Introduction
Agrobacterium tumefaciens-mediated gene transformation (AMT) is widely used for gene transformation in plants. Comparing to PEG (poly ethylene glycol), electroporation or bombardment transformation techniques, AMT allows efficient insertion of stable, unarranged, and single-copy sequences into the plant genome (Barkat et al. 1997, Shou et al. 2004). After the report by Hiei et al. (1994), the AMT method is widely applied to most of the japonica rice varieties. Though successful AMT technology has been reported in standard rice cultivar Nipponbare (NP), efforts need to be made to optimize the transformation efficiency genotype by genotype basis.
In our laboratory, a rice variety Gimbozu (GM) has become a valuable material for its highly activated transposable element
mPing. This makes it possible to establish mPing tagging system
in rice. Furthermore, high copy number (>1000) of mPing insertion sites offers a unique molecular marker, named mPing SCAR (Sequence characterized amplified region) marker (Monden et al. 2009). mPing SCAR markers are useful to analyze the populations between two closely related jaoponica rice varieties. It is worth to mention that mPing copy number is around 50 in most of the japonica varieties. When mutant lines originated from GM are crossed with NP, almost 1000 mPing
SCAR markers are available for gene mapping. With the use of
mPing SCAR markers, we have already identified several mutant
genes induced in GM.
However, the very low efficiency in trasformants recovery of GM prevents the complementation test of the candidate genes. Lower regeneration ability of GM and its high susceptibility to
Agrobacterium are the limiting factors in getting large number
trasformants. We could observe the GUS staining several calli after co-cultivation which indicated successful transformation of T-DNA. Those calli, however, turned brown soon after co- cultivation and could not regenerate. Browning of callus after co-cultivation is believed to be caused by excess proliferation of
Agrobacterium (Ozawa 2009). In this study, we tried to find a
way to optimize AMT method for GM and established the effective procedure for screening and regeneration of transformant.
Materials and methods
Agrobacterium-mediated transformationMature seeds of rice varieties GM and NP were manually dehusked. They were sterilized with 70% ethanol for 30 sec and then with 50% sodium hypochlorite for 40 min, followed by 3 rinses in sterile distilled water. Seeds were washed again with distilled water for 10 min in shaking. After three rinses with sterilized water, they were placed on solid N6 media (Chu et al.
A highly effective Agrobacterium-mediated transformation method for
a rice variety Gimbozu
Siddika Ayesha, Yoshihiro Yoshitake, Takayuki Yokoo, Mustafa Kamal, Masayoshi Teraishi, Takuji Tsukiyama and Yutaka Okumoto
Graduate school of Agriculture, Kyoto University(Kitashirakawa, Sakyo, Kyoto 606 - 8502, Japan)
Summary: An efficient gene transformation is a crucial for complementation test to identify the agronomically important genes. However, significant varietal difference was observed in the efficiency of Agrobacterium-mediated transformation in rice. Gimbozu has become a valuable research material for its highly activated transposable element, mPing. With the use of mPing SCAR markers and mPing tagging system, we have successfully identified several mutant genes induced in Gimbozu. However, the very low transformation efficiency in Gimbozu prevents the complementation test and functional analysis of the candidate genes. Here, we report an effective Agrobacterium-mediated transformation method applicable to Gimbozu. It was very difficult to recover viable calli from Gimbozu after the co-cultivation with Agrobacterium to obtain the transformants, as it is very sensitive to the Agrobacterium infection. To overcome this difficulty, we used a very low density of Agrobacterium (OD600=0.001) for infection and a high amount of meropenem at first screening. It was also important to keep infected calli dry by blotting a callus on paper towel and air drying very carefully at the time of co-cultivation. As a result, we got a sufficient number of transgenic plants in a time (90 plants from 5 resistant calli). Transformation efficiency became 14.3% and the regeneration efficiency was more than 90%.
Key words: Agrobacterium, Gimbozu, meropenem, transformation, viable calli
Acccepted : April 19, 2012
1975) supplemented with 2, 4-D (2 mg/L), casamino acids (0.3 g/L), sucrose (30 g/L), and gelrite (4 g/L). The cultures were incubated at 30℃ using a 12 hr light and 12 hr dark cycle. After 3-4 weeks, desirably proliferated calli shaped globular and compact with clear yellow color were subcultured on the same fresh medium for three days.
Agrobacterium strain EHA105 harboring pMLH7133 (Fig. 1) was cultured on 523 medium containing Kanamycin (50 mg/L) and Hygromycin (50 mg/L) for two days at 25℃ in the dark. A little amount of bacteria (OD600=0.001) were collected with toothpick and suspended in AA medium (AA salts, vitamins, amino acids, sucrose 20 g/L and 2,4-D 1 mg/L) containing acetosyringone (15 mg/L) (Toriama and Hinata 1985). Then, calli were immerged for 2 min and the excess bacterial suspension was removed by blotting on sterile paper towel. The calli were transferred on to a single filter paper placed on solid AA or AAM medium supplimented with 2 mg/L 2, 4-D (Toki et
al. 2006) for three days in dark at 25℃. Then, calli were washed
five times with sterilized water and last two times with sterilized water containing with meropenem (20 mg/L) to remove
Agrobacterium. Calli were blotted on paper towel for
desiccation. After washing, co-cultivated calli were placed on KSP (Tsugawa and Suzuki 2000) screening media containing meropenem (25 mg/L) and hygromycin (50 mg/L) for 3-4 weeks at 30℃ (12 hr dark and 12 hr light). At every 12-15 days, calli were transferred to fresh KSP media. During screening, volume of antibiotics was gradually decreased (Table 2). For regeneration, calli were transferred to DKN (Daigen et al. 2000) regeneration media containing NH4NO3 instead of (NH4)2 SO2
supplemented with casamino acids (2 g/L), sucrose (30 g/L), gelrite (6 g/L), indol-3 acitic acid (IAA-2 × 10-4mg/L) and
buteric acid (BA-5 × 10-4 mg/L) cultured for 3-4 weeks. At 7-10
days later, when green spots and primordia had seen only good part of calli from first regeneration media were transferred to fresh DKN media without meropenem and hygromycin. Regenerated shoots were transferred to MS (Murashige and Skoog-manufactured powder 4.4 g/L) medium supplemented with sucrose (30 g/L), sorbitol (30 g/L), gelrite (4 g/L) and casamino acids (2 g/L) for rooting and then seedlings were grown in controlled growth chamber. Petridishes for bacteria culture and co-cultivation were sealed with parafilm (Bemis
Flexible Packaging) whereas for induction, screening and regeneration sealed with surgical tape (Micropore).
Transformation efficiency and regeneration ability
After subculture, 35 calli of GM and 36 calli of NP were used for inoculation. At the end of final screening procedure, five resistant calli were observed in both the varieties. The transformation efficiency (%) was calculated by dividing the number of Hyg-resistant calli with the number of calli initially inoculated. Regeneration ability (%) calculated by dividing the number of GUS inserted plant with the number of Hyg-resistent calli inoculated.
PCR analysis
Putative transgenic plants were screened by the polymerase chain reaction (PCR) for the presence of Hyg-genes. About 2 cm leaf tip was cut from each seedling, grown on MS media. They were frozen in liquid N2, and grind with multi-beads shocker.
Supernatant was taken after treated with TPS buffer. After treating with 1 : 1 phenol-chloroform, DNA was isolated. PCR analysis to detect Hyg-gene was performed using Ex-Taq polymerase (Takara) with the primer Hyg-F1 5’-TTTCTGATCG AAAAGTTCGACAGCGTCT-3’ and Hyg-R1 3’-GGCAGTT CGGTTTCAGGCAGGTCTT GCAA -5’.
Histochemical assay
GUS buffer was prepared just before the sampling. Sample was taken into two 15 ml tubes (control and transgenic) of 90% acetone and kept on ice. Leaves and shoots were washed with 100 mM NaPO4 buffer. Then they were put in the GUS staining
buffer solution and desiccated the air by desiccators for 15 minutes. Samples were incubated with gentle shake for 2 hr at 37℃. After appearing the blue spot on the leaves and shoots, GUS buffer was discarded. A series of ethanol 10%, 30%, 50% and 70% were used for remove the staining buffer for 30 minutes each at 37℃. Assayed tissues were observed under a microscope and then photographed.
Results and discussion
Browning of callus during callus induction might be the consequent of production of phenolic compounds and harmful nitrite ions (NO2-) in culture medium. We didn’t observe any
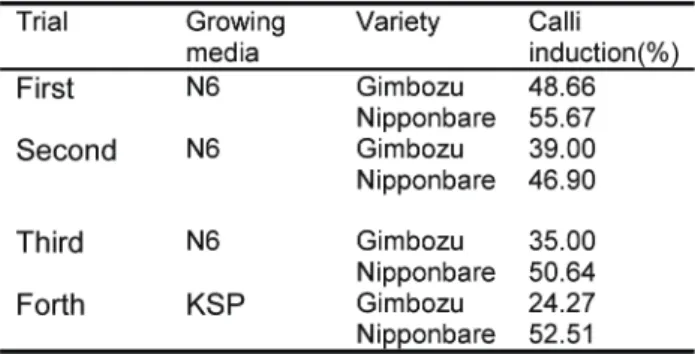
browning at callus induction stage. In most cases, the synthesis of secondary products is lost when the cells are differentiating and growing rapidly in culture (Ozeki and Komamine 1981). In comparison of N6 with KSP medium for calli induction, it was found that N6 medium exhibited better performance. Calli induction rate of GM was found to be slightly lower than that of NP in N6 medium and was much lower than that of NP in KSP medium (Table 1). It is reported that N6D medium for calli induction is applicable to both high regeneration and low
medium.
We selected calli with compact and round shape for subculture (Fig. 2a). Those calli became swollen on subculture medium in three days. They tended to show vigorous growth at the screening and regeneration stages.
GM is very sensitive to the Agrobacterium infection. It has been suggested that Agrobacterium induces browning and programmed cell death in non-host plant cells (Khanna et al. 2007). In this study, we successfully applied a very low density of Agrobacterium (OD600=0.001) for infection to GB, although Ozawa (2009) reported that the optimal concentration for
Agrobacterium infection in rice was OD600 of 0.04 and 0.2.
High bacteria concentration in GM might increase the number of transformation events, but increased bacteria inhibit viable cell growth.
In AMT method, suppression of overgrowth of the bacteria is a prerequisite to reduce their detrimental effects. Therefore, we used higher concentration of meropenem (25 mg/L) for 1st screening than that in previous work (12.5 mg/L). Then, the concentration was gradually decreased to 12.5 mg/L, 8.75 mg/ L, and 6.25 mg/L for 2nd and 3rd screening, and 1st regeneration media, respectively (Table2). In addition, application of meropenem-a β-lacam antibiotic-was reported to improve transformation efficiency (Ogawa and Mii 2007). Bacteria do not grow well on dried plates containing large amounts of a gelling agent (Ozawa 2009). We used media which were left for two days after preparation also followed the air drying treatment during inoculation and co-cultivation. After infection with Agrobacterium the calli were blotted on sterilized paper towel then air dry for 5-10 min. After three days of co- cultivation, the calli were washed and blotted on paper towel and kept for air drying 5-10 min. It was reported that desiccation
treatments greatly enhanced T-DNA delivery (Cheng et al. 2003). Nitrates are commonly used as nitrogen sources for plant culture. The reduction of nitrate to nitrite, then nitrite to ammonium is catalyzed by two enzymes NR (Nitrate reductase) and NiR (Nitrite reductase), respectively. Regeneration ability of GM is very low, because it has lower NiR activity and it cannot uptake NO2- from the medium. The accumulation of NO2- in
medium causes toxicity to the calli (Nishimura et al. 2005, Ogawa et al. 1999).The accumulation of NO2- may be explained
by either excess production of NO2- by NR or slowed reduction
of NO2- by NiR (Ogawa et al. 1999). As GM has lower NiR
activity, GM prefers a medium containing optimum nitrate source with lower NO2-. In this type media, GM cannot produce
excess amount of NO2- catalyzing by NR due to lower amount
of nitrate source. KSP and DKN medium were suitable for GM than NP calli (data not shown) for the following stages of co-cultivation as they reduce NO2- toxicity to the calli by
balancing NR and NiR activity. Moreover, a frequent change of media (three times during screening and two times in regeneration) was preferable for the reduction NO2-
accumu-lation.
Selection system is very important for identifying transformed calli. We used 3 times selection which required 48-50 days and maintained an effective amount of antibiotics up to first regeneration step (Table 2). At the end of the selection, all the non transformed calli turned very very black. We could observe vigorously growing calli with clear yellow color at the end of the 3rd selection. These calli were later proved to be from transformed cells and all the regenerated plants originated from those calli were transgenic plants (Fig. 2). Kumar et al. (2005) also reported that actively growing calli from 3rd round of selection found to be transformed.
We got only five resistant calli out of 35 inoculated calli. They incresed in size on first regeneration medium. Then, the calli generating green spots and small roots were transferred to 2nd regeneration medium using 2-3 plates from one calli. We didn’t add any meropenem or hygromycin in the 2nd regeneration medium. On the 2nd regeneration medium, we found many shoots which indicate high totipotent performance of calli. This indicated that a fresh medium without antibiotics could make GM calli highly totipotent. The transgenic plant was identified by PCR and Gus test (Fig. 2). Though transformation efficiency was not considerably high (14.29%) in GM, a large number of total transgenic plants (90 plants from 5 resistant calli) were Table 1. Performance of Gimbozu and Nipponbare on N6
and KSP calli induction medium
obtained from GM in compensation (Table 3). Each single callus can produced about 18 plants. According to the sectioning method by Hiei and Komari (2008), they successfully obtained 10-18 independent transgenic plants from a single callus in rice This indicated that the present protocol is optimal for AMT to GM. Obtaining viable calli after co-cultivation was the main obstacle for AMT applied to GM. We successfully produced viable calli after co-cultivation and a large number of regenerated plants. In the optimized method, the transformation efficiency of GM was not considerably high. Therefore, we need to get a larger number
of transformants from a single viable callus to compensate the lower transformation efficiency. To increase transformation efficiency, it is worth trying another combination of bacteria strain and vector type. It was reported that, Agrobacterium strain and vector type combination influence transformation efficiency of monocot species (Cheng et al. 2003). In addition, osmotic condition of inoculation and co-cultivation media could be optimized as it also might have important role in transformation efficiency.
References
Barkat, A., N. Carels and G. Bernardi(1997)The distribution of genes in the genome of Gramineae. Proc. Natl. Acad. Sci. USA 94:6857 - 6861.
Cheng, M., T. Hu, J. Layton, C. N. Liu and J. E. Fry(2003) Desiccation of plant tissues post-Agrobacterium infection enhances T-DNA delivery and increases stable transformation efficiency in wheat. In Vitro Cell. Dev. Biol. Plant 39:595 - 604.
Chu, C. C., C. C. Wang, C. S. Sun, C. Hsu, K. C. Yin, C. Y. Chu and F.Y. Bi(1975)Establishment of an efficient medium for anther culture of rice through comparative experiments of nitrogen sources. Sci. Sin.18:659 - 668.
Daigen, M., O. Kawakami and Y. Nagasawa(2000)Efficient anther culture method of the japonica rice cultivar Koshihikari. Breed. Sci. 50:197 - 202.
Hiei, Y., S. Ohta, T. Komari and T. Kumashiro(1994)Efficient transformation of rice (Oryza sativa L.) mediated by
Agrobacterium and sequence analysis of the boundaries of the
T-DNA. Plant J. 6:271 - 282.
Hiei, Y. and T. Komari(2008)Agrobacterium-mediated trans-formation of rice using immature embryos or calli induced from mature seed. Nat. Protocols 3:824 - 834.
Khanna, H. K., J. Y. Paul, R. M. Harding, M. B. Dickman and J. L. Dale(2007)Inhibition of Agrobacterium-induced cell death by antiapoptotic gene expression leads to very high trans-formation efficiency of banana. Mol. Plant. Microbe Interact. 2:1048 - 1054.
Kumar, K. K., S. Maruthasalam, M. Loganathan, D. Sudhakar and P. Balasubramanian(2005)An improved Agrobacterium-mediated transformation protocol for recalcitrant elite Indica rice cultivars. Plant Mol. Biol. Rep. 23:67 - 73.
Monden, Y., K. Naito, Y. Okumoto, H. Saito, N. Oki, T. Tsukiyama, O. Ideta, T. Nakazaki, S. R. Wessler and T. Tanisaka.(2009)High potential of a transposon mPing as a marker system in japonica × japonica cross in rice. DNA Res.
16:131 - 140.
Nishimura, A., M. Ashikari, S. Lin, T. Takashi, E. R. Angeles, T. Yamamoto and M. Matsuoka(2005)Isolation of a rice regeneration quantitative trait loci gene and its application to transformation systems. Proc. Natl. Acad. Sci. USA 102: 11940 - 11944.
Nishimura, A., I. Aichi and M. Matsuoka(2006)A protocol for
Agrobacterium-mediated transformation in rice. Nat. Protocols
1:2796 - 2802.
Ogawa, T., H. Fukuoka, H. Yano and Y. Ohkawa(1999) Relationships between nitrite reductase activity and genotype-dependent callus growth in rice cell cultures. Plant Cell Rep. 18:576 - 581.
Ogawa, Y. and M. Mii(2007)Meropenem and moxalactam : Novel β-lactam antibiotics for efficient Agrobacterium-mediated transformation. Plant Sci. 172:564 - 572.
Ozawa, K.(2009)Establishment of a high efficiency
Agrobacterium- mediated transformation system of rice (Oryza sativa L.). Plant Sci. 176:522 - 527.
Ozeki, Y. and A. Komamine(1981)Induction of anthocyanin synthesis in relation to embryogenesis in a carrot suspension culture : Correlation of metabolic differentiation with morphological differentiation. Physiol. Plant 53:570 - 577. Shou, H., R.B. Frame, A. Steven, S.A. Whitham and K. Wang
(2004)Assessment of transgenic maize events produced by particle bombardment or Agrobacterium-mediated transfor-mation. Mol. Breed. 13:201 - 202.
Toki S, N. Hara, K. Ono, H Onodera, A. Tagiri, S. Oka and H, Tanaka(2006)Early infection of scutellum tissue with
Agrobacterium allows high-speed transformation of rice. Plant
J. 47:969 - 976.
Toriyama, K and K. Hinata(1985)Cell suspension and protoplast culture in rice. Plant Sci. 41: 179 - 183.
Tsugawa, H., and M. Suzuki(2000)A low-temperature method for maintaining plant regeneration activity in embryogenic callus of rice (Oryza sativa L.) Plant Cell Rep. 19:371 - 375.
イネ品種銀坊主に適した高効率アグロバクテリウム形質転換系の開発
Siddika Ayesha・吉竹良洋・横尾敬行・Mustafa Kamal・寺石政義・築山拓司・奥本 裕
京都大学大学院農学研究科(〒 606 - 8502 京都市左京区北白川追分町)要旨:農業上重要な遺伝子を同定するための相補性検定において,形質転換効率の改善は重要である.しかしながら, アグロバクテリウムを介したイネ形質転換効率において大きな品種間差異が存在する.銀坊主には,非常に活性化され た転移因子 mPing が内在しており,価値の高い研究材料である.我々は,mPing SCAR マーカーおよび mPing タギング システムを利用し,銀坊主に誘発された突然変異遺伝子を同定することに成功してきた.しかしながら,銀坊主の非常 に低い形質転換効率は,候補遺伝子の相補性検定および機能解析の妨げとなっている.本報では,銀坊主に適した効果 的なアグロバクテリウム形質変換法を報告する.銀坊主は,アグロバクテリウム感染に対する耐性が極めて弱く,アグ ロバクテリウムとの共存培養後に活性のあるカルスを再生することが困難であった.これを克服するために,感染には 非常に低濃度のアグロバクテリウムを,一次スクリーニングでは高濃度のメロペンを使用した.また,共存培養時に感 染したカルスを紙タオルで水分を拭き取って乾燥した状態を保つことも重要であった.その結果,一度に十分量の形質 転換体を獲得することが可能となった.形質転換効率は 14.3%であり,再生効率は 90%以上であった. キーワード:アグロバクテリウム,銀坊主,メロペン,形質転換,カルス形成 作物研究 57:33 - 38(2012) 連絡責任者:奥本 裕([email protected])