昼も夜も動くキツネザル:周日行性の系統発生と至近メカニズム,
および適応的意義をさぐる
佐藤宏樹 京都大学大学院アジア・アフリカ地域研究研究科 1.はじめに 霊長類学,とくに日本の霊長類学が目指してきた目 標の一つとして,「人間性やホミニゼーションの理解」 が挙げられる。そのため,ヒト(Homo sapiens)に最も 近縁な類人猿や複雑な社会構造を持つ直鼻猿亜目[日 本の場合はニホンザル(Macaca fuscata)]が古くから研 究対象として取り上げられてきた(山極,2009)。一方, 霊長目にはヒトが属さないもう一つのグループである 曲鼻猿亜目が存在し,筆者はそのうちのマダガスカル 島に生息するキツネザル下目を対象に研究してきた。 この研究対象は霊長目の中ではヒトと近縁ではないた め,「人間」を理解するには直鼻猿亜目に比べて不向き かもしれない。では,キツネザル研究の何が面白いの だろうか。直鼻猿亜目の研究だけでは至ることのでき ない,「霊長類」への理解を深めるような形質がキツネ ザル下目にはいくつか存在しているのである。例えば, キツネザル下目でみられる単独性から群居性に至る多 様な社会構造は霊長類社会の進化過程を理解する上で 適 し た 研 究 対 象 に な っ て い る し(Müller & Thalmann, 2000; Shultz et al., 2011),性的二型の欠如やメス優位社 会などの特異的な雌雄間関係は直鼻猿亜目が辿らな かったもう一つの霊長類の進化として注目されている (Kappeler & Fichtel, 2015; van Schaik & Kappeler, 1993)。こうしたキツネザル研究の魅力的なトピックの中から, 本稿では筆者が取り組んできたキツネザルの活動パ ターンに関する研究について紹介してみたい。 1.1. 霊長類学における活動パターン研究の意義 現生の霊長類の活動パターンを分類群ごとに整理し ていくと,キツネザル下目を除く曲鼻猿亜目,すなわ ちロリス下目は夜行性であり,直鼻猿亜目はメガネザ ル下目と広鼻猿下目のヨザル属(Aotus)以外は昼行性 である(Griffin et al., 2012; Santini et al., 2015)。霊長類 の共通祖先は夜行性であり,霊長類全体における進化
の流れとしては,夜行性から昼行性へと移行してきた と 考 え ら れ て い る(Griffin et al., 2012; Santini et al., 2015)。霊長類における昼行性化は,群れや複雑な社会 性の進化を伴い(Shultz et al., 2011),視覚や脳の発達に もつながる一大イベントであったと考えられる(Barton, 1998)。よって,霊長類における活動パターンの進化は 霊 長 類 学 の メ イ ン ト ピ ッ ク の ひ と つ に な っ て き た (Heesy & Ross, 2001; Martin, 1990; Martin et al., 2003)。
活動パターンの進化と適応的意義を理解する上で最 も重要になる研究対象は,分類群内で活動パターンが 多様化しているキツネザル下目だろう。とくに夜行性 とも昼行性とも異なる,昼も夜も動く周日行性を示す 分類群は同トピックにおいて欠かすことのできない対 象となっている。 1.2. 周日行性とは 周日行性の発見は,1960 年代にまで遡る。Petter(1962) はエリマキキツネザル属(Varecia),チャイロキツネザ ル属(Eulemur),ジェントルキツネザル属(Hapalemur) が示す昼行性とも夜行性とも定義しにくい活動パター ンを発見し,「薄明薄暮活動性(crepuscular)」という言 葉で表現している。さらにキツネザル類の野外生態研 究が本格化した 1970 年代には,コモロ諸島のマヨット 島に人為的に導入されたチャイロキツネザル(Eulemur fulvus)を対象として,日中にも夜間にも相当の活動量 が記録された(Tattersall, 1979)。やはり夜行性とも昼行 性とも適用しがたかったために,Tattersall(1987)はギ リシャ語の“Kata(= Through)”と“Hemera(= Day)” を用いて“Cathemerality”という言葉を考案し,今日ま で使用されている。日本語では小山・高畑(2009)に したがって“周日行性”と呼ぶことにする。周日行性 とは採食や移動などのまとまった活動が 24 時間を通し て,もしくは日中と夜間に同程度配分される活動パター ンを指し,夜行性の動物が日中に巣穴からわずかな時 間 だ け 出 る と い っ た 行 動 は 周 日 行 性 に 含 ま れ な い 3 2017 年 3 月 31 日受付,2017 年 5 月 8 日受理,2017 年 6 月 20 日早期公開(J-STAGE) *e-mail: [email protected] 総 説 〈高島賞受賞記念〉
佐藤宏樹 4 (Tattersall, 1987)。 周日行性の活動パターンを把握するには終日終夜を 通した長時間かつ,季節変動を検出するために十分な 期間の行動観察が不可欠である。野外調査の成果が蓄 積されて,その至近要因および究極要因に関する論争 が本格化したのは,ラジオテレメトリー法等で効率的 に対象を追跡できるようになった 1990 年代以降である。 近年では加速度計データロガーが内蔵された首輪をキ ツネザルに装着し,5 分間隔の活動状態を数か月間にわ たって自動的に計測する技術も導入されている(Erkert & Kappeler, 2004; Kappeler & Erkert, 2003)。そして現在 までに,周日行性の至近メカニズムと適応的意義,進 化的な起源を探るために,行動生態学だけでなく,時 間生物学,系統分類学,形態学など,多様な分野にお ける成果が蓄積されてきた。本総説では 30 年近くに及 ぶ周日行性についての研究成果と論争を整理し,同研 究分野が目指すべき将来の課題について考えてみたい。 哺 乳 類 に お け る 周 日 行 性 は 食 肉 目 や 有 蹄 類(van Schaik & Griffiths, 1996),ネズミ目やトガリネズミ目
(Halle & Stenseth, 2000)などの広い分類群で確認されて いる一方,霊長目ではまれで,直鼻猿亜目では唯一ヨ ザ ル 属 で 確 認 さ れ て い る だ け で あ る(Aotus azarai Fernandez-Duque, 2003; A. trivirgatus Wright, 1989)。曲鼻 猿亜目では既述の 3 属だけでなく,近年ではジェント ルキツネザル属に近縁なオオジェントルキツネザル属
(Prolemur)が周日行性に位置づけられた(Curtis, 2006;
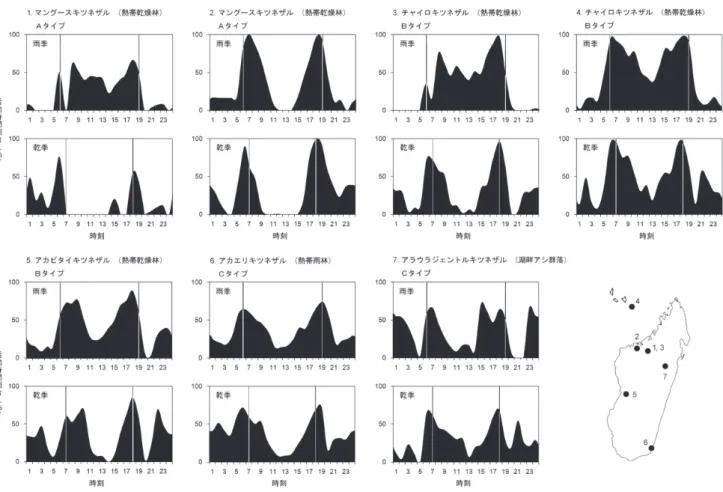
Curtis & Rasmussen, 2006)。さらに,厳格な昼行性であ ると定義されていたワオキツネザル(Lemur catta)でも 夜間活動が確認されおり(Donati et al., 2013; LaFleur et al., 2014; Parga, 2011),キツネザル下目における周日行 性の分布と起源が大きく見直されている(系統発生に おける周日行性の進化的起源は 2.2.1 節で詳述する)。 周日行性キツネザルの活動パターンは一様ではなく, 種や地域によって変異が確認されており,大きく 3 つ のタイプに分けることができる(Curtis & Rasmussen, 2002)(図 1)。
A タイプ:雨季(夏:長日)にほぼ昼行性としてふる まい,乾季(冬:短日)に活動パターンが移行してほ
図 1 周日行性キツネザルの活動パターン。各個体群の24 時間の活動時間割合を雨季と乾季に分け,種名,生息地タイプ,活動パターン タイプ(A-C)を表記した。各個体群の番号は右下の調査地ポイントの番号に該当している。グラフ中の 6-7 時,18-19 時の縦線は それぞれ日の出と日の入りのタイミングを示している。1, 3 のグラフは Rasmussen(1999),2 は Curtis et al.(1999),4 は Tarunaud (2006),5 は Donati et al.(1999)および Curtis & Rasmussen(2002),6 は Donati & Borgognini-Tarli(2006),7 は Mutschler(1998)
およびCurtis & Rasmussen(2002)に基づいて作成した。
46
1846
1847
図
1
1848
1849
1850
1851
1852
1853
1854
1855
1856
1857
1858
1859
1860
1861
1862
1863
1864
1865
1866
1867
1868
1869
ぼ 夜 行 性 に な る(Curtis et al., 1999; Rasmussen, 1999, 2005; Tattersall, 1987)。雨季と乾季が明瞭な熱帯乾燥林 にみられる。図 1-2 は年間を通して薄明薄暮に活動ピー クがあるが,乾季になると日中の活動停止が顕著化し, 夜間の活動が増えることからA タイプに分類された (Curtis & Rasmussen, 2002)。
B タイプ:雨季(夏:長日)にほぼ昼行性を示すが,乾 季(冬:短日)に夜間活動の割合が増すことで昼も夜 も 活 動 が み ら れ る 周 日 行 性 に な る(Rasmussen, 1999, 2005; Tarnaud, 2006)。雨季と乾季が明瞭な熱帯乾燥林に みられる。 C タイプ:1 年を通して昼も夜も活動する恒常的な周日 行性を示す(Colquhoun, 1998; Donati & Borgognini-Tarli, 2006; Schwitzer et al., 2007a)。季節が不明瞭な熱帯雨林 にみられる。 これらの共通点や差異を生じさせる至近要因と究極 要因については,後ほど触れていく。 2. 周日行性は昼行性化の途中か? 霊長類の共通祖先が夜行性であることは長らく主張 されており,近年の研究においても強く支持されてい る(Griffin et al., 2012; Santini et al., 2015)。我々ヒトを 含め,直鼻猿亜目のほとんどは昼行性であることから, 霊長類は夜行性から昼行性に進化してきたといえる (Santini et al., 2015)。一方,原始的な霊長類の特徴を多 く残すキツネザル下目において夜行性・周日行性・昼 行性の種が混在していることから,「周日行性はもとも と夜行性だった種による昼行性化の途中であり,進化 的に不均衡な状態なのか?」それとも「周日行性は適 応進化した完成型なのか?」という問いかけがなされ てきた。本節では近年になって終結しつつある,系統 発生上の論争をまとめていきたい。 2.1. 進化的不均衡仮説 マダガスカル島では 2300-1200 年ほど前にヒトが移入 して以降(Cox et al., 2012; Tofanelli et al., 2009),それま で日中の生態的ニッチを独占していた現生キツネザル の競争相手となりうるジャイアントリーマー[8 属から 成り体重 10-200 kg もあったとされる大型キツネザル類 の 総 称(Godfrey & Jungers, 2002; Jungers et al., 2002)] と 天 敵 と な り う る 大 型 の 猛 禽 類(Aquila 属 や Stephanoaetus 属 : Goodman 1994; Goodman & Muldoon, 2016)が狩猟によって絶滅したとされている(Goodman & Jungers, 2014)。van Schaik & Kappeler(1996)はこう したマダガスカル島における歴史的背景を考慮して, 近年に生じた競争と捕食によるリスクからの解放が きっかけとなって夜行性キツネザルの一部に昼行性化 がおこり,その途中段階としてみられる昼夜の活動が 周日行性であるとする進化的不均衡仮説を提唱した。 夜行性から昼行性への移行の途中段階としての周日 行性は,哺乳類においてはいくつか報告がある。南米 に 生 息 す る タ テ ガ ミ ナ マ ケ モ ノ(Bradypus torquatus: Chiarello, 1998),アフリカ南東部に生息するミナミキノ ボ リ ハ イ ラ ッ ク ス(Dendrohyrax arboreus: Milner & Harris, 1999),南太平洋の島々のサモアオオコウモリ (Pteropus samoensis: Brooke, 2001),南米のミナミヨザル
(Aotus azarai:Wright, 1989)など,これらの動物は本来 夜行性だが,捕食者である猛禽類が不在になっている 地域個体群においては日中の活動が可能となり,周日 行性の活動パターンが確認されている。よって,進化 的不均衡仮説は普遍的に支持されうるアイデアなので ある。 2.2. 進化的不均衡仮説の検証 すでに過去に起こった競争者と捕食者の絶滅を根拠 とする進化的不均衡仮説を直接検証することは不可能 である。しかし,「現行の周日行性キツネザル類の夜行 性からの逸脱は過去 2000 年内に起こった」とする本仮 説の前提を検証することは可能である。 2.2.1. 系統分類からの検証 野外観察で確認された各種の活動パターンを種間の 系統分類関係と対応させ,夜行性・周日行性・昼行性 の進化的な起源を推定する研究が近年行われている (Griffin et al., 2012; Santini et al., 2015)。Griffin et
al.(2012)は霊長類の共通祖先およびキツネザル下目 の共通祖先は夜行性であると推定し,現生のインドリ 科とキツネザル科からなる単一の分岐群が夜行性の主 要分岐群から分かれる際にまず昼行性となり,その後, インドリ科およびキツネザル科内のエリマキキツネザ ル属とワオキツネザル属(Lemur)がそのまま昼行性を 引き継ぎ,チャイロキツネザル属とジェントルキツネ ザル属は昼行性から周日行性に進化したと推定した(図 2a)。よって,周日行性の起源はチャイロキツネザル属 の共通祖先が出現する 1300 万-900 万年前まで遡ること になる(Yoder & Yang, 2004)。
しかし,Griffin et al.(2012)による結果は,解析対象 となった活動パターンデータと系統分類データが古い。 チャイロキツネザル属は 1990 年代から長期的な野外行 動観察によって周日行性と位置づけられてきたが,ジェ ントルキツネザル属(Eppley et al., 2015)やワオキツネ ザ ル 属(Donati et al., 2013; LaFleur et al., 2014; Parga, 2011)における周日行性の位置づけは 2010 年代に入っ てからである。さらに近年の系統解析ではキツネザル 下目の共通祖先の主要分岐群からまずキツネザル科が, 次にインドリ科が独立に分岐したと考えられている (Horvath et al., 2008)。これらの新しいデータを用いて
佐藤宏樹 6 各活動パターンの起源を探ったSantini et al.(2015)は, キツネザル科の共通祖先で夜行性から周日行性が進化 し,インドリ科ではアバヒ属(Avahi)を除くインドリ 属(Indri)およびシファカ属(Propithecus)それぞれの 共通祖先で夜行性から昼行性が進化したと結論づけて いる(図 2b)。キツネザル科の中で現在もエリマキキツ ネザル属だけが昼行性の扱いとなっているが(Santini et al., 2015),エリマキキツネザル属では夜間活動の研究 が乏しく,今後の研究次第で周日行性への位置づけの 可能性は十分にあり得るという(Donati et al., 2013)。よっ て周日行性の起源はキツネザル科の共通祖先が出現し た 4600-2000 万年前まで遡ることになる(Donati et al., 2013; Santini et al., 2015)。
Griffin et al.(2012)にしても Santini et al.(2015)に しても,系統解析による周日行性の起源の推定年代は 進化的不均衡仮説の前提を否定することになる。さら に,どちらの研究にしても周日行性は活動パターンの 移行における最終段階であり,周日行性から他の活動 パターンへの移行は起こっていない。マダガスカル島 はサイクロンや干ばつ,植物の資源生産の不規則なパ ターンを原因とする不安定な環境であるため,柔軟に 活動時間帯を変更できる周日行性が安定的な適応戦略 として進化したと位置づけられる(Santini et al., 2015)。 よって周日行性を昼行性化の途中段階とする進化的不 均衡仮説の主張は否定されている。 2.2.2. 眼の形態と機能からの検証 日中と新月の夜では環境の光強度に 1000 億倍も差が あるとされ(Dusenbery, 1992),そうした光環境に対応 する動物の眼は,一般的に暗い環境では視覚の感度 (sensitivity)を上げる機能が発達し,明るい条件では視 覚 の 精 度(acuity) を 上 げ る 機 能 が 進 化 す る(Kirk, 2004)。昼夜両方の光環境を活動する周日行性について 進化的に安定な形質とする立場と不均衡な形質とする 立場では,予想される眼の構造や機能が異なる。すな わち,前者では周日行性に特殊化した形質が,後者で は夜行性種の特徴を色濃く残す形質が検出されると予 想される。 キツネザル下目における眼球サイズに対する角膜サ イズの比率は夜行性種の方が昼行性種よりも大きいが, 周日行性種は中間的な比率が検出された(Kirk, 2006)。 また,網膜上の視細胞における視覚精度を上げる錐体 細胞と視覚感度を上げる桿体細胞の密度は,周日行性 種(アカビタイキツネザル Eulemur rufifrons 17,800 個 / mm2 : 200,000-430,000 個 /mm2 )は夜行性種(ハイイロネ ズミキツネザル Microcebus murinus 7,600-10,900 個 /mm2 : 500,000-630,000 個 /mm2 )と昼行性種(ヴェローシファ カ Propithecus verreauxi 34,000 個 /mm2 : 225,000-370,000 個/mm2 )の中間的な密度となった(Peichl et al., 2017)。 よって周日行性キツネザルの眼の形態は昼夜両方の活 動を可能にするために適応進化した形質であると考え られる(Kirk, 2006)。 図 2 キツネザル下目における活動パターンの進化プロセスの系統解析による推定。(a)Griffin et al.(2012)による結果に基づき作成した。 (b)Santini et al.(2015)による結果に基づき作成した。系統樹末端の円は現生属の活動パターンを示し,黒,灰,白はそれぞれ夜行性, 周日行性,昼行性となる。系統樹内部の円グラフは現生属の活動パターンから推測される結節点(共通祖先)の活動パターンを示す。 円グラフ内の3 色の割合は各活動パターンの事後確率を表している。
47
1870
1871
図
2
1872
1873
1874
1875
1876
1877
1878
1879
1880
1881
1882
1883
1884
1885
1886
1887
1888
1889
1890
1891
1892
1893
1894
1895
1896
1897
霊長目では昼行性の狭鼻猿類やホエザル属(Alouatta) では三色型色覚が発達しており,ホエザル以外の昼行 性広鼻猿類は多型三色型色覚とされる(Jacobs et al., 1996)。多型三色型色覚では,X 染色体上にコードされ るオプシン遺伝子に中波長および長波長の色覚を介在 する 2 つのタイプが存在するため,異なる遺伝子がコー ドされたX 染色体を 2 本もつヘテロ型のメスのみが三 色型色覚を有することになる。キツネザル下目では, 昼 行 性 で あ る エ リ マ キ キ ツ ネ ザ ル 属 2 種(Varecia variegate, V. rubra) と コ ク レ ル シ フ ァ カ(Propithecus coquereli),さらに周日行性のアオメクロキツネザル
(Eulemur flavifrons)で X 染色体上の中 / 長波長オプシン
遺伝子の多型が確認され,多型三色型色覚の可能性が 示唆されている(Jacobs & Deegan, 2003; Surridge et al., 2003; Tan & Li, 1999; Veilleux & Bolnick, 2009)。一般に霊 長類における三色型色覚は夜行性昆虫食ニッチから昼 行性果実食ニッチに移行する際に進化したと考えられ ており(Heesy & Ross, 2001),さらに多型三色型色覚の 進化と維持には進化的に安定した日中活動が必要とな るため(Jacobs & Deegan, 2003),少なくともこれら 3 属での日中活動の起源は古くまで遡ると考えられる (Veilleux & Bolnick, 2009)。
霊長類では上記のようにそれぞれの活動パターンご とに視覚に関する形質は特殊化しているため,その特 殊化が短期間での活動パターンの移行を妨げていると 考えられる(Bearder et al., 2006; Kirk, 2006; Santini et al., 2015)。よって,目の構造および機能の検証からも進化 的不均衡仮説は否定されるのである。 一方で現在みられる活動パターンと一致していない 形態的特徴も確認される。夜行性動物に発達している 視覚感度を上げる脈絡層タペタム(反射膜)は,キツ ネザル科では昼行性とされるエリマキキツネザル属に は存在せず,周日行性のジェントルキツネザル属とワ オキツネザル属には確認される(Pariente, 1979)。チャ イロキツネザル属では種によって有無の報告が異なる (Pariente, 1979; Schwitzer et al. 2005)。インドリ科では夜 行性のアバヒ属はもちろん,昼行性のシファカ属にも タペタムが確認されている(Martin, 1990)。さらに錐体 細胞が密集する網膜上の中心窩(視覚精度が最も高い 領域)は,エリマキキツネザル属,ジェントルキツネ ザル属,ワオキツネザル属には存在するが,チャイロ キツネザル属では欠落している(Pariente, 1979)。各種 における活動パターンとタペタムおよび中心窩の有無 の矛盾については,いまだに納得のいく説明がなされ ていない。 3. 時間生物学からみる周日行性の至近メカニズム これまで述べたように,現在では周日行性は適応的 な獲得形質ととらえる考え方が主流となっている。そ のため,現在の行動解析に基づくアプローチは周日行 性の至近メカニズムもしくはその適応的意義を追及す る研究に分かれている。本節では至近メカニズムを追 及する,時間生物学における研究成果についてレビュー する。 生物には環境サイクルの長さに似た周期で自律的に あらゆる生理機能のリズムを振動させる体内時計が存 在しており,恒常的な環境条件にさらされた哺乳類で は約 24 時間の周期(=概日リズム)が確認されること が多い(Aschoff, 1965)。しかし,夜行性動物は暗期に, 昼行性動物は明期に活動時間を調節する必要があるた め,1 日に起こる実際の明暗位相と少しずれる体内時計 を同調させる必要がある(位相変化機構)。このずれを 調 整 す る 外 部 環 境 同 調 因 子 を ツ ァ イ ト ゲ ー バ ー [zeitgeber(独語)= time-giver(英語)]と呼ぶが(Aschoff, 1954),哺乳類では光が最も強力なツァイトゲーバーと なり,網膜にある光受容体で光を感じることで脳視床 下部視交叉上核を中枢とする体内時計が実際の明暗位 相に同調される(Refinetti, 2006)。そのため,日の出入 りが活動リズムの制御因子となっており(Aschoff, 1966; Aschoff et al., 1982; Gwinner, 1986; Reppert & Weaver, 2002),例えば夜行性霊長類のモホールガラゴ(Galago moholi)は日の入りが活動開始,日の出が活動停止の キ ュ ー と な る(Bearder et al., 2006)。さらにキタリス (Sciurus vulgaris)のように日中活動の開始を日の出で 制御している場合,季節的な日長の変化は日中の活動 量の季節変化と相関する(Wauters, 2000)。 周日行性を示すキツネザル類では,哺乳類に普遍的 に認められる活動パターンの概日リズムや光環境への 反応はどうなっているのだろうか。さらに季節によっ て活動パターンに変化が生じる現象はどのように説明 されるのだろうか。 3.1. 日の出入りと日長の機能 野外における行動観察では,周日行性のキツネザル 各種に共通して日の出と日の入りに活動のピークを迎 える双峰型の活動パターンが検出されている(図 1. Curtis et al., 1999; Donati et al., 2009; Donati & Borgognini-Tarli, 2006; Kappeler & Erkert, 2003; Schwitzer et al., 2007a)。一方で,夜間の活動には目立ったピークはみら れない(Kappeler & Erkert, 2003)。これは日の出と日の 入りがツァイトゲーバーとして機能していることを示 唆 し て お り(Donati & Borgognini-Tarli, 2006; Donati et al., 2013),夜行性のキツネザルにも共通している(Warren & Crompton, 1997)。
佐藤宏樹 8 マダガスカル島は赤道から南に離れて位置するため, 日の出入りのタイミングは季節的に変化する。ワオキ ツネザルでは,日の出入りのタイミングの季節的な変 化とともに日中の活動の開始と停止のタイミングもず れるため,長日化とともに日中活動量が増加する(Donati et al., 2013)。昼行性のヴェローシファカも同様の傾向 がみられたが,日の出入りの時刻の季節変動以上に, 活 動 の 開 始 と 停 止 の 時 刻 の 季 節 変 動 が 大 き か っ た (Erkert & Kappeler, 2004)。短日条件では日の出後(朝遅 く)に活動が開始し,日の入り前(夕方早く)に活動 が停止する一方で,長日条件では日の出前(朝早く) に活動が開始し,日の出後(夕方遅く)に活動が停止 する。これは日長によって日の出入りへの反応を変化 させていることを示している。より顕著な例が,チャ イロキツネザル属で報告されている周日行性のA タイ プとB タイプである(図 1)。A タイプを示すマングー スキツネザル(Eulemur mongoz)は長日条件で昼行性, 短日条件で夜行性へと移行していることが示唆される (Curtis et al., 1999)。B タイプでは,アカビタイキツネ ザル(Eulemur rufifrons)が長日条件で日中の活動時間 割合を,短日条件で夜間の活動時間割合を高くしてい る(Kappeler & Erkert, 2003)。Donati & Borgognini-Tarli (2006)は日長の変化に伴うアカエリキツネザル(Eulemur collaris)の活動パターンの移り変わりを詳細に分析し ている。日長が長くなって日の出時刻が早まると朝の 活動開始が早まり,かつ午前の活動停止が遅くなる。 同様に長日化に伴って日の入りの時刻が遅くなるほど, 午後の活動開始時間は早まる。よって,長日で昼行性 活動が増え,短日で夜間活動が増えるという結果にな る(Donati & Borgognini-Tarli, 2006; Donati et al., 2009)。 これらの結果は昼と夜の時間が等しくなる春分と秋分 を境に昼行性と夜行性の活動時間割合を変えているこ とを示しており,時間生物学的に言い換えると日長が 季節的な活動パターンの移行を制御するツァイトゲー バーとなっている可能性を示唆している(Kappeler & Erkert, 2003; Donati & Borgognini-Tarli, 2006)。
3.2. 月光による影響
チャイロキツネザル属の野外行動観察では満月の夜に 夜間活動が増える現象や(Colquhoun, 1998; Donati et al., 1999; Harrington, 1975; Schwitzer et al., 2007a),夜間の光 強度が強いほど夜間活動が多くなる現象が確認されてい る(Curtis et al., 1999; Donati et al., 2001, 2009; Donati & Borgognini-Tarli, 2006; Eppley et al., 2015)。満ちていく月 の夜は 20-23 時頃に,満月の時は 23-2 時頃に,欠けて いく月の夜は 2-5 時頃に天頂付近に位置して最も明るい 時間帯を迎えるが,これらの時間に合わせて夜間活動が ピークを迎えることが報告された(Kappeler & Erkert, 2003)。さらに満月時に起こった皆既月食では,月食前
に活発だったアカビタイキツネザルは月食に伴って活動 を停止し,月食終了時に活動を再開したことも記録され ている(Donati et al., 2001)。また,翌日の日中活動につ いては,満月の夜の翌日は活動量が減り,新月の夜の翌 日は増える傾向が確認されている(Donati & Borgognini-Tarli, 2006)。夜間の光強度増加に伴う夜間活動の増加と 翌日の日中活動量への影響はヨザルにおいても同様の傾 向がみられる(Fernandez-Duque, 2003)。 上記の一連の報告は,月光による夜間の光刺激には日 の出入り(ツァイトゲーバー)によって体外環境に同調 された概日リズムを改変する効果があることを示してい る。このように本来の規則正しい一定のリズムを隠す効 果をマスキング効果という(Aschoff et al., 1982)。この マスキング効果の適応的意義としては光強度の強い環境 の方が採食や捕食者検出の効率が上がるためだと解釈で きるが(Gursky, 2003; Kappeler & Erkert, 2003),まだ確 定的な説明とは言い切れない(Bearder et al., 2006)。 夜間は月明かりの届く林冠部での活動が多くなるこ と や(Andrew & Birkinshaw, 1998; Curtis et al., 1999; Donati et al., 1999; Rasmussen, 2005),乾季に落葉するマ ダガスカル西部の乾燥林では雨季よりも乾季に夜間の 活動が増えること(Colquhoun, 1998),季節的な落葉が ない東部の降雨林では月光と夜間活動の関係が不明瞭 であることが報告されている(Overdorff, 1988, Overdorff & Rasmussen, 1995; た だ し Donati & Borgognini-Tarli, 2006)。森林内の夜間における光強度を左右する環境要 因による夜間活動への影響からも,月光によるマスキ ング効果が示唆されている。 3.3. 室内行動実験における分析 人工的に光条件を設定できる実験室での行動分析は 時間生物学分野においてよく用いられるアプローチで あり,周日行性キツネザル類を対象とした室内行動実 験もわずかに行われている。マングースキツネザル(周 日行性),ワオキツネザル(周日行性),コクレルシファ カ(昼行性),アカエリマキキツネザル(Varecia rubra: 昼 行 性 ), ク ロ シ ロ エ リ マ キ キ ツ ネ ザ ル(Varecia variegata: 昼行性)を実験室で人工的に自然条件での明 期(光強度 95-179 lx)と暗期(光強度の記載なし)に さらしたところ,いずれの種も明期に活動,暗期に休 息のリズムに調整され,昼行性的な活動パターンが検 出された(Rea et al., 2014)。このことは野外行動観察と 同様に,明暗サイクルがツァイトゲーバーとして機能 していることを示している。
さらに示唆に富む実験がErkert & Cramer(2006)によっ て,シロビタイキツネザル(Eulemur albifrons)の飼育 個体を対象に行われている。この研究では明期 12 時間, 暗期 12 時間のサイクルで実験室が管理され,明期は 145 lx の光強度に設定され,暗期の光強度は 0.15 lx(満
月の夜間光強度),0.0002 lx(新月の夜間光強度),暗順 応できない完全な暗闇の 3 条件に設定されて,活動パ ターンをモニタリングしている。その結果,24 時間内 の暗期の活動時間割合の配分は,満月条件で 89%(夜 行性的),新月条件で 51%(周日行性的),暗闇で 31%(昼 行性的)となった。野外観察と同様に明暗サイクルが ツァイトゲーバーとなることや,暗期の強い光がマス キング効果をもつことが明らかになった。また,最も 活動が集中した満月条件を恒常的に継続すると,25.2 時間で活動と休息が繰り返される体内時計周期が検出 された。さらに明期と完全暗闇のサイクルにさらして 明期に活動させるリズムに順化させた後でも,満月恒 常条件にすると,もともと暗期だった時間帯に活動ピー クを移行させて 25.2 時間周期での活動を見せることか ら,シロビタイキツネザルの体内時計は進化的に夜行 性種に由来すると主張している。このことは昼行性を 経ない,夜行性から周日行性への進化プロセスを主張 したSantini et al.(2015)の見解に一致する(図 2b)。 また,Erkert & Cramer(2006)の報告による異なる光 条件での活動量の差異が示すシロビタイキツネザルの 光環境の嗜好性は,既述の眼の構造とも一致するだろ う。本実験で設定された 4 つの光強度条件では,満月 条件で最も活動量が多くなった。しかし,野外では光 環境は連続的に変化し,最も活動する光環境は日の出 入り時の薄明るい条件である。昼行性種と夜行性種の 中間的な特徴をもつ目の構造を考えると,満月の夜や 薄明薄暮の光環境を選択した結果の周日行性活動なの かもしれない。光強度が活動パターンに及ぼす至近的 なメカニズムの解明は,光環境を人工的に設定できる 実験的検証が今後も有効となるだろう。 4. 周日行性の適応的意義 マダガスカルでは東部は年中湿潤であるが,北部, 西部,南部は日長が長い時期は雨季,日長が短い時期 は乾季に重なっており,気温や水資源,食物資源など の環境も雨季と乾季で大きく異なる(Dewar & Richard, 2007; Jury, 2003)。日長や明暗サイクルを至近要因とし てとらえる一方で,同時に季節変化する環境変数を究 極要因として周日行性の適応的意義を説明するアプ ローチが 1990 年代から現在までに試みられている。究 極要因を追及する仮説は現在までに 4 つ挙げられてお り,本節では各仮説とその検証研究について紹介して いく。 4.1.1. 捕食リスク回避仮説 肉食動物による捕食リスクを回避するために活動す る時間帯を変化させることは多くの哺乳類で指摘され てきた(van Schaik & Griffiths, 1996)。例えば,タテガ
ミナマケモノが夜間に活動するのは,大型の昼行性猛 禽類による捕食リスクを回避するためだと考えられて い る(Chiarello, 1998)。ハタネズミ亜科(Arvicolinae) の場合,植生被覆が少ない開けた生息地では捕食リス クが上がるため,植生被覆が多い生息地に比べて夜間 に 活 動 す る 傾 向 が あ る(Halle, 2000)。 イ タ チ 亜 科 (Mustelinae)の一部では,小型種は猛禽類によるリスク を回避するために夜行性になり,大型種はキツネによ るリスクを回避するために昼行性になることが知られ ている(Zielinski, 2000)。また,本来は夜行性に分類さ れるヨザル属は昼行性の猛禽類がおらず夜行性のアメ リカワシミミズク(Bubo virginianus)がいる生息地では 夜間の捕食リスクを回避するために日中の活動が増え て 周 日 行 性 に な っ て い る こ と が 示 唆 さ れ た(Wright, 1989)。 マダガスカルではマダガスカルオオタカ(Accipiter henstii) や マ ダ ガ ス カ ル チ ュ ウ ヒ ダ カ(Polyboroides radiatus),マダガスカルノスリ(Buteo brachipterus)な ど の 昼 行 性 猛 禽 類 や(Brockman, 2003; Karpanty, 2006; Karpanty & Goodman, 1999; Karpanty & Grella, 2001),3 種 の ボ ア(Acrantophis madagascariensis; A. dumereli; Sanzinia madagascariensis: Goodman, 2003; Rakotondravony et al., 1998),キツネザル食のスペシャリストとして認 識されることもあるマダガスカル最大の食肉目フォッ
サ(Crypoprocta ferox)がキツネザル類にとっての主な
捕食者となりうる(Colquhoun, 2006; Wright et al., 1997)。 そのため,上記で述べた哺乳類の例と同様に,キツネ ザル類においても捕食リスクを回避する戦略として周 日行性が進化したとする仮説が提唱された(Colquhoun, 2006; Rasmussen, 1999, 2005)。 猛禽類の場合は日中の捕食圧が想定され,マダガス カル南東部の調査では 1 年間でチャイロキツネザル属 個体群の 5.8-7.2%にあたる個体が昼行性猛禽類に捕食 されていると推定された(Karpanty, 2006)。とくに,落 葉が起こる乾季は上空から林内への視界がよくなるた め,捕食リスク回避のためにキツネザルは夜間へ活動 を移行すると予想された(Rasmussen, 1999, 2005)。 一方,フォッサは周日行性である。昼も夜も動くこ とのできるフォッサに対しては,時間的に狙いを定め させずに捕食リスクを分散する戦略として,被食者の キツネザル類では周日行性が安定するだろうと考えら れた(Colquhoun, 2006)。 4.1.2. 捕食リスク回避仮説の検証 乾季になると落葉するマダガスカル西部の熱帯乾燥 林で行われた検証研究では,林冠の植生密度が減少す るにしたがってチャイロキツネザル属が日中の活動量 を減らし,夜間の活動を増加させることが確認された (Curtis et al., 1999; Rasmussen 1999, 2005)。さらに,昼
佐藤宏樹 10
は森林内の下層で活動し,夜間は樹冠近くで活動する 傾 向 も 報 告 さ れ て い る(Andrew & Birkinshaw, 1998; Curtis et al., 1999; Donati et al., 1999, 2007; Rasmussen, 2005)。これらの結果は,日中の猛禽類による上空から のリスクおよび夜間の食肉目による地上からのリスク を回避する行動戦略として解釈されうる。しかしなが ら,他の要因を排除できるほどの盤石な検証結果では ない。例えば,落葉した森林には月光が良く届くために, 月光によるマスキング効果が夜間活動を増加させた可 能性は捨てきれない(Kappeler & Erkert, 2003)。また, 落葉の度合いは森林環境の乾燥化の指標にもなりうる ため,後に述べるように体内水分の損失を最小化する ために乾燥した季節は暑い日中の活動を回避している とも解釈できる(Sato, 2012)。さらに時間帯ごとの垂直 利用の変化についても,日中は暑熱ストレスが特に高 くなる林冠部の利用を避けるという体温調節のための 行 動 戦 略(e.g., Kosheleff & Anderson, 2009; Takemoto, 2004)である可能性も排除できないのである。体温調 節についての仮説は後に触れる。 一方,群れの構造と活動パターンの移行の関係から の検証も試みられている。クロキツネザル(Eulemur macaco)は,メスが出産後の小さなアカンボウを抱い ている時期(9-10 月)に夜間活動を増やしたことから, アカンボウへの猛禽類による捕食リスクの回避を主張 している(Andrew & Birkinshaw, 1998)。また,アカエ リキツネザルでは,群れサイズの大きな地域個体群の 方が群れサイズの小さな地域個体群に比べ,日中の活 動時間割合が高くなった(Campera et al., 2014)。同様に, 7 か月以上の昼夜行動データが蓄積されている 13 個体 群を対象としたメタ解析でも,乾季が長く,葉食の割 合が低く,群れサイズが大きい個体群で日中の活動時 間割合が高くなった(Donati et al., 2016)。乾季の長さ と葉食の割合との関係は後に述べる採食時間延長仮説 を支持する一方,群れサイズが示す効果は大きな群れ では猛禽類による捕食リスクが軽減されるためだと説 明された。雨季では昼行性で乾季に夜行性に移行する A タイプ(図 1-1, 1-2)を示した個体群はいずれもマン グースキツネザルである(Curtis et al., 1999; Rasmussen 1999, 2005)。マングースキツネザルは複雄複雌群が多 いチャイロキツネザル属の中でも雌雄ペアを基本とす る小さな群れを形成するため,捕食リスクに対応して A タイプのように極端な活動パターンの移行を示すの かもしれない。 捕食リスク回避仮説に関して逆説的に検証する場合, 捕食者がいなければ周日行性は成り立たないと予想さ れる。コモロ諸島のマヨット島ではマダガスカルから 人為的に導入されたチャイロキツネザルが生息してい るが,捕食者となる猛禽類やフォッサは生息していな い。しかし,マヨット島でも乾季に日中活動を制限し て夜間活動を増やすことから,少なくともこの個体群 においては捕食リスクが現在にみられる周日行性の要 因になっていないと考えられる(Tarunaud, 2006)。また, 乾季に落葉する乾燥林で周日行性のアカビタイキツネ ザルと同所分布しているヴェローシファカは年中昼行 性を維持している(Erkert & Kappeler, 2004)。シファカ 属はチャイロキツネザル属より体サイズは大きいが, オトナ個体でもチュウヒダカなどの猛禽類に捕食され る(Karpanty & Goodman, 1999)ので,捕食リスク回避 仮説はシファカ属が昼行性を維持している事実につい て説明できないことになる。これまでのところ,本仮 説に関する検証は間接的な因果関係からの解釈を試み ているにすぎず,説得力に欠けている。しかしながら, 捕食-被食に関する行動データの収集は困難であるた め,直接的な検証はいまだに行われていない。 4.2.1. 採食競合緩和仮説 競合する動物種のニッチが分化するメカニズムは, 生息地の分化(すみわけ),食性の分化(食べわけ)と 共に活動時間の分化が考えられ(Schoener, 1974),実際 に同所分布する哺乳類が種間競合を緩和するために活 動時間を分けている例が報告されている(ネズミ目 Jones et al., 2001; 食肉目 Marcelli et al., 2003)。マダガス カル北部と東部では,チャイロキツネザル属 2 種が同 所分布することから(Markolf & Kappeler, 2013; Tattersall & Sussman, 1998),競合を緩和しながら共存するメカニ ズムとして活動時間の分化が進む過程で,周日行性が 発達したとする仮説が提唱された(Curtis & Rasmussen, 2006; Tattersall & Sussman, 1998)。
4.2.2. 採食競合緩和仮説の検証 Rasmussen(1999)はマダガスカル北西部の熱帯乾燥 林に同所分布しているマングースキツネザルとチャイ ロキツネザルの活動パターンの季節変化を比較分析し, 前者をA タイプ(図 1-1),後者を B タイプに分類して いる(図 1-3)。すなわち,両者では乾季になると活動ピー クの時間帯が異なってくるのだが,同時に両者が利用 する食物の重複度合いは乾季の方が低くなった。資源 欠乏期になると同所で共存するチャイロキツネザル属 2 種が異なる資源を利用する傾向は,東部の熱帯雨林の アカビタイキツネザルとアカハラキツネザル(Eulemur rubriventer)でも報告されている(Overdorff, 1993)。よっ て,食べわけが成立しているために,「食物資源をめぐ る競合が活動時間の分化を促進する」というストーリー にはなっていない。マダガスカル北部では同所分布す るサンフォードキツネザル(Eulemur sanfordi)とカン ムリキツネザル(Eulemur coronatus)との間に食物資源 の重複利用が確認されたが,両者では同様の活動パター ンを示すことから,時間的なニッチ分化が採食競合を
緩和していることを支持するデータは得られなかった (Freed, 1996)。 本仮説はチャイロキツネザル属内だけでなく,果実 食性キツネザル同士であれば適用可能である。Donati ら は,周日行性のチャイロキツネザル属と昼行性の傾向 が強いワオキツネザルとの共存に着目し,それぞれの 活動パターンについて分析を行っている(Donati et al., 2009, 2013)。チャイロキツネザル属に着目した検証で は,ワオキツネザルと共存している個体群の方が,チャ イロキツネザル属が単独で生息している個体群よりも 夜間の活動時間割合が多いと予想されたが,この仮説 は支持されなかった(Donati et al., 2009)。両種間で起 こる敵対的な交渉の頻度も昼夜の活動時間割合を説明 する変数にはなっていない(Donati et al., 2009)。一方, ワオキツネザルに着目し,チャイロキツネザル属と共 存する個体群とワオキツネザルが単独で生息する個体 群で活動パターンを比較したところ,共存する個体群 で日中の活動時間割合が高かった(Donati et al., 2013)。 チャイロキツネザル属は昼に長く休息するため,競合 種が活動を停止している昼間にワオキツネザルは活動 時間を移行させている可能性が示唆された。 上記のワオキツネザルの昼間の活動増加の事例を除 き,本仮説を全面的に支持する野外データは得られて いない。また,果実資源をめぐる競合種が存在しない 生息地においてもチャイロキツネザル属は周日行性を 示すことから(e.g., Donati et al., 2009; Kappeler & Erkert, 2003; Tarunaud, 2006),採食競合の緩和は現在みられる 周日行性を単独で成立させる要因にはならないと考え られる。 4.3.1. 体温調節仮説 哺乳類は体温を一定範囲内に保つ内温動物であり, 温度中性域(thermoneutral zone:血液循環の調節だけで 体温を維持できる外気温の範囲)の範囲外の温度環境 にさらされた場合,寒冷条件では筋肉のふるえ産熱に よって体温を上げ,暑熱条件下では発汗や呼吸による 気過冷却によって体温を下げる生理的な体温調節機構 が発動する(Schmidt-Nielsen, 1997)。このとき,生理的 な体温調節にはエネルギーと水分を消費することにな るが,これらの生理的体温調節にかかるコストは複数 個体で抱き合ったり(Hanya et al., 2007),日陰で休んだ りする(Kosheleff & Anderson, 2009)などの行動によっ て軽減できる。この行動性体温調節には活動の量や時 間帯の調整も含まれる。例えば,草食性の大型哺乳類 (Grenot, 1992)やタテガミナマケモノ(Chiarello, 1998),
ハ リ モ グ ラ(Tachyglossus aculeatus: Abenspergtraun & Deboer, 1992)では,日中の暑さや夜間の寒さによる気 温のストレスがかかる時間帯の活動を制限することが 報告されている。
霊長類においても寒い状況で熱生産のためのエネル ギーを保存したり(Hanya, 2004),暑い中での急激な体 温上昇を回避したり(Baldellou & Adan, 1998; Hill, 2006; Huang et al., 2003; Kosheleff & Anderson, 2009),水分損失 を最小化したりするため(Campos & Fedigan, 2009)に 活動を制限することが報告されている。周日行性のキ ツネザル類は体重が 700 g から 2.5 kg と体サイズが小さ いために(Glander et al., 1992; Sato et al., 2016)外部の 温度環境の影響を受けやすい一方,代謝速度はKleiber モデルによって体重から予測される哺乳類の基準値よ りもはるかに低い(Daniels, 1984; Müller, 1985)ために 生理的な体温調節能力も高くはないと予想される。そ のため,行動による体温調節が重要な役割を果たすと 考えられる。チャイロキツネザルの体温は 38.2℃であり, 温度中性域は 22-30℃である(Daniels, 1984)。マダガス カルでは生息地によっては最低気温が 18℃,最高気温 は 40℃になり,周日行性キツネザルの温度中性域から 大幅に逸脱するため,気温に対応して活動パターンを 調節している可能性は十分にありえる。そのため,体 温調節のための行動戦略が周日行性の成り立ちを説明 するという仮説が提唱されている(Curtis & Rasmussen, 2006)。 4.3.2. 体温調節仮説の検証 雨季と乾季が明瞭なマダガスカル北西部の熱帯乾燥 林に生息するマングースキツネザルは,夜間の気温が 下がる乾季に夜間活動を増やすことが確認された(Curtis et al., 1999)。この生息地では乾季の最低気温が 18-21℃ と冷え込むため,夜間に運動することで体温を維持し ていると考えられた。しかし,他のチャイロキツネザ ル属個体群における観察では,夜間の寒い時間帯は抱 き合ってじっとしていることが多く(Donati et al., 1999; Donati et al., 2011; Ostner, 2002),ワオキツネザルでも 21℃以下に気温が下がる寒い夜は夜間の活動が減った (LaFleur et al., 2014)。活動量の減少は熱を生産するエネ ルギーを保存するための行動であると解釈できる。よっ て,Curtis et al.(1999)が主張する体温維持のための夜 間運動はどの個体群にも当てはまるわけではない。 林冠の植生が乏しい湖畔のアシ群落に生息するアラ ウラジェントルキツネザル(Hapalemur alaotrensis)は, 日中の気温が高い季節に昼間の活動を回避し,夜間の 活動を増やす(図 1-7. Mutschler et al., 1998)。マダガス カル南部の乾燥地に生息するワオキツネザルも同様に, 日中の気温が 40℃を超えると活動を停止する(Ellwanger & Gould, 2011)。雨季と乾季が明瞭なマダガスカル北西 部の熱帯乾燥林に生息するチャイロキツネザルは,水 資源が豊富な雨季は日中も継続して活動するが,乾季 になると真昼の休息が顕著化し,最高気温が高くなる にしたがって日中の休息時間は増加した(Sato, 2012)。
佐藤宏樹 12 乾燥条件下で暑熱ストレスにさらされた場合,体内水 分の損失を最小にしつつ体温の上昇を回避するために 活動の休止が最善の体温調節行動になると考えられた。 同様の傾向はマダガスカル西部に生息する昼行性の ヴェローシファカにも当てはまり,やはり乾季に最高 気温が高くなると日中の休息が顕著化し,さらに最低 気 温 が 低 く な る と 朝 夕 に 活 動 が 集 中 す る(Erkert & Kappeler, 2004)。その結果,乾燥して寒暖差が激しい時 期に日中の活動量が減ることから,エネルギーと水分 を保存する戦略であると考えられる。 上記で示されたように,暑熱ストレス下で日中の活 動を回避し,涼しい夜間に活動を移行するのであれば, 気温が高い環境の方が夜間活動の割合は高くなると予 想される。しかしながら,気温が異なる 2 つの地域でチャ イロキツネザル属の活動パターンを比較した研究では, この予想ははずれた(Donati et al., 2009)。また,気温 に大きな季節変化がない地域でも活動パターンの季節 的な変化がみられることから,体温調節戦略が周日行 性の適応的意義を必ずしも説明するわけではないよう だ(Curtis, 2006)。 4.4.1. 採食時間延長仮説 小型哺乳類や草食性の有蹄類では,食物の消化吸収 効率の制限を克服するために長時間にわたって採食す る必要があり,昼も夜も活動する要因となることが示 唆されている(有蹄類 Ensing et al., 2014; ハタネズミ Halle, 2006; トガリネズミ Merritt & Vessey, 2000; イタチ Zielinsky, 2000)。
果実食者であるチャイロキツネザル属は乾季になる と果実量の季節的な減少に伴って花や葉などの繊維を 多く含む低質なFallback food の利用を増やす傾向が各 個体群で報告されている(Ossi & Kamilar, 2006; Sato et al., 2016)。チャイロキツネザル属は繊維を消化するた めの特殊化した消化器官をもたず(Campbell et al., 2000, 2004),かつ体サイズも小さいために一度に大量に摂食 できない。このような小型の果実食動物が低質な食物 からエネルギーや各種栄養素の要求量を満たそうとす る場合,長時間にわたり採食する必要があると考えら れる。すなわち,果実が欠乏する乾季は低質なFallback food を昼も夜も食べ続けるため,周日行性活動に移行 するという仮説が提唱されている(Engqvist & Richard, 1991)。 4.4.2. 採食時間延長仮説の検証 本仮説に対しては観察から得られる採食時間だけで なく,食物の栄養分析を組み合わせて摂取した各栄養 素やエネルギーの量を考慮することによって精度が高 い検証が行われてきた。 コモロ諸島のマヨット島に生息するチャイロキツネ ザルは,雨季は昼行性活動が中心となり,乾季は日中 の活動量が減って夜間活動が増えるB タイプの活動パ ターンの移行を示す(図 1-4. Tarnaud, 2006)。摂取した 食物の重量には季節差はないものの,乾季に成熟葉の 採食量が増えるためにタンパク質と糖質の摂取量が 減って繊維の摂取量が増えたことから,エネルギーを 温存しながら活動時間の幅を広げることによって繊維 質の食物を消化する時間を増やしていると考えられた (Tarnaud, 2006)。マダガスカル南東部のアカエリキツネ ザルでも,昼行性から周日行性への季節的な移行が確 認された(Donati et al., 2007)。同個体群のアカエリキ ツネザルは年中果実食で季節的な葉食の増加はみられ ないが,果実資源の繊維量は季節的に変動しており, 食物の質の低下が採食時間の延長を引き起こし,夜間 活動の促進につながったと考えられる(Donati et al., 2007)。 さらにDonati et al.(2009)ではマダガスカル南東部 熱帯雨林および南部乾燥地の川辺林に生息するチャイ ロキツネザル属個体群を対象に,昼夜活動比を目的変 数,採食時と休息時の観察対象個体の高さ,他群や他 種に対する敵対的行動の頻度,気温,繊維摂食量の 4 つの要因を説明変数として多変量解析を行った。この 4 つの説明変数は,順に捕食リスク回避仮説,採食競合 回避仮説,体温調節仮説,採食時間延長仮説を検証す る変数となっている。解析の結果,繊維摂取量だけが 有意な変数となり,繊維摂取量の増加に伴って夜間の 活動時間割合が増加することが示された。また,2 個体 群間の比較では繊維摂取量が多い南東部の個体群の方 が, 夜 間 の 活 動 時 間 割 合 が 多 か っ た。 こ のDonati et al.(2009)の研究は,4 つの仮説を包括的に検証し,採 食時間延長仮説を支持する結論に至っている。また, Donati et al.(2016)では 7 か月以上の終日終夜行動デー タを蓄積したチャイロキツネザル属 13 個体群を対象に 昼夜活動比を説明するメタ解析を行っており,ここで もやはり葉食時間割合が高い個体群で夜間活動時間割 合が高いことが示されている。すでに述べたが,同解 析では群れサイズも有効な説明変数となっており,採 食時間延長仮説だけでなく捕食リスク回避仮説も支持 されている。 上記の報告からは,繊維を消化吸収できないチャイ ロキツネザル属は採食量を増やすことで栄養やエネル ギーの要求量を満たすPower feeding strategy を採用して いると考えられる(Donati et al., 2007; Sato et al., 2016)。 その結果,昼夜に及ぶ採食活動を行うことで周日行性 が成り立っていると主張されるのである。しかしなが ら,食物および繊維摂取量(Curtis et al., 1999)や森林 内 の 果 実 資 源 量(Rasmussen, 1999; Schwitzer et al., 2007b)の季節変化が活動パターンの季節変化と相関し ないチャイロキツネザル属個体群もいくつかあること
から,採食時間延長仮説がすべての個体群の周日行性 の適応的意義に当てはまるわけではない。 採食時間延長仮説はチャイロキツネザル属のように 繊維の消化能力が備わっていないことが前提となって いる。しかし,周日行性を示すジェントルキツネザル 属は(Eppley et al., 2015),繊維の多いタケやササの葉 やシュート,タケノコを採食するスペシャリスト(e.g., Overdorff et al., 1997; Tan, 1999)として消化管が発達し ている(Campbell et al., 2000, 2004)。ジェントルキツネ ザル属の周日行性に本仮説を当てはめた場合,体重が 1100 g と小型であるために昼夜にわたって葉を食べ続 ける必要があるとするアイデアが挙げられているが (Eppley et al., 2015),同様の体サイズで葉食性のイタチ キツネザル属(Lepilemur)やアバヒ属が夜行性である こと(Ganzhorn, 1988; Thalmann, 2001)を踏まえると説 得力に欠ける。キツネザル科の共通祖先で獲得したと される周日行性をジェントルキツネザル属が単に保持 しているだけなのか,採食戦略として周日行性が機能 するのかについては,さらに検証が必要である。 5. 周日行性研究の今後 本稿第 2 節で述べたように,周日行性の起源はキツ ネザル科の共通祖先の出現にまで遡るほど古く,進化 的に安定した戦略としての獲得形質であると現時点で は理解されている。これまで行われてきた研究は,現 生のキツネザルを対象としてどのような適応的意義が 当てはまるかを検証するアプローチが多かったが,第 4 節でまとめたようにどの仮説にも支持・不支持があり, 単一の究極要因を周日行性の適応的意義として結論づ けることは難しい。また今日みられる周日行性を成立 させる要因は,キツネザル科の祖先が周日行性を獲得 した際の適応的意義を説明するとは限らない。その逆 に現生種で究極要因として支持されない説明が祖先種 における周日行性獲得の淘汰圧だった可能性もあり得 るだろう。これまでの研究成果からいえることは,「各 仮説が提唱したようなあらゆる環境要因に対して,周 日行性は柔軟に対応しうる」ということである。Curtis (2006) やDonati et al.(2016),Santini et al.(2015)が 主張するように,キツネザル科に進化した周日行性は マダガスカルの特徴的な環境要因とされるサイクロン, 干ばつ,食物資源の量と質の低さ,年変動の大きな植 物フェノロジーなどが重なる,予測不可能で厳しい環 境 変 動(Dewar & Richard, 2007; Ganzhorn et al., 1999; Wright, 1999)に対して有利に働く柔軟な戦略として進 化していったと捉えるべきなのかもしれない。こうし た発想の転換が起こりつつある今,今後はどのような アプローチで周日行性研究に挑戦していくべきなのか を本節で考えてみたい。 5.1. 人為的攪乱に対応しうる周日行性 周日行性が予測不可能な環境変動にも対応しうるた めの柔軟な行動戦略として解釈される場合,近年新た に起こっている人為的な環境攪乱に対しても活動パ ターンを変化させて対応している可能性があるだろう。 この予想に対する検証はすでに始まっている。 Donati et al.(2016)は,人為的な攪乱の少ない生息 地に生息する個体群と攪乱を受けている生息地の個体 群の活動パターンを比較し,攪乱を受けている生息地 では日中の活動時間割合が低く,より周日行性的な活 動パターンになることを報告している。攪乱林では成 熟葉などの低栄養で繊維が多い食物の利用が増えるた め(Donati et al., 2011),採食時間延長仮説が唱える適 応的意義によって周日行性が成り立っていると主張さ れた。この傾向はチャイロキツネザル属 13 個体群を対 象としたメタ解析でも得られており,人為的な攪乱を 受けている生息地の方が日中の活動時間割合が低い (Donati et al., 2016)。 アオメクロキツネザルでは,人為的な攪乱を受けて いる森林に生息する個体群の方が昼夜全体の活動量が 多くなり,かつ夜間活動の割合も高くなった(Schwitzer et al., 2007a)。夜間活動増加の背景には,攪乱を受けた 二次林では林冠層の植生密度が低いため,猛禽類によ る捕食リスク回避のメリットが挙げられた。さらに, 二次林における夜間の光強度は植生が密な一次林より も強いため,月光のマスキング効果で夜間活動が促進 されることも考えられた(Schwitzer et al., 2007a)。 上記 2 事例では報告によって活動パターンを変化さ せる要因は異なるが,周日行性キツネザルは人為的な 環境攪乱にも柔軟に対応していることがわかってきた。 環境破壊が顕著なマダガスカルでは,人為的な攪乱と 周日行性パターンとの関係を分析する研究例が今後も 増えると考えられる。 5.2. 複数要因による段階的な説明 4 つの究極要因は互いに排他的ではないため,複数の 変数が周日行性を説明する要因として示唆されるケー スはいくつか報告がある(Andrew & Birkinshaw, 1998; Curtis et al., 1999; Donati et al., 1999)。4.4.2 節で紹介した ような,すべての究極要因を説明変数として同一の多 変量解析モデルで検証するアプローチ(Donati et al., 2009)によって,複数の究極要因が同時に提示される 可能性はあり得るだろう。現在のところ,複数の要因 の効果は並列的に提示され「要因A も要因 B も季節的 な周日行性への移行に寄与している」と説明されるこ とが多い。しかしながら,段階的な提示,すなわち「要 因A が日中活動を減少させるために,要因 B が生じて 夜間活動が増加し,周日行性になる」という説明はこ れまでにない。
佐藤宏樹 14 4.3.2 節で述べたようにマダガスカル北西部の熱帯乾 燥林に生息するチャイロキツネザルは乾季の昼間に気 温が高くなると活動を休止する(Sato, 2012)。さらに同 調査で実施した採食活動の解析からは,乾季は朝夕に 採食活動が集中し,多肉植物の葉を噛みしめて水分を 摂取していることが明らかになった(Sato et al., 2014)。 昼間の休息および朝夕の水分摂取は,乾燥条件下の暑 熱ストレスに対して体内水分を最大限に保存しつつ体 温の上昇を回避するための行動性体温調節であると解 釈できる。この時期,他の季節に比べて日中の果実採 食時間は少なくなる一方で,夜間の果実採食時間は多 くなった(Sato et al., 2014)。これらの報告をまとめると, 明瞭で長い乾季が存在するマダガスカル北西部に生息 するチャイロキツネザルでは「乾季になると日中の乾 燥暑熱に対しては体温調節のために水分摂取と休息に 従事し,その結果エネルギー不足が生じる。そのため 涼しい夜間は果実採食を行うことでエネルギー不足を 補完し,周日行性が成り立つ」という,昼夜で異なる 要因による段階的な説明が浮かび上がってくる(図 3: 佐藤, 2012)。上記の仮説は現在検証中であるが,多く の研究が解析対象としてきた昼夜の活動時間割合比率 だけでなく,昼夜別の休息や採食といった個々の活動 の根拠を明らかにすることで,これまで報告がなかっ た複数要因による段階的な周日行性活動の成り立ちが 明らかになるかもしれない。 5.3. 昼行性キツネザルとの比較 キツネザル科における周日行性がマダガスカルに特 徴的な予測不可能で厳しい環境変動に有利に働く柔軟 な戦略として進化した形質であるならば,同環境に生 息するインドリ科(夜行性のアバヒ属を除く)ではな ぜ昼行性が進化したのだろうか。両者の活動パターン を比較することにより,周日行性や昼行性の適応的意 義が浮かび上がってくる可能性がある。 特にキツネザル科のチャイロキツネザル属とインド リ科のシファカ属は多様な種に分化しながら広域に分 布しており,互いに同所分布することが多い。同じ環 境要因にさらされる両者で大きく異なるのは採食戦略 である。これら 2 属の複数個体群を対象としたメタ解 析によって両属の採食戦略の違いを検証した結果,チャ イロキツネザル属は繊維消化能力に乏しい消化管を持 つために果実に固執するスペシャリスト的な食性を示 す一方,シファカ属は繊維消化機能を兼ね備えた消化 管を持つために多様な食物を柔軟に利用できるジェネ ラリスト的な食性を持つと位置づけられた(Sato et al., 2016)。Sato et al.(2016)によると,チャイロキツネザ ル属は食性に柔軟性がない分,資源量が減少した際は 活動と遊動を柔軟に対応させながら果実資源獲得に固 執するPower feeding strategy を採用していると考えられ る。一方のシファカ属は資源量が減少した際は柔軟に 利用可能な低質食物に切り替え,活動を抑えてエネル ギーを保存する戦略を採用していると思われる。周日 行性か昼行性かは,このような食性と活動パターンの 相反する柔軟性によって説明が可能かもしれない。 もう一点,両属で大きく異なると考えられるのは水 分要求量である。チャイロキツネザル属は乾季に多肉 植物の葉の汁を舐めたり(Sato et al., 2014),数 km も離 れた水場まで水を飲みに行くこともあるが(Scholz & Kappeler, 2004),シファカ属は乾燥した生息地で水場が あ っ て も 水 を 飲 む 行 動 は 観 察 さ れ な い(Scholz & Kappeler, 2004; Simmen et al., 2003)。哺乳類が暑熱スト レス下で体温を下げるには水分を利用した気過冷却シ ステムは不可欠だが,水分の要求量が少ないにもかか わらず冷涼な夜間への活動パターンの移行を起こさな いシファカ属の生理的な体温調節機構はチャイロキツ ネザル属のそれとは大きく異なると予測できる。 Curtis(2006)が指摘するように,これまでの周日行 性研究は行動学・生態学分野からのアプローチに偏っ ている。上記に述べたような採食戦略と活動戦略との 関係や,生理的な体温調節機構と活動戦略との関係を 解明するには,行動学・生態学分野での解析と同時に, エネルギーや水分の収支をはじめとする生理学的なア プローチを組み合わせることで今後の周日行性や霊長 類の活動パターン研究を大きく進展させることができ るだろう。 謝辞 本稿は 2014 年度日本霊長類学会高島賞の受賞を機に 執筆いたしました。「高島賞」の存在と受賞された研究 者の方々は私にとって大学院時代からの憧れであり, 目標となって幾度も励まされた存在でした。本賞の創 図 3 マダガスカル北西部の乾季におけるチャイロキツネザル (Eulemur fulvus)の周日行性メカニズムの仮説。活動パター ンはRazanaparany & 佐藤(未発表データ)に基づいて作成 した。