博士学位論文
植物の低温保存における 微小電流を用いた
ダメージ低減に関する研究
平成 28 ( 2016 )年 12 月
神奈川工科大学
大学院工学研究科機械工学専攻
蜷川 貴子
i 目次
記号表
第1章 はじめに ... 1
1.1 本研究の背景... 1
1.2 低温保存技術について ... 5
1.2.1 食品品質が低下する要因への低温の影響 ... 5
1.2.2 緩慢冷却による細胞の凍結挙動 ... 6
1.2.3 急速冷却による細胞の凍結挙動 ... 7
1.2.4 細胞内のガラス化 ... 7
1.3 植物細胞と動物細胞 ... 9
1.4 凍結・解凍処理後に起こる食品素材の品質の低下 ... 11
1.5 低温環境と植物... 12
1.6 これまでの低温保存技術に関する研究 ... 13
1.6.1 細胞内凍結に関する研究 ... 13
1.6.2 植物の低温保存に関する研究 ... 14
1.6.3 食品中に形成される氷結晶を制御する凍結保存技術の研究 ... 15
1.7 本研究の目的と特徴 ... 16
1.8 本論文の構成... 17
第2章 高速度カメラを用いた細胞内氷結晶形成挙動の評価 ... 19
2.1 緒言 ... 19
2.2 実験装置および実験方法 ... 20
2.2.1 低温顕微鏡システム ... 20
2.2.2 試料 ... 20
2.2.3 試料への電流負荷方法 ... 23
2.2.4 実験方法... 23
2.3 Tukey-Kramer法による取得データの検定について ... 26
2.4 細胞外および細胞内凍結発生時の温度 ... 26
2.4.1 細胞外および細胞内凍結発生時の温度の定義 ... 26
2.4.2 電流負荷しない場合 ... 28
2.4.3 電流負荷した場合 ... 28
ii
2.5 高速度カメラで捉えた細胞内の氷結晶形成過程 ... 31
2.6 細胞内氷結晶の出現位置 ... 34
2.6.1 細胞内氷結晶の出現位置のパターン定義 ... 34
2.6.2 電流負荷しない場合 ... 36
2.6.3 電流負荷した場合 ... 36
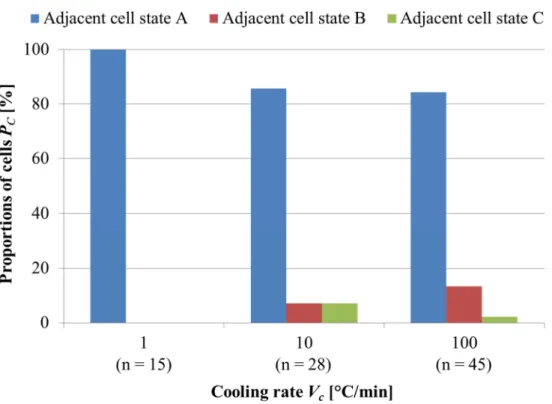
2.7 細胞内氷結晶の出現位置に隣接する細胞の様子 ... 38
2.7.1 細胞内氷結晶の出現位置に隣接する細胞の様子について ... 38
2.7.2 電流負荷しない場合 ... 40
2.7.3 電流負荷した場合 ... 40
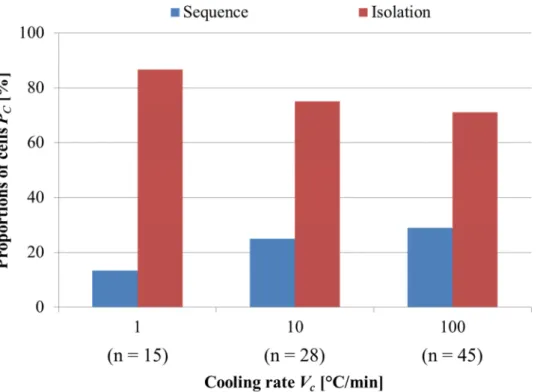
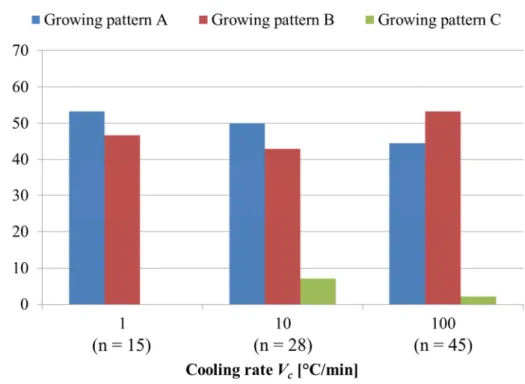
2.8 細胞内凍結の発生パターン ... 42
2.8.1 細胞内凍結の発生パターンの定義 ... 42
2.8.2 電流負荷しない場合 ... 43
2.8.3 電流負荷した場合 ... 43
2.9 細胞内氷結晶の成長パターン ... 45
2.9.1 細胞内氷結晶の成長パターンの定義 ... 45
2.9.2 電流負荷しない場合 ... 45
2.9.3 電流負荷した場合 ... 46
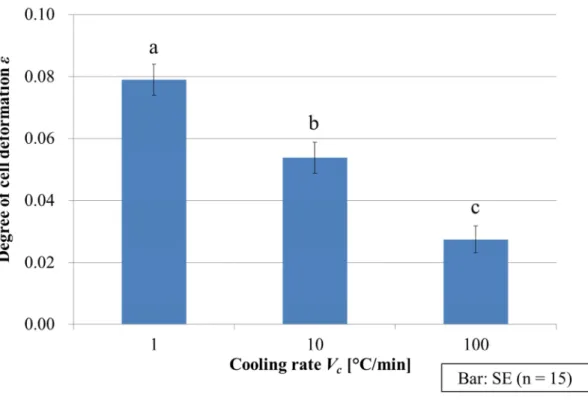
2.10 細胞の変形度 ... 49
2.10.1 細胞の変形度の評価方法 ... 49
2.10.2 電流負荷しない場合 ... 51
2.10.3 電流負荷した場合 ... 51
2.11 細胞内氷結晶の成長速度 ... 54
2.11.1 細胞内氷結晶の成長速度の評価方法 ... 54
2.11.2 電流負荷しない場合 ... 54
2.11.3 電流負荷した場合 ... 56
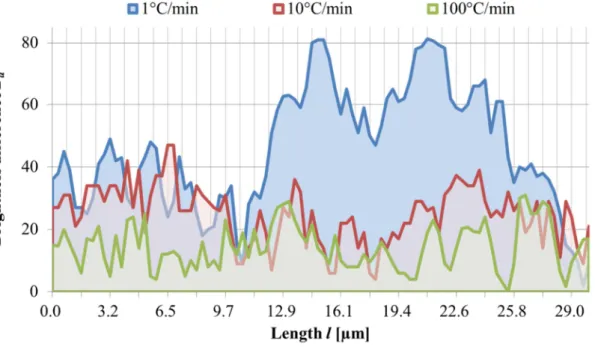
2.12 細胞内に形成される氷結晶の輪郭 ... 58
2.12.1 細胞内に形成される氷結晶の輪郭の評価方法 ... 58
2.12.2 電流負荷しない場合 ... 62
2.12.3 電流負荷した場合 ... 66
2.13 本章で得られた結果および知見に基づく考察 ... 70
2.13.1 細胞内凍結の発生について ... 70
2.13.2 電流負荷による細胞内の氷結晶形成挙動の変化について ... 73
iii
2.14 結言 ... 76
第3章 冷却の進行を伴う植物細胞内pHの推移 ... 79
3.1 緒言 ... 79
3.2 物質量濃度(モル濃度)の単位表記について ... 80
3.3 蛍光によるpH計測のメカニズムについて ... 80
3.4 実験装置および実験方法 ... 84
3.4.1 低温顕微鏡システム ... 84
3.4.2 試料 ... 84
3.4.3 試料への蛍光試薬の導入方法 ... 88
3.4.4 試料への電流負荷方法 ... 92
3.4.5 蛍光画像の撮影方法 ... 92
3.4.6 実験方法... 92
3.5 蛍光強度比の算出方法の検討 ... 95
3.5.1 蛍光強度比の算出方法について ... 95
3.5.2 蛍光画像の輝度から蛍光強度比を算出 ... 95
3.5.3 レシオ画像を用いた蛍光強度比の算出 ... 99
3.5.4 蛍光強度比の算出方法 ... 101
3.6 pH検量線の作成 ... 108
3.6.1 pH検量線の作成方法 ... 108
3.6.2 作成したFDAおよびBCECF-AMのpH検量線 ... 110
3.6.3 作成したCDCFDAのpH検量線 ... 110
3.7 蛍光によるpH計測の検証 ... 113
3.7.1 0°C下にあるpH標準液のpH計測 ... 113
3.7.2 16°C一定温度下の細胞内pHの推移 ... 113
3.8 植物組織の冷却過程における細胞内pH推移 ... 115
3.8.1 電流負荷しない場合 ... 115
3.8.2 電流負荷した場合 ... 120
3.9 電流負荷による細胞内pHの変化についての考察 ... 127
3.10 結言 ... 129
iv
第4章 実用的見地からの電流負荷効果の検討 ... 131
4.1 緒言 ... 131
4.2 電流負荷による細胞内過冷却の持続性 ... 131
4.2.1 細胞内過冷却の持続性の評価について ... 131
4.2.2 実験装置および実験方法 ... 134
4.2.3 細胞内過冷却の持続時間 ... 134
4.3 0°C保存下の細胞活性への電流負荷効果の持続性 ... 137
4.3.1 0°C保存下の細胞活性の評価について ... 137
4.3.2 実験装置および実験方法 ... 138
4.3.3 細胞活性の評価方法 ... 140
4.3.4 0°C保存下の細胞活性の推移 ... 142
4.4 葉への電流負荷効果 ... 151
4.4.1 葉への電流負荷効果の評価について ... 151
4.4.2 重量減少率の定義 ... 151
4.4.3 実験装置および実験方法 ... 152
4.4.4 葉への電流負荷効果 ... 154
4.5 結言 ... 160
第5章 総括 ... 161
5.1 各章のまとめと結論 ... 161
5.2 微小電流負荷による植物細胞の変化についての考察 ... 164
5.3 微小電流を用いた植物の低温保存における今後の課題 ... 166
参考文献 ... 167
謝辞 ... 180
v
記号表
Ab 細胞内凍結が発生する前の細胞の面積 [µm2] Ac 細胞内凍結が完了した直後の細胞の面積 [µm2] Ai 細胞内氷結晶の面積 [µm2]
B 8bitグレースケール画像の輝度
Bd 8bitグレースケール画像の輝度の変化量 ca 出現回数
d 実験開始からの経過日数 [day]
dg 細胞内氷結晶先端の代表粒径 [µm]
di 細胞内氷結晶先端の粒径 [µm]
FI 蛍光強度
Io 負荷電流値 [µA]
L リットルの単位表記 [= 10−3 m3] l 長さ [µm]
M 物質量濃度(モル濃度)の単位表記 [= mol/L]
n サンプル数
P Tukey-Kramer法による多重比較を行った際の有意水準
PC パターンを示した細胞の割合 [%]
Pd 確率密度
pHi 細胞内pH
pHm0 作製したpH標準液の0°CでのpH
pHs20 作製したpH標準液の20°CでのpH
pHv 液胞pH
Rf 蛍光強度比
SD 標準偏差
SDBd 輝度の変化量の標準偏差
vi
SE 標準誤差
T 温度 [°C]
t 経過時間 [min]
t' 細胞内に氷結晶が発現する直前からの経過時間 [msec]
tc 冷却開始からの経過時間 [sec]
ts 過冷却維持を検討する温度に到達してからの経過時間 [sec]
Tr 過冷却解除温度 [°C]
Ts 過冷却度 [°C]
RWD 重量減少率 [%]
Vc 冷却速度 [°C/min]
Wb 実験開始前の葉の重量 [g]
Wc 実験終了後の葉の重量 [g]
ε 細胞の変形度 λem 蛍光波長 [nm]
λex 励起波長 [nm]
1
第1章 はじめに
1.1 本研究の背景
今日,世界的な食糧危機が懸念されている.世界人口は爆発的に増加しており,2050 年 には97億人を超えると推計されている(1).一方,2001年以降,1981~2010年の30年平均 値に対する世界の年平均気温(陸域における地表付近の気温と海面水温の平均)の偏差は 徐々に上昇しており(2),1981~2010年平均基準に対する2015年の世界の年平均気温の偏差
は+0.42°Cと最も高い値となった(3).また,2015年は世界各地で異常高温と異常多雨もしく
は少雨が発生した(4,5).このような異常気象は今後も続くと見られることから,近い将来,
農作物の安定供給が厳しくなると見込まれている.
ところが,日本は農作物の自給率が低い.Figure 1.1に,主要31ヶ国の2011年もしくは 2013 年における人口(6)に対する穀類,砂糖類,いも類,豆類,野菜類,果実類を合わせた 主要農産物の自給率(7)を示す.31 ヶ国には,アジアからは日本,イラン,インド,インド ネシア,韓国,サウジアラビア,タイ,中国,トルコ,パキスタン,フィリピン,マレー シア,北アメリカからはアメリカ合衆国,カナダ,メキシコ,南アメリカからはアルゼン チン,コロンビア,ブラジル,ヨーロッパからはイギリス,イタリア,オランダ,スペイ ン,ドイツ,フランス,ポーランド,ロシア,アフリカからはアルジェリア,エジプト,
ナイジェリア,南アフリカ,オセアニアからはオーストラリアが選ばれている.この図よ り,日本は人口に対する主要農産物の自給率が世界的に見ても非常に低いことがわかる.
一方,日本では収穫可能な農産物の全てが市場へ出荷されておらず,損失が発生してい ると考えられる.Figure 1.2に,2015年に日本で生産された各農産物の収穫量および出荷量 と,収穫量に対して収穫量から出荷量を差し引いた未出荷量の割合を示す.Fig 1.2(A)に示 す指定野菜14品目(8)(ダイコン,ニンジン,ハクサイ,キャベツ,レタス,ネギ,キュウ リ,ナス,トマト,ピーマン,タマネギ,ばれいしょ(ジャガイモ),サトイモ,ホウレン ソウ)の総収穫量は890万2300 tであったのに対し,総収穫量のおよそ14.5%にあたる128 万6500 tは出荷されていなかった(9,10).また,Fig 1.2(B)に示す特定野菜(8)を含む26品目(カ ブ,ゴボウ,レンコン,やまのいも(ヤマイモ),ナガイモ,コマツナ,チンゲンサイ,フ
2
キ,ミツバ,シュンギク,ミズナ,セルリー,アスパラガス,カリフラワー,ブロッコリ ー,ニラ,ニンニク,カボチャ,スイートコーン,シシトウ,サヤインゲン,サヤエンド ウ,グリーンピース,ソラマメ,エダマメ,ショウガ)においても,総収穫量は186万9200 tであったのに対し,総収穫量のおよそ17.1%にあたる31万9700 tは出荷されていなかった
(9,10).さらに,Fig 1.2(C)に示す果実的野菜および果樹17品目(イチゴ,メロン,スイカ,
ミカン,リンゴ,日本なし,西洋なし,カキ,ビワ,モモ,スモモ,おうとう(サクラン ボ),ウメ,ブドウ,クリ,パインアップル,キウイフルーツ)においても,総収穫量は325
万9200 tであったのに対し,総収穫量のおよそ11.4%にあたる37万2300 tは出荷されてい
なかった(11).未出荷分に農家の自家用分が含まれていることは考えられるものの,その多 くは需要を上回る供給を抑えて,農産物の市場価格を安定させるために出荷が抑えられた 結果だと推測される.農産物は食べ頃を過ぎると腐敗が始まることから収穫可能な時期は 限られており,収穫時期を過ぎた農産物は出荷できず,損失が生ずる.それゆえ,農産物 の損失を出す前に収穫して長期保存することができれば,懸念されている食糧危機への方 策の一つとなりえる.
食料を長期保存するため,昔から塩漬け(塩蔵)・砂糖漬け(糖蔵)・酢漬け・乾燥(干 物)・燻製・発酵・瓶詰・缶詰といった数多くの食品加工技術がある.しかし,これらの食 品加工技術は,食品素材そのものの性質を大きく変化させる.一方,低温保存は食品素材 そのものの性質の変化を抑えることができ,特に冷凍保存は長期間保存できる.しかし,
一般的に用いられる冷却速度で生体の温度を下げていくと生体中の水分が凍結し,細胞外 凍結後,細胞内の過冷却を経て,細胞内凍結が発生する.細胞内凍結の発生は細胞にとっ
て致死的(12‒16)であり,また,肉や魚といった食品素材に対しては凍結・解凍後のドリップ
(17,18)や食感の低下(19‒21)を引き起こす.特に,野菜や果物はそのまま冷凍保存すると,解凍
後に特有のシャキシャキとした食感が失われる.それゆえ,野菜や果物の食感を失わずに 長期に保存できる技術の実現に向けて様々な研究がなされており,低温保存技術は更なる 向上が望まれている.
3 Figure 1.1 Self-sufficiency rate in agricultural product toward population in each country.
4
(A) Designated vegetables (14 items)
(B) Vegetables including specified vegetables (26 items)
(C) Fruity vegetables and fruits (17 items)
Figure 1.2 Crop yields, shipments and un-shipped rates in agriculture products.
5
1.2 低温保存技術について
1 . 2 . 1 食品品質が低下する要因への低温の影響
食品素材の品質の低下は,野菜や果物といった農作物は収穫直後,魚は漁獲直後,肉は 屠殺・解体直後から始まる.一部においては,熟成させることでおいしさを増すものはあ るものの,その後は時間の経過とともに品質は低下する(22).食品素材の品質の低下は,そ れまでの生育環境や状態が変化した結果,食品素材に生物的,微生物的,物理的,もしく は化学的に大別される変化過程が示されていると考えることができる(23).
生物的過程の変化(23)とは,野菜や果物が収穫後も呼吸や蒸散などの生命維持活動を続け
(24),蓄積されている栄養素を消費してエネルギー源に変換されることによって,味が悪く なるなどの品質低下が起こることである.生命維持活動にはさまざまな酵素が必要であり,
その一部は野菜や果物の品質の低下に関与している.しかし,低温に置くことで酵素活性 は低下し,呼吸や蒸散などの生命維持活動が抑制される.
微生物的過程の変化(23)とは,微生物の繁殖による腐敗が起こることである.食品の品質 変化には,微生物のうち主に細菌が関与している.細菌には多くの種類があり,生物学的 分類とは関係なく,発育温度帯から高温細菌,中温細菌,低温細菌に分類される.しかし,
低温細菌でさえも−10°Cになると発育は抑制される.
物理的過程の変化(23)の例としては,乾燥が挙げられる.食品の乾燥現象は飽和水蒸気圧 の差によって起こる.しかし,飽和水蒸気圧は温度の関数として変化し,温度の低下に伴 い小さくなることから,低温に置くことで乾燥は抑制され,生ずる品質低下は抑制される.
化学的過程の変化(23)とは,化学反応による酸化や褐変などが起こることである.しかし,
化学反応は温度の低下に従って進行速度が低下することから,温度が低いほど化学反応の 基質となる食品成分の変化も少なくなる.
このような原理から,食品品質の低下は低温に置くことで抑えることができる.食品の 品質保持期間と温度の関係は,温度の変化に応じてほぼ指数関数的に変化する(23).それゆ え,保存期間はわずかな温度の違いによって大きく変化し,温度が低いほどより長くなる(23).
一方,水は生体が安定して生存するために欠かせない存在であり(25),生体には多量の水
6
分が含まれる.水は氷点(0°C)下へ冷却していくと過冷却後に凍結して氷になるが,細胞 は冷却速度に依存して凍結に至る挙動が変化する.Figure 1.3(A)は冷却速度の違いによる細 胞内の凍結挙動の変化を示しており,横軸は冷却開始からの時間,縦軸は温度を示してい る.冷却速度に依存した細胞内の凍結挙動は,植物・動物を問わず,全ての細胞に共通す る特徴である(26).そして,細胞内の凍結挙動は,解凍後の細胞の生存率に影響を及ぼす(26).
Figure 1.3(B)は細胞内の凍結挙動の違いによる細胞の生存率の推移を示しており,横軸は冷
却速度,縦軸は細胞の生存率を示している.
1 . 2 . 2 緩慢冷却による細胞の凍結挙動
細胞を冷却していくと,まず,細胞外に氷結晶が形成される.細胞外に形成された氷結 晶は細胞内外の環境を隔てている細胞膜によって細胞内への侵入(植氷)を妨げられるた め,細胞内の水分は凍結せずに一時的に過冷却する(26).温度が同じ場合の氷と比較すると,
過冷却水の方が蒸気圧は高いことから,細胞外の氷結晶と細胞内の水分との間に化学的な ポテンシャルエネルギーの差ができる.冷却速度が低い場合,細胞内の水分はこの化学的 なポテンシャルエネルギー差を埋めるように細胞外へ排出される一方で,温度は低下する ため,細胞内は脱水された後に凍結する(12,28).このような細胞内の凍結挙動を細胞外凍結と 呼ぶ(26).冷却速度が非常に低い場合,温度低下は非常に緩慢になるため,細胞内の脱水は 続く.その結果,細胞内の水分がほぼ脱水される(Fig. 1.3(A)‒a)と,細胞は生存できない
(Fig. 1.3(B)‒a).しかし,細胞内の脱水が部分的に抑えられる(Fig. 1.3(A)‒b)と,細胞の
生存率は上昇する(Fig. 1.3(B)‒a).
この特性を活かした保存方法は緩慢凍結法と呼ばれ,医療や畜産・酪農の分野で用いら
れてきた(29‒31).しかし,緩慢凍結法は凍結・融解後の細胞の生存率などに課題が多い(32‒34)
ことから,現在は後述のガラス化保存が主流となっている(35).また,冷却に時間がかかる ことから,緩慢凍結法は新鮮な状態での保存が望まれる食品に対しては適さないと言える.
7
1 . 2 . 3 急速冷却による細胞の凍結挙動
冷却速度が高くなると温度の低下が早まるため,細胞内が脱水される前に細胞内の過冷 却度は増加する.この時,細胞外からの氷核の侵入(植氷; Fig. 1.3(A)‒c)や,細胞内での自 発的な核生成(不均質核生成, 均質核生成; Fig. 1.3(A)‒d, e)によって細胞内の過冷却が解除 されると,細胞内は瞬時に凍結する.このような細胞内の凍結挙動を細胞内凍結と呼ぶ(26). 細胞内凍結の発生は細胞にとって致死的である(Fig. 1.3(B)‒c, d, e).
肉や魚,野菜や果物といった含水量の多い食品の凍結保存において,細胞内凍結の発生 を防ぐことは困難である.含水量の多い食品を凍結させる場合,−1~−5°C の温度帯では食 品中の水分の凍結(氷結)と氷結による潜熱の発生が繰り返され,冷却の大部分は氷結に よる潜熱を除くことに使われるため,温度の低下は緩やかになる(36).この−1~−5°Cの温度 帯を最大氷結晶生成帯と呼ぶ(36).一方,最大氷結晶生成帯を素早く通過させる急速冷凍は,
凍結・解凍後のドリップや食感の低下を抑えることができる.そこで,マグロの急速冷凍 が知られているように,肉や魚,加工食品の長期保存には急速冷凍が用いられている.
1 . 2 . 4 細胞内のガラス化
冷却速度をさらに高くしてガラス転移温度を素早く通過させると,過冷却している細胞 内の水分は結晶化せずに,ガラスに転移する(ガラス化; Fig. 1.3(A)‒f).ガラス化とは熱力 学的に非平衡である準安定状態の非晶質(アモルファス)になることであり,過冷却液体 からガラスに転移することをガラス転移と呼ぶ(37,38).また,このような細胞内の凍結挙動は ガラス化と呼ばれ(26),ガラス化によって細胞の生存率は急激に上昇する(Fig. 1.3(B)‒f).
細胞内をガラス化させて保存する方法はガラス化凍結保存と呼ばれ(30),一般的には DMSO やグリセロールといった凍結保護剤(39‒41)の中に細胞を入れ,液体窒素を用いて行わ れている.ガラス化凍結保存は,医療や畜産・酪農,学術研究などの幅広い分野で卵子(42)
や精子(43,44),角膜(45),種子(46)など様々な生体を対象に用いられている.しかし,肉や魚,
野菜や果物といった含水量の多い食品は,最大氷結晶生成帯が障壁となり,食品中の細胞
8
(A) Different in intracellular freezing behavior with respect to a cooling rate
(B) Transition of cell survival rate with respect to intracellular freezing behavior
Figure 1.3 Intracellular freezing behavior and cell survival rate.
9 内をガラス化させることは不可能だと言える.
1.3 植物細胞と動物細胞
本研究は植物の低温保存をテーマにしているため,本節では植物細胞を中心に細胞の基 本構造を簡潔に述べる.
Figure 1.4に,植物細胞と動物細胞の模式図を示す.細胞は,植物・動物を問わず細胞膜
によって細胞内外の環境が隔てられており,細胞膜は細胞内の環境を一定に保つ役割を持 つ.細胞内には,細胞核と細胞内小器官(オルガネラ)があり,細胞核と細胞内小器官以 外の空間はコロイド状の細胞質基質で満たされている.細胞内の細胞核以外の領域,言い 換えると,細胞内小器官と細胞質基質を併せた領域は細胞質と呼ばれている.細胞内小器 官は植物細胞と動物細胞に共通して存在するものと,植物細胞のみ,もしくは動物細胞の みに存在するものとがある.
細胞膜は脂質二重層と呼ばれる構造をしており,細胞の活動に必要な物質のみを細胞外 から細胞内へ取り入れたり,不要になった物質を細胞内から細胞外へ排出したりといった 細胞内外の物質を移動させるのにきわめて精巧な機構を持つ(48‒51).細胞の活動に必要なイ オンの一つに水素イオン(プロトン,以下,H+)があり(25,51),細胞内pH(以下,pHi)は細 胞膜によって7.2~7.4の弱アルカリ性に保たれている(52,53).それゆえ,pHiは細胞活性を示 す指標の一つとなっている(54,55).なお,一般的にpHiは細胞質基質のpHを指している.
液胞は植物細胞特有の細胞内小器官であり,液胞内液のpHは一般的に5.0~6.0の酸性で,
果物の果汁として知られている(56).また,液胞は成長した植物細胞の体積の 90%以上を占 める巨大な器官であり,老廃物を貯蔵するだけでなく,細胞の膨圧を維持して細胞の内側 から細胞壁を支える役割もある(56).
また,細胞外構造ではあるが,植物にはセルロースやペクチンを主成分とする細胞壁が ある(25).細胞壁は細胞膜の外側を覆う形で存在しており,その役割には細胞の強度を高め る(25),細胞の大きさや形の決定する(57)などが挙げられる.
10
Figure 1.4 Schematic illustrations of plant and animal cells(47).
11
1.4 凍結・解凍処理後に起こる食品素材の品質の低下
Love は,魚の切り身の凍結前後および解凍後の断面を撮影し,冷却速度が高くなるほど 食品中に形成される氷結晶が小さくなること明らかにした(17).その後,Fennema らは,凍 結速度と細胞構造の破壊程度の関係は形成される氷結晶サイズに依存すると報告した(20). これらの報告に基づき,凍結・解凍処理後に起こる食品品質の低下は,凍結時に細胞内外 に形成された氷結晶によって細胞膜や細胞壁に機械的なダメージが与えられた結果,解凍 後に細胞内液が漏れ出るドリップや食感の低下が起こると考えられるようになったと推測 される.また,急速冷凍は食品中に形成される氷結晶を小さくすることから,凍結・解凍 処理後の食品品質の低下を抑えると知られるようになったと考えられる.
しかし,野菜や果物といった植物性食品素材をそのまま急速冷凍しても,凍結・解凍処 理後の組織軟化は著しく,特有のシャキシャキした食感が失われる.Fuchigamiら(58)は解凍 処理後のニンジン組織を凍結前と比較すると,細胞壁の主成分の一つである水溶性ペクチ ンの量が破断強度の低下と共に減少することを報告した.さらに,Fuchigamiら(59)やRoyら
(60)は,ニンジン組織を非常に緩慢な冷却条件で凍結・解凍処理した後に細胞間に隙間が見 られ,細胞壁に剥離が生じていることを報告した.それゆえ,凍結・解凍後の野菜の組織 軟化は,細胞壁の変化に起因していると考えられてきた.
一方,宮脇は,凍結・解凍処理後の植物性食品素材の組織軟化は細胞壁の変化のみによ って起こるのではなく,細胞膜の水の透過性の変化によっても起こると考察した(61).この 考察を踏まえて,安藤らは核磁気共鳴法(NMR)を用いてタマネギ組織の凍結前および凍 結・解凍後の細胞膜の水透過性を測定・比較したところ,細胞膜の水透過性が凍結・解凍 後に増大したことから,凍結・解凍後の野菜組織特有の軟化は細胞内の水分が細胞外へ流 出することによって起こると考察した(62).
野菜や果物といった植物性食品素材は,動物性のものよりも水分を多く含む.また,
Ishikawa らはコメとブドウから採取したプロトプラストの細胞膜の水の透過率を誘電率か
ら測定した結果,植物細胞の細胞膜の水の透過率は動物細胞と比較して低いことを明らか にした(63).それゆえ,植物細胞は凍結に伴う細胞内の水分移動が起こりにくいため,細胞
12
内凍結が発生しやすく,細胞内凍結による細胞膜へのダメージは大きいと考えられる(61).
1.5 低温環境と植物
1.2.1で述べたように,野菜や果物は収穫された後も呼吸や蒸散などの生命維持活動が 行われている.しかし,低温に置くことで酵素活性は低下し,生命維持活動は抑制される.
一方,野菜や果物にはさまざまな産地・性質・育ち方があり,種類によっては温度が一定 以下に低下すると,凍結温度に至らなくても褐変やピッティング(腐食)といった低温傷 害が発生する(64).特に,熱帯・亜熱帯原産の植物は一般的に低温感受性が高いことから,
低温傷害を起こしやすい(65).GrahamとPattersonは,低温傷害を受けた植物にはタンパク質 の変化や代謝の低下が見られることを報告した(66).
また,植物は低温ストレスを受けると pHiが低下する.Murai と Yoshida は,キクイモ
(Helianthus tuberosus L.)塊茎組織の凍結過程においてpHiが低下して酸性化するとともに,
細胞の生存率も低下することを明らかにした(54).それゆえ,低温ストレスによる植物のpHi
低下は,細胞の生存率へ影響を及ぼすことが知られている.
しかし,温帯以北に自生する植物は,地域によっては氷点下にもなる寒い冬を超えなけ ればならない.そのため,温帯以北に自生する植物は秋から冬にかけての温度低下と日照 時間の短縮を感知して,低温や凍結による傷害発生を回避する耐寒性や耐凍性と呼ばれる 能力を獲得する.このような植物が低温耐性を獲得していく過程を低温馴化と呼ぶ(65,67). Yoshidaらは低温馴化させた冬コムギ(Triticum aestivum L. cv. Chihokukomugi)と低温馴化さ せていないものの葉から単離したプロトプラストを用いて,凍結過程のpHi推移および細胞 の生存率の推移を比較した(55).その結果,低温馴化させていないプロトプラストは温度低 下とともにpHiが低下して酸性化し,細胞の生存率も低下したものの,低温馴化させたプロ トプラストは凍結過程の pHi と細胞の生存率の低下が抑えられていたことを明らかにした
(55).
13
1.6 これまでの低温保存技術に関する研究
1 . 6 . 1 細胞内凍結に関する研究
植物の凍結保存において,細胞内凍結の発生は凍結・解凍後の品質を左右するキーであ り,凍結による細胞へのダメージを低減する低温保存技術の開発のためには,細胞内の氷 結晶形成挙動を把握することが非常に重要である.
細胞内に形成される氷結晶および氷結晶形成挙動に関する研究は,1930 年代から行われ てきた(68).光学顕微鏡で細胞内凍結の発生を観察すると,細胞内が暗くなる現象を見るこ とができる.この現象はフラッシングと呼ばれており,細胞内の暗化は細胞内に形成され た氷結晶によって光が散乱され,通過しにくくなるために起こる(69,70).
動画による細胞内凍結の撮影はModilibowskaとRogers(71)によって初めて行われ,その後 の細胞内凍結に関する研究には動画が用いられるようになった(72‒74).その中でも,白石ら は,タマネギを用いた細胞内凍結の発生において細胞核周りから氷結晶が出現して成長す る様子を捉えたことを報告し,また,観察された細胞内氷結晶の成長過程を模式図で表し て,フラッシングの形が多様であること示した(74).
近年では高速度カメラの登場およびその精度の向上に伴い,高速度カメラを用いて細胞 内凍結の発生メカニズムや進行過程を明らかにする研究が行われてきた.上村らはシロイ ヌナズナ葉とキクイモ塊茎から単離したプロトプラストを用いて,細胞内の氷結晶が細胞 膜近傍から出現して広がっていく様子を報告した(75).また,StottとKarlssonはウシ肺動脈 内皮細胞を用いて,細胞内凍結が進行する際の細胞内のグレースケール値の変化から細胞 内氷結晶の成長速度を求める手法を検討した(76).
上述したように,細胞内凍結の発生および細胞内の氷結晶形成挙動に関する研究はこれ までにも行われてきた.また,これまでの研究から,細胞内凍結の発生は細胞外からの氷 核の侵入によって起こることが知られている(77,78).しかし,細胞内に氷結晶が出現する瞬間 は未だ捉えられておらず,また,冷却速度の違いが細胞内の氷結晶形成挙動に与える影響 は検討されていない.それゆえ,細胞内凍結の発生および細胞内の氷結晶形成挙動を理解 するためには,更なる研究が必要である.
14
1 . 6 . 2 植物の低温保存に関する研究
植物を保存する目的は多様であり,保存目的や保存する植物の種類によって保存方法は 異なる.例えば,植物遺伝資源の保存を目的とする場合は,種子として保存したり,自然 植生が行われたりしている.また,植物の培養細胞や茎頂のガラス化保存も行われている(79). 生鮮野菜や生花などの流通を目的に短期貯蔵する場合は,氷点よりも高い温度帯を用いた 冷蔵保存を行う.これらの保存方法は,いずれも保存後の植物が生きた状態であることを 望まれる場合に用いられる.
一方,野菜や果物を食用目的に長期保存する場合には,保存後の生死は必要とされない.
しかし,サラダやカットフルーツとして生食することも多いため,凍結・解凍処理後に起 こる野菜や果物特有の食感の低下は大きな課題となっている.それゆえ,植物性食品素材 の長期保存に関する研究においては,凍結・解凍処理後の食感の低下を抑えることに関心 が集まっている.
中川らは,キュウリとタマネギをソルビトールといった低分子糖,デキストリンといっ た高分子糖もしくはその両方を浸透させた後に凍結保存する方法を考案し,味に改良の余 地はあるものの,解凍処理後の歯ごたえの評判がよかったことを報告した(80).
また,凍結による植物組織へのダメージを低減する方法として,浸透圧を利用して農作 物を脱水した後,凍結保存する浸透圧脱水凍結法が知られている(81).しかし,浸透圧脱水 凍結法による組織軟化の抑制効果については,OhnishiとMiyawakiが力学物性と電気物性か ら評価した結果,ニンジンとブロッコリーではその効果が見られたものの,ジャガイモで は見られなかったことが報告された(81).さらに,安藤らによっても,浸透圧脱水凍結法を 適用したタマネギとニンジンに対して核磁気共鳴法(NMR)を用いた細胞膜の水透過性測 定とレオメータを用いたテクスチャーの測定が行われ,浸透圧脱水凍結法は細胞壁に関す るテクスチャーの保持には有効であるものの,細胞膜に関するテクスチャーや水透過性の 変化は抑えられないことが報告された(82).
近年では,植物性食品素材を凍結させずに長期保存する技術としてガス・クラスレート・
ハイドレート(gas clathrate hydrate; 気体包接化合物,ガスハイドレート)が着目され,安
15 藤らはキセノンガスハイドレート利用した野菜の低温保存の研究を行ってきた(83,84).ガスハ イドレートとは,水分子によって形成された籠状構造の中にガス分子が入った結晶のこと である(85).しかし,植物組織にキセノンハイドレートを形成・解離後,細胞膜の水透過性 に著しい増大が示され,キセノンハイドレートが形成・成長する際に細胞膜へダメージを 与えることが報告された(86).それゆえ,組織軟化を抑制しながら植物性食品素材を長期に 保存できる新たな技術が必要とされている.
1 . 6 . 3 食品中に形成される氷結晶を制御する凍結保存技術の研究
食品の急速冷凍は,Birdseyeが−45.5°Cの低い気温下で急速冷凍された魚を数ヶ月後に解 凍しても新鮮な品質が保持されていたことを発見し,商業化したことから始まったと言わ
れている(87,88).その後,食品の急速冷凍に関する研究は盛んになり,1960 年代には,冷却
速度が高くなると食品中に形成される氷結晶は小さくなること(17)や最大氷結晶生成温度帯 の存在(89)が知られるようになった.それゆえ,食品の凍結保存技術の開発においては,冷 凍食品が商業化されてからしばらくの間は,いかに冷却速度を高めて食品から熱を奪うか に注目が集められていた.
一方,Miyawaki らは,大豆タンパク質ゲルを同じ冷却条件下で凍結する実験を行った際
に,偶発的に過冷却を伴ったサンプル群に生成された氷結晶は,過冷却を伴わない場合と 比較すると微細であったことを報告した(90).この報告により,食品中に形成される氷結晶 を小さくする方法として過冷却現象が着目され,小林らは過冷却現象が食品凍結時の氷結 晶の形態およびドリップロスに及ぼす影響を検討した結果,過冷却解除温度が低いほど食 品中に形成される氷結晶が小さくなり,食品のドリップは少なくなることが明らかにした
(91).しかし,過冷却は非常に不安定な状態であり,維持することが難しいことから,過冷 却を用いた凍結保存の実用化に向けた課題として,過冷却の制御法に言及している(91). 近年では,外部から電気エネルギーを付与することによって,食品中に形成される氷結 晶を制御する技術の研究も行われている.吉本らは2 kVおよび8 kVの交流電場で魚類の凍 結を試み,冷凍による品質低下が抑えられることを見込んだものの,有意な結果を得るま
16
でには至らなかった(92).しかし,Xanthakisらは3~12 kVの直流電場で豚肉を凍結させた結 果,凍結温度は上昇した一方,食品中に形成される氷結晶は小さくなったことを報告した(93). また,Itoらは,低周波の10 kVの交流電場でウニを凍結させた結果,鮮度が長期間維持さ れ,凍結・解凍後のドリップが抑えられたことを報告した(94).しかし,これらの研究は,
形成された氷結晶や食品からのドリップ量を比較することによって電気エネルギー付与に よる効果を検討しているものの,そのメカニズムの解明までには至っていない.
1.7 本研究の目的と特徴
生体は,細胞の活動によって細胞膜や液胞膜といった生体膜(48,95)を介して電荷が異なるイ オンの分布に差が生じるため,生体膜の内外で膜電位と呼ばれる電位差が生じている(48,96). それゆえ,生体中には電気が生じていることから,治療に電気刺激が用いられる場合があ る(97).その中でも,マイクロカレント(微小電流)を用いた電気刺激は損傷した骨格筋の 再生や術創部の回復の促進(98‒100),靭帯損傷の急性期におこる腫脹の軽減 (101)が見られるこ とから,微小電流には細胞を活性化させて,組織修復を促す効果があると考えられている.
本研究は,植物の新しい低温保存技術の可能性を探るため,外部からの付与エネルギー として細胞活性を期待できる微小電流に着目し,低温保存において植物組織へ微小電流(最 大出力電圧は10 V)を負荷することによる植物のダメージ低減を検討するために行われた.
まず,細胞内の過冷却解除がどのように起こっているのか,また,細胞内の氷結晶形成 挙動において凍結による細胞へのダメージの低減につながる手掛かりはあるのかを把握す るため,高速度カメラを用いて細胞外から細胞内凍結が発生する瞬間を撮影し,細胞内の 氷結晶形成挙動の評価にパターンを用いた評価を新たに導入した.急速冷凍は凍結・解凍 後の食品品質の低下を抑えることから,まず,電流負荷なしの場合で冷却速度の変化によ る細胞内の氷結晶形成挙動の変化を評価した後,電流負荷ありの場合で冷却速度の変化に よる細胞内の氷結晶形成挙動の変化を評価した.
次に,低温ストレスによって引き起こされるpHi低下による細胞へのダメージに対する微 小電流負荷の効果を検討するため,まず,電流負荷なしの場合で冷却速度を変化させて,
17 冷却の進行を伴う植物組織のpHi推移を計測した後,電流負荷ありの場合で冷却速度を変化 させて,冷却の進行を伴う植物組織のpHi推移を計測した.
このように,微小電流負荷が細胞内の氷結晶形成挙動およびpHi推移へ及ぼす影響を微視 的に明らかにした後,実用的見地から微小電流負荷の効果を評価した.
1.8 本論文の構成
本論文は5つの章から成り立っている.
第1章である本章では,序章として本研究の背景および目的を述べた.
第2章では,微小電流負荷が細胞内凍結の発生や細胞内の氷結晶形成挙動に及ぼす影響 を検討した.細胞内凍結の発生や細胞内の氷結晶形成挙動を把握するため,まず,電流負 荷なしで冷却速度を変化させて植物組織を冷却し,細胞内凍結の発生を高速度カメラで撮 影した.得られた高速度カメラ画像を用いて,細胞外から細胞内凍結が発生する瞬間およ び細胞内の氷結晶形成挙動をパターン化し,冷却速度の変化によるこれらのパターン変化 を評価した.また,高速度カメラ画像を用いて細胞内の氷結晶による細胞の変形度,細胞 内の氷結晶の成長速度および輪郭を評価する手法を新たに検討し,冷却速度の変化が細胞 内に形成される氷結晶へ及ぼす影響を微視的に評価した.次に,電流負荷した場合の細胞 内凍結の発生を高速度カメラで撮影し,電流負荷なしの場合と比較して,微小電流負荷が 細胞内凍結の発生や細胞内の氷結晶形成挙動に及ぼす影響を検討した.章の最後に,得ら れた結果と知見から,細胞内凍結の発生および電流負荷による細胞内の氷結晶形成挙動の 変化について考察した.
第3章では,微小電流負荷が冷却中の植物組織のpHi推移へ及ぼす影響を検討した.まず,
蛍光pH指示薬を用いたpH計測方法を検討し,pH検量線の作成を行った.次に,蛍光pH 指示薬を用いたpH計測値の妥当性を確認するため,pHメーターと蛍光pH指示薬を用いて 0°CのpH標準液のpHを計測し,その結果を比較した.また,蛍光の退色および電流負荷 によるpHi計測への影響を確認するため,16°C一定温度下のpHi推移を計測し,電流負荷し た場合としない場合とで比較した.こうして,電流負荷なしで冷却速度を変化させて冷却
18
の進行を伴う植物組織のpHiを計測し,冷却速度の変化が植物組織のpHiへ及ぼす影響を評 価した.その後,電流負荷した場合の冷却の進行を伴う植物組織のpHiを計測し,電流負荷 なしの場合と比較して,微小電流負荷が冷却中の植物組織のpHi推移へ及ぼす影響を検討し た.章の最後に,得られた結果と知見から電流負荷によるpHi推移の変化について考察した.
第4章では,第2章と第3章で得られた結果に基づいて,実用的見地から植物の低温保 存における微小電流負荷の効果とその持続性を評価した.まず,電流負荷した場合に細胞 内凍結が発生するまでの過冷却度が増加したことに着目し,電流負荷が細胞内の不安定な 過冷却を安定的に維持する可能性を検討した.微小電流を負荷しながら,もしくは負荷な しで冷却速度1°C/minで室温から−8°Cまで試料を冷却した後,−8°Cを保持して細胞内凍結 が発生するまでの時間を計測して,比較した.次に,電流負荷した場合に低温ストレスに よるpHi低下が抑えられていたことに着目し,低温ストレスによる細胞活性の低下を抑制す る電流負荷の効果の持続性を検討した.微小電流を負荷しながら,もしくは負荷なしで試 料を0°Cで5日間保存し,FDAの蛍光強度から細胞活性の推移を比較して,相対的に評価 した.最後に,微小電流を用いた植物の低温保存の実現性を検討するため,試料を切り出 した植物組織から葉へスケールアップして電流負荷の効果を評価した.微小電流を負荷な がら,もしくは負荷なしで葉を−15°C,−10°C,−5°Cにそれぞれ1時間置いた後,22°Cに1 日以上置いた.このようにしてダメージが与えられた葉の表面をマイクロスコープで撮影 して細胞の外観を観察し,また,実験前後の葉の重量を計測して求めた重量減少率を用い て,細胞がダメージを受けることによる水分流出を比較・評価した.
第5章では,総括として各章のまとめと,本研究を通して得られた結果および知見に基 づいて総合考察を行い,最後に,微小電流を用いた植物の低温保存に関する今後の課題を 述べた.
19
第2章 高速度カメラを用いた細胞内氷結晶形成挙動の評価
2 . 1 緒言
非常に短い時間の中で被写体を大量に撮影できる高速度カメラは,現象の変化に対して 時間精度の高い情報を提供することができる.以前の高速度カメラは,連続撮影枚数を増 やすと分解能が低くなり,分解能を高くすると連続撮影枚数が少なくなるという問題を抱 えていた.しかし,近年のカメラ技術の向上によって,分解能と連続撮影枚数の両方が向 上したカメラが登場した.そこで,冷却ステージを取り付けた倒立型顕微鏡に高速度カメ ラを取り付け,試料の選定や撮影速度の調整を繰り返しながら植物組織中の細胞内凍結の 発生を撮影し続けた結果,細胞内に形成される氷結晶を明瞭に撮影することに成功した.
本章では,高速度カメラ画像を用いて細胞外から細胞内凍結が発生する過程および細胞 内の氷結晶形成挙動をパターン化し,さらに,細胞内に形成される氷結晶の評価方法を検 討して,微小電流負荷が細胞内凍結の発生や細胞内の氷結晶形成挙動へ及ぼす影響を検討 した.試料には,細胞内の氷結晶形成挙動を観察しやすい単一細胞層の表皮組織を生きた 状態で得ることができる植物組織として,ユキノシタの葉の表皮組織を用いた.
まず,電流負荷なしで冷却速度1°C/minから100°C/minで試料を冷却して,細胞内凍結が 発生した瞬間を高速度カメラで撮影した.併せて,細胞外および細胞内凍結発生時の温度 を記録した.得られた高速度カメラ画像から細胞内の氷結晶が出現した位置を同定し,細 胞内の氷結晶が出現した位置と接する細胞の状態を確認して,細胞外から細胞内凍結が発 生する過程をパターン化した.また,細胞内での氷結晶の成長もパターン化した.さらに,
高速度カメラ画像を用いて細胞内氷結晶による細胞の変形度合いを示す細胞の変形度,細 胞内に氷結晶が出現してからの経過時間毎の氷結晶の面積に基づいた細胞内氷結晶の成長 速度を求めた.また,細胞内に形成されている氷結晶先端の輪郭と粒の大きさを求める手 法を検討して評価した.
次に,電流負荷しながら試料を冷却して,細胞内凍結が発生した瞬間を高速度カメラで 撮影し,細胞外から細胞内凍結が発生する過程および細胞内の氷結晶形成挙動を評価した.
こうして,電流負荷した場合を負荷なしの場合を比較して,微小電流負荷による細胞内凍
20
結の発生および細胞内の氷結晶形成挙動の変化を検討した.
2 . 2 実験装置および実験方法
2 . 2 . 1 低温顕微鏡システム
Figure 2.1に,本研究で使用した低温顕微鏡システムの概略を示す.低温顕微鏡システム
は,倒立型顕微鏡(IX71, OLYMPUS)に冷却ステージ(HCD301, INSTEC)と高速度カメラ
(Phantom V4.2, VISION RESEARCH)と通常のCCDカメラ(SHC-721A, SAMSUNG)を取 り付けて構築した.Table 2.1に装備品の詳細を記載する.
高速度カメラは512×512 pixelsの解像度の画像を最大2000 frames per second(fps)で撮 影することができ,細胞内凍結の発生を撮影するために使用した.通常の CCD カメラは,
試料の冷却開始から解凍が完了するまでの一連の様子を録画するために使用した.冷却は 液体窒素とヒーターからの熱を混合して冷却ステージに流し込み,冷却速度の制御はコン ピュータ(STC200, INSTEC)によって行われた.なお,冷却ステージ内を観察する観察窓 が結露によって曇るのを防止するため,冷却ステージ上には窒素ガスを流した.
2 . 2 . 2 試料
Figure 2.2に,本研究で使用した植物の画像と植物組織の顕微鏡画像を示す.試料にはユ
キノシタ(Saxifraga stolonifera Curtis)の葉を用い,葉の表皮組織を10 mm×10 mmの大き さに切り出して使用した.ユキノシタの葉は,2014年12月下旬から2015年1月中旬に神 奈川工科大学キャンパス内で生育していたものを実験前に葉柄から切り取った後,乾燥を 防ぐために水を張ったトレーの中に置いた.
21 Figure 2.1 Schematic illustration of the cryomicroscopic system.
22
Table 2.1 Component list of the cryomicroscopic system
装置名 装備品 メーカー 製品名/型番 備考 光学顕微鏡 倒立型リサーチ
顕微鏡
Olympus IX71
ハロゲンランプ ↑ 12V100WHAL- L
100W
ハロゲンランプ ハウス
↑ U-LH100-L
ハロゲンランプ 電源装置
↑ TH4-100
ユニバーサルコン デンサ
↑ IX2-LWUCD NA: 0.55, WD: 27 mm
×40対物レンズ ↑ SLCPlanFl 40x Na 0.55, WD: 7.7 mm 高速度カメラ Phantom Vision
Research
V4.2
カメラ コントロール ソフトウェア
↑ Phantom
Camera Control (Version9.0.640.
0-C PhCon:640)
コントロールPC DELL Latitude D531 HDD: 120 GB
CPU: AMD Turion™ 64×2 Mobile Technology TL-50 1.6 GHz
メモリ: 800 MHz, 1GB OS: Windows XP Professional SP2
CCDカメラ - SAMSUNG SHC-721A
冷却ステージ 温度制御装置 INSTEC STC200 加熱装置(Heater) ↑ LN2-P 冷却装置(Cooling
pump)
↑ LN2-F2
Hot/cold stage ↑ HCD301
温度制御ソフトウ ェア
↑ WinTemp for
STC200 V1.2.111024 電気刺激装置 - 日本光電 SEN-3401
アイソレータ ↑ SS-203J
23
2 . 2 . 3 試料への電流負荷方法
Figure 2.3に,試料への電流負荷の方法を示す.まず,カバーガラスの両端にアルミシー
ルを貼り,その上に電線を取り付けて固定した.電線を取り付けたカバーガラス上に試料 と乾燥を防ぐための蒸留水を置き,その上に周囲にシリコングリースを塗布したカバーガ ラスを置いて,蓋をした(Fig. 2.3(A)).カバーガラスの外側に出ている電線は,それぞれ電 気刺激装置(SEN-3401, Nihon Kohden)と繋げた(Fig. 2.3(B)).
本章の負荷電流値は,電気刺激装置のアイソレータ(SS-203J, Nihon Kohden)で設定した 値(1 µAおよび10 µA)を示している.また,電流負荷は,0.5 msec毎に0.1 msecの間試料 へ電流を負荷し,60 sec毎にその極性を反転させる波形パターンを用いた(Fig. 2.3(C)).
2 . 2 . 4 実験方法
室温(22°C)に設定した冷却ステージ上に試料と乾燥を防ぐための蒸留水を挟んだカバ ーグラスを置き,電流を負荷しながら,もしくは負荷なしで冷却ステージの温度を下げて,
細胞内凍結が発生するまで試料を冷却した.冷却速度の設定は1°C/min,10°C/min,100°C/min を用いた.本章の冷却速度は,設定した値を示している.
撮影は,試料の冷却過程を観察して,細胞内凍結が発生した瞬間に高速度カメラのシャ ッターボタンを押して行った.これをトリガーにした前後1秒ずつが1000 fpsで撮影された.
24
(A) Leaf of strawberry geranium
(B) Microscopic image of epidermal tissue of strawberry geranium leaf
Figure 2.2 Leaf and cell of strawberry geranium (Saxifraga stolonifera Curtis).
25 (A) Preparation of cover glasses for electric current load on a specimen
(B) Schematic illustration of electric current load on a specimen between cover glasses
(C) Waveform of electric current loading with reversal of the polarity
Figure 2.3 Method of electric current load on a specimen between cover glasses.
26
2 . 3 Tukey-Kramer 法による取得データの検定について
本研究において得られたデータは,母数の有意差を確認するため,一元分散分析(ANOVA)
を行った後,Tukey-Kramer法を用いて多重比較を行った.ANOVAと多重比較はR(102)を用 いて行い,図中の小文字のアルファベットは多重比較の結果を示している.アルファベッ トの並びは平均の並びに対応しており,平均間に有意差がない場合に同じアルファベット が付与される.また,アルファベットに付与されているアスタリスク(*, **)は,有意水準
(P)の違いを示している.有意水準が低いほど,結果は要因の影響を受けていると考えら れる.
2 . 4 細胞外および細胞内凍結発生時の温度
2 . 4 . 1 細胞外および細胞内凍結発生時の温度の定義
まず,細胞外および細胞内凍結発生時の温度を評価した.Figure 2.4にCCDカメラで撮影 した試料の冷却過程を示す.試料と蒸留水を挟んだカバーガラス(Fig. 2.4(A))を冷却して いくと,カバーガラスの間にある蒸留水が凍結した後(Fig. 2.4(B)),細胞内凍結の発生を示 す,フラッシングと呼ばれる現象(69,70)を試料中に順次見ることができた(Fig. 2.4(C)).
本研究では,細胞外および細胞内凍結発生時の温度をそれぞれ次のように定義した.細 胞外凍結発生時の温度は,カバーガラスの間にある蒸留水の相変化を目視で確認した時の 温度と定義した.細胞内凍結発生時の温度は,カメラの観察下にある細胞の半分以上(お よそ 7 個)でフラッシング現象を確認した時の温度と定義した.これは,同一組織内の細 胞であっても細胞内凍結は同じタイミングで発生せず,また,カメラによって観察できる 範囲は組織のごく一部に限られることに基づいて定義した.なお,細胞外および細胞内凍 結発生の温度は,水の凍結温度(0°C)から過冷却解除の温度(Tr)を差し引いた過冷却度
(degree of supercooling: Ts)を用いて表記した.
= 0−
27 (A) Before cooling
(B) Extracellular freezing
(C) Intracellular freezing
Figure 2.4 Occurrence of extracellular and intracellular freezing captured with an ordinary CCD camera.
28
2 . 4 . 2 電流負荷しない場合
Figure 2.5に,電流負荷なしで冷却速度1°C/min,10°C/min,100°C/minで試料を冷却した
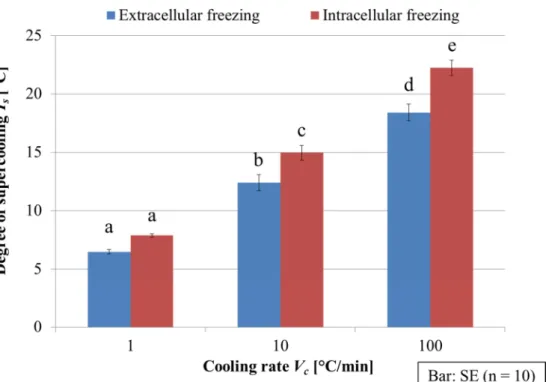
際の細胞外および細胞内凍結の発生までの過冷却度を示す.横軸は冷却速度(Vc),縦軸は 過冷却度(Ts)を示している.細胞外および細胞内凍結の発生までの過冷却度はそれぞれの 冷却速度において10回ずつ実験して得られた結果の平均値を示しており,バーは標準誤差 を示している.また,小文字のアルファベットはTukey-Kramer法による多重比較の結果(P
< 0.05)を示している.アルファベットの昇順はデータ平均の昇順に対応しており,平均間 に有意差がない場合に同じアルファベットが付与されている.この結果から,冷却速度が
1°C/min の場合における細胞外および細胞内凍結の発生までの過冷却度以外には有意差が
認められたことがわかる.
冷却速度が1°C/minの場合,細胞外凍結発生までの過冷却度はおよそ6.5°C,細胞内凍結 発生までの過冷却度はおよそ7.9°Cであったのに対し,冷却速度が10°C/minの場合はそれ ぞれおよそ12.4°Cと15.0°C,冷却速度が100°C/minの場合はそれぞれおよそ18.4°Cと22.2°C であった.また,細胞外凍結が発生してから細胞内凍結が発生するまでの過冷却度の差は,
冷却速度が1°C/min の場合はおよそ 1.4°C,10°C/minはおよそ 2.6°C,100°C/min はおよそ
3.8°Cであった.この結果から,電流負荷なしで冷却速度が高くなると,細胞外および細胞
内凍結発生までの過冷却度と,細胞外凍結が発生してから細胞内凍結が発生するまでの過 冷却度の差は共に増加すると言える.
2 . 4 . 3 電流負荷した場合
Figure 2.6に,電流負荷しながら冷却速度1°C/minで試料を冷却した際の細胞外および細
胞内凍結の発生までの過冷却度を示す.横軸は負荷電流値(Io),縦軸は過冷却度(Ts)を示 している.細胞外および細胞内凍結の発生までの過冷却度はそれぞれの負荷電流値におい て 10 回ずつ実験して得られた結果の平均値を示しており,バーは標準誤差を示している.
なお,電流負荷なしの場合と比較するため,0 µAの結果も示している.また,小文字のア
29 ルファベットはTukey-Kramer法による多重比較の結果(P < 0.05)を示している.アルファ ベットの昇順はデータ平均の昇順に対応しており,平均間に有意差がない場合に同じアル ファベットが付与されている.この結果から,細胞外凍結の発生までの過冷却度には電流 負荷の有無による有意差は認められなかったものの,細胞内凍結の発生までの過冷却度に は電流負荷によって有意差が認められたことがわかる.
電流負荷なしの場合,細胞外凍結発生までの過冷却度はおよそ6.5°C,細胞内凍結発生ま での過冷却度はおよそ7.9°Cであった.一方,1 µAの電流負荷した場合はそれぞれおよそ
6.9°Cと8.6°C,10 µAの電流負荷した場合はそれぞれおよそ7.1°Cと9.7°Cであった.また,
細胞外凍結が発生してから細胞内凍結が発生するまでの過冷却度の差は,電流負荷なしの 場合はおよそ1.4°Cだったのに対し,1 µAの電流負荷した場合はおよそ1.8°C,10 µAの電 流負荷した場合はおよそ2.6°C であった.この結果から,負荷電流値が高くなるにつれて,
細胞内凍結発生までの過冷却度と,細胞外凍結が発生してから細胞内凍結が発生するまで の過冷却度の差は共に増加することがわかった.それゆえ,冷却速度が低くとも電流負荷 することによって,細胞内凍結が発生するまでの過冷却度は電流負荷なしで冷却速度が高 くなった場合と同様の傾向を示すと言える.
30
Figure 2.5 Degrees of supercooling for extracellular and intracellular freezing in no electric current load.
Figure 2.6 Degrees of supercooling for extracellular and intracellular freezing in electric current load at the cooling rate of 1°C/min.
31
2 . 5 高速度カメラで捉えた細胞内の氷結晶形成過程
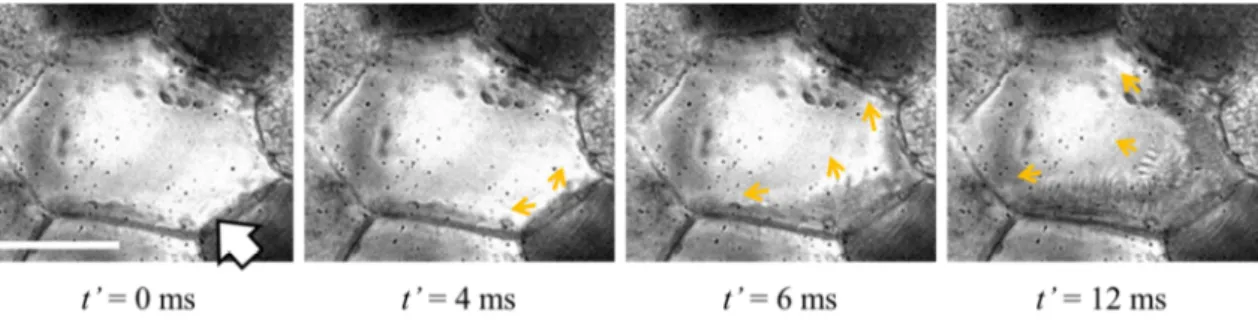
Figure 2.7に,冷却速度1°C/minで試料を冷却して高速度カメラで撮影した,細胞内で氷
結晶が成長する過程を示す.図中の白線は25 µm,白抜きの矢印は氷結晶が出現した位置,
小さい矢印は氷結晶の成長方向,t' は氷結晶が出現する直前からの経過時間を示している.
Fig. 2.7(A)から,細胞内の氷結晶が細胞壁に接した箇所から出現して,細胞の輪郭に沿って 成長していく過程がわかる.また,Fig. 2.7(B)からは,細胞内の氷結晶が細胞壁に接した箇 所から出現して,細胞内部を横切って成長している過程がわかる.これらが示すように,
本研究では細胞内の氷結晶形成挙動を明瞭に捉えた画像を得ることができた.
また,Figure 2.8に,10 µAの電流を負荷しながら,もしくは負荷なしで冷却速度1°C/min で試料を冷却して高速度カメラで撮影した,細胞内で氷結晶が成長する過程を示す.図中
の白線は25 µm,白抜きの矢印は氷結晶が出現した位置,小さい矢印は氷結晶の成長方向,
t' は氷結晶が出現する直前からの経過時間を示している.この図から,10 µAの電流負荷し た場合(Fig. 2.8(A))の細胞内の氷結晶は,電流負荷なしの場合(Fig. 2.8(B))と比較して粒 が微細なことがわかる.そこで,得られた高速度カメラ画像を用いて,微小電流負荷が細 胞内凍結の発生や細胞内の氷結晶形成挙動へ及ぼす影響を多面的に検討した.
32
(A) Progress of intracellular ice crystal growing along the configuration of the cell
(B) Progress of intracellular ice crystal growing across the center of the cell
Figure 2.7 Growth of intracellular ice crystal in the epidermal tissues of strawberry geranium (Saxifraga stolonifera Curtis) leaves.
33 (A) Intracellular ice crystal with electric current load of 10 µA
(B) Intracellular ice crystal without electric current load
Figure 2.8 Comparison of intracellular ice crystal with and without electric current load.
34
2 . 6 細胞内氷結晶の出現位置
2.6.1 細胞内氷結晶の出現位置のパターン定義
まず,細胞内の過冷却解除がどのようにして起こっているのかを把握するため,細胞内 の氷結晶の出現位置を確認して,パターン化した.Figure 2.9に,ユキノシタの葉の表皮組 織中の細胞内に氷結晶が出現する様子を捉えた高速度カメラ画像を示す.図中の白線は 25 µm,白抜きの矢印は氷結晶の出現位置,t’ は氷結晶が出現する直前からの経過時間を示す.
細胞内の氷結晶の出現位置は,細胞壁に接する箇所(Fig. 2.9(A))と細胞の中央部(Fig. 2.9(B)) に分けることができた.氷結晶が細胞の中央部から出現した場合においては,出現位置を 特定するためにさらに観察を繰り返したが,観察面もしくは観察面の反対側(底部)にあ る細胞壁に接する箇所から氷結晶が出現したのか,細胞内部から俄かに出現したのかは判 別できなかった.
Figure 2.10は,細胞内の氷結晶が出現した位置の模式図である.Fig. 2.10(A)は細胞内の氷
結晶が細胞壁に接した箇所から出現したパターン,Fig. 2.10(B)は細胞内の氷結晶が細胞の中 央部から出現したパターンを示している.そこで,電流負荷なしで冷却速度を高くした場 合と電流負荷した場合の細胞内の氷結晶の出現位置パターンの発生率を調査して,微小電 流負荷による影響を検討した.
35 (A) Intracellular ice crystal appearing at the cell wall
(B) Intracellular ice crystal appearing at the center of the cell
Figure 2.9 Locations where intracellular ice crystals appeared in the epidermal tissues of strawberry geranium (Saxifraga stolonifera Curtis) leaves.
Figure 2.10 Schematic illustrations of appearance locations of intracellular ice crystals.
(A) The cell wall (B) The center part of the cell