IgGレ
セプ ター を介 す る好塩 基 球 の
活性 化機構 に関 す る研 究
岡 山大 学 医 学部 第二 内科 学教 室(指 導:木 村郁 郎 教 授)
河
田
典
子
(平 成5年11月15日 受 稿)
Key words: KU812-F, Basophil, Ca2+ response, FcγRII, IgG antibody
緒 言 好 塩 基 球 ・肥 満 細 胞 系 に は, I型 ア レ ル ギー 反 応 に 関 与 す るIgEレ セ プ タ ー が 存 在 し, IgE 抗 体 を介 した レ セ プ タ ー のcross-linkingに よ り 化 学 伝 達 物 質 が 遊 離 さ れ て,種 々 の ア レ ル ギ ー 疾 患 を惹 起 す る こ と は よ く知 ら れ て い る.し か し な が ら,本 細 胞 系 はIgEに よ る ア レ ル ギ ー 反 応 の み な らず,成 人 の 非 ア トピ ー 型 喘 息 の 病 態 へ の 関 与 が 指 摘 さ れ て お り,広 くア レ ル ギ ー 反 応 全 般 に 関 係 し て い る こ とが 推 定 さ れ1), IgE系 以 外 の 機 序 に よ る ア レ ル ギ ー 反 応 に つ い て の 解 明 が 必 要 と考 え ら れ る. 教 室 の 木 村 ら2,3),小 林4)は,血 清IgE低 値 の 気 管 支 喘 息 患 者 の 好 塩 基 球 が, anti-IgGに 反 応 し て 形 態 変 化 を示 す こ と に 注 目 し,続 い て 木 村 ら5),松 岡6)は,免 疫 走 査 電 顕 法 を 用 い て 気 管 支 喘 息 患 者 の 好 塩 基 球 上 に 結 合 したIgG抗 体 を 半 定 量 す る こ とに よ りIgGレ セ プ ター の 動 態 を分 析 し,非 ア トピ ー 型 の 喘 息 患 者 で は 好 塩 基 球 が IgG抗 体 を 介 して ア レ ル ギ ー 反 応 に 関 与 し て い る可 能 性 を 報 告 し た. さ らに 最 近, IgGレ セ プ タ ー に は3つ の サ ブ タ イ プFcγRI(CD64), FcγRII(CD32w), FcγRIII(CD16)が 存 在 す る こ と が 明 らか に さ れ,各 々 に 対 す る モ ノ ク ロ ー ナ ル 抗 体 との 反 応 か ら,細 胞 表 面 上 のFcγ レ セ プ ター の 同 定 が 可 能 に な っ た こ と よ り7),教 室 の 高 橋 ら8)は,好 塩 基 球 上 のIgGレ セ プ タ ー をflow cytometry
(FCM)を 用 い て 分 析 し,好 塩 基 球 細 胞 表 面 上 のFcγRIIの 選 択 的 発 現 を 証 明 し た.加 え て, 抗IgG1,抗IgG4モ ノ ク ロ ー ナ ル 抗 体 を用 い て, 喘 息 患 者 の 好 塩 基 球 表 面 上 にIgG1, IgG4が 結 合 して い る こ と も明 ら か に し,気 管 支 喘 息 患 者 の 病 態 にIgG抗 体 が 何 らか の 役 割 を 果 た して い る 可 能 性 を示 唆 し た. こ の よ う に, IgGを 介 し た 好 塩 基 球 の 反 応 は 非 ア ト ピー 型 喘 息 の メ カ ニ ズ ム を 解 明 す る 上 で 重 要 と考 え られ る が,好 塩 基 球 は そ の 絶 対 数 が 極 め て 少 数 で あ る た め,実 験 に 際 し て は 種 々 の 制 限 を伴 わ ざ る を 得 な い.そ こ で 今 回 著 者 は, 末 梢 血 好 塩 基 球 の 代 わ りに,最 近 本 邦 で 樹 立 さ れ た ヒ ト好 塩 基 球 性 白 血 病 細 胞 株KU8129)を 用 い, IgGレ セ プ タ ー を介 し た 刺 激 に よ っ て,細 胞 内 情 報 伝 達 が どの よ う に 行 わ れ て い る か を, 細 胞 内Ca2+濃 度 の 変 化 を 指 標 と し て 分 析 し,併 せ て ヒ ス タ ミン 遊 離 に つ い て も検 討 し た の で 報 告 す る. 材 料 と 方 法 1.細 胞 実 験 に は,慢 性 骨 髄 性 白 血 病 患 者 末 梢 血 よ り 樹 立 さ れ た 細 胞 株KU812のsubcloneで あ る KU812-F(理 化 学 研 究 所 よ り供 与)を 使 用 し た. 10% fetal calf serum(FCS)を 含 むRPMI 1640(Gibco社)を 細 胞 培 養 液 と し て 用 い,細
胞 濃 度1-2×106/ml, 5%CO2, 37℃ で培 養 し, 3日 毎 に 培 養 液 を交 換 し た.
2.方 法 1) KU812-F細 胞 表 面 上 の 免 疫 グロ ブ リン レ セ プ タ ー 発 現 の 検 討 ①IgGレ セ プ タ ー 細 胞 をPBSで 洗 浄 後, 0.1% BSAを 含 む PBS(BSA-PBS)に1×106/ml濃 度 で 浮 遊 させ, モ ノ ク ロー ナ ル 抗 体 と反 応 させ る ため0.2mlず つ 5本 のtubeに 分 注 し た.ま ず1本 のtubeに 抗 FcγRIモ ノ ク ロ ー ナ ル 抗 体 で あ るFITC標 識 32.2(20μg/ml, Medarex社)を,さ ら に 別 の tubeに 抗FcγRIIIモ ノ ク ロ ー ナ ル 抗 体 で あ る FITC標 識Leu 11a(50μg/ml, Becton-Dickinson社)を 加 え, FcγRI, FcγRIIIの 発 現 を 検 討 し た.な お, FITC標 識mouse IgG1
(20μg/ml, Becton-Dickinson社)を 加 え たtube をcontrolと し て 用 い,こ れ ら のtubeを 各 々 氷 上 で30分 間 反 応 させ, PBSで2回 洗 浄 した 後 再 浮 遊 させ た.ま たFcγRIIに つ い て は,抗Fcγ RIIモ ノ ク ロー ナ ル 抗 体 で あ る2E1(20μg/ml, Immunotech社)並 び にcontrolと し てmouse IgG2a(20μgml, Becton-Dickinson社)を 氷 上
で30分 反 応 させ た 後PBSで2回 洗 浄 し,再 度 BSA-PBSに 浮 遊 さ せ,二 次 抗 体 と してFITC 標 識goat F(ab')2 anti-mose lgG(GAM, Tago 社)を40μg/mlの 濃 度 で 浮 加 して 氷 上 で30分 間 反 応 後,洗 浄 しPBS 0.5mlに 再 浮 遊 させ た.以 上 の 検 体 を, FACStar(Becton-Dickinson社)で 分 析 した. ②IgEレ セ プ タ ー RPMI1640(10%FCS加)に1×106/ml濃 度 で 浮 遊 させ た 細 胞 にhuman IgE(25μg/ml, Japan Immuno-monitoring Center)を 加 え て
37℃, 4時 間incubateし た 後 洗 浄 し, PBSに 再 浮 遊 させ, FITC標 識anti-IgE(10μg/ml, Tago社)を 添 加 し て 氷 上 で30分 間 反 応 させ て
か ら 洗 浄 し, FACStarで 分 析 し た.な お, human IgEを 加 え な い 細 胞 に, FITC標 識anti-IgEを
添 加 し て 反 応 さ せ た も の をcontrolと し て 用 い た. 2)細 胞 内Ca2+濃 度 の 測 定 fluo-3(Dojindo社)は,励 起 波 長500nm, 最 大 蛍 光 波 長520nmとFCMのargon laser (488nm)で の 検 出 に 適 して お り, acetoxymeth ylester化 の 形(fluo-3/AM)で 容 易 に 細 胞 内 に 取 り込 ま れ, Ca2+と 結 合 す る と 蛍 光 強 度 が40 倍 に 増 強 さ れ,し か もCa2+と の 親 和 性 が 低 い こ と よ り,細 胞 内Ca2+濃 度 の 変 化 を 捉 え や す い た め10,11), calcium indicatorと し て 使 用 し た. 細 胞(2×106/ml)をHBSSに 浮 遊 させ, fluo -3/AM 2μMを 加 え,暗 所,室 温 で40分 間 反 応 させ て2回 洗 浄 し, HBSS(Ca2+, Mg2+を 含 む)に 再 浮 遊 させ37℃ で10分 間incubateし た 後.各 種 刺 激 を加 え て30分 以 内 にFACStarで 分 析 し た. IgGレ セ プ ター を介 す る刺 激 と し て Aggregated human IgG(Agg-IgG)を 用 い た が,こ れ は 実 験 の 度 にchromatographically purified human IgG(Zymed社)を63℃, 60
分 間 熱 処 理 を加 え た 後10000×g, 10分 間 遠 沈 し large aggregatesを 除 去 し て 作 成 し た もの を 使 用 し た.な を, fluo-3の 蛍 光 測 定 は525nm(FL -1)band pass filterを 通 し てlinear scaleで 分 析 し,経 時 的 に5000個 の 細 胞 のmean fluores cenceを 求 め た. 細 胞 内Ca2+濃 度 の 較 正 の た め, Bromo-A 23187(10μM, Molecular Probes社)添 加 時 の 値 を 最 大 値F maxと し12), MnCl2を 添 加 し た と き の 値 よ り,最 小 値F minを 算 出 し た13). こ れ ら の 値 よ り,下 記 の 式 を 用 い て 細 胞 内Ca2+ 濃 度 を 求 め た14). [Ca2+]i(nM)=Kd(F-Fmin)/(Fmax-F) F:検 体 のmean fluorescence Fmin=Fmax-(Fmax-FMnC12)×1.25 Kd=400nM(fluo-3の 解 離 定 数) な お,較 正 は 実 験 検 体 ご と に 行 っ た. 4)ヒ ス タ ミ ン遊 離 率 の 検 討 HBSS(Ca2+, Mg2+を 含 む)に,細 胞 を1× 105/ml濃 度 で 浮 遊 さ せ, IgGレ セ プ タ ー を 介 す る 刺 激 並 び に 非 生 理 的 刺 激 物 質 と してCalcium ionophore(A23187, 1μg/ml)を 加 え て37℃, 45分 間incubateし た の ち, 4℃, 400×gで 10分 間 遠 心 し,上 清 中 の ヒ ス タ ミ ン をRIA (Immunotech社)で 測 定 し た15).ま た, human IgE(25μg/ml)を 加 え て4時 間incubateし た 細 胞 をHBSSに 浮 遊 させ, anti-IgEを 添 加 し 上 記 と 同 様 に し て 反 応 させ た.刺 激 物 質 の 代 わ り にHBSSを 加 え 同 様 にincubateし て 得 られ
た も の を 自然 遊 離 量 と し て 求 め,全 ヒ ス タ ミ ン 量 は 細 胞 浮 遊 液 を100℃, 10分 間 熱 処 理 した も の を遠 沈 後,上 清 を 測 定 に 供 し た. RIA法 に よ る 測 定 は す べ てduplicateで 行 い,ヒ ス タ ミン 遊 離 率 は,下 記 の 式 に よ り算 出 して 求 め た.な お, 自然 遊 離 量 は 全 ヒ ス タ ミン 量 の10%以 下 で あ っ た. ヒ ス タ ミ ン遊 離 率(%)=
刺 激 に よ る遊 離 量-自 然遊 離 量/
全 ヒス タ ミン量-自 然 遊 離 量
×100 次 に,ヒ ス タ ミ ン 遊 離 を 増 強 さ せ る 目 的 でrecom binant human interleukin-3(rhIL-3, Genzyme 社)を100U/mlの 濃 度 で 加 え て10分 間prein cubateし た 後16),刺 激 物 質 を 添 加 し て ヒ ス タ ミ ン 遊 離 を 検 討 し た . 結 果 1. KU812-Fの 免 疫 グ ロ プ リ ン レセ プ ター 発 現 に つ い て KU812-Fの 細 胞 膜 表 面 上 に は, FcγRI, FcγRIIIは ご く僅 か な 発 現 か,あ る い は ほ と ん ど 発 現 が 認 め ら れ な か っ た の に 対 し て, FcγRIIで はFig. 1Aに 示 す 如 く明 か な発 現 が み られ, IgGレ セ プ タ ー に 関 し てFcγRIIの 選 択 的 な 発 現 を 示 し た ヒ ト末 梢 血 好 塩 基 球 と 同 様 の 結 果 で あ っ た17,18). 一 方IgGレ セ プ タ ー(Fig . 1B)に つ い て は, FcγRIIの よ う な 明 瞭 な 発 現 で は な く,軽 微 な 発 現 で あ っ た.
FcγRI
FcγRII
FcγRIII
Log fluorescence
intensity
Log fluorescence
intensity
Fig.1 KU812-Fに おけ る免 疫 グ ロプ リン レ セプ ター 発 現 の検 討 A(上): IgGレ セ プ タ ー FcγRIIで は, control(…)に べ て 蛍 光 強 度 に 顕 著 な差 を認 め,同 レセ プ ター の 明 か な発 現 が み られ た. B(左): IgEレ セ プ ター
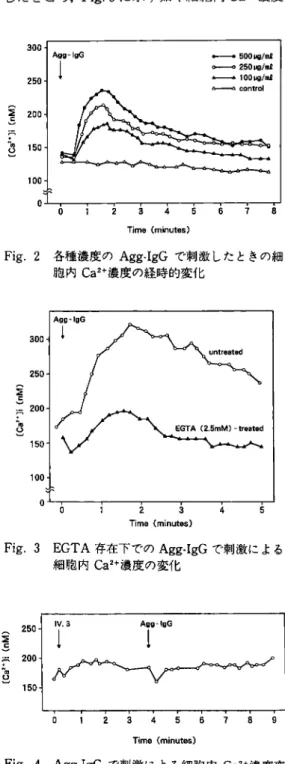
2. Agg-IgG刺 激 に よ る 細 胞 内Ca2+濃 度 の 動 態 分 析 1)時 間 的 経 過 まず,刺 激 前 の 細 胞 内Ca2+濃 度 を1×104個 の KU812-F細 胞 に つ い て 測 定 し,続 い て 種 々 の 濃 度(100, 250, 500μg/ml)のAgg-IgGで 刺 激 し て5-10秒 ご とに5000個 測 定 し た.ま たcon trolと し てHBSSを 添 加 し た 細 胞 に つ い て 同 様 の 検 定 を行 っ た.そ の 結 果Fig. 2に 示 す 如 く Agg-IgG刺 激 で はcontrolに 比 し添 加 後30-40 秒 で 細 胞 内Ca2+濃 度 の 上 昇 が み られ, 1分30秒 -2分 後 に ピー クに 達 し ,そ の 後 徐 々 に 低 下 し て10分 前 後 で 元 の 濃 度 に 復 し,し か もAgg-IgG の 容 量 依 存 性 に 増 強 さ れ る傾 向 が 認 め ら れ た. な お, Agg-IgG 500μg/mlで 刺 激 し た 場 合,ピ ー ク時 の 細 胞 内Ca2+濃 度 は 刺 激 前 の1.4±0.16倍 (mean±SD, n=13)に 達 し た. 2)細 胞 外 カ ル シ ウ ム の 影 響 細 胞 外 か ら のCa2+の 流 入 を 遮 断 す る 目的 で細 胞 浮 遊 液 に2.5mMのEthyleneglycol-bis-(2-aminoethyl)-tetraacetate(EGTA)を 加 え て 処 理 した 後, Agg-IgGで 刺 激 し, EGTAを 加 え て い な い 対 照 群 と比 較 し た.そ の 結 果, Fig. 3 の 如 く刺 激 後 に は 対 照 群 と同 様30-40秒 で 細 胞 内Ca2+濃 度 の 上 昇 が み ら れ た が,上 昇 の ピ ー ク は 低 く,し か も3分 以 内 に 元 の レベ ル に 戻 っ た. こ の 短 時 間 の 僅 か な 上 昇 は,細 胞 内 の 貯 蔵 カ ル シ ウ ム か ら の 放 出 に よ る も の と考 え ら れ た, 3)細 胞 内Ca2+濃 度 の 上 昇 に お け るFcγRII を 介 す る関 与 KU812-F細 胞 膜 表 面 上 にFcγRIIの 選 択 的 な 発 現 が 認 め ら れ た こ と よ り,か か る 受 容 体 を 介 す る 刺 激 で 細 胞 内Ca2+濃 度 の 上 昇 が 起 こ り う る か 否 か を, Agg-IgGを 用 い て 検 討 し た.ま ず, FcγRIIへ のhuman IgGの 結 合 を 遮 断 す る 抗
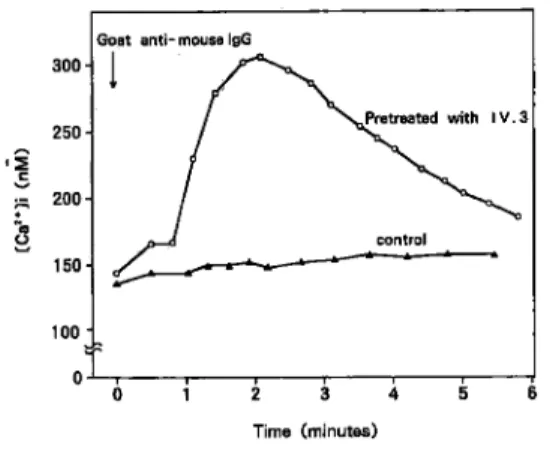
FcγRIIモ ノ クロー ナ ル 抗 体IV. 3(Medarex社) を 用 い て, Agg-IgG刺 激 とFcγRIIの 結 合 の 特 異 性 を み た.す な わ ちfluo-3/AMで 処 理 した 細 胞 浮 遊 液 にIV. 3 Fab(10μg/ml)を 添 加 し, 続 い てAgg-IgG(500μg/ml)で 刺 激 し た と こ ろ, Fig. 4に 示 す よ うに 細 胞 内Ca2+濃 度 の 上 昇 は 検 出 され な か っ た.従 っ て, IV. 3に よ っ てAgg-IgG の 刺 激 伝 達 は 完 全 に 遮 断 され る こ と が 判 明 した . 次 に, FcγRIIをcross-linkさ せ た 際 の 細 胞 内Ca2+濃 度 の 変 化 に つ い て 検 討 し た.ま ず,前 記 のIV. 3を 細 胞 浮 遊 液 に 加 え て4℃, 30分 間 incubateし て 洗 浄 処 理 し た 細 胞 にgoat anti mouse IgG(GAM, Tago社)25μg/mlを 添 加
し た と こ ろ, Fig. 5に 示 す 如 く細 胞 内Ca2+濃 度
Fig. 2 各 種 濃 度 のAgg-IgGで 刺 激 した と きの 細 胞 内Ca2+濃 度 の 経 時 的 変 化
Fig. 3 EGTA存 在 下 で のAgg-IgGで 刺 激 に よ る 細 胞 内Ca2+濃 度 の 変 化
Fig. 4 Agg-IgGで 刺 激 に よ る 細 胞 内Ca2+濃 度 変 化 に 及 ぼ す 抗FcγRII抗 体IV. 3の 影 響
Fig. 5 抗FcγRII抗 体IV. 3をGoat anti-mouse IgGでcross-linkさ せ た と きの 細 胞 内Ca2+
濃 度 の 変 化
Fig. 6 Human IgEでpreincubateし た 後, Anti-IgEで 刺 激 した と き の 細 胞 内Ca2+濃 度 の 変 化 は30-40秒 後 に 急 峻 な 上 昇 を示 し,ピ ー ク も 刺 激 前 の3.05±0.45倍(mean±SD, n=5)と 著 し い 変 化 が 認 め ら れ た. 3. IgE刺 激 に よ る 細 胞 内Ca2+濃 度 の 動 態 human IgE(25μg/ml)で4時 間preincubate し た 細 胞 にfluo-3/AMを 同 様 に し て 取 り込 ま せ た 後, anti-IgE(20μg/ml)を 添 加 して 細 胞 内 Ca2+濃 度 を 測 定 し たが, Fig. 6の 如 く同 刺 激 に よ る 細 胞 内Ca2+濃 度 に 変 化 は 認 め られ な か っ た. 4.ヒ ス タ ミ ン 遊 離 各 種 刺 激 に よ る 本 細 胞 の ヒ ス タ ミ ン 遊 離 に つ い て 検 討 し た と こ ろ, Table 1の 如 くA23187に よ る刺 激 で は28.6±4.7%と 有 意 な ヒ ス タ ミン 遊 離 が 見 られ た の に 対 して, Agg-IgG刺 激 で は100, 250, 500μg/mlの 各 種 濃 度 で 刺 激 し て も 有 意 な ヒ ス タ ミ ン遊 離 は 認 め ら れ な か っ た. 次 に, IV. 3(10μg/ml)を 加 え てincubateし 洗 浄 後,二 次 抗 体GAM(25μg/ml)を 添 加 し て FcγRIIをcross-linkさ せ た 際(IV. 3+GAM) の ヒ ス タ ミ ン遊 離 を 検 討 し た が,細 胞 内Ca2+濃 度 の 著 し い 上 昇 に も拘 ら ず,ヒ ス タ ミン 遊 離 は 10%以 下 で あ っ た. ま た, human IgEでpreincubateし た 細 胞 をanti-IgE(0.2-2μg/ml)で 刺 激 し て も 有 意 な ヒ ス タ ミ ン 遊 離 は み ら れ な か っ た. さ ら に, rhlL-3を 添 加 し てpreincubateし た 後 同 様 の 刺 激 を加 え て 検 討 し た と こ ろ, A23187 で は ヒ ス タ ミン遊 離 の 増 強 が み ら れ た が, IgGレ Table 1 各 種 刺 激 に よ るKU812-Fか らの ヒ ス タ ミン遊 離率 の 検 討
Mean±SD N. D.=Not Done (n=3) * Goat anti-mouse IgG
セ プ タ ー を 介 す る刺 激 で は ヒ ス タ ミン 遊 離 の 増 強 は 認 め ら れ な か っ た. 考 察 好 塩 基 球 はIgEを 介 す るI型 以 外 の ア レ ル ギ ー 反 応 に も関 与 す る こ とが 想 定 さ れ て い る こ と か ら,本 編 で はIgGを 介 し た 好 塩 基 球 の 活 性 化 機 構 を解 明 す る 目 的 で,ヒ ト好 塩 基 球 性 白 血 病 細 胞 株KU812-Fを 用 い て,細 胞 表 面 上 のIgG レ セ プ タ ー の 発 現,並 び に か か る レ セ プ タ ー を 介 し た細 胞 の 活 性 化 を細 胞 内Ca2+濃 度 の 動 態 分 析,及 び ヒ ス タ ミ ン 遊 離 能 の 観 点 か ら検 討 を加 え た.そ の 結 果, KU812-Fに は 末 梢 血 好 塩 球 と 同 様, FcγRIIの 選 択 的 発 現 が 認 め られ,さ ら に 同 レ セ プ ター を介 す る刺 激 に よ って,細 胞 内Ca2+ 濃 度 の 明 ら か な 上 昇 が み ら れ た が,ヒ ス タ ミン 遊 離 は 認 め ら れ ず,好 塩 基 球 に お い て もIgGレ セ プ タ ー が ヒ ス タ ミ ン 遊 離 以 外 の 細 胞 活 性 化 に 関 与 して い る こ とが 判 明 し た. 今 回 検 討 を加 え たKU812-Fは,慢 性 骨 髄 性 白血 病 患 者 の 末 梢 血 よ り樹 立 さ れ た 細 胞 株 で, 芽 球 の 形 態 を と り,一 部 に トル イ ジ ン ブ ル ー に 異 染 性 を 示 す 好 塩 基 球 顆 粒 を 持 ち,ヒ ス タ ミ ン の 含 有 も認 め ら れ て い る9).ま た,好 塩 基 球 と類 似 の 表 面 抗 原 を 有 し19,20),特 にIgEレ セ プ タ ー に つ い て は,高 親 和 性IgEレ セ プ ター で あ るFcε RIの αサ ブ ユ ニ ッ トが ク ロ ー ニ ン グ され て お り21),好塩 基 球 系 へ の 分 化 の 可 能 性 が 示 唆 され て い る,今 回flow cytometryを 用 い てIgGレ セ プ ター の サ ブ タ イ プ を 検 討 し た 結 果,ヒ ト末 梢 血 好 塩 基 球 と 同 様, FcγRIIの 選 択 的 発 現 が 認 め ら れ17,18), IgGを 介 す る 好 塩 基 球 の 刺 激 伝 達 を 検 討 す る 上 で 有 用 な モ デ ル と考 え ら れ た. FcγRIIは, 40kDaの 分 子 量 を も つ レ セ プ タ ー で,単 球,好 中 球,好 酸 球, Bリ ン パ 球 な ど 幅 広 く白 血 球 に 存 在 す る ほ か,血 小 板 上 に も発 現 が 認 め られ て い る が, IgGに 対 す る 親 和 性 が 低 い た めmonomeric IgGは ほ と ん ど結 合 せず, 免 疫 複 合 体 や 多 価 のIgGの み 結 合 す る と い う性 質 を も っ て い る22,23).今 回Agg-IgGを 用 い て IgGの 刺 激 伝 達 機 構 の 検 討 を行 っ た と こ ろ,刺 激 後 速 や か に 細 胞 内Ca2+濃 度 の 上 昇 が み られ た. さ ら に, EGTAを 用 い た 実 験 結 果 よ り,刺 激 直 後 の 細 胞 内Ca2+濃 度 の 上 昇 に は あ る程 度 細 胞 内 の 貯 蔵 カ ル シ ウ ム か ら の 放 出 が 関 与 し て い るが, よ り 高 度 の 濃 度 上 昇 並 び に 持 続 に 関 し て は 細 胞 外 か ら の カ ル シ ウ ム の 流 入 が 必 要 と考 え ら れ た. ま た,抗FcγRII抗 体 で あ るIV. 3に よ っ て Agg-IgGに よ る 細 胞 内Ca2+濃 度 の 上 昇 が 完 全 に ブ ロ ッ ク さ れ た こ と か ら,細 胞 内 シ グ ナ ル 伝 達 はFcγRIIを 介 し て 起 こ っ て い る こ と が 確 認 され た.さ ら に, IV. 3の 単 独 刺 激 で は 細 胞 内Ca2+ 濃 度 は 変 化 し な か っ た の に 対 し, IV. 3で 処 理 し た 細 胞 に 二 次 抗 体 を 加 え てFcγRIIをcross -linkさ せ る と,細 胞 内Ca2+濃 度 が 著 明 に 上 昇 し た こ と よ り, Agg-IgGはFcγRIIに 単 に 結 合 す る だ け で な く,レ セ プ ター のclusteringを 惹 起 し,そ れ に よ っ て,細 胞 の 活 性 化 が 誘 導 さ れ る も の と推 察 さ れ た.以 上 の 結 果 は,生 体 内 に お い て も 好 塩 基 球 が 免 疫 複 合 体 な ど に よ っ て 刺 激 を 受 け, IgGレ セ プ タ ー を 介 し て 細 胞 内Ca2+濃 度 の 上 昇 を初 め と す る細 胞 の 活 性 化 が 起 こ る 可 能 性 を 示 唆 し て い る も の と思 わ れ た. 従 来 よ りIgEレ セ プ ター を 介 した 細 胞 の 活 性 化 機 構 に つ い て は,好 塩 基 球 ・肥 満 細 胞 系 を 中 心 に 研 究 が 行 わ れ て お り, FcεRIのcross -linkingに よ っ て,イ ノ シ トー ル リ ン 脂 質 代 謝 の 亢 進,細 胞 内Ca2+濃 度 の 上 昇 が 起 こ り,ヒ ス タ ミン な ど種 々 の 化 学 伝 達 物 質 が 遊 離 さ れ る こ と が 明 らか に さ れ て い る が, IgGレ セ プ ター に 関 し て も,近 年,様 々 な 細 胞 に お け るFcγRサ ブ タ イ プ の 同 定,並 び に そ れ ら を 介 し た シ グ ナ ル 伝 達 機 構 の 解 明 が 行 わ れ て い る.
FcγRII, FcγRIIIの2種 類 のIgGレ セ プ タ ー を持 つ 好 中 球 で は,細 胞 活 性 化 に 際 して 両 レ セ プ ター 間 で の 協 同 作 用 が 報 告 さ れ て お り24),ま た 血 小 板 の よ う にFcγRIIの み の 発 現 が 認 め ら れ て い る 細 胞 で は, FcγRIIのcross-linkingに よ っ て,イ ノ シ トー ル リ ン脂 質 代 謝 の 亢 進,細 胞 内Ca2+濃 度 の 上 昇 が 起 こ り,最 終 的 に 血 小 板 凝 集 が 引 き 起 こ さ れ る こ と が 認 め られ て い る25). 今 回 のKU812-Fの 実 験 系 で も, FcγRIIの cross-linkingに よ り細 胞 内Ca2+濃 度 が 上 昇 し, 細 胞 内 シ グ ナ ル 伝 達 が 行 わ れ て い る こ とが 確 認 さ れ た が,か か る シ グナ ル が い か な る 機 序 で 細 胞 を 活 性 化 し,ど の よ う な 化 学 伝 達 物 質 を 産 生 ・
放 出 して い る か を 検 討 して い く必 要 が あ る.例 え ば 好 塩 基 球 のIgEレ セ プ ター をcross-linkさ せ た と き に み られ る ヒ ス タ ミ ン遊 離 が こ の 場 合 に も 認 め ら れ る の か 否 か は, IgG抗 体 に よ る好 塩 基 球 の 活 性 化 を 考 え る 上 で 大 変 興 味 深 い 問 題 で あ る. KU812-Fの ヒ ス タ ミ ン 含 有 量 は,ヒ ト 好 塩 基 球 に 比 し少 量 で あ るが9,19), A23187, PMA な ど の 非 生 理 的 刺 激 に よ っ て ヒ ス タ ミ ン が 遊 離 す る こ と が 認 め られ て お り19),今 回 の 実 験 で も, A23187に よ る 明 らか な ヒ ス タ ミン 遊 離 が み ら れ た.し か し な が ら, Agg-IgGを 添 加 し た 場 合, 並 び にIV. 3を 結 合 させ 二 次 抗 体 でcross-linkさ せ た 場 合 の い ず れ に お い て も ヒ ス タ ミン は 遊 離 せ ず, IgGレ セ プ タ ー を 介 す る刺 激 で は 有 意 な ヒ ス タ ミ ン の 遊 離 は 認 め ら れ な い と い う 結 果 で あ っ た. 従 来 か らIgG抗 体 に よ る好 塩 基 球 ・肥 満 細 胞 系 か ら の ヒ ス タ ミ ン 遊 離 に つ い て は,動 物 の 種 に よ っ て 差 の あ る こ とが 知 ら れ て い る.例 え ば, モ ル モ ッ ト,マ ウ ス の 肥 満 細 胞 で は, IgG1抗 体 に よ る ヒ ス タ ミン 遊 離 がFcγRを 介 して 起 こ っ て い る こ と が 確 認 さ れ て い る が,ヒ トに お い て は も と よ り異 論 の あ る と こ ろ で あ り,未 だ に 一 定 の 知 見 が え ら れ て い な い, 1970年Parish26)が, 即 時 型 反 応 を 起 こす ヒ トIgGの 存 在 を示 し, IgG short-term sensitizing antibody(IgG-STS)の
概 念 を提 唱 し て 以 来, anti-lgGに よ っ て ヒ ト好 塩 基 球 か ら の ヒ ス タ ミ ン 遊 離 が み られ た と す る 実 験 結 果 が 相 次 い で 出 さ れ た27,28,29).特 に モ ノ ク ロ ー ナ ル 抗 体 を 用 い た 実 験 で, anti-IgG4抗 体 に よ り好 塩 基 球 か ら ヒ ス タ ミ ン遊 離 が 認 め ら れ た と す る報 告 が 複 数 の 研 究 者 か ら 出 さ れ, IgG4 抗 体 とI型 ア レ ル ギ ー との 関 連 が 注 目 さ れ る よ うに な っ た28,29).一 方 こ れ と は 逆 に, IgG抗 体 は 好 塩 基 球 に 固 着 し得 ず,ヒ ス タ ミ ン 遊 離 は 起 こ ら な い とす る 報 告 も い くつ か み られ30,31),ま た, anti-IgGに よ る ヒ ス タ ミン 遊 離 の 機 序 に つ い て も不 明 な 点 が 多 く,ま だ 仮 説 の 域 を 出 て い な い の が 現 状 で あ る. Ishizakaら32)は,ラ ジ オ ア イ ソ トー プ を 用 い た 実 験 で,好 塩 基 球 に は monomeric IgGは 結 合 せ ず, Agg-IgGな ど の 多 価IgGの み 結 合 し う る が, Agg-IgGが 結 合 し て も ヒ ス タ ミ ン 遊 離 は 起 こ ら な い こ と を 報 告 し て い る.さ ら に, Lichtensteinら33)は ア ト ピ ー 患 者 及 び 健 常 者 の 好 塩 基 球 と血 清 を 用 い て 実 験 を 行 い, anti-IgGに よ る ヒ ス タ ミ ン 遊 離 は, IgEに 対 す る 自 己 抗 体(IgG)を 介 し て 間 接 的 に 好 塩 基 球 上 のIgGレ セ プ タ ー をcross-linkし た 結 果 起 こ っ た もの で あ る可 能 性 が 強 く, FcγR IIの 関 与 は 考 え に く い と し て い る. KU812-F細 胞 のFcγRIIを 介 す る刺 激 で 細 胞 内Ca2+濃 度 の 上 昇 が み ら れ た に も拘 ら ず ヒ ス タ ミン 遊 離 が 起 こ ら な か っ た と い う今 回 の 実 験 結 果 は, Lichten steinら の 見 解 を裏 付 け る も の と考 え られ,好 塩 基 球 に お い て, FcγRIIを 介 し たanti-IgGに よ る ヒ ス タ ミ ン遊 離 の 機 序 を 想 定 す る こ と は,些 か 困 難 が あ る と思 わ れ る.た だ し, KU812-F自 体 未 熟 な 細 胞 で あ り, IgEレ セ プ タ ー の 明 ら か な 発 現 が み られ な い た め,そ れ を 介 す る 刺 激 伝 達 を確 認 す る こ とが で き な か っ た こ と を 考 え る と,脱 顆 粒 に 至 る シ グ ナ ル 伝 達 機 構 が 未 成 熟 で あ る 可 能 性 も否 定 で き ず,今 後,細 胞 を 分 化 成 熟 させ, IgEレ セ プ タ ー を 増 加 さ せ た 段 階 で の 検 討 が 必 要 と思 わ れ る. ま た, FcγRIIを 介 す る 刺 激 が ヒ ス タ ミン 遊 離 に 直 接 関 与 し て い な い と し て も,産 生 機 構 の 異 な る他 の 化 学 伝 達 物 質,あ る い は サ イ トカ イ ン の 放 出 に 関 与 す る 可 能 性 が 想 定 さ れ る.実 際, 好 塩 基 球 と 同 じ くFcγRIIの 選 択 的 発 現 が 認 め ら れ て い る好 酸 球 で は, IgG刺 激 で ロ イ コ ト リ エ ン の 産 生 が 報 告 さ れ て お り34),ま た マ ウ ス の 脾 臓,骨 髄 由 来 の 細 胞 を用 い た 実 験 で,好 塩 基 球 ・ 肥 満 細 胞 系 と考 え ら れ るFcεRI陽 性 細 胞 の FcγRIIをIgG抗 体 でcross-linkさ せ る と, IL -4が 産 生 さ れ る と の 報 告 も み ら れ る35).今 後, IgG刺 激 に よ る好 塩 基 球 の 活 性 化 機 構 解 明 の 一 助 と して, KU812-Fで の これ らの 化 学 伝 達 物 質, サ イ ト カ イ ン を 含 め た 幅 広 い 細 胞 活 性 化 の 検 討 が 必 要 と思 わ れ る. 結 論 IgE以 外 の ア レ ル ギ ー 反 応 に お け る 好 塩 基 球 の 役 割 を解 明 す る 研 究 の 一 環 と し て, IgGレ セ プ タ ー を介 す る 細 胞 の 活 性 化 機 構 を 明 ら か に す る 目 的 で,ヒ ト好 塩 基 球 性 白 血 病 細 胞 株KU812 -Fを 用 い て 検 討 し ,以 下 の 結 果 を 得 た.
1) KU812-Fに は,ヒ ト末 梢 血 好 塩 基 球 と同 様, IgGレ セ プ タ ー の サ ブ タ イプ の 一 つ で あ る FcγRIIの 明 ら か な 発 現 が 認 め られ た. 2) KU812-FをAgg-IgGで 刺 激 す る と,細 胞 内Ca2+濃 度 の 有 意 な 上 昇 が み ら れ た.こ の 上 昇 は 抗FcγRII抗 体IV. 3に よ っ て 阻 止 さ れ, ま たIV. 3を 加 え た後,二 次 抗 体 でcross-linkさ せ る と著 明 な細 胞 内Ca2+濃 度 の 上 昇 が 認 め られ た. 3) KU812-FにIgGレ セ プ タ ー を 介 す る刺 激 を加 え て も,有 意 な ヒ ス タ ミ ン 遊 離 は 認 め ら れ な か っ た. 以 上 の 結 果 よ り, KU812-Fで はFcγRIIの cross-linkingに よ っ て 細 胞 内 シ グ ナ ル 伝 達 が 行 わ れ,何 ら か の 細 胞 の 活 性 化 が 起 こ っ て い る と 推 察 さ れ, IgGレ セ プ ター を介 す る 好 塩 基 球 の 機 能 発 現 を 考 え る 上 で 示 唆 に 富 む 重 要 な 所 見 で あ る と思 わ れ た. 稿 を終 え るに あ た り,終 始 御 指 導 御 校 閲 を賜 っ た 恩 師 木 村 郁 郎 教 授 に 深 甚 の 謝 意 を表 す る と共 に,直 接 御 指 導 載 い た 高 橋 清 講 師 に 深 謝 い た し ます. (本論 文 の 要 旨は 第42回 ア レ ル ギ ー 学 会 総 会 に て 発 表 した.)
文
献
1) Kimura I, Tanizaki
Y, Sato S and Takahashi
K: Differences in response to anti-IgE and to anti-IgG
in basophils from patients with bronchial asthma.
Clin Allergy
(1981) 11, 31-36.
2) 木 村 郁 郎,谷 崎勝 朗,高 橋 清:気 管 支 喘 息 に お け る抗 免疫 グ ロ ブ リン に 対 す る好 塩 基 球 の 反 応 性 に つ い て -抗 ヒ トIgGに 対 す る反 応 性 か ら み た 分 類 の 可 能 性- .ア レ ル ギー(1976) 25, 70-75. 3) 木 村 郁 郎,谷 崎 勝 朗,佐 藤 周 一,高 橋 清,上 田 暢 男,斉 藤 勝 剛: In vitro免 疫 下 に お け る好塩 基 球 の 生 態 観 察-気 管 支 喘 息 患 者 の 抗 ヒ トIgGに 対 す る 反 応性-.ア レル ギー(1976) 25, 489-492. 4) 小 林 誠:免 疫 反 応 下 に お け る好 塩 基球 の 形 態 学 的 研 究 第2編 抗 ヒ ト免 疫 グ ロブ リに よ る好 塩 基 球 の 形 態 学 的 変 化 の 免 疫 走 査 電 子 顕 微 鏡 観 察.岡 山 医 誌(1979) 91, 749-761.
5) Kimura
I, Takahashi
K,
Soda R, Kishimoto
T and Matsuoka
T: Basophils
immunoglobulin
receptors in asthmatics
under immuno-scanning
electron microscopy.
Clin Allergy
(1985) 15, 9
-15.
6) 松 岡 孝:気 管 支 喘 息 患 者 好 塩 基 球 の免 疫 グ ロ ブ リン レセ プ ター とIgG抗 体 の 作 用 機 序 に 関 す る研 究 第 1編 重 症 難 治 性 喘 息 に お け るIgG抗 体 の役 割 に関 す る検 討.岡 山 医 誌(1979) 98, 525-535.
7) Huzinga TWJ, Roos D and Von dem Borne AEGKr: Neutrophil FcƒÁ Receptors: A two-way bridge
in the immune system. Blood (1990) 75, 1211-1214.
8) Takahashi K, Takata M, Suwaki T, Kawata N, Tanimoto Y, Soda R and Kimura I: New flow
cytometric method for surface phenotyping basophils from peripheral blood. J Immunol Methods
(1993) 162, 17-21.
9) Kishi K: A new leukemic cell line with Philadelphia chromosome characterized as basophil precur
sors. Leuk Res (1985) 9, 381-390.
10) Minta A, Kao JPY and Tsien RY: Fluorescent indicators for cytosolic calcium based on rhodamine
fluorescein chromophores. J Biol Chem (1989) 264, 8171-8178.
11) Kao JPY, Harootunian AT and Tsien RY: Photochemically generated cytosolic calcium pulses and
their detection by fluo-3. J Biol Chem (1989) 264, 8179-8184.
12) Deber CM, Tom-Kun J, Mack E and Grinstein S: Bromo-A 23187: a non fluorescent calcium
ionophore for use with fluorescent probes. Anal Biochem (1985) 146, 349-352.
13) Hesketh TR, Smith GA, Moore JP, Taylor MV and Metcalfe JC: Free cytoplasmic calcium
14) Vandenberghe PA and Ceuppens JL: Flow cytometric measurement of cytoplasmic free calcium in
human peripheral blood T lymphocytes with fluo-3, a new fluorescent calcium indicator. J Immunol
Methods (1990) 127, 197-205.
15) McBride P, Bradely D and Kaliner M: Evaluation of a radioimmunoassay for histamine measure
ment in biological fluids. J Allergy Clin Immunol (1988) 82, 638-646.
16) Hirai K, Morita Y, Misaki Y, Ohta K, Takaishi T, Suzuki S, Motoyoshi K and Miyamoto T:
Modulation of human basophil histamine release by hemopoietic growth factors. J Immunol (1988)
141, 3958-3964.
17) Anselmino LM, Perussia B and Thomas L: Human basophil selectively express the FcƒÁRII (CDw
32) subtype of lgG receptor. J Allergy Clin Immunol (1989) 84, 907-914.
18) Stain C, Stockinger H, Scharf M, Jaerer U, Goessinger H, Lechner K and Bettelheim P: Human
blood basophils display a unique phenotype including activation linked membrane structures. Blood
(1987) 70, 1872-1879.
19) Fukuda T, Kishi K, Ohnishi Y and Shibata A: Bipotential cell differentiation of KU-812: evidence
of a hybrid cell line that differentiates into basophils and macrophage-like cells. Blood (1987) 70,
612-619.
20) Valent P, Besemer J, Kishi K, Kaltenbrunnrer R, R, Kuhn B, Maurer D, Lechner K and Bettelheim
P: IL-3 promotes basophilic differentiation of KU812 cells through high affinity binding sites. J
Immunol (1990) 145, 1885-1889.
21) Kochan J, Pettine LF, Hakimi J, Kishi K and Kinet JP: Isolation of the gene coding for the alpha
subunit of the human high affinity IgE receptor. Nucleic Acids Res (1988) 16, 3584.
22) Ravetch JV and Kinet JP: Fc Receptors. Annu Rev Immunol (1991) 9, 457-492.
23) Van de Winkel JGJ and Anderson CL: Biology of human immunoglobulin receptors. J Leukocyte
Biol (1991) 49, 511-524.
24) Kimberly RP, Ahlstrom JW, Click ME and Edberg JC: The glycosyl phosphatidylinositol-linked
FcƒÁIII Rpmn mediates transmembrane signaling events distinct from FcƒÁRII. J Exp Med (1990)
171, 1239-1255.
25) Anderson GP and Anderson C: Signal transduction by the platelet Fc receptor. Blood (1990) 76,
1165-1172.
26) Parish W: Short-term anaphylactic IgG antibodies in human sera. Lancet (1970) 2, 590-592.
27) Grant JA and Lichtenstein LM: Reversed in vitro anaphhylaxis induced by anti-IgG: specificity of
the reaction and comparison with antigen-induced histamine release. J Immunol (1972) 109, 20
- 25.
28) Nakagawa T, Stadler BM, Heiner DC, Skvaril F and DeWeck AL: Flow cytometric analysis of
human basophil degranulation. II. Degranulation induced by anti-IgE, anti-IgG 4 and the calcium
ionophore A23187. Clin Allergy (1981) 11, 21-30.
29) Fagan DL, Slaughter CA, Capra JD and Sullivan TJ: Monoclonal antibodies to immunoglobulin G
4 induce histamine release from human basophils in vitro. J Allergy Clin Immunol (1982) 70, 399 -404 .
30) Van Toorenenbergen AW and Aalberse RC: IgG 4 and passive sensitization of basophil leukocytes.
Int Arch Allergy Appl Immunol (1981) 65, 432-440.
31) Devey ME and Panzni R: The IgG subclass antibodies to castor bean allergen in patients with
(1975) 5, 353-361.
32) Ishizaka T, Sterk AR and Ishizaka K: Demonstration of FcƒÁ receptors on human basophil
granulocytes. J Immunol (1979) 123, 578-583.
33) Lichtenstein LM, Sobotka AK, White JM and Hamilton R. G: Anti-human IgG causes basophil
histamine release by acting on IgG-IgE complexes bound to IgE receptors. J Immunol (1992) 148,
3929-3936.
34) Shaw RJ, Walsh GM, Cromwell O, Moqbel R, Spry CJF and Kay AB: Activated human eosinophils
generate SRS-A leukotriens following IgG-dependent stimulation. Nature (1985) 316, 150-152. 35) Seder RA, Paul WE, Ben-Sasson SZ, LeGros GS, Sobotka AK, Finkelman FD, Pierce JH and Plaut
M: Production of interleukin-4 and other cytokines following stimulation of mast cell lines and in
The mechanism
of human
basophil
activation
associated
with
IgG receptor:
Analysis
of calcium
mobilization
in the human
basophilic
leukemia
cell line
KU812-F
Noriko
KAWATA
Second Department
of Internal
Medicine,
Okayama
University
Medical
School,
Okayama
700, Japan
(Director:
Prof. I. Kimura)
To clarify the mechanism of human basophil activation via IgG receptors, calcium mobiliza tion in response to IgG antibodies was analyzed in the human basophilic leukemia cell line KU812-F, using flow cytometry. KU812-F cells as well as human basophils selectively expres sed the FcƒÁRII subtype of IgG receptor. After stimulation with aggregated IgG, an obvious increase in [Ca2+] 1 was observed, but the increase was completely inhibited by IV. 3 (anti FcƒÁRIImAb). Moreover, IV. 3 elicited a [Ca2+]1 rise only when cross-linked on the cell surface with anti-mouse IgG. No significant histamine release was observed after any IgG stimulation and the biologic function of the FcƒÁRII-induced [Ca2+] 1 rise remains unclear.
These findings suggest that the cross-linking of FcƒÁRII in KU812-F cells induces signal transduction events and initiates cell activation with the exception of histamine release, and human basophils also may be activated in vivo by IgG antibodies.