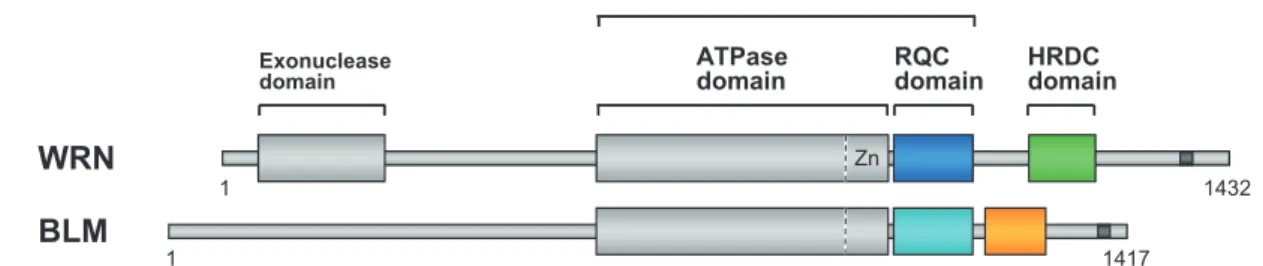
133 1.はじめに 1.1 RecQ ファミリーヘリカーゼ 細胞が正常に分裂するためには, 2 本の遺伝子 DNA が ねじれ合わさった二重らせん構造がいったんほどかれ, それぞれがコピーされる必要がある. このさいに DNA を ほどく(巻き戻す)という重要な作用をするのが, ヘリカ ーゼとよばれる一群のタンパク質酵素である. RecQ ファ ミリーヘリカーゼは, 大腸菌からヒトまで広く保存され ているヘリカーゼの一種で, ゲノム DNA の安定性維持に 必須な役割を果たしている. ヒトには 5 種類の RecQ ヘリカーゼが存在するが, その うち WRN(Werner syndrome protein)と BLM(Bloom syndrome protein)が変異によって機能を失うと, それぞれ 日本人に多い早老症のウェルナー症候群と, ガン多発のブ ルーム症候群が引き起こされる. いずれも常染色体劣性の まれな遺伝病で, 治療法の見つかっていない難病である. WRNと BLM は一般的な複製ヘリカーゼと異なり, 通 常の二重らせんに加えて, ホリデイジャンクションとよ ばれる十字型の組換え中間体や, 染色体末端を保護してい るテロメアなど, 特殊な構造をとった DNA を解きほぐす ことができる. この優れたヘリカーゼ活性こそ, WRN と BLMが細胞を, 早期老化とガン化から守ることができる 理由と考えられているが, なぜ可能なのか, 仕組みはわか っていなかった. 筆者らは, WRN と BLM の C 末側に保 存された 2 つのドメインに, 活性の秘密が隠されているの ではないかと考えて, 構造解析の研究を進めてきた.1)-4) 本稿では, 一連の研究から明らかとなった, RecQ ファミ リーヘリカーゼの作用の仕組みについて解説する. 1.2 WRN と BLM のドメイン WRNと BLM は, いずれも約 1,400 アミノ酸からなる大 きなマルチドメインタンパク質である(図1). モーター として働く ATPase ドメインに続いて, 特徴的な 2 つの領 域, RQC ドメイン(RecQ C-terminal)と HRDC ドメイン (helicase-and-ribonuclease D-C-terminal)を有している. RQCドメインは名前のとおり, RecQ ファミリーだけが 有するドメインで, ほかのタンパク質には見つかってい ない.
The RecQ family of DNA helicases play a key role in protecting the genome against deleterious changes. In humans, mutations in the members WRN(Werner syndrome protein) and BLM(Bloom syndrome protein)respectively lead to rare genetic diseases associated with accelerated aging and cancer predisposition. Recently we determined the 3D structures of
human WRN and BLM including a crystal structure of the RecQ C-terminal(RQC)domain
bound to a DNA duplex, the first structure of the RecQ-DNA complex. In the complex, the
β-wing of the RQC winged-helix motif acts as a scalpel to induce unpairing of a Watson-Crick base pair, an explanation for the unique activities of RecQs toward recombination and repair intermediates such as Holliday junctions.
図1 ヒト WRN と BLM のドメイン構成.(Domain diagram of human WRN and BLM.)本研究では両タンパク質 の, RQC ドメインと HRDC ドメインの立体構造を決定した. ATPase ドメインに付加された亜鉛(Zn)結合モチ ーフは, RQC ドメインとの連結部として働く. C 末端の暗く塗りつぶした領域は, 核移行シグナル.
図2に, 筆者らが構造決定した, WRN1),2) と BLM3),4) の ドメイン立体構造(計 4 個)を示した. 両タンパク質間で 領域の相同性は 10 ∼ 20 %と低いものの, RQC ドメイン (A, C)と HRDC ドメイン(B, D)は, それぞれ似たフォ ールディング構造をとっていた. ただし各ドメインの境界 (N 末端と C 末端の長さ)には WRN と BLM で違いがみら れ, これが結晶化のさい, 精製コンストラクトの検討に時 間を費やした理由であった. 現在はいずれのドメインも大 腸菌を使って大量発現させて, 高純度精製できるようにな っている. RQCドメインは, DNA 結合モジュールの 1 つとして有 名な winged-helix 構造5) をとっていた. 一方で HRDC ドメ インは, 6 本のヘリックスが組み合わさった楕円体状の構 造を形成していた. それぞれのドメインに対して合成 DNAとの共結晶化を試みてきたが, 複合体の結晶が得ら れたのは WRN RQCドメインであった(図2A). まず, この RecQファミリー初の DNA 複合体構造について解説する. 2.RQC ドメインの構造と機能 2.1 WRN RQC ドメイン− DNA 複合体の結晶解析 共結晶化の実験では, ヘリカーゼの一般的な基質である フォーク DNA を使ってスクリーニングを始めたが, 結晶 が得られたのは意外にも, 単純な二本鎖 DNA を用いたと きであった. DNA の長さを変化させつつ結晶化条件の精 密化を行ったところ, 14 塩基対の二本鎖 DNA を用いた ときに, 高分解能(1.9 Å)の反射を与える結晶を得るこ とができた. X 線回折データの収集は, 大型放射光施設ス プリング 8 のビームライン BL41XU, 44XU で行った. 構 造解析の位相決定は, 結晶に水銀試薬(ethyl mercuric phosphate)をソーキング法で浸透させて, 多波長異常分散 法(MAD)で行った. 決定した複合体の結晶構造(図2A)1) では, まず驚いた 北野 健 134
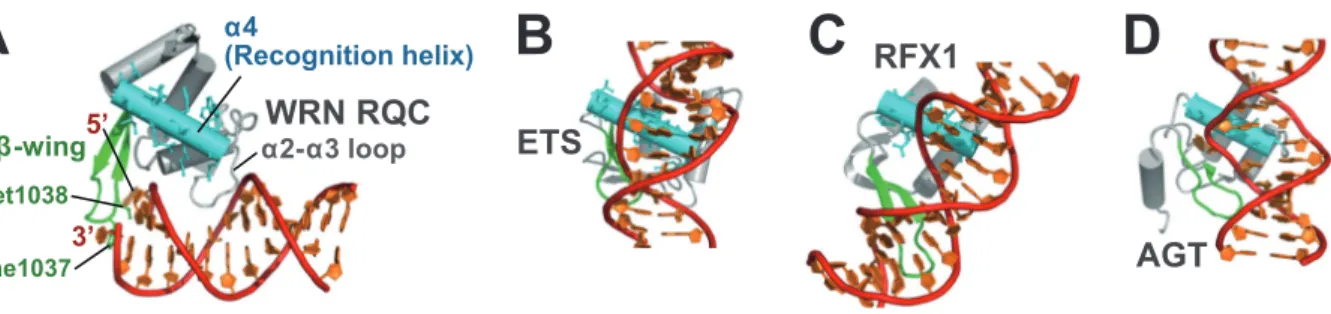
図2 ヒト WRN と BLM のドメイン立体構造.(3D structures of human WRN and BLM domains.)(A)WRN RQC ドメイン−二本鎖 DNA 複合体(PDB ID:3AAF).1)(B)WRN HRDC ドメイン. 2 つの異なる結晶系(2E1E, 2E1F)2) で構造を決定した.(C)BLM RQC ドメイン. リン酸およびヒ酸イオン結合型(3WE2, 3WE3)3) の構造 を決定した. イオンの位置は DNA 結合サイトの 1 つを表している.(D)BLM HRDC ドメイン. 唯一, NMR 法 で構造を決定した(2RRD).4)
ことに, 2 分子の WRN RQC ドメインが二重らせんの両末 端に結合して, ワトソン・クリック塩基対を 1 塩基ずつほ どいているのが確認できた. つまりほかのドメインがすべ て切り落としてあるにもかかわらず, WRN による DNA 巻き戻しの反応が, 結晶内で再現されていたわけである. これほど小さなドメインが, 単独で, ATP 加水分解のエ ネルギー供給もなしに, 一塩基とはいえ DNA をほどくの は予想外の結果であった. 2.2 RQC ドメインによる DNA 巻き戻しの仕組み 図3に, DNA 巻き戻し部位の拡大図を示した. 立体構造 の仕組みを詳しく調べると, RQC ドメインの端から突き 出したヘアピン構造(winged-helix ドメインでβ-wingとよ ばれる二次構造)が,“DNA 鎖分離ヘアピン”として, マ イナーグルーブの隙間から二重らせんに割り込むように 結合することで, 塩基対の引きはがしを巧妙に実現してい ることがわかった. なかでもβ-wingの先端にある Phe1037 と Met1038 の疎水性側鎖が, 新たに露出した 2 番目の塩基 対 G13’−C2にそれぞれ疎水性スタッキングすることで, 巻 き戻し構造の安定化に働いていた. また同じくβ-wing上 の Tyr1034 が, 巻き戻しによってフリップアウトした 5’-側の塩基 A1に疎水性スタッキングすることで, 3’-側の塩 基 T14’(ディスオーダーしていた)と再アニーリングでき ない位置に追いやっていた. β-wingがこうして DNA の末端と疎水性相互作用を形 成している一方で, 二重らせんのリン酸骨格への結合は, α2-α3ループという部分によって担われていた. 特にα 2-α3ループ上の Arg993 が, β-wingと反対側のメジャーグ ルーブから側鎖を伸ばして, 巻き戻し点である A1と C2 のあいだのリン酸基と強いイオン性水素結合を形成して いた. 実際, Arg993 をアラニンに変異させると, WRN RQCドメインの DNA 結合能は 1/20 にまで落ちた.1) 本結晶構造で興味深いのは, WRN RQC ドメインのβ -wingが, まるで指先にもった外科用メスのような格好で, ドメインの端から細長く突き出している点である. BLM RQCドメインの立体構造(図2C)にも, 同じく伸びたβ -wingが存在する. 後述するように WRN と BLM は, 通常 のヘリカーゼでは近づくことのできない DNA のわずかな 隙間からでも巻き戻しを開始できるが, その活性の秘密 は, このβ-wingの尖ったかたちにあったと考えられる. RecQファミリーヘリカーゼは, β-wingを DNA の間に差 し込んだまま, 二重らせんに沿って回転していくことで, 二本鎖を一本鎖にほどくと考えられる. 2.3 RQC ドメインは亜種 winged-helix 図4に WRN RQCドメイン(A)と, 一般的な winged-helix ドメイン(B-D)の DNA 結合様式の違いを示した. Winged-helixドメインは, 転写因子などのタンパク質に見られる DNA結合モチーフの 1 つで, helix-turn-helix スーパーファ ミリーに分類される.5) 認識ヘリックス(recognition helix) とよばれる 1 本のαヘリックスを二重らせんの溝にはめ 込んで, 決まった配列の塩基対とのみ水素結合を形成する ことで, 高い配列特異性を実現している. このときβ-wing も, DNA のほかの部位との相互作用に用いられる. 一方で RQC ドメインの場合は, 同じ winged-helix 構造 をとっているにもかかわらず, 認識ヘリックスが DNA か ら 5 Å 以上も離れた場所に位置していて, 結合に関与して いないことが明らかとなった. 一般にヘリカーゼは DNA を, 塩基配列とは無関係にほ どかなければならない. にもかかわらず RecQ ファミリー 135 図3 結晶中に捉えられたヘリカーゼ反応.1)(DNA-unwinding event captured in the crystal structure.)電子密度マッ
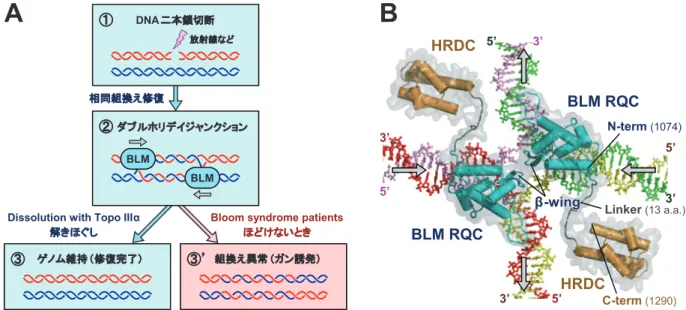
プ(composite omit map)をタンパク質は青色, DNA は赤色で示した. 末端のワトソン・クリック塩基対 A1− T14’がβ-wingによって巻き戻されている.
の活性部位に, 配列特異的な DNA 結合モチーフである winged-helix構造が含まれているのは, 研究者らを悩ませ る矛盾であった. 今回, 複合体の構造が得られたことで, 初めてその謎が解明された. RQC ドメインは winged-helix 構造をとっているものの, 認識ヘリックスを除外した新し い作用面を用いることで, 塩基の部分とは水素結合を形成 することなく, 配列無関係な DNA 結合を実現していたの である. また RQC ドメインの結合様式を見ると, 突き出したβ -wingが立体障害を起こすため, 二重らせんにはその末端 部分への結合のみ可能であることがわかる. 実際, 全長の WRNと BLM は DNA にランダムに結合することはなく, 二本鎖と一本鎖の分岐点, すなわち巻き戻しの開始点を正 しく認識して結合することが知られている. この高い構造 特異性は, 後述する解きほぐしのサイトを認識するのに重 要である. このように RecQ ファミリーヘリカーゼの機能に必要 とされる, 分岐点構造の認識(recognition), 結合(binding), そして巻き戻し(unwinding)の仕組みは, 実はすべて小さ な RQC ドメインに詰まっていたのである. 3.HRDC ドメインの構造と機能 RecQファミリーの RQC ドメインがヘリカーゼ反応に 直接関与しているのと対照的に, C 末側に続く HRDC ド メインの機能は, まだよくわかっていない. これまでは同ドメインも, DNA 結合モチーフの 1 つと 考えられてきた. しかし筆者らが構造決定した WRN HRDCドメイン2) と BLM HRDC ドメイン4) の立体構造 (図2B, D)を見比べてみると, DNA リン酸基と相互作用 できるアミノ酸(ドメイン表面に露出した塩基性側鎖)が, まったく保存されていないことが判明した. BLM HRDC ドメインでは, 逆に多くの酸性アミノ酸が表面に露出して おり(ドメインの等電点も 5.1 と低い), DNAとむしろ反 発し合うのではと考えられた. これらの知見を裏付けるよ うに, 精製した WRN HRDC ドメインと BLM HRDC ド メインは, いずれも溶液中で DNA に結合しなかった.2),4) 4.特殊な DNA 構造の解きほぐし 4.1 BLM とダブルホリデイジャンクション それでは立体構造を基にして, WRN と BLM がヒトの 健康を守る仕組みについて考えてみよう. 私達の細胞のゲノムは, 放射線や活性酸素が引き起こす さまざまな DNA 損傷を受けている. なかでも複製時に生 じやすい DNA 二本鎖切断(図5Aの①)は, 二重らせんが 途中で切れてしまうため, 細胞にとって最も重篤なゲノム 損傷と言われている. この二本鎖切断は, 通常はもう片方 の姉妹染色体を鋳型に使うことで, 相同組換え修復によっ てつなぎ合わせられる. しかし厄介なのが, このとき必然 的に生じる“ダブルホリデイジャンクション”(②)という, 二重らせんの絡まりである. この組換え中間体は名前のと おり, 十字型のホリデイジャンクションが 2 つ並んだ構造 をとっていて, ほどかずに切れてしまうと(③’), 組換え 異常の蓄積, ひいては細胞のガン化を誘発することが知ら れている. 私達の細胞で, このダブルホリデイジャンクションの 解きほぐし(dissolution;②→③の反応)を担っているの が, BLM と, トポイソメラーゼ IIIαのペアである.6) この 解きほぐし反応では, BLM がまず 2 つのホリデイジャン クションに結合して, 互いの分岐点を近付ける向きに素 早く移動させていく. 2 つが融合するまで近付いたら, トポイソメラーゼ IIIαが BLM と協調的に働いて, 脱カテ ナン化(decatenation)を行うことで 2 本の二重らせんを分 離させる(③). 図5Bに, BLM とホリデイジャンクションの複合体モ デル3) を示した. これはダブルホリデイジャンクションの 片方に相当するが, それぞれの分岐点移動は 2 回軸で関係 づけられる対称な反応であるため, 1 つのホリデイジャン クションに対して 2 分子の BLM が必要と考えられる. 本 北野 健 136
図4 RQCドメインの DNA 結合様式はこれまでの winged-helix とまったく違う.1)(DNA-interaction mode of RQC domain is distinct from all known examples of the winged-helix proteins.)(A)WRN RQC−DNA 複合体. 認 識ヘリックスに相当する部分を水色, β-wingを緑色で示した.(B)転写因子 ETS の winged-helix ドメイン(PDB ID:1PUE).(C)転写因子 RFX1 の winged-helix(1DP7).(D)修復タンパク質 AGT の winged-helix(1T38). 編集部注:カラーの図はオンライン版を参照下さい.
モデルでは, 2 つの BLM RQC ドメインが左と右の DNA に結合して, それぞれ外側に向けて二重らせんを巻き戻そ うとしている. ここで興味深いのは, RQC ドメインのβ-wingが, ドメ インの端から真下に向かって伸びているため, 2 分子の BLMは互いに立体障害を起こすことなく, それぞれのβ -wingを分岐点の小さな穴へ同時挿入することが可能な点 である. β-wingによってほどかれた左右の二本鎖は, ATPaseドメイン(図6A右に模式的に示した)の牽引力 によって一本鎖ずつ上下に移動して, 新しいペアと二本鎖 を形成するのであろう. このように BLM は, おそらくホ リデイジャンクションの穴を小さく保ったまま, 巻き戻し と再アニーリングを同時進行させることで, 効率的な分岐 点移動を実現しているのではと考えられる. ところで溶液中のホリデイジャンクションは, 通常はこ のように開いた十字構造ではなく, 逆平行に積み重なった コンパクトな状態(図6A左)にあることが知られてい る.7) この閉じた構造では, 分岐点は立体構造的に固定さ れているため, 移動させることができない. つまり BLM は解きほぐしのさい, 何らかの方法でホリデイジャンクシ ョンを開いた状態に保っているはずである. ここで RQC ドメインに続く HRDC ドメインの推定位 置を考えると(図5B), ちょうどホリデイジャンクション の十字の隙間, かつ ATPase ドメインの下(図6A右)に位 置することが予想される. BLM HRDC ドメインが負電荷 に富むことも考えると, この HRDC ドメインが, ホリデ イジャンクションを開くためのくさびとして使われてい るのではないかと推察される.3) 実際, HRDC ドメインを 137 図5 BLMとダブルホリデイジャンクション.(Dissolution of double Holliday junctions by BLM.)(A)BLM はトポ
イソメラーゼ IIIαと協調的に働いて, 修復中間体ダブルホリデイジャンクションを解きほぐす.6) ②では 2 つの ホリデイ構造は十字型でなく, 折れ曲がった状態で描かれている.(B)BLM −ホリデイジャンクションの複合 体モデル.3) 矢印は DNA 鎖の移動方向.
図6 RecQファミリーヘリカーゼによる分岐点移動のモデル図.1),3)(Proposed models for branch migration by RecQ-family helicases.)(A)BLM によるホリデイジャンクションのコンフォメーション開化と分岐点移動.(B)WRN によるテロメア T-ループの解きほぐし(仮想モデル). T-ループの点線枠内が右図に相当する.
欠損させると, BLM の解きほぐし活性はなくなることが 報告されている.6) BLMが変異したブルーム症候群では, ダブルホリデイ ジャンクションが解きほぐせないためヌクレアーゼで切 断するしかなく(③’), その結果, ガンを誘発する組換え異 常がゲノムに蓄積すると考えられる. 4.2 WRN とテロメア WRNと BLM は, 2 つの遺伝病の存在が示すとおり, そ れぞれの変異欠損を互いに補い合うことができない. すな わち細胞で異なる役割を担っていると考えられる. BLM がダブルホリデイジャンクションの解きほぐしに必須で あるのに対して, WRN の本来の基質がどのような DNA 構造なのか, はっきりとはわかっていない. しかし特に関 係性が指摘されているのは, 染色体末端のテロメアで ある. テロメアはその長さが細胞寿命に関係することから, “老化時計”ともよばれる重要なゲノム部位である. 一方 で, DNA 複製が特に困難な領域としても知られている. そ の理由の 1 つが, テロメアループ(T-ループ)とよばれる 投げ縄のような構造(図6B左)である. T-ループは, 損傷 に弱い DNA の最末端をループ内に隠して守る目的で形成 されているが, その特殊な構造ゆえ, 通常のヘリカーゼに はほどくことができない. この T-ループを, 複製時に解きほぐしているタンパク 質の候補として, WRN が有力視されている. 実際, WRN は試験管内で, T-ループの分岐点構造(図6B右;D-ルー プとよばれる)をほどくことができる.8) この分岐点をホ リデイジャンクションと見比べてみると, 実は構造的に似 ていて, たしかに RQC ドメインのβ-wingを差し込めば, 解きほぐしが可能ではと思われる. WRNが変異したウェルナー症候群では, 複製時の T-ル ープ解きほぐしがうまくいかずに, テロメア長の欠落が進 みやすく, 早老症の一因となっているのかもしれない. 5.おわりに WRNと BLM の構造解析が進んだことで, RecQ ファミ リー特有のヘリカーゼ反応の仕組みが明らかとなってき た. 特に, ユニークな形状のβ-wingを備えた RQC ドメイ ンが, 細胞の健康維持に重要な役割を果たしていることが わかってきた. ヒトでは, このように DNA 鎖の狭い隙間 に作用できるタンパク質が限られているため, WRN と BLMのいずれかに変異が起こると, 染色体の随所に不具 合が残ってしまい, その結果, 早老症やガンの病気が引き 起こされるのであろう. また今回は触れられなかったが, WRN と BLM は, グ アニン 4 重鎖(G-quadruplex;G4)とよばれる特異な DNA 構造も巻き戻すのに加えて, ほかの核内タンパク質と相互 作用ネットワークを形成することで, さらに広範な DNA 修復反応に関与することが明らかとなってきている.6) 引 き続き, DNA およびほかの結合タンパク質との複合体結 晶化が期待されている. 最後に, WRN と BLM は難病の原因タンパク質である 一方で, 最近では, 一般人に対する新しい抗ガン剤ターゲ ットとしても注目を集めている. ガン細胞では DNA 複製 が活発なため, WRN と BLM による解きほぐしの反応も, 通常の細胞より高頻度に要求される. 実際, 多くのガン細 胞では両タンパク質の高発現がみとめられる. したがって 片方だけでもヘリカーゼ活性を阻害できれば, ガンの増殖 抑制につながると期待されるわけである. 実際すでに, 欧 米のグループによって WRN と BLM の特異的阻害剤(低 分子化合物)が見出されており,9),10) 細胞レベルでのガン 抑制効果が報告されている. 今後, ドラッグデザインに 向けた阻害剤との複合体結晶化も, 重要な研究テーマと なろう. 文 献
1)K. Kitano, S. -Y. Kim and T. Hakoshima: Structure 18, 177 (2010).
2)K. Kitano, N. Yoshihara and T. Hakoshima: J. Biol. Chem. 282,
2717 (2007).
3)S. -Y. Kim, T. Hakoshima and K. Kitano: Sci. Rep. 3, 3294 (2013).
4)A. Sato, M. Mishima, A. Nagai, S. -Y. Kim, Y. Ito, T. Hakoshima,
J. -G. Jee and K. Kitano: J. Biochem. 148, 517 (2010). 5)G. M. Harami, M. Gyimesi and M. Kovacs: Trends Biochem.
Sci. 38, 364 (2013).
6)N. B. Larsen and I. D. Hickson: Adv. Exp. Med. Biol. 767, 161
(2013).
7)Y. Liu and S. C. West: Nat. Rev. Mol. Cell Biol. 5, 937 (2004).
8)P. L. Opresko, et al.: Mol. Cell 14, 763 (2004).
9)M. Aggarwal, J. A. Sommers, R. H. Shoemaker and R. M.
Brosh: Proc. Natl. Acad. Sci. U.S.A. 108, 1525 (2011). 10)G. H. Nguyen, et al.: Chem. Biol. 20, 55 (2013).
プロフィール
北野 健 Ken KITANO
奈良先端科学技術大学院大学バイオサイエンス 研究科
Graduate School of Biological Sciences, Nara Institute of Science and Technology
〒 630-0192 奈良県生駒市高山町 8916-5
8916-5, Takayama, Ikoma, Nara 630-0192, Japan e-mail: [email protected] 最終学歴:京都大学大学院理学研究科 博士課程 専門分野:タンパク質と DNA の結晶構造解析 現在の研究テーマ:ゲノム疾患の構造生物学 趣味:テニス, 釣り 北野 健 138