iじ
課 程 博 学 位 論 文
︑・.ノ
逢﹂
・生
日員丁牛㌃耳41〆
r・.
1‑,.‑lr̲1.{一一 一 .一ゴー 」̲
三 一 、惑 一 ノ 、\ す.一
{/一
﹂⊥
邪 京 東
コケ仁ケ\伝田二詳川
接 触 お よ び 光刺 激 に よ って 誘 導 され る
葉緑 体運動の解析
ニレく τヨ
ロ
韮臼一.
勝
良
一f‑t一
謄
佐
ヒヨ
π̀τ
ノヤコ 吐÷
冒一トレコ査審
矯 〜 /中
.!ノ
π1.留 一
ノ.
イ
l!.い 〕
!'
・/\
﹁︑9
、 r昌1
コ
〆?
/\i
9
員
一一 求一‑A丁 一
一
一̀
,
.'
の
レL、 .7
̀.ブ
fi午
ロキも口呉
委 昌へ
.一 「 一
妻
)
す 定 判
と各L智が﹁
︑電﹂
△に
ヒ 言己 σつ 言命 .:'J〔∠を
日6
ヨロリ一=︑rトノ
3
一二丁.︑ノ⊥
﹂ 戎
/3
ア一qコ
東 京 都 立 大 学 大 学 院 理 学 研 究 科 教 授 会
へG
り,一
夕 添
︑ー
︑19
そ
}\
タタ
で
長研 究 科
m In ri 1-) —
U LTJQN
n'0TDT-r‘ 1L-;fCE
TOKYO METROPOLITAN m -TiANT -1-1\TT-,.\/:117DCTT
TITLE: Dire
ctional movement of chloroplasts
is induced by mechanical stimulation as well as light stimulation
ALITIOR:
Yoshikatsu Sato
ETAMINFD BY
Examiner
mi mci
t-,xanuner
ill chief/
( • )
QUALIFIED BY THE GRADUATE SCHOOL OF SCIENCE
TOKYO METROPOLITAN UNIVERSITY
Dean /4..
Date "X,1 fr(_A.
Directional movement of chioroplasts
is induced by
mechanical stimulation as well as light stimulation
Yos h i kats u Sato
A dissertation presented to the Faculty of the Graduate School of the Tokyo Metropolitan University in candidacy for the Degree of Doctor of Science
ACKNOWLEDGMENTS
My thanks are due to Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists and the Sasagawa Scientific Research Grant from the Japan Science Society.
I am deeply indebted to Prof. Wada under whose guidance this study has been carried out. His interest in my work was a great encouragement.
His criticism and advice during this study are most greatly acknowledged.
I especially appreciate Associate Prof. Kadota for his valuable suggestions, completion of this thesis, and continuous encouragement throughout the study.
I am also grateful to Assistant Prof. Kanegae for his thoughtful suggestion and encouragement.
I greatly appreciate to Mr. Imaizumi for his warm encouragement and his kind friendship.
Special thanks must be given to Prof. Kazama of the International Christian University for discussion and encouragement during the course of this study.
I also thank Mr. Sugiyama and Ms. Yasuki for their kindness to supply spores of ferns throughout the present study.
I thank Prof. Hasebe of National Institute of Basic Biology for the gift of Physcomitrella patens.
I wish to express my gratitude to other members of Laboratory of Plant
TABLE OF CONTENTS
GENERAL INTRODUCTION
Section I:
veneris ... ...
Directional ment of chloroplasts in the fern Adiantum capillus- 1
Chapter 1.
stimulation
1-1. Abstract
1-2.
1-3.
1-4. Results.
1-5.
Chapter 2. Roles of e>
relocation
2-1. Abstract
2-2.
2-3.
2-4. Results..
Chloroplast relocation movement induced by mechanical
Introduction
Materials and methods
6
7
7
8
8
Discussion
Roles of external Ca2+ in signaling of mechano- and photo-
relocation of chloroplasts ...
Introduction
Materials and methods
13
29
34
34
36
36
40
-ii-
2-5. Discussion 52
Section II: Directional movement of chloroplasts in the moss
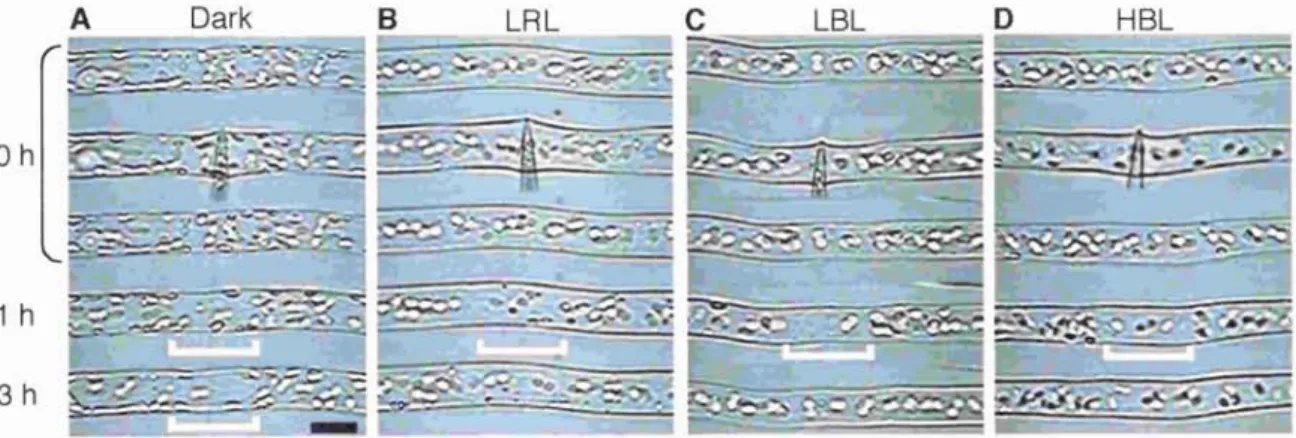
Chapter 3. Chloroplast relocation movement induced by light stimulation Physcomitrella patens
3. Chloroplast relocatioi
3-1. Abstract ... .
3-2.
3-3.
3-4.
3-5.
Materials and methods Introduction
Materials ar
Results...
Discussion
58
59
59
60
60
64
Chapter 4. Chloroplast relocation movement induced by mechanical
stimulation.... ...
82
4-1.
4-2.
Abstract
Introduction
Materials and methods...
4-4. Results and discussion ... .
CONCLUDING REMARKS...
91
91
92
93
95
REFERENCES
104
109
ABBREVIATIONS
APM: amiprophosmethyl
BL: blue light
BDM: 2, 3-butanedione monoxime
[Cal: cytosolic Ca2+ concentration HBL: high fluence rate blue light HFR: high fluence rate response HRL: high fluence rate red light IR: infrared light
LBL: low fluence rate blue light LFR: low fluence rate response LRL: low fluence rate red light
MS solution: modified Murashige and Skoog's mineral salt solution MF(s): actin microfilament(s)
MT(s): microtubule(s)
PBS: phosphate buffered saline
PIPES: piperazine-N,N'-bis (2-ethane-sulfonic acid) RL: red light
-iv-
GENERAL INTRODUCTION
Immobile plants are obliged to suffer from environmental changes and must cope with them to survive, while animals can move away from the hazard condition to the appropriate location. Among many environmental factors, light and mechanical stimulation are the most significant stresses, and plants use these as signals for regulation of their own development and morphology to adapt to the environment. Responses caused by these two environmental factors are known as a term of photomorphogenesis and of thigmomorphogenesis (Jaffe, 1973; Kendrick and Kronenberg, 1994). In addition to the macroscopic responses, such as the morphology of individuals, plants also respond rapidly and reversibly at microscopic level to these fluctuant signals. At the subcellular level, the movement of intracellular organelles is very active and plant cells adapt to various environmental stimuli by changing their distribution. One of the most prominent phenomena is a directional
movement of chloroplasts.
1. Light-induced chloroplast movement
Light plays an important role in the life of a plant; namely, it is used for an environmental signal as well as for the energy source. Directional movement of chloroplasts induced by light has been known over a hundred years and was
Chloroplasts move to appropriate position where they can perceive light of moderate fluence rate as much as possible for efficient photosynthesis, but they move away from the place of potentially hazardous light condition where a fluence rate is too strong. Chloroplast photo-movement is one of the responses
observed in cells of many species and is known to occur throughout the plant kingdom. Studies on the response are divided into two main areas: the light perception mechanism and the force generation system for movement.
1-1. Photoreceptors involved in chioroplast movement
The direction of chioroplast movement, whether toward or away from the light, depends on light intensity of effective wavelength. There are two types of photo-sensory pigment involved in chioroplast relocation movement. In most plants, the effective wavelength of light for chioroplast translocation is restricted to the blue band region of the spectrum, and the response is controlled by a blue light (BL) receptor (Zurzycki, 1980). In some plant species, such as the fern Adiantum capillus-veneris and the green alga Mougeotia scalaris, red light (RL) as well as BL also induces chioroplast movement. The response to RL is mediated by another photoreceptor, phytochrome (Haupt and Scheuerlein, 1990; Wada et al., 1993). Recently, we showed for the first time in bryophytes that phytochrome as well as a BL receptor also regulates chioroplast relocation in the moss Physcomitrella patens (Kadota et al., 2000).
-2-
1-2. Motile system of chioroplast movement
The involvement of two types of motility system is well known for organelle movement in eukaryotic cells. One is a microtubule (MT)-based system and the other an actin microfilament (MF)-based system. Force-generation in chioroplast movement has been investigated in various plant cells. The mechanism of movement in all cells so far investigated, where chloroplasts show directional movement toward or away from the light, has been considered as an exclusively MF-based system but not MT-based system (Wagner et al., 1972;
Izutani et al., 1990; Kadota and Wada, 1992; Tlalka and Gabrys, 1993). On the contrary, organelle movement and vesicle transport in animal cells had been considered to be an exclusively MT-based system for many years. Since Kuznetsov et al. (1992) showed that single axoplasmic organelles move along both MTs and MFs tracks in vitro, however, previous views of the roles of the two cytoskeletal filaments have been changing, and it now appears that they functionally cooperate during organelle transport (Langford, 1995; Goode et al ., 2000; Rogers and Gelfand, 2000). It seems clear that melanosome movement and axonal transport of mitochondria involves both MTs and MFs in vivo, although they had previously been thought as a model system of MT-based movement (Morris and Hollenbeck, 1995; Rodionov et al., 1998; Rogers and Gelfand, 1998; Bridgman, 1999). Rodionov et al . (1998) mentioned the possibility that the coordinated action of these filaments might be a general
2. Mechanically induced chloroplast movement
As mentioned above, mechanical stresses, such as touch, rain and wind, are important natural factors in the life of a plant because they can physically threaten survival at any time. A number of physiological responses caused by mechanical stimulation have been known for a range of plants since the last century (Darwin, 1882). However, studies of these phenomena at the cellular or subcellular level have been limited because multicellular tissues of higher
plants have been used as experimental systems.
To elucidate the mechanisms of mechano-perception and mechano- transduction in plants, it is necessary to know how cells respond to mechanical stimulation at the subcellular level. A few studies of this nature have been
performed. In a dark-grown, suspension-cultured parsley cell, intracellular nuclear migration toward the site of fungal penetration has been studied (Gross et al., 1993). In epidermal cells of Tradescantia virginiana, nuclear movement toward a site touched with a microneedle was recently reported (Kennard and Cleary, 1997) . No other organelles, even chloroplasts, whose movement can be induced readily by light (Nagai, 1993; Wada et al., 1993; Williamson, 1993) were found to move in response to mechanical stresses in vascular plants.
In my master course, I discovered the mechanically induced chloroplast movement in protonemal cells of A. capillus-veneris (Chapter 1).
-4-
3. The purpose and the constitution of this thesis
This thesis is divided into two parts. In Section I, chloroplast relocation movement was investigated in the fern A. capillus-veneris. Directional movement of chloroplasts is induced by mechanical stimulation as well as light stimulation in fern A. capillus-veneris. To elucidate the regulation of chloroplast
movement, motile system responsible for the mechano-relocation was examined (Chapter 1). Subsequently, I studied on the relationship of mechano- and photo-relocation movement of chloroplasts (Chapter 2). In section II, using the moss P. patens, I investigated the coordinated action of MFs and MTs for chloroplast movement (Chapter 3). In Chapter 4, mechano-relocation of chloroplasts was examined in P. patens. To examine the signal tranduction pathway, requirements of external Ca' on mechano- and photo-relocation movement of chloroplasts were also examined in both cells of the fern and the moss (Chapter 2, 4).
Section I: Directional movement of chioroplasts
in the fern Adiantum capillus-veneris
-6-
Chapter t stimulation
Chloroplast relocation movement induced by mechanical
Abstract
Cell response to mechanical stimulation was investigated at a subcellular level in protonemal cells of the fern Adiantum capillus-veneris by means of pressing a small part of the cell with a microcapillary. In cells receiving local stimulation, the chioroplasts moved away from the site of stimulation, whereas the nuclei failed to show such avoidance movement. Mechanical stimulation for a period as short as 0.3 min was enough to induce the avoidance response to a maximal level. The avoidance movement of chloroplasts started within 30 min and the plateau level of avoidance was attained around 2 h after stimulation. By tracing the movement of chloroplasts during the response, it was shown that the mobility of chioroplasts near the stimulation site increased transiently within 1 h after the stimulation. After 2 to 3 h, it slowed down to the control level without stimulation.
The avoidance response was inhibited by 0.1 mM cytochalasin B and 25 mM 2,3-butanedione monoxime (BDM) but not by 3.3 pM amiprophosmethyl (APM) or 5 mM coichicine. These findings indicate that the protonemal cells are very sensitive to mechanical stimulation and that chloroplasts move away from the mechanically stimulated site through the actomyosin motile system.
Introduction
Protonemal cell of a fern, A. capillus-veneris, has a simple structure of linearly- arranged cells. In these cells, organelles, including chloroplasts, can be readily observed and chioroplast photo-movement has been examined in previous studies (Wada et al., 1993). Furthermore, mechanical stimulation can be applied to part of an intact cell by using a micromanipulator. The site of contact with a needle can be seen under the microscope as the perception site of stimulation. Thus, this type of cell appeared to be ideal for the investigation of the mechano-responses at the subcellular level. Using this system, subcellular response to mechanical stimulation was examined. I found that chloroplasts moved away from the site of mechanical stimulation induced by pressing with a microcapillary. In this chapter, the details of this phenomenon are reported.
Materials and methods
Plant materials and aseptic culture
Spores of Adiantum capillus-veneris L. were collected in a greenhouse of Tokyo Metropolitan University in 1993 and stored at about 4°C until use. Spores were sterilized for 7-8 min with a 0.5% (v/v) sodium hypochlorite solution (Wako Pure Chemical Industries, Ltd., Osaka, Japan) containing 0.1% (w/v) Triton X-100 (Wako), and washed three times with sterilized distilled water. The sterilized spores were sown in a line between two layers of agar-gelatin film on a cover slip. The film was made from 0.5% (w/v) BACTO-agar (Difco Laboratories ,
-8-
Detroit, MI) and 0.05% (w/v) gelatin (Koso Chemical Co. Ltd., Tokyo, Japan).
Spores were cultured under continuous RL of about 0.5 W m2 for 9 days in 0.1- strength, modified Murashige and Skoog's mineral salt solution (MS solution;
Wada and Furuya, 1970). The resultant protonemata were irradiated with white light of about 4.5 W m-2 for 6 h and then kept in the dark for 2 days. During the dark period, cell division occurred in the apical region of each protonema, giving rise to short apical cell and long basal cell. The basal cells of the protonemata were used for the present study. All the cultural and the experimental procedures were conducted at 25 ± 1°C.
Light sources
Fluorescent lamps (FL40SD or FL10D; Toshiba Lighting and Technology Corp., Tokyo, Japan) were used as the source of white light. Red light was obtained from the lamp through a red plastic filter (Shinkolite A, #102; Mitsubishi Rayon Co., Ltd., Tokyo, Japan).
Mechanical stimulation
Protonemata on the cover slip on which they were cultured, were placed in a hand-made chamber constructed from a glass bottomed dish and then anchored with 1% (w/v) agarose VII (Sigma Chemical Co., St. Louis, MO). This chamber was filled with MS solution and placed on a stage of an inverted microscope
condenser. Mechanical stimulation of individual cells was performed under the microscope using a microcapillary connected to a joystick-controlled micromanipulator (MO-202, Narishige Co., Ltd., Tokyo, Japan). The microcapillary was prepared from a 1.0 mm diameter borosilicate glass tube (GD-1, Narishige) using a vertical two-step puller (PP-84, Narishige). Each cell was pressed with the capillary until the deformation of the cell was observed
under the microscope. The apical 100-300 pm region of basal cell was used for this study.
Time-lapse video observation
Movement of chloroplasts was monitored under the microscope with infrared light (IR) obtained through an IR-transmitting filter (IR85; Hoya Co., Akishima, Japan). The microscope was equipped with an IR-sensitive video camera (C2400-07ER; Hamamatsu Photonics, Hamamatsu, Japan) coupled to a time- lapse video recorder (AG-6730; Panasonic, Osaka, Japan), a video monitor (PVM-14420; SONY, Tokyo, Japan) and a video copy processor (SCT-P66, Mitsubishi Electric Co., Ltd., Tokyo, Japan). For observations, a long working- distance objective (Zeiss Achroplan, x40 LD, NA 0.60) was used.
Quantification of chloroplast avoidance response
To analyze mechanically induced chloroplast relocation quantitatively, the avoidance response was determined as the percentage of chloroplasts that had
-10-
moved away from the stimulus region. The numbers of chloroplasts at the stimulus site, before and after mechanical stimulation, were counted for each cell. The following equation was used.
Avoidance response (%) = {(No - Nt)/No} x 100
where No and Nt are the numbers of chloroplasts before and t hours after mechanical stimulation, respectively within 50 pm in length of the cell (i.e. 25 µm in both the apical and basal directions from the stimulation point).
Inhibitor treatment
Cytochalasin B (Sigma) was employed as an MF-depolymerizing agent.
Amiprophosmethyl (APM) and colchicine (both from Sigma) were used to disrupt MTs. 2,3-butanedione monoxime (BDM; Sigma), known as an inhibitor of myosin ATPase, was also applied.
Cytochalasin B and APM were dissolved in DMSO as stock solutions of 10 mg ml 1 and 200 pg ml-1, respectively. Final concentrations of cytochalasin B and APM were 50 µg ml-1 (0.1 mM) and 1 µg ml-1 (approx. 3.3 µM), respectively. For treatment with BDM, culture medium containing 25 mM BDM was prepared from a 5 M stock solution in DMSO. The final concentration of DMSO was 0.5% (v/v) in all three cases. Colchicine was dissolved in deionized-distilled water as a stock solution at the concentration of 500 mM and for use, the stock solution was diluted with the culture medium to 5 mM. To
Pluronic F-127 (Sigma) was added in MS solution. Cells were incubated with each drug solution for 2 h before mechanical stimulation. After that, mechanical stimulation was applied for 1 min and the avoidance response was assessed 2 h after the stimulation.
Nucleic acid staining
Nucleic acid staining was carried out with SYTO 11 (Molecular Probes Inc., Eugene, OR). Cells were incubated with 5 pM SYTO 11 in MS solution for 1 h.
Fluorescence microscopy
Samples were mounted on a very thin cover slip of 0.06-0.08 mm in thickness (No. 00, Matsunami Glass Ltd., Osaka, Japan). The specimens were observed under a confocal laser scanning microscope (LSM 410 invert, Zeiss). The excitation wavelength for SYTO 11 was 488 nm. The beam splitter used was FT 488/543. A barrier filter, BP 515-525 and an additional beam splitter, FT560 were used for observation. Chlorophyll fluorescence, after excitation with either the 488 nm or 543 nm laser line, was obtained using OG 665 as a barrier filter.
The pinhole size was 20. Serial optical sections were obtained every 0.75 pm in the Z-axis. Images were acquired through a x 63 Zeiss Plan-Apochromat objective (NA 1.4) at a maximum resolution of 512 x 512 pixels. Every optical section was gained after averaging 4 scans.
-12-
Results
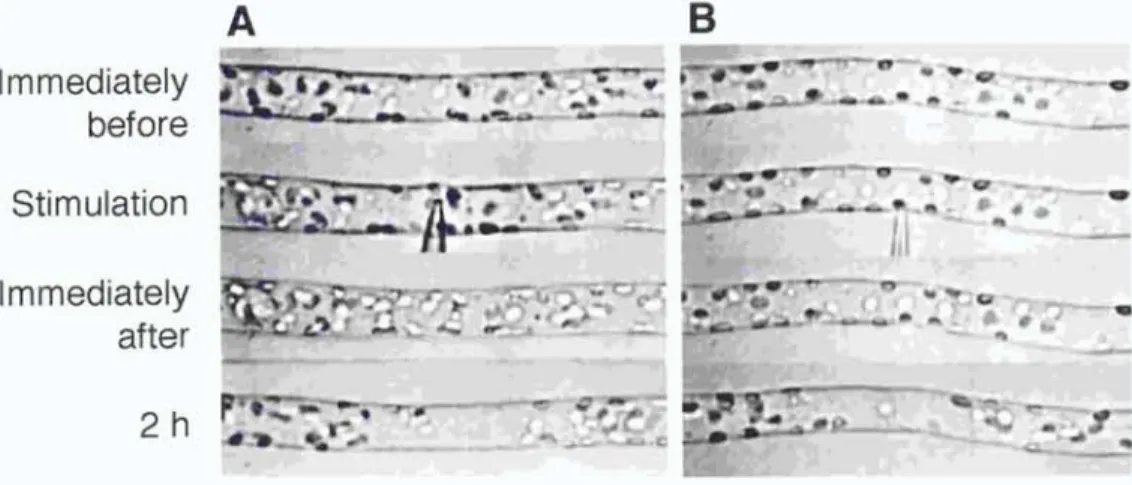
Avoidance response of chloroplasts induced by mechanical stimulation Chloroplasts are distributed randomly throughout the periphery of the long cylindrical basal cell of two-celled A. capillus-veneris protonema when cells are kept in the dark for 2 days, but relocate to the site of optimal light condition for photosynthesis when irradiated (Yatsuhashi et al., 1987). Therefore, in this study, experiments were carried out in the dark to eliminate the effects of light on chloroplast relocation. When mechanical stimulation was applied to part of the basal cell using a microcapillary, chloroplasts around the stimulated area moved away. Whenever mechanical stimulation was applied to the cell either from above with the flank of the capillary (Fig. 1A) or from the side with the tip of the capillary (Fig. 1 B), avoidance responses were observed within 2 h after the onset of mechanical stimulation. The distribution of chloroplasts immediately after the stimulation was not significantly different from that before the stimulation. Subsequently, as a result of chloroplast movement to avoid the stimulated area, a clear zone which had fewer chloroplasts compared to the adjacent areas appeared. This mechanically induced chloroplast relocation was also observed in protonemata of Dryopteris filix-mas, Onoclea sensibilis and Matteucia struthiopteris (data not shown). Mechanical stimulation in the subsequent studies was applied from above with the flank of capillary to avoid cell penetration with the tip.
Immediately before Stimulation Immediately after
2h
AB
1-'" -..1*w•.1.
14p
flti 7111 8...'"...1111---} ... --7 71,11/17— 1. .
„. 00111.ir..., ._,....,....,. rapap..i..., JA._,..., 0 ,,,,,
ga...4....„ (3T"- ,•77E wr -- - "
Sr
^ -
N••
.0
Figure 1. Avoidance response of chloroplasts induced by mechanical stimulation in A. capillus-veneris. A protonemal cell was stimulated either vertically from above with the flank of a microcapillary (A) or horizontally from side with the tip of the capillary (B). Note the deformation of the protonemal cell at the stimulation site in (A). The protonemal tip was on the right. Bar, 20 pm.
-14-
Relationship between stimulation periods and the avoidance response of chloroplasts
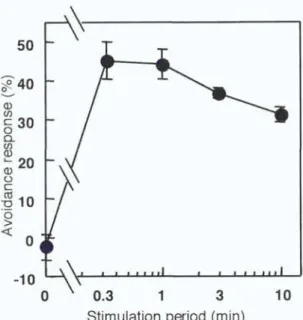
The relationship between the duration of mechanical stimulation and the degree of avoidance response of chloroplasts was investigated. Cells were stimulated for various periods of time with a microcapillary and the avoidance response was
quantified after 2 h. The results in Figure 2 show that maximal avoidance response was obtained even with 0.3 min, the shortest period examined. A stimulation period of 1 min was used in the subsequent experiments.
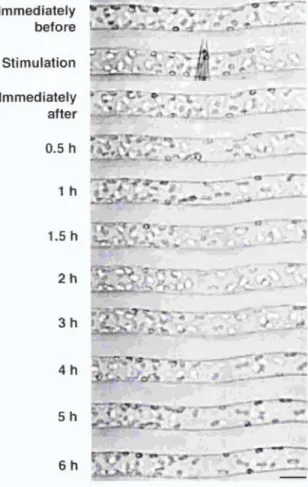
Protonemal cells were mechanically stimulated for 1 min and chloroplast distribution was continuously observed under the microscope with IR. Time course images of a cell during the chloroplast avoidance response are shown in Figure 3. It was found that a clear zone became apparent after 0.5 h and the zone spread in both the apical and basal direction until 2 h after mechanical stimulation. The time course for the avoidance response, shown in Figure 4, revealed that the response reached the maximum level at around 1-2 h and remained at a relatively constant level thereafter.
Effects of cytoskeletal inhibitors
I examined the effects of cytoskeletal inhibitors on the avoidance response of chloroplasts to determine the motile system responsible. After 2 h preincubation in an inhibitor solution, cells were mechanically stimulated and the
50
40 a) u)
0 30
Q a@ a
~ 20
0 C
~1
00 Q 0
-10
0.3 1 3
Stimulation period (min)
10
Figure 2. Relationship between stimulation periods and the chioroplast avoidance response. Protonemata were pushed for various periods from above with a microcapillary. The avoidance response was assessed 2 h after the stimulation. See materials and methods for quantification of the response.
Each point represents a mean ± SE obtained from 5-10 protonemata.
-16-
Immediately before Stimulation Immediately after
0.5 h
1h
1.5 h
2h
3h
4h
5h
6h
r -7-
47titiF.,~.
~ .t
#'4`6r~y-L
mtsikir.. II
~a, J,.4. .
23-4".°17`,4-lig"r li'.: a
Mtriti-a--47- ., 16" -0 -., .,
~.
~,L
1
ooV •V I • gi•
Figure 3. Time course images of a cell during the chloroplast avoidance response. Part of a cell was stimulated with a microcapillary for 1 min. The cell was continuously observed under IR through the IR-sensitive video system.
The protonemal tip was on the right. Bar, 20 pm.
40 30 Q)
c 20
0 a
a~10
0 c (cs 70 0 O Q
-10
0 1 2 3 4 5
Time after stimulation (h) 6
Figure 4. Time course of the chioroplast avoidance response induced by 1 min mechanical stimulation. Protonemata were pushed for 1 min with a microcapillary from above (0). Control results without mechanical stimulation are also presented (a). Each point represents a mean ± SE obtained from 4-5
protonemata.
-18-
inhibitor. The cytochalasin B concentration used is known to disrupt the MF architecture in A. capillus-veneris protonemata within 2 h (Kadota and Wada, 1995) and the concentrations of APM and colchicine are also sufficient to disturb the MT organization within 2 h (Murata and Wada, 1989). As shown in Figure 5, avoidance responses were inhibited by 0.1 mM cytochalasin B and 25 mM BDM but not by 3.3 µM APM or 5 mM colchicine. These results suggest that the response of chloroplasts after mechanical stimulation was mediated by the actomyosin motile system.
Behavior of each chioroplast during avoidance response
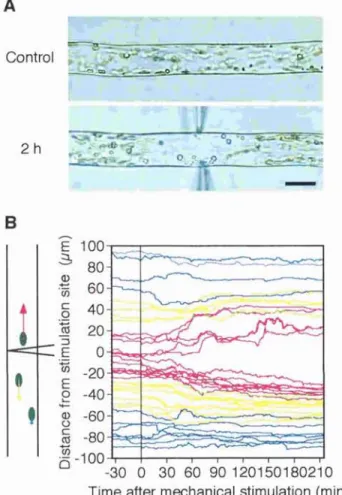
Chloroplasts were tracked individually to analyze the movement of chloroplasts during the avoidance response. The paths of movement of chloroplasts in a single cell for 1 h after stimulation are shown in Figure 6. Chloroplasts in an unstimulated cell moved randomly at a relatively constant velocity. On the other hand, in the stimulated cell, it is clear that the movement of chloroplasts near the stimulated site was activated in an axial direction, while that of chloroplasts far from the site remained at the unstimulated level. Time courses for chloroplast mobility in the regions around the stimulus point were
quantitatively examined over intervals of 1 h as shown in Figure 7. The mobility of chloroplasts in the stimulus region was greatest during the first 1 h after the stimulation. Then it gradually slowed down to the control level during 1-3 h
MS solution alone Colchicine APM Cytochalasin B
BDM DMSO
0 10 20 30 40
Avoidance response (%)
50
Figure 5. Effects of cytoskeletal inhibitors on the chioroplast avoidance response. Cells were treated with 5 mM colchicine, 3.3 pM APM, 0.1 mM cytochalasin B, or 25 mM BDM in MS solution containing 0.2% Pluronic F-127.
APM, cytochalasin B and BDM were dissolved with 0.5% DMSO and the control result with 0.5% DMSO alone is also presented. Each bar represents a mean ± SE obtained from 11-17 protonemata.
-20-
A
B
i --
Figure 6. Tracking of chioroplast movement during the avoidance response. A, Start and stop positions of chioroplast tracks made over 1 h after a 1 min mechanical stimulation are linked by arrows. B, Movement of chioroplasts in the control cell without the stimulation are also presented. The diagram of needle tip
indicates the site of the stimulation. The protonemal tip is on the right. Bar, 20 pm.
A
(a)
50 pm
(b) (c)
B
20
(a) 15-
E U10+
C co
51
0' 0-1 1-2 2-3 3-4 4-5 Time after stimulation (h)
i
E
co U) 0
20
15
10
5
0'
(b)
0-1 1-2 2-3 3-4 4-5 5-6 11
Time after stimulation (h)
20
(c) E 15
w V
c 10 0
5
0 ii
0-1 1-2 2-3 3-4 4-5 5-6 Time after stimulation (h)
-22-
Figure 7. Time course of chloroplast mobility after mechanical stimulation. A, Chloroplasts are divided into three groups with respect to their locations before
mechanical stimulation, namely those located in the basal region (a), in the stimulus region (b), and in the apical region (c). Each region is 50 pm in length.
B, Distances over which chloroplasts moved were measured in these three regions every 1 h during a period of 0-6 h after stimulation. Number of chloroplast analyzed are 14 for (a), 23 for (b), 17 for (c), and 63 for the control.
Data were obtained from 5 protonemata. Control data from unstimulated cells are presented as a line with a SE in each graph.
chloroplasts also increased transiently during the first 1 h, although to a lesser extent than seen in the area of stimulation. Figure 8 shows the direction of chloroplast movement at hourly intervals after mechanical stimulation. In mechanically stimulated cells, chloroplasts in the stimulus region (Fig. 8b) tended to move toward the base of the cell. Chloroplasts in regions both apical (Fig. 8c) and basal (Fig. 8a) to the stimulated region moved away from the stimulus for 2 h after the stimulation. In the later periods examined (4-6 h after stimulation), however, chloroplasts exhibited a tendency of recovery movement back to the stimulated area (Fig. 8).
Nuclear movement after mechanical stimulation
A nucleus can readily be identified in a single cell under the microscope. During the above experiments, in which nuclei were located at various distances from the stimulation point, no apparent movement of the nucleus in response to mechanical stimulation was observed. Therefore, the behavior of nuclei after mechanical stimulation was further investigated. Mechanical stimulation was applied for 1 min above the nuclear region of the cell, in the same way as for chloroplast avoidance analysis. No apparent avoidance response of nuclei was seen within 6 h after mechanical stimulation, while the chloroplasts around the nuclei moved away from the stimulation site. We can clearly observe a nucleus in the clear zone (Fig. 9). No significant change in the velocity or the direction of nuclear movement was detected during these experiments (Fig. 10).
-24-
(a)
^0-1
c ....z_2°1 E 2-3 i 3-4
a) Co 4-5 a) •
5-6 ^
^
I---I I I 7.1
0 0 0 0 0
100 75 50 25 0 25 50 75 100 BasalApical
Direction (%)
(b)
0-1 C
1-2 74
E2-3
3-4 a) 15 4-5 m 5-6
4.3 0 0 0 0 4.3
100 75 50 25 0 25 50 75 100 BasalApical
Direction (%)
(C)
r 0-1 c 2 1-2
Co
E 2-3 L 3-4
a~ •-.
a 4-5 a)
5-6 100 75
Basal
8
50 25 0 25 50 75 100 Apical Direction (%)
0 0 0 0 0
(d)
1h I F7,---7'2
100 75 Basal
50 25 0 25 50 75 100 Apical Direction (%)
0
Figure 8. Direction of chloroplast movement after mechanical stimulation.
Direction of chloroplast movement in the basal region (a), in the stimulus region (b) and in the apical region (c) as shown in Figure 7A, was analyzed hourly after mechanical stimulation. Chloroplasts which moved toward the apex (N) and toward the base (u) are shown as percentages. Percentage of chioroplasts which showed no movement was also shown on the right outside of the column.
A control result is also shown in (d). Data were obtained from the same group of 5 protonemata as in Figure 7.
-26-
Figure 9. Fluorescence micrographs of nuclei and chloroplasts after mechanical stimulation at the nuclear region. A, A cell was stained with SYTO 11 2 h after mechanical stimulation in the nuclear region. B, A cell without the stimulation was also stained and presented as a control. Micrographs are shown as combined images of fluorescence of SYTO 11 (green) and of chlorophyll (red). An arrow indicates the site of the stimulation. Bar, 20 pm.
20
_ 15 E a)
c10 a1 76
0
5
0
0-1 1-2 2-3 3-4 4-5 5-6 Time after stimulation (h)
Figure 10. Time course of nuclear movement after mechanical stimulation. After stimulation in the nuclear regions of protonemata, distances over which nuclei moved were measured hourly during the period of 0-6 h after stimulation. Each bar represents a mean ± SE obtained from 7 protonemata. Control data from unstimulated cells is presented as a line with a SE (n = 7).
-28-
Discussion
Touch-induced chioroplast avoidance response in A. capillus-veneris protonemal cells
In this chapter, cellular responses to mechanical stimulation in fern protonemal cells were identified. I have clearly shown that the avoidance response of chloroplasts was induced by a stimulus applied from outside of the cell. It should be noted that no impalement with a capillary was necessary for the
induction of the response. These results demonstrate, for the first time, that chloroplasts migrate away from a mechanically-stimulated site. This
"chloroplast avoidance response" was induced to a maximum degree even with a 0.3 min stimulus, the shortest period used in this study (Fig. 4), showing that the protonemal cells are very sensitive to mechanical stresses. It would be useful to know the threshold period for induction of the response. However, it was difficult to apply the stimulus for time periods shorter than 0.3 min using the
present experimental system. It was also not possible to quantitate the strength of mechanical stimulation.
The motile system mediating chioroplast avoidance movement
I found that cytochalasin B and BDM were effective, and colchicine and APM were both ineffective, inhibitors of the mechanically induced avoidance response
streaming within 2 h at concentrations of 0.1 mM and 25 mM, respectively, the streaming recovered subsequently after replacement of the cells in inhibitor-free media (data not shown). BDM is an agent that has been demonstrated to inhibit actomyosin ATPase and actin-myosin interaction in vitro (Horiuti et al., 1988; Higuchi and Takemori, 1989; Osterman et al., 1993; McKillop et al., 1994) but not to affect the organization of actin (Cramer and Mitchison, 1995; Grabski et al., 1998). Thus it seems that the avoidance movement of chloroplasts depends on the actomyosin motile system, but not on the MT system. Photo- relocation movement of chloroplasts is also due to MF-based motile system in A.
capillus-veneris (Kadota and Wada, 1992). The velocity of chioroplast movement during the avoidance response was close to that observed during photo-relocation movement (Kagawa and Wada, 1996). Taken together, these data indicate that it is plausible that the same motile system is used both in the photo-relocation movement and the mechano-relocation movement of chloroplasts.
Chloroplast avoidance response is a directional movement in A. capillus- veneris protonemal cells
Recently, chloroplasts in cells of the diatom, Pleurosira laevis, were found to translocate and gather around the nucleus following contact stimulation with a needle (Makita and Shihira-Ishikawa, 1997). In this cell, chloroplasts are located all around the cell periphery before the stimulation but after the
-30-
stimulation, they move toward the nucleus, which resides in the center of the cell.
The direction of chloroplast movement was always from the cell periphery toward the nuclear region, independent of the site of stimulation. In A. capillus- veneris protonemal cells, in contrast, only the chloroplasts which were located near the stimulated site responded and moved away from the site. It is evident that the chloroplast response in a protonemal cell is a directional movement, avoiding the site of stimulation. Further, the time required for translocation of chloroplasts was shown to be different between P. laevis and A. capillus-veneris.
It took only 5-10 sec in P. laevis to complete the response, while 0.5-2 h was necessary in A. capillus-veneris. The period in A. capillus-veneris was close to that required for photo-relocation movement of chloroplasts in the same cell (Kadota and Wada, 1992) . These results suggest that an unknown signal, which is generated by mechanical stimulation, arises and diffuses from the stimulated site and that chloroplasts respond to this signal, resulting in the avoidance movement. Chloroplast avoidance seemed to be more rapid and extent when the cell was touched by the flank of a needle than by the tip of it (Fig.
1). It may reflect the dependence of the amount of the signal on the all surface area contacted.
Behaviors of other organelles after mechanical stimulation
Movement of nuclei toward a wound site is known as "traumatotactic nuclear
pressure focused on a small part of the cell causes nuclear migration to the site within 10-50 min after the onset of continuous stimulation. In A. capillus-
veneris protonemal cells, migration of nuclei under chioroplast photo-movement conditions was reported (Kagawa and Wada, 1993). On the other hand, no apparent change in the location or mobility of a nucleus was observed within 6 h after mechanical stimulation. This result indicates that nuclei show neither
"traumatotactic" migration toward the stimulated site
, nor avoidance movement from the site in A. capillus-veneris protonemal cells. Further, no apparent change in cytoplasmic streaming could be observed before or after mechanical stimulation, and I could observe the movement of small vesicles through the clear zone (data not shown). Thus it seems that chloroplasts respond to mechanical stimulation independently from other organelles in A. capillus- veneris protonemal cells.
Possible mechanisms and purpose of the response
The fact that only chioroplasts escape from the stimulated area implies that a specific motor system associated with chioroplasts is regulated by a gradient of some signal generated by mechanical stimulation. However, the molecular mechanism of mechanical perception, and transduction of the avoidance response of chioroplasts are not understood at present. Ion channels which are activated by mechanical stimulation have been suggested to be a component of mechano-transduction steps (Morris, 1990). Mechano-induced
-32-
transient increases in cytosolic Ca' concentration ([Ca2+];) have been detected at the tissue level in higher plants (Knight et al., 1991; Knight et al., 1992;
Trewavas and Knight, 1994; Legue et al., 1997). Using the present experimental system, we might be able to monitor [Ca2}]; before and after mechanical stimulation at the cellular level.