* 連絡先著者(Corresponding author):〒214-8571 神奈川県川崎市多摩区東三田 1-1-1 E-mail:[email protected]
0 論文
ORIGINAL ARTICLE
春植物の地上部消失後の植生と光環境
大塚勇哉
*1)・倉本 宣
2)1) 明治大学大学院農学研究科 Graduate School of Agriculture, Meiji Univercity
2) 明治大学農学部 School of Agriculture, Meiji Univercity
摘要:春植物が消長したあとどのようになっているのか,植生を把握することを目的とし,神奈川県境川沿いにある2つの異なる 河畔林緑地で調査を行った。各調査地の春植物生育地内に2 m×2 m のコドラートを合計 21 カ所設置し,コドラート内の植生と光 環境の季節変化を追った。調査の結果,春植物が消長したあとの植生として,高茎草本などの特定の種が優占するコドラートと, 特定の優占種は存在しないコドラート,そしてほぼ何も生えてこないコドラートの3 つのパターンがみられた。
キーワード:ニリンソウ,相対光量子束密度,クラスター分析,DCA
OTSUKA, Yuya and KURAMOTO, Noboru: Vegetation and light environment after disappearance of aboveground parts of spring ephemerals
Abstract: We conducted the study in two forests along the Sakai River in Japan to determine emergent vegetation after spring ephemerals complete their seasonal cycle. We set up 21 quadrats (2 m×2 m) in two study areas and monitored seasonal changes in vegetation and light. From the results, quadrats were covered with either a specific species, or multiple species, or were without vegetation.
Key words: Anemona flaccida F.Schmidt, Photosynthetic photon flux density, Cluster Analysis, DCA
1. はじめに
春植物,スプリング・エフェメラルとも呼ばれる植物群は, 関東地方の雑木林において,落葉広葉樹に依存する生活史を 持っている。しかし,雑木林の放置・減少,宅地開発や大量 採掘によって,その自生地は著しく減少している。神奈川県 ではヒメニラ(Allium monanthum Maxim.)が絶滅危惧Ⅰ 類,東京都ではレンプクソウ(Adoxa moschatellina L.)が絶 滅危惧Ⅰ類に指定されている。それ以外の春植物も少しずつ 分布を狭めており,絶滅危惧に指定はされていないものの, もはや身近な山野草とはいえなくなってきた。また,春植物 は都市のレクリエーション林の中で高い観賞的価値を持って いる。こうした,希少な種としても,自然観賞の対象として も強い関心を集めている春植物の保全は重要な意義を持つ。 春植物の生態研究の一例として,養父(1988)の報告がある
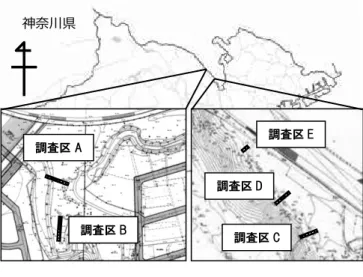
が,これはカタクリ(Erythronium japonicum Decne.)の生育 適地を競合,あるいは共生している他の植物との関係に焦点 をあてたものであった 5)。また,鈴木(1986)も各環境要因や 森林植生の年変化について調べているが,これも研究対象種 は カ タ ク リ で あ っ た 2)。 ニ リ ン ソ ウ(Anemone flaccida F.Schmidt)については,基礎的な生態を明らかにした倉本 (1984)や,宮元・桑原(2009),屋祢ら(2014)などのごく最近 の報告もある1,3,6) が,カタクリ以外の春植物について植生の 年変化や競合種,共存種を明らかにした報告はない。春植物 と春植物を取り巻く植生環境を季節の変化と共に追うこと は,春植物を含めた林床植生の保全計画を立てるうえで重要 な意義を持つ。本研究では,春植物周辺の植生から,その秋 までの植生変化のパターンを把握し,そのパターンの違いを 決定づける要因をさぐることを目的として,神奈川県の河川 沿いの春植物生育地で調査を行った。 2. 材料ならびに方法 2.1 調査地 調査は東京都町田市と神奈川県相模原市の間を流れる境川 沿いの緑地,高木道正山河畔林(以下高木道正山)と古淵鵜野 森公園(以下古淵鵜野森)の 2 ヶ所で行った(図-1)。両緑地間の 距離はおよそ 4.8 km であり,面積は高木道正山では約 1.5 ha,古淵鵜野森では約 1.6 ha である。この 2 つの調査地は, 同じボランティア団体によって管理されている。 2.2. 調査区の設定 それぞれの調査地に2 つずつ,2 m×20 m の帯状の調査区 を設置し,高木道正山に調査区A,B,古淵鵜野森に調査区 C, D,E を設定した(図-1)。それらの調査区の中に,端から 2 m ×2 m のコドラートを 2 m 間隔で設置,それぞれにコドラー ト番号を振り分けた。また,調査区C,D は通路を跨って設 置してあるが,コドラートD3 のみ通路上にかかってしまう
論 文
図-1 調査地概況および調査区設置図 (左)高木道正山 (右)古淵鵜野森
Fig.1 Overview of the study area and diagram of plot layout (Left)Takagidoshoyama (Right)Kobuchiunori ため,春植物は存在するものの全植被率が極端に低いコドラ ートD3 を削除した。調査区 E は平坦面上にコドラートを 2 箇所設置した。 2.3 調査区の環境 高木道正山の調査区A は平坦な地形であり,主な樹種はム
クノキ(Aphananthe aspera (Thunb.) Planch.),アブラチャ ン(Lindera praecox (Siebold et Zucc.) Blume)と,すべて落
葉広葉樹で構成されている。調査区B も平坦な地形であるが, その樹種構成は常緑針葉樹のサワラが多く占めており,つい でアブラチャン,エノキと落葉広葉樹が生えている。そのた め,春季でも比較的暗い環境となっている。古淵鵜野森の調 査区C,D については,C1,D1 のコドラートは斜面上に設 置し,コドラート番号の順に斜面が緩やかになり,C3 以降, D4 以降はほぼ平坦となる。斜面上部に位置する C1,C2, D1,D2 付近は,シラカシなどの常緑広葉樹が多く,平坦面 部のコドラートよりも暗くなっている。調査区E は,平坦な
地形で,コクサギ(Orixa japonica Thunb.),ミズキ(Cornus controversa Hemsl. ex Prain),アブラチャンなど構成樹種の すべてが落葉広葉樹である。そのため,樹木の構成種は異な るものの,調査区A に近い環境といえる。 2.4 調査方法 1) 群落調査 コドラート内に見られる植物種とその植被率,他に全植被 率や群落高を定期的に記録した。植被率及び全植被率は1 % 以下,5 %,以降は 10 %刻みで記録した。群落高に関しては コドラート内でもっとも植被率が高い種について,平均的な 高さの個体を目視で選択し,10 cm 刻みで記録した。調査は 2015 年の 4 月から 12 月の間,毎月 1 回ずつ行った。 2) 光環境調査 各コドラートの光環境を把握するために,春季(2015 年 3 月)と夏季(2015 年 9 月)の相対光量子束密度を求めた。被陰さ れない条件の場所として,古淵鵜野森公園の近くのマンショ ン屋上にロガー(LI-1400)と光量子センサーを設置し,調査の 間30 秒間隔で計測した。 一方で現地での測定では,2 m×2 m のコドラートを 9 等 分し,ライトメーター(LI-250)を用いてそれぞれの光量子束 密度の瞬間値を計測し,時刻と共に記録した。計測する高さ に関しては,春季(3 月)の調査では春植物に焦点を当てるた め,地上から5 ~10 cm ほどで計測した。夏季(9 月)の調査 では,夏季に群落内の多くを占める背の高い草本層に焦点を 当てるため,高茎草本と同程度の高さ (150 cm ほど)で計測 した。現地で測定した光量子束密度(µmol/m2/s)は,計測時刻 で照らし合わせ,ロガーで継続的に計測した光量子束密度と の商を求め,相対光量子束密度(%)(以下光量)とした。 2.5 解析 各コドラートを種組成からタイプ分けし,その傾向を把握 するため,夏季の種組成および植被率によるクラスター分析 を行った。また,ブラウン−ブランケ法における被度階級Ⅱ (10-25%)を基準とし,4 月の調査結果で春植物の植被率が合 計10 %以上を占めるコドラートを選択して解析を行った。ま ず森下の Cλ指数4)に基づいたコドラート間の類似度指数を 算出し,その結果をもとにクラスター分析を行った。分析に はCλの非類似度を距離としてウォード法を採用し,解析ソ フトにはPC-ORD ver.6.08 を使用した。なお,異なるフェノ ロジーを持つ種による影響を除くため,6 月~9 月の非類似 度を平均して用いた。 クラスター分析で分類したパターンの違いを決定づける要 因を把握するため,夏季の群落データを DCA により序列化 し,各軸と光環境とのスピアマンの順位相関係数を求めた。 また,春植物が存在するコドラートは,春季において他のコ ドラートに比べて光量が高いかどうか,t 検定で評価した。 これらの統計解析についてはR(ver.3.13)を用いた。 3. 結果 3.1 植生 表-1 に各コドラートの 4 月から 9 月までの春植物と主な優 占種の植被率の季節変化を示す。なお,優占種は植被率と出 現頻度から上位5 種を選出した。 春植物は,ニリンソウ,イチリンソウ(Anemone nikoensis Maxim.) , ジ ロ ボ ウ エ ン ゴ サ ク (Corydalis decumbens (Thunb.) Pers.),分布は限られているがヒメニラ,レンプク ソウがみられた。
A1 ではイチリンソウとニリンソウ,A2,A3 ではニリンソ
ウが確認された。また,A2 ではジロボウエンゴサクとヒメニ
ラも確認された。これらのコドラートでは春植物以外ではハ ナ ウ ド(Heracleum sphondylium L. var. nipponicum (Kitag.) H.Ohba)が主な優占種となっており,春植物の地上 調査区 A 調査区 A 調査区 B 調査区 E 調査区 D 調査区 C 神奈川県 神奈川県 81 春植物の地上部消失後の植生と光環境
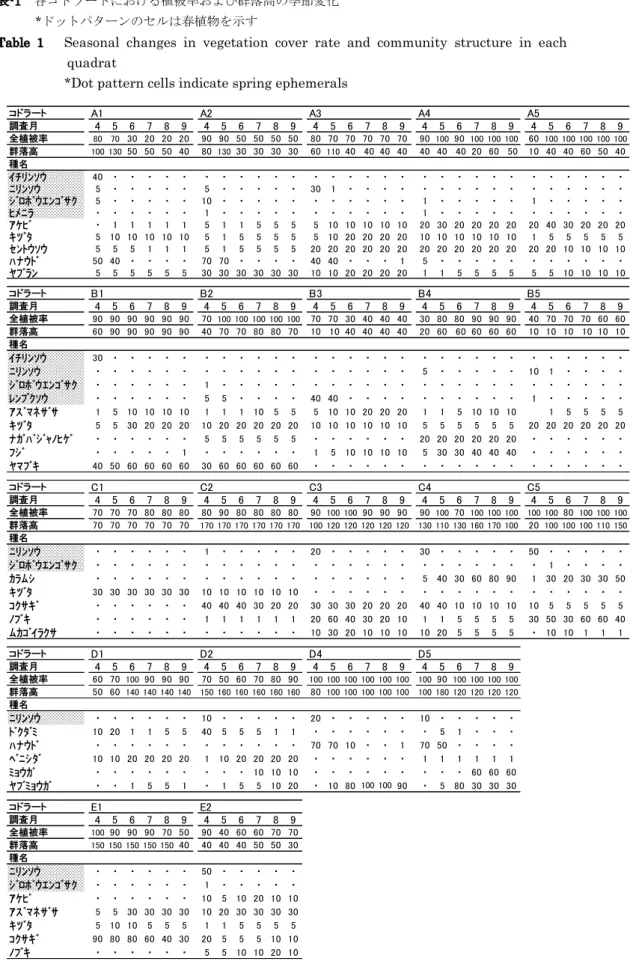
表-1 各コドラートにおける植被率および群落高の季節変化 *ドットパターンのセルは春植物を示す
Table 1 Seasonal changes in vegetation cover rate and community structure in each quadrat
*Dot pattern cells indicate spring ephemerals
コドラート A1 A2 A3 A4 A5 調査月 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 全植被率 80 70 30 20 20 20 90 90 50 50 50 50 80 70 70 70 70 70 90 100 90 100 100 100 60 100 100 100 100 100 群落高 100 13050 50 50 40 80 130 30 30 30 30 60 110 40 40 40 40 40 40 40 20 60 50 10 40 40 60 50 40 種名 イチリンソウ 40 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ニリンソウ 5 ・ ・ ・ ・ ・ 5 ・ ・ ・ ・ ・ 30 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ジロボウエンゴサク 5 ・ ・ ・ ・ ・ 10 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ ヒメニラ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ アケビ ・ 1 1 1 1 1 5 1 1 5 5 5 5 10 10 10 10 10 20 30 20 20 20 20 20 40 30 20 20 20 キヅタ 5 10 10 10 10 10 5 1 5 5 5 5 5 10 20 20 20 20 10 10 10 10 10 10 1 5 5 5 5 5 セントウソウ 5 5 5 1 1 1 5 1 5 5 5 5 20 20 20 20 20 20 20 20 20 20 20 20 20 20 10 10 10 10 ハナウド 50 40 ・ ・ ・ ・ 70 70 ・ ・ ・ ・ 40 40 ・ ・ ・ 1 5 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ヤブラン 5 5 5 5 5 5 30 30 30 30 30 30 10 10 20 20 20 20 1 1 5 5 5 5 5 5 10 10 10 10 コドラート B1 B2 B3 B4 B5 調査月 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 全植被率 90 90 90 90 90 90 70 100 100 100 100 100 70 70 30 40 40 40 30 80 80 90 90 90 40 70 70 70 60 60 群落高 60 90 90 90 90 90 40 70 70 80 80 70 10 10 40 40 40 40 20 60 60 60 60 60 10 10 10 10 10 10 種名 イチリンソウ 30 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ニリンソウ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 5 ・ ・ ・ ・ ・ 10 1 ・ ・ ・ ・ ジロボウエンゴサク ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ レンプクソウ ・ ・ ・ ・ ・ ・ 5 5 ・ ・ ・ ・ 40 40 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ アズマネザサ 1 5 10 10 10 10 1 1 1 10 5 5 5 10 10 20 20 20 1 1 5 10 10 10 1 5 5 5 5 キヅタ 5 5 30 20 20 20 10 20 20 20 20 20 10 10 10 10 10 10 5 5 5 5 5 5 20 20 20 20 20 20 ナガバジャノヒゲ ・ ・ ・ ・ ・ ・ 5 5 5 5 5 5 ・ ・ ・ ・ ・ ・ 20 20 20 20 20 20 ・ ・ ・ ・ ・ ・ フジ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ ・ 1 5 10 10 10 10 5 30 30 40 40 40 ・ ・ ・ ・ ・ ・ ヤマブキ 40 50 60 60 60 60 30 60 60 60 60 60 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ コドラート C1 C2 C3 C4 C5 調査月 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 全植被率 70 70 70 80 80 80 80 90 80 80 80 80 90 100 100 90 90 90 90 100 70 100 100 100 100 100 80 100 100 100 群落高 70 70 70 70 70 70 170 170 170 170 170 170 100 120 120 120 120 120 130 110 130 160 170 100 20 100 100 100 110 150 種名 ニリンソウ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ 20 ・ ・ ・ ・ ・ 30 ・ ・ ・ ・ ・ 50 ・ ・ ・ ・ ・ ジロボウエンゴサク ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ カラムシ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 5 40 30 60 80 90 1 30 20 30 30 50 キヅタ 30 30 30 30 30 30 10 10 10 10 10 10 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ コクサギ ・ ・ ・ ・ ・ ・ 40 40 40 30 20 20 30 30 30 20 20 20 40 40 10 10 10 10 10 5 5 5 5 5 ノブキ ・ ・ ・ ・ ・ ・ 1 1 1 1 1 1 20 60 40 30 20 10 1 1 5 5 5 5 30 50 30 60 60 40 ムカゴイラクサ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 10 30 20 10 10 10 10 20 5 5 5 5 ・ 10 10 1 1 1 コドラート D1 D2 D4 D5 調査月 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 4 5 6 7 8 9 全植被率 60 70 100 90 90 90 70 50 60 70 80 90 100 100 100 100 100 100 10090 100 100 100 100 群落高 50 60 140 140 140 140 150 160 160 160 160 160 80 100 100 100 100 100 100 180 120 120 120 120 種名 ニリンソウ ・ ・ ・ ・ ・ ・ 10 ・ ・ ・ ・ ・ 20 ・ ・ ・ ・ ・ 10 ・ ・ ・ ・ ・ ドクダミ 10 20 1 1 5 5 40 5 5 5 1 1 ・ ・ ・ ・ ・ ・ ・ 5 1 ・ ・ ・ ハナウド ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 70 70 10 ・ ・ 1 70 50 ・ ・ ・ ・ ベニシダ 10 10 20 20 20 20 1 10 20 20 20 20 ・ ・ ・ ・ ・ ・ 1 1 1 1 1 1 ミョウガ ・ ・ ・ ・ ・ ・ ・ ・ ・ 10 10 10 ・ ・ ・ ・ ・ ・ ・ ・ ・ 60 60 60 ヤブミョウガ ・ ・ 1 5 5 1 ・ 1 5 5 10 20 ・ 10 80 100 100 90 ・ 5 80 30 30 30 コドラート E1 E2 調査月 4 5 6 7 8 9 4 5 6 7 8 9 全植被率 10090 90 90 70 50 90 40 60 60 70 70 群落高 150 150 150 150 15040 40 40 40 50 50 30 種名 ニリンソウ ・ ・ ・ ・ ・ ・ 50 ・ ・ ・ ・ ・ ジロボウエンゴサク ・ ・ ・ ・ ・ ・ 1 ・ ・ ・ ・ ・ アケビ ・ ・ ・ ・ ・ ・ 10 5 10 20 10 10 アズマネザサ 5 5 30 30 30 30 10 20 30 30 30 30 キヅタ 5 10 10 5 5 5 1 1 5 5 5 5 コクサギ 90 80 80 60 40 30 20 5 5 5 10 10 ノブキ ・ ・ ・ ・ ・ ・ 5 5 10 10 20 10 82 大塚 勇哉・倉本 宣
部が消える頃にハナウドの植被率と群落高がピークに達して
いた。ハナウドが枯れる6 月以降は,優占種はないか,ヤブ
ラン(Liriope muscari (Decne.) L.H.Bailey)のような常緑の
草本が残った。一部は表-1 にも挙げられているが,他にセン
トウソウ(Chamaele decumbens (Thunb.) Makino)やジャノ ヒゲ(Ophiopogon japonicus (Thunb.) Ker Gawl.),アオキ (Aucuba japonica Thunb. var. japonica),アケビ(Akebia quinata (Houtt.) Decne.)も多く見られた。しかしいずれも植
被率は低く,背の高い草本もないので,A1~A3 の夏季におけ る全植被率は平均50 %程度,群落高は平均 39 cm 程度であ った。調査区B ではニリンソウは B4, B5 に各 5, 10%(4 月) みられるのみであった。イチリンソウを30 %程度含む B1 で は,イチリンソウが消えたタイミングでヤマブキ(Kerria japonica (L.) DC.)が展葉し,以降秋まで覆い続けており,5 月以降全植被率は90 %,群落高は 90 cm から変化していな い。古淵鵜野森の調査区C,D では,斜面上部(C1,D1)には ニリンソウは存在しなかったが,それ以外の箇所すべてで確 認された。C3・C4 では落葉低木のコクサギも優占種として 挙げられており,春季でも群落高が100 cm を越えている。 これらのコドラートでは夏になるとカラムシ(Boehmeria
nivea (L.) Gaudich. var. concolor Makino f. nipononivea (Koidz.) Kitam. ex H.Ohba)あるいはノブキ(Adenocaulon himalaicum Edgew.)がコドラートを覆う。そのため夏季(6 月~7 月)の全植被率は 70 ~100 %,群落高は 100 cm 以上で
あった。D4,D5 では,春季にハナウドが優占する点で,コ
ドラートA1,A3 と同様だが,ハナウドが枯れた後,ヤブミ
ョウガ(Pollia japonica Thunb.)やミョウガ(Zingiber mioga (Thunb.) Roscoe)がコドラート全体を被覆する点で異なって いる。調査区C 同様,全植被率 90 ~100 %,群落高も 100 cm 以上とともに高いのはこのためである。E2 ではニリンソウが 50 %と高い植被率で確認されている。夏季には調査区 A の コドラートと同様,林冠が閉鎖され暗くなり,調査区 C,D のように優占するような草本は見られず,夏季における全植 被率は60 ~70 %程度,群落高は 30 ~50 cm であった。 3.2 光環境 春季と夏季の各コドラートの光量を図-2 に示す。全体の平 均光量(±標準偏差)は,3 月では 33 %(±13.7),9 月では 7 %(±6.1)であった。また,ニリンソウ,イチリンソウ,ジロ A1 A2 A3 A4 A5 B1 B2 B3 B4 B5 C1 C2 C3 C4 C5 D1 D2 D4 D5 E1 E2 0 10 20 30 40 50 60 70 コドラート 相対光量子束密度 (% ) A1 A2 A3 A4 A5 B1 B2 B3 B4 B5 C1 C2 C3 C4 C5 D1 D2 D4 D5 E1 E2 0 5 10 15 20 コドラート 相対光量子束密度 (% ) 図-2 春季と夏季における相対光量子束密度 *上の横線から最大値,第 3 四分位数,中央値,第 1 四分位数,最小値を示す。図中の丸印は外れ値を表す。 Fig. 2 Relative photosynthetic photon flux density in spring and summer
* Horizontal lines indicate, from top to bottom, the sample maximum, the upper quartile, the median, the lower quartile, and the sample minimum. Circles in the figure indicate outlier.
3 月 March 9 月 September 83 春植物の地上部消失後の植生と光環境
図-3 クラスター分析によるコドラートの分類 Fig.3 Classification of quadrats by cluster analysis
ボウエンゴサクを合計 10 %以上含むコドラート(A1~A3, B1,C3~C5,D2~D5,E2)では春季の平均光量 39 %(±11.7) であり,それ以外のコドラートの平均光量25 %(±12.8)に比 べて有意に高かった(p<0.01)。春植物がまったく分布しない C1,D1,E1 の春季の平均光量は 19 %(±7.1)程度であった。 調査区A 全体での光環境は春季に平均 50 %(±9.2)の光量 が記録されているが,夏季には平均光量3 %(±1.1)と対称的 に暗くなっていた。調査区B は春季,夏季通して他の調査区 に比べて暗く,イチリンソウが分布するB1 のみ春季は平均 光量33 %(±10.3)だが,それ以外は平均光量 17 %(±5.6)で あり,夏季になると全体で平均光量3 %(±1.2)まで低下した。 また,C3,C4,C5,D4,D5 の夏季に特定の草本が優占す るコドラートでは,春季は平均光量39 %(±4.0),夏季におい ても平均17 %(±3.7)の光量を示していた。これらを除いたコ ドラートの夏季の平均光量は4 %(±2.4)であり,夏になり全 体的に光量が下がる中でもこれらのコドラートは比較的明る いといえる。ニリンソウの 50 %と高い植被率が確認された E2 では,春季は平均光量 34 %(±10.4),夏季には平均光量 4 %(±1.3)と,調査区 A と同様の光環境の変化がみられた。 3.3 植生と光環境 クラスター分析結果のデンドログラムを図-3 に示す。クラ スターはまず大きく 2 つのグループに分かれた。ひとつは A1,B3,B5,E2,A2,A3 を含む,夏になると光量が急減 する(平均光量 3 %)グループ(タイプ 1 とする),もうひとつは C3,C4,C5,D2,D4,D5 を含む,夏季に光量が急減しな い (平均光量 17 %)グループ(タイプ 2)である。そこからさら に区切ると前者のクラスターからは3 グループ,後者からは 2 グループに区分された。すなわち,春植物が消えた後は特 に何も生えず植生が変化しない A1,B3,B5,E2 のグルー プ(タイプ 1-1),一部の夏緑性草本は生えてくるが,それほ ど植生が変化しないA2,A3 のグループ(タイプ 1-2),ヤマ ブキが秋まで覆い続けるB1 のみのグループ(タイプ 1-3), ノブキ,カラムシなどが優占し覆っているC3,C4,C5 を 含むグループ(タイプ 2-1)とヤブミョウガおよびミョウガが 主に優占するD2,D4,D5 のグループ(タイプ 2-2)である。 DCA スコア第 1 軸と第 2 軸の散布図を図-4 に示す。各コ ドラートの種組成のパターンはクラスター分析の結果にタイ プ1 とタイプ 2 のコドラート群は,DCA スコアにおいても 従った分布を示した。デンドログラムにおける最初の分離, 明確に分離していることがわかる(図-4)。タイプ 1 は第 1 軸 の数値が高い方に,タイプ2 は第 1 軸の数値が低い方に位置 づけられた。また,第1 軸と夏季の光量の間には高い負の相 関(ρ= -0.791, p<0.001)がみられた。全植被率および群落高 においても,光量とDCA 第 1 軸ともに負の相関があった(表 -2)。また,第 2 軸は光量と弱い相関がみられるが,今回計測 したのは光環境のみであり,より高い相関を持つ他の環境要 因が存在する可能性がある。以上のことから,タイプ1 とタ イプ2 における植生の変化のパターンは光環境が大きな要因 であることが示唆された。 4. 考察 B3 と B2 には,ニリンソウ,イチリンソウは分布しないが, レンプクソウは見られた。これらのコドラートは,ほかの春 植物が分布しているコドラートよりも暗く,レンプクソウは 注:**は 1%有意水準,*は 5%有意水準 光量(夏) 全植被率 群落高 第 1 軸 -0.79** -0.63** -0.68** 第 2 軸 -0.37** -0.46** -0.47** 第 1 軸 図-4 各コドラートの DCA による序列化 Fig.4 DCA ordination diagram of each quadrat
表-2 DCA スコアと各要因との順位相関係数
Table 2 Spearman's rank Correlation Coefficients between DCA score and each variable 0 100 200 0 100 200 300 400 Axis1 Axi s2 plot2 A1 A2 A3 B1 B3 B5 C3 C4 C5 D2 D4 D5 E2 第 1 軸 第 2 軸 タイプ 1 タイプ 2 84 大塚 勇哉・倉本 宣
耐陰性がある,あるいは暗い場所を好むと推定される。C3, C4,C5 の春季における優占種はコクサギであった。また, 優 占種ではないものの,B5 および D4,D5,E1,E2 でもコク サギが確認された。このコクサギはカタクリを指標する植物 として挙げられており 5),ニリンソウにおいても同様に指標 できるものと考えられる。 管理ボランティアへの聞き取りによると,古淵鵜野森の調 査区C,D 付近では,数年前に樹木の伐採が行われている。 そのため,平面部のC3~C5,D4,D5 付近の林冠は開けて おり,夏季の林床は平均光量 17 %(±3.7)で,それ以外のコ ドラートの平均光量5 %(±2.4)に比べても平均光量 12%ほど の差がみられた。このことは光量の確保にもつながり,春季 に春植物の植被率を高める効果があると考えられるが,一方 で夏季にノブキ,カラムシ,ヤブミョウガなどが優占する一 因となっていると考えられる。調査区Aでは春季は明るいが, 夏季になると林冠が閉鎖され,十分な光量が林床に差し込ま ない。周辺の樹種構成がほぼ落葉広葉樹であることと,調査 区C,D に比べ,それらが葉をつけた際に林冠を覆うように 密に生えているためである。また,調査区B では常緑針葉樹 が多く分布しており,春季,夏季通して他の調査区に比べ暗 い。そのため,植生は春植物の地上部が消えたあとも大きく 変化することはなく,少量の夏緑性草本が生えてくるか,常 緑の草本が維持されて残るのみである。 クラスター分析では,2 つの群落タイプが確認された。ほ とんどは調査地の違いにより二分されたが,E2 のみ高木道 正山と同じクラスターに吸収された。これらタイプ1 とタイ プ2 の区分は,図-4 および表-2 より光環境によるものだと推 察できる。しかし,さらにその先の区分,1-1,1-2,1-3 お よび2-1,2-2 の分類は光環境だけで説明するのは難しい。1-1 と1-2 の分類は春植物の地上部が消えてから植生はほぼ変化 しないか,夏緑性草本がわずかに生えてくるかの違いである。 しかし,夏季の平均光量は1-1 では 3 %(±1.3),1-2 では 2.2 %(±0.5)であり,むしろ夏緑性草本が分布する 1-2 の方が暗 いため,光環境が影響しているとは考えにくい。また,1-3 はヤマブキのみが優占していることから,単独で別のグルー プに区分されたが,ヤマブキが優占する要因は光環境だけで なく今回計測していない別の環境要因が複雑にからみ合って 成立しているものと考えられる。一方,2-1 と 2-2 の区分は, 優占する種の違いである。図-4 より,その区分は第 1 軸より も第2 軸の方が影響していそうであるが,第 2 軸は光量とは 弱い相関しかみられず(表-2),土壌水分量や地温など他の環 境要因が影響している可能性が高い。 今回の研究では2-1 と 2-2 の種組成の違いに影響する要因 を明らかに出来なかったが,夏季の光量が高く,背の高い草 本が優占するという点で,本研究においては本質的には同じ タイプと結論付ける。したがって,春植物の地上部が消えた 後の植生のパターンとしては,特定の優占種は存在しないタ イプ(1-2),そしてほぼ何も生えてこず,植生が変化しないタ イプ(1-1,1-3),そして高茎草本などの特定の種が優占する タイプ(2-1,2-2)の 3 つのパターンが存在し,特に植生が変 化しないグループ(タイプ 1)と高茎草本が優占するグループ (タイプ 2)の種組成の違いには光環境が影響している可能性 があるということがわかった。 5. おわりに 植生については,春植物の地上部が消えた後,夏に特定の 草本が優占するタイプと,特定の優占種がなく植被率の低い タイプ,そしてほぼ何も生えてこないタイプの3 種類の植生 の変化がみられた。これらは主に光環境によるものであり, 光量次第では春植物と同じ空間を夏緑性の草本が夏から秋に かけて利用する,季節のすみわけを行っていることが明らか になった。今後の課題として,夏緑性草本が春植物とすみわ ける,共存のメカニズムを把握する必要がある。また,春植 物が休眠している間の地下部での根の分布や相互作用など, さらなる検討が必要である。 謝辞: 本研究を行うにあたり,NPO 法人境川の斜面緑地を 守る会の方々には多くの協力をいただいた。この場を借りて, 感謝の意を表したい。 引用文献 1) 倉本宣 (1984) 都市公園における春植物ニリンソウ保全の ための基礎的研究. 造園雑誌, 47(5): 101-105. 2) 鈴木由告 (1986) 多摩川中流域におけるカタクリ群落の分 布と生態および保護育成に関する研究. 公益財団法人とう きゅう環境財団 学術研究, 87: 1-127. 3) 宮本太・桑原美代子 (2009) 東京農業大学厚木キャンパス (神奈川県)における早春植物ニリンソウ(Anemone flaccida Fr. Schm.)の季節消長とその生育環境. 東京農大農学集報, 54 (3): 175-181.
4) Morisita, M. (1959) Measuring of interspecific association and similarity between communities. Men. Fac. Sci. Kyushu Univ. Ser. E.(Biol.), 3: 65-80.
5) 養父信乃夫 (1988) カタクリ個体群の形成並びにその個体 群の育成管理上の指針. 造園雑誌, 51(4): 228-236. 6) 屋 祢 下 亮 ・ 北 脇 優 子 ・ 猪 熊 千 恵 (2014) ニ リ ン ソ ウ (Anemone flaccida Fr. Schm.)の生育に影響を及ぼす環境 要因. 日緑工誌, 40(1): 243-246. (2016 年 6 月 21 日受理) 第 1 軸 タイプ 2 85 春植物の地上部消失後の植生と光環境