊛䉲䉸䉡ർ㑐᧲
㩷 㩷 㩷 㩷 䇭䇭䊙䉿䊛䉲䉸䉡Ꮊ
㩷 㩷 㩷 㩷 㩷 䇭䇭䇭䇭䊚䉦䊪䊙䉿䊛䉲䉸䉡䋱 㩷 㩷 㩷 㩷 䇭䇭䇭䇭䊚䉦䊪䊙䉿䊛䉲䉸䉡䋲 㩷 㩷 㩷 㩷 㩷 䇭䇭䇭㩷䊚䉦䊪䊙䉿䊛䉲䉸䉡䋳 㩷 㩷 㩷 㩷 䇭䇭䇭䊙䉿䊛䉲䉸䉡྾࿖
㩷 㩷 㩷 㩷 䇭䇭䊙䉿䊛䉲䉸䉡㐳㊁䋲
㩷 㩷 㩷 㩷 㩷 㩷䊙䉿䊛䉲䉸䉡Ꮊ䋲
㩷 㩷 㩷 㩷 㩷 䇭䇭䇭䉸䊅䊧䊙䉿䊛䉲䉸䉡䋱 㩷 㩷 㩷 㩷 㩷 䇭䇭䇭䉸䊅䊧䊙䉿䊛䉲䉸䉡䋲 㩷 㩷 㩷 㩷 䉺䉦䊈䊙䉿䊛䉲䉸䉡ᷰ
㩷 㩷 㩷 㩷 䇭䇭䇭䉺䉦䊈䊙䉿䊛䉲䉸䉡ධർ䉝䊦䊒䉴
㩷 㩷 㩷 㩷 㩷 㩷 㩷 䇭䇭䇭䇭䉺䉦䊈䊙䉿䊛䉲䉸䉡⊕ጊ
㩷 㩷 㩷 㩷 䇭䇭䉺䉦䊈䊙䉿䊛䉲䉸䉡྾࿖
㩷 㩷 㩷 㩷 㩷 䇭䇭㩷䊆䉟䉺䉦䊙䉿䊛䉲䉸䉡䋱
㩷 㩷 㩷 㩷 㩷 㩷 㩷 䇭䇭䇭䊆䉟䉺䉦䊙䉿䊛䉲䉸䉡䋲 㩷 㩷 㩷 㩷 䇭䇭䇭䇭䇭䉣䉹䊙䉿䊛䉲䉸䉡㕍
tschiliensis aff. ᴪᶏᎺᶏጯ 㩷 㩷 㩷 䇭䉣䉹䊙䉿䊛䉲䉸䉡ർᶏ䋱
㩷 㩷 㩷 䇭䇭䇭㩷䉣䉹䊙䉿䊛䉲䉸䉡ർᶏ䋲 mansenensis㖧࿖
tschiliensis aff. ᴪᶏᎺౝ㒽 tschiliensis aff. ᴪᶏᎺౝ㒽䋲
tschiliensisਛ࿖
ᄖ⟲䋺䊣䊷䊨䉾䊌䊶䉝䊐䊥䉦䇭§Scabiosa
᧲ࠕࠫࠕ↥
§Prismakena
東アジアにおける日本産マツムシソウ属植物の系統 に関する考察
今回の解析においてマツムシソウ(狭義)のサン プルは纏まったクレードを形成せず,ミカワマツム シソウ,ソナレマツムシソウのクレードに対して地 域的関連が薄いと思われる様な結果となった。これ は,同一変種内の地域間という非常に近縁な系統関 係の比較を行っているため,系統推定を行うための 共有派生的変異の情報が不足していることによると 思われる。北海道から九州まで広く分布する狭義の マツムシソウの各サンプルの塩基配列のアライメン トデータを見ると,塩基の挿入・欠失や固有派生で ある1塩基の置換が個々に存在する(核と葉緑体 を合わせた全体の約1.7%)一方で,共有派生変異 は非常に少ない(同全体の約0.7%)。アライメン トデータを検分して日本産マツムシソウ種群のデー タから固有派生変異を除くと,マツムシソウ(狭義)
10サンプル中の4サンプルは同じ配列となる。こ
の様に日本産マツムシソウ属植物内において系統樹 の分解能が低い一方で,東アジア全体の中での比較 からはマツムシソウの属するマツムシソウ節とエゾ マツムシソウの属するエゾマツムシソウ節の間には 興味深い系統関係が判明した。現時点で示された分 子系統樹をもとに,日本産マツムシソウ属植物の自 然史を次に述べる。
エゾマツムシソウはマツムシソウの変種ではなく 別節の独立種:分子系統解析の結果,エゾマツムシ ソウはロシア沿海州の海岸地域で採集したサンプル と単系統になり,実は大陸に起源を持つ 東アジア でメジャーなグループの一員 であることが示唆さ れた。従来のエゾマツムシソウとマツムシソウの識 別点である葉裂片先端や被萼冠状部裂片の形態には 一見両者の中間型が多く,各地の標本庫では誤同定 がしばしば見受けられた。しかし分子データを基に 両者の違いを詳細に再検討した所,両者の果実形態 には明らかに差異があり,別節に分類されることを
Fig. 5. Phylogeny of east AsianScabiosainfered from nDNA(ITS)and cpDNA [the petB intron, the trnG(UCC)
intron, the petD-rpoA intergenic spacer, the trnV(UAC)-trnM(CAU)]. Maximum likelihood tree(Suyama and Ueda 2005 b, Suyama and Ueda in prep.).核ITS領域および葉緑体イントロン・遺伝子間領域を用いた最尤法によ る東アジア産マツムシソウ属植物の系統樹.図中で省略した種名を以下に示す. Abbreviations of sample name in fig- ure : tschiliensis =Scabiosa tschiliensis, tschiliensis aff. = a plant akin toS. tschiliensis, mansenensis =S. mansenensisf.pinnata.
113
発見した。また,マツムシソウ(広義)は一回繁殖 型多年草であるのに対してエゾマツムシソウは多回 繁殖型多年草であることや,マツムシソウ(広義)
は冬季をロゼットで過ごすのに対してエゾマツムシ ソウは越冬芽を形成するという生態的特徴を新たに 発見した。以上のことからエゾマツムシソウをマツ ムシソウとは異なる別節の独立種と認め,学名は Scabiosa jezoensis Nakaiが正名となる(Suyama and Ueda 2007印刷中)。
また,エゾマツムシソウとマツムシソウ(狭義)
は,両者を明確に区別できる形態特徴が指摘されて おらず,国内外の植物標本庫各所においてエゾマツ ムシソウとマツムシソウ(狭義)の間の誤同定が数 多く見られた。これに関連してマツムシソウ(狭義)
が北海道に分布する(Hara 1952;大井1965;北 村他1979;北村1981;奥 山1983)か,分 布 し な い(中井1943 ; Hurusawa 1951 ; Yamazaki 1993)
かについても長年意見の相違が見られた。しかし,
本研究により,果実の被萼の形態で両者を明確に区 別することが出来るようになった。改めて北海道で 採集された標本を調査したところ,その大半はエゾ マツムシソウであったが,確かにマツムシソウ(狭 義)と同定できる標本を少数ながら確認したことか ら,マツムシソウ(狭義)は北海道まで分布するこ とが明らかとなった(Japan, Hokkaido : Prov.
Ishikari, Sapporo, S. Tamaki(TI); Hakodate, Akamura, Oct. 1915, Greatrex(SAPS); 29 July.
1890, Faurie 5766(K); between Yunokawa to Akamura, 4 Sept. 1916, Greatrex(SAPS))。ただ し,いずれの標本も採集時代が古い(1890―1916 年)ため,マツムシソウが北海道に現存しているか どうかは,再調査が必要である。
タカネマツムシソウはマツムシソウが 登山 し たものではない:従来一般的に,タカネマツムシソ ウは マツムシソウの高山型 であると理解されて きた。しかし,分子系統解析(Fig. 5)で示された
「日本の高山帯には,マツムシソウより分岐年代が 古く,かつ系統の異なるタカネマツムシソウが個々 に生育している」という事実と,林の中や高茎草本 群落中には分布出来ないという本植物の生育特性か ら,タカネマツムシソウはマツムシソウよりも先に 日本へ進出してきた,系統の異なるグループである と考える。この知見は「タカネマツムシソウとは,
高所へ分布拡大してその環境に適応したマツムシソ ウである」との漠然とした考えを大きく覆すもので ある。
タカネマツムシソウは,本州・四国の高山帯に生 育し,果実の萼刺が4 mm以上の長さであること により他の分類群から区別されてきた(Yamazaki
1993)。従来マツムシソウと同定されつつも疑問視
されることもあった佐渡島の金北山一帯に分布する 集団は,分子系統解析ではマツムシソウよりもタカ ネマツムシソウに類縁性が高いという結果となった。
そこで,改めて標本観察および現地調査を行った所,
佐渡のマツムシソウは萼刺が3 mm程度の個体が 多いが,地上高が低く,茎は短く,葉に光沢が有り やや厚い等の形態特徴がタカネマツムシソウと類似 しており,大佐渡山地上部の稜線にある裸地にのみ 生育することが判った。タカネマツムシソウは本州
では2000 mを越す山岳地に見られる 高山植物
としての知名度が高く,四国の東赤石山等の産地は
「1700 m程度の低い場所」であることが注目され てきた。しかし今回の再検討により 佐渡島では標 高が更に低い,僅か1000 mでタカネマツムシソウ が存在している という新たな事実を認識すること が出来た。
ソナレマツムシソウは単なる海岸生品種ではな い:ソナレマツムシソウは中井(1943)の記載時 より一貫して 海岸で矮小化したマツムシソウ で あり,マツムシソウの品種として捉えられていた。
今回の分子系統解析では2産地のソナレマツムシ ソウが,他のマツムシソウ種群とは異なる遺伝的変 異を共有していた。現地調査および標本の再検討を 行った所,海岸から離れた山中でも短い茎・肥厚し た葉・先の丸い葉裂片といったソナレマツムシソウ の形態特徴を持つ集団が多く存在することが判明し,
神奈川県・千葉県においてこれまでマツムシソウと 思われていた集団の多く,および静岡県東部の山地 帯に生育するものの一部はソナレマツムシソウであ ることを発見した。ソナレマツムシソウは種子から の肥育栽培を行っても,形態特徴に変化は見られな かった。また,栽培条件によっては多回繁殖を行う 個体もあることを確認している。以上の特性から,
ソナレマツムシソウはマツムシソウの単なる品種で はなく,少なくとも変種とするべきであろう。
マツムシソウ属植物は,日本へ南から北からやっ てきた:今回比較した東アジア産マツムシソウ属植 物の中で,日本産マツムシソウ(広義)に最も近縁 で あ っ た の は 台 湾 産 の ニ イ タ カ マ ツ ム シ ソ ウ Scabiosa lacerifolia Hayataであった。本種は台 湾 の 標 高2000―3600 mの 山 地 に 固 有 な 種 で(Li
1998),長年マツムシソウ属の一員とされてきたが,
近年の研究においては幾つかの異なる見解が示され て い る。Verlaque(1986 a)は こ の 種 をSuccisa 属とした(但し,組み替えの記載が命名規約に則っ た正しいものではなかったため,ここで行った組み 替えは無効である)。しかし,アジア産マツムシソ ウ属植物の一部を研究したBurtt(1999)はニイタ 114
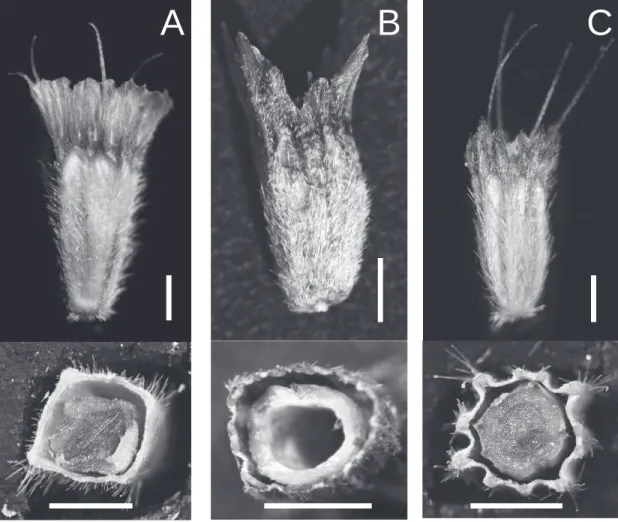
A B C
カマツムシソウについて,「形態はSuccisa属には 当てはまらず,マツムシソウ属で良いが,マツムシ ソウ属植物の中でも形態が特異であることから,詳 細研究後の結果によっては新たな節に分ける必要性 があるかも知れない」としている。その後,東アジ ア 産 の 数 種 の マ ツ ム シ ソ ウ 科 植 物 を 観 察 し て Pterocephalodes属を新たに記載したMayer and Ehrendorfer(2000)は,ニイタカマツムシソウを このPterocephalodes属に含めた。しかしこの属は Burtt(1999)が記載した新属Bassecoiaとタイプ 種(バシオニム:Scabiosa bretschneideriBatalin)
が同一であることから,命名規約上では非合法名と なる。この様に分類議論の的となっているニイタカ マツムシソウは,マツムシソウ節とエゾマツムシソ ウ節の中間的な被萼の形態を持っている(Fig. 6)。 分子系統解析においても,マツムシソウ科の解析
(Fig. 2)ではエゾマツムシソウ節,東アジア産マツ ムシソウ属の解析(Fig. 5)ではマツムシソウ節の,
それぞれのクレード基部に位置する結果になったこ とから,ニイタカマツムシソウは東アジア地域のマ ツムシソウの種分化を考える際の鍵となる種であろう。
この分子系統解析の結果と植物の分布様式を考え 合わせて,マツムシソウ属植物が過去において日本 へ進出した概略は,現在の所,以下の様に考えてい る。マツムシソウ(広義)は昔の台湾から中国南部 にかけての,日本列島より南方の地域でマツムシソ ウ節から分化した。その中でタカネマツムシソウは 比較的古いグループであり,複数回にわたって日本 へ分布を拡大した一方で,狭義のマツムシソウは最 も遅く日本へ侵入し急速に列島を北上して分布を広 げた。また,エゾマツムシソウは朝鮮半島北部から 中国東北部,沿海州地域にかけての地域に生育して いたエゾマツムシソウ節の一群から派生し,現在の 北海道・青森北部に伝播した集団である。エゾマツ ムシソウ節とマツムシソウ節は,マツムシソウ科の 分子系統樹上において互いに姉妹群となって最末端
Fig. 6. Fruits morphology ofScabiosa plants.マツムシソウ属植物の果実形態. A:エゾマツムシソウ;B:ニイタカ マツムシソウ;C:マツムシソウ.上段:側面;下段:横断面.図中スケールバーは1 mm. A : S.jezoensis; B : S. lacerifolia; C : S.japonica. Upper : lateral view ; lower : transection. Scale bars = 1 mm.
115
B A
に位置しており,系統樹からは分化の順番がどちら が先であるかの判断は出来ない。しかし,両者の分 布を見ると,マツムシソウ節はヨーロッパからアジ ア大陸を横断して東アジア、および中央アジアから アフリカ大陸の南端まで達する広域分布グループで あるのに対して,エゾマツムシソウ節の分布は極東 アジアに限られていることから,恐らく後者は前者 から派生して生じた一群と考えている。ここで,東 アジアでは日本のマツムシソウ(広義)が唯一のマ ツムシソウ節植物であり,他は全てエゾマツムシソ ウ節であることが興味深い。
日本産マツムシソウ属植物 分類ノート 日本産マツムシソウ属植物の分類研究に際しての 新知見,および特筆事項を記す。
マツムシソウの生活史:マツムシソウ(広義)は,
一般に二年草(大井1965 ; Yamazaki 1993;清水 1994)あるいは越年草(北村他1979;北村1981;
奥山1983)とされている。しかし,2年以上生育
する個体の存在を中井(1943)が指摘し,また清 水(1982)はタカネマツムシソウを「越年草また は一稔性の多年草」としている。私は日本産マツム シソウ属植物の生育を確認するために,金沢大学植 物園において2000年から6年以上継続して栽培実 験を行っている。観察しているのは,1)野外に自 生する若い個体を採取し,植木鉢あるいはプランタ
ーで栽培したもの,2)野外に自生する個体に結実 していた種子を播種育成したもの,3)前述1)2)
により植物園で栽培した個体に結実した種子を播種 育成したものである。
この結果,上記1)〜3)の全ての栽培条件にお いて,マツムシソウ(広義)ではロゼットのままで 2年以上生育する個体が多数観察された。このよう な個体の観察をさらに続けたところ,開花・結実し た個体はその後全て枯死した。特に十分な養分を与 えずに育てると年数を経ても植物体は小さいままで,
開花することができない。しかし,野外観察では,
花茎を付けた花期の途中で花茎を切除されたり,踏 みつけや草刈り等に遭い地上部が大きく損傷した様 な場合には,開花個体が次年まで生育を続けること もあった。また,ソナレマツムシソウは金沢で栽培 している個体は開花後全て枯死しているが,自生地 では大株となって数年にわたり多数の花を付ける個 体も見られる(Fig. 7 A)。これらのことからマツ ムシソウ(広義)は基本的には一回繁殖型の多年生 草本(monocarpic perennial herb)であり,開花 に至る年数は生育条件によって変化するということ がわかった。一方,エゾマツムシソウは開花結実し た個体のほぼ全てが枯死しなかったことから,多回 繁殖型の多年草(polycarpic perennial herb)であ ることが判明した。また,エゾマツムシソウは地上 部付近の地下茎から新たなシュートが生じて株は多
Fig. 7. Large individuals ofScabiosa japonicavar.littoralis(A)andS.jezoensis(B). Both having multi-headed root(shown by arrowhead in B).地下茎が多頭性を示すソナレマツムシソウ(A)およびエゾマツムシソウ(B)の 大型個体.どちらも地下茎が多分岐している(B中に矢印で示す).
116
1 2 2
3 3
4 4
5
5 6 6
7 7
A B
頭性を示し,生育年数の長い個体は花茎が叢生する
(Fig. 7 B)。これに対してマツムシソウ(広義)で は根茎が多頭になることはないため,一個体の花茎 は一本のみである。エゾマツムシソウの生育型の特 徴は中井(1943)も記述していたが,その後の文 献や図鑑類では全く言及されていない(大井1965;
北 村 他1979;北 村1981;奥 山1983 ; Yamazaki 1993;清水1994).
また,通年栽培による観察の結果,マツムシソウ
(広義)とエゾマツムシソウは越冬状態に相違があ ることを新たに確認した。マツムシソウ(広義)は,
種子から発芽した後,ロゼットで生長を続け,成熟 すると花茎を形成して開花後に枯死する。花期は夏
〜秋である。開花しなかった未成熟個体はロゼット のまま越冬する(Fig. 8 A―C)。しかし,エゾマツ ムシソウは,花期はマツムシソウ(広義)と同様に 夏〜秋であるが,越冬前の晩秋には,非開花個体・
開花個体共に地上部が枯死して越冬芽を形成するこ とが新たに判明した(Fig. 8 D)。越冬芽による越 冬はエゾマツムシソウだけに見られた。このことは,
金沢大学植物園において3年間継続して観察され た。またこのエゾマツムシソウの越冬芽による越冬 は,北海道様似郡様似町の自生地においても確認さ れている(田中正人氏,私信)。
種子は秋に地上に散布される。成熟種子を採り播 きしてその発芽はターンを観察すると,すぐに発芽 するものと翌年の春に発芽するものが混在している。
さらに長期にわたって休眠するものがあるようであ る。林を切り開いて草原に戻した所,近隣に分布し ていないにも関わらずマツムシソウが出現したとい う事例をしばしば聞く。ここで,本植物の種子は比 較的大型な痩果であるため種子散布は自然落下であ り,鳥や風などによる長距離分散は無いと考えられ ていることから(Ulbrich 1928 ; Ehrendorfer 1964 a, 1964 b ; Verlaque 1986 b ; Mayer 1995),マツ ムシソウの種子は比較的長期の休眠が可能であると 思われる。
花序の形態と開花順:前述の様にマツムシソウ属 植物は十分に成長した個体は夏〜秋にかけて花茎を 形成する。日本のマツムシソウは通常1個体に1 つ,エゾマツムシソウ(稀にソナレマツムシソウ)
は根茎の地上部が多頭となるため,1から複数の花 茎をつける。花茎は節で二出分枝し,複数の頭花を つける(Fig. 9 A)。一個体に複数の花を持つ花序 を形成する植物のほとんどは,有限花序か無限花序 のいずれかである。仮軸分枝によって形成される集 散花序の形態を持つものは,一般に花は頂花から下 へ向かって咲き進む有限花序(遠心性花序)であり,
単軸分枝で形成される総房総状花序の形態を持つも のは,花茎の下から上へと先進む無限花序(求心性 花序)である(Gifford and Foster 1989;長谷部
他監訳2002)。マツムシソウ属では1個体が形成
する花序全体中での頭花間の開花順は一定であり,
Fig. 9. Terminate inflorescence ofScabiosaplants and its order to open. A : S.japonicavar.alpina; B : opening order of heads in terminate inflorescence. マツムシソウ属植物の花序とその頭花の開花順.A:タカネマツムシソ ウ;B:花序中における頭花の開花順.花序全体は有限的に開花する.
117
A
C D
B
A B
C
Fig. 8. Wintering form ofScabiosaplants in Japan. A-C : S.japonica(A : var.japonica; B : var. alpina; C : var.littoralis), D : S. jezoensis. 日本産マツムシソウ属植物の越冬形態.A:マツムシソウ;B:タカネマツムシソ ウ;C:ソナレマツムシソウ;D:エゾマツムシソウ.
Fig. 10. Flowering stages of the head of dichogamousScabiosa japonica. A : opening stage(photo shows the case that outer ligules and inner florets in a head opening at the same time);B : male stage ; C : female stage
subsequent to male stage. マツムシソウ頭花の開花段階と雌雄異熟性.A:開花開始期(写真は頭花外側の舌状
花と中心部の小花が同時に咲き始めた場合);B:雄性期;C:雄性期に続く雌性期.
118
A B C
Fig. 9 Bに示す通り,分岐順の早い頭花から遠心的
に開花していく。1個体の開花期間は,付ける頭花 の数により異なるが,2―3週間から,長いものでは
1―1.5ヶ月程である。また,イギリス王立キュー植
物園の野外圃場で栽培されているヨーロッパ,アフ リカのマツムシソウ属植物約20種(Scabiosa afri- cana, S. argentea, S. atropurpurea, S. canes- cens,S.columbaria,S.cosmoides,S.graminifo- lia, S. incisa, S. lucida, S. maritima, S. mi- noana, S. ochroleuca, S. prolifera,S. si- lenifolia,S.songorica,S. speciosa, S. triandra 他)を2002年5月から2003年1月の期間に観察 した結果,花期は日本のものと同様に夏から秋であ った。
マツムシソウの頭花は多数の小花からなり,中心 の小花は筒状花,周辺の小花は,花冠裂片の一部が 長く伸びた舌状花となる。マツムシソウの頭花にお ける小花の開花順を見ると,周辺の舌状花から開花 が始まり中心へと咲き進むことが多いが,頭花の中 心と周辺の舌状花が同時に咲き出すのも多数観察さ れる(Fig. 10 A)。このことは,マツムシソウの花 茎全体が集散花序であって,花茎主軸からの分岐順 の早いものから遅いものへと遠心的に開花していく 一方で,頭花内の小花の開花順は,頭花ごとの開花 順と同様であるならば中心から周囲へと遠心的に咲 き進むべきところを,頭花周辺の舌状花がより早く 開花していると捉えることができる。また,マツム シソウ科ナベナ属では花茎全体はマツムシソウ属同 様に集散花序である。中でもナベナ節植物の頭花は マツムシソウ属植物と比べて1頭花あたりの小花 数がより多く,円錐形で大型となり(Fig. 11 A), 頭花周辺の小花は舌状花とはならない。ナベナ属植
物では頭花間の開花順はマツムシソウ属と同様に遠 心的である一方で,観察例は少数であるが,頭花内 での小花の開花順がFig. 11 B, Cに示した様々な 咲き方をするうえに,個体内で一定していない場合 も観察している。
雄性先熟と自家和合性:マツムシソウ属植物の筒 状花・舌状花は共に両性花であり,どちらも良く結 実する。頭花内の小花が開花していく様子を観察し た結果,この属の植物は明瞭な雄性先熟であること を確認した。則ち,小花は開花時に先ず雄蕊を花冠 より長く伸張させ,葯が裂開して花粉が散布された 後に葯が花糸より脱離して花糸が萎れる。一方,雌 蕊は葯が脱離する頃までは花冠の内部にあるが,そ の後急激に花冠の外へ伸張する(Fig. 10 B, C)。 それぞれの開花ステージの正確な時間数のデータを 測定してはいないが,1頭花中の最初の小花が開き 始めてから全ての小花が咲き揃うまでに2―3日か かり,咲き始めから雄性期の終了までが4―5日間 あり,その後雌性期が数日間続いた後,柱頭と花柱 が萎れていく。この進行状況は,気温や日照条件に 大きく左右される。先に述べた様に1個体には複 数の頭花が付き,それらの開花時期には差があるた めに,結果として1個体に未開花で蕾の頭花,雄 性期の頭花,雌性期の頭花,および結実した頭花が 混在する時期が長い。
また,日本産マツムシソウにおける自家不和合性 の有無を調べる目的で人工授粉実験を2000年に行 った。訪花昆虫を避けるために頭花全体を薄いパラ フィン紙で被い,条件を変えた以下の受粉実験を行 った。
Fig. 11. Head ofDipsacus fullonum(A), and diagrams of florets opening in a headof Dipsacusplants(B, C). Two different opening sequences were observed. オニナベナの頭花(A),およびナベナ節植物の頭花における小花の開 花順(B, C).異なる二通りが見られる.
119