人工容器を用いた
ウズラ胚の培養法に関する研究
2014年
目次 第1章 序論 1 第2章 培養開始時期の違いと培養容器の空気層の有無が胚に与える影響 13 第3章 人工容器での培養開始時期が胚の外部計測値とカルシウムおよび マグネシウムの含有量に与える影響 30 第4章 人工容器による培養開始時期の違いが奇形の発生に及ぼす影響 46 要旨 61 Summary 66 謝辞 70 引用文献 71
1 第1章 序論 1節 生物の多様性 人間の活動範囲の広がりに伴う生息域の分断、環境汚染、外来生物による捕 食や競合種の移入等生息環境の悪化により絶滅及び絶滅の危機に瀕している野 生動物は日々増加している。 国際自然保護連合(IUCN)など 130 カ国 1,800 人以上の研究者が参加した国際チ ームは、世界中の 5487 種の哺乳類が危機的な状態にあり、データ不足(DD: Data Deficient)の 836 種を除いた残りの 4651 種のうち、西暦 1500 年から現在まで、 76 種が絶滅(EX: Extinct)、2 種が野生絶滅 (EW: Extinct in the Wild)、188 種が絶 滅寸前(CE: Critically Endangered)、448 種が絶滅危機 (EN: Endangered)、505 種が 危急 (VU: Vulnerable) および 1141 種が絶滅危惧種であることを報告している (Schipper et al., 2008)。また、鳥類においても同様に、多くの種が絶滅や絶滅の 危機に瀕しており、西暦 1500 年から現在までに 279 種が絶滅したという試算が ある(Szabo et al., 2012)。
このような現状を踏まえ、1993 年に発効された生物の多様性に関する条約 (Convention on Biological Diversity: CBD)により生物多様性の保全が行われてい る。生物の多様性に関する条約において、生物多様性の保全は、生態系や自然 の生息地を保全する「生息域内保全」を原則とし、「生息域外保全」はその補 完的措置として位置づけられている。しかしながら、上記のように多くの動物 が絶滅の危機に瀕している現在、生息域内保全だけではなく動物園や保護施設 などでの生息域外保全も積極的に行われている。 このような野生動物の保護や保全に対しては、畜産学が培ってきた様々な知 識や技術の活用が求められている。
2 2節 哺乳類および鳥類の人工繁殖技術 哺乳類の人工繁殖技術における最初の科学的記録は、イタリアの生物学者 Spallamzani (1781)が犬の人工授精に成功したことに始まる。その後、ロシアの生 理学者である Ivanov (1907)が各種の家畜について人工授精を試み貴重な成果を 上げた。第 2 次世界大戦後には、乳肉の需要の高まりとともに家畜の人工授精 に関する基礎的および応用的研究が発展した。特に、Polge and Rowson (1952)が 牛精子の凍結保存に成功して以来、畜産技術の発達とともに様々な動物の生殖 細胞(精子、卵子および受精卵)が凍結保存できるようになり、凍結精子を用いた 人工授精や凍結受精卵の移植や凍結した配偶子間での体外受精が可能となった。
野生動物における人工授精は、Kraemer and Cruz (1972)がヒヒで初めて成功し て以来、多くの野生動物で試みられている。また、胚移植は Kraemer et al.(1976) がヒヒで成功し、体外受精は Clayton and Kuehl (1984)がヒヒで成功している。 Martha et al.(2004)は、アフリカヤマネコの体細胞クローンの作出に成功し、その クローン同士から自然繁殖で子ネコを得ている。また、2011 年にはサンディエ ゴ動物園においてキタシロサイとマンドリルの近縁種において iPS 細胞が樹立 されたという報道がされている(Ben-Nun et al., 2011)。
一方、哺乳類におけるトランスジェニック動物は、Jaenisch and Mintz (1974) がマウスを対象として初めて作出した。その後、Gordon et al. (1980)によりマウ スの受精卵前核に DNA を注入する方法がはじめて実践された。ラット、ウサギ およびスンクス等の実験動物の胚を体外で培養する全胚培養法も確立されてい る。また、マウス(Evans and Kaufman, 1981)やヒト(Thomson et al., 1998)において ES 細胞が樹立され、マウスでは樹立した ES 細胞を用いたノックアウトマウス の作出が盛んに行われている。
このように、哺乳類では実験動物や家畜だけでなく、野生動物に対しても発 生工学的な手法が活用されている。
3 これに対し、鳥類では古くから孵卵器を用いた人工孵化が行われている。家 禽では人工授精や精液の凍結保存などの技術は確立しており、シチメンチョウ やホロホロチョウでは人工授精が産業的に行われている。しかしながら、生殖 細胞の凍結保存技術は精子に関してのみであり、卵子や受精卵については大量 の卵黄物質を含んでいる巨大細胞であることから凍結保存の方法は確立されて いない。また、受精卵を活用したクローン技術も確立されていない。 一方、ニワトリでは生殖細胞の幹細胞である始原生殖細胞(Primodial Germ Cell: PGC)が孵卵48~55時間後(Stage13-15)をピークとして胚の血液中に存在し ている(Tajima et al., 1999)ことから、胚盤葉期胚の胚盤葉明域や孵卵48~55時間 後の胚の血液中に存在するPGCs(Primordial Germ Cells)を採取し、ホスト胚の血 液中に注入して生殖系列キメラを作成(図1-1)し、成長した後にドナーと同種の 鳥を交配することにより希少種を復元、増殖する技術がニワトリにおいて確立 されている(Tajima et al.,1993)。また、Pain et al. (1996)は、ニワトリのES細胞を 樹立し、生殖細胞へと分化させている。一方、発生工学において必須である細 胞培養の手法として、Kuwana et al. (1996)は高カルシウムの培養液(KAv-1)を作 成し、その培養液を用いることでそれまで困難であった鳥類の体細胞やPGCsの 長期培養および増殖法を確立している。 以上のことから、生殖系列キメラを用いた個体復元、体細胞やPGCsの長期培 養や増殖法の確立、また、ES細胞樹立等の技術により、将来的には凍結保存し た希少鳥類の細胞から個体復元が可能になると期待されている。将来の個体復 元や増殖を目指し、国立環境研究所では、遺伝子資源保存のため平成14年より 開始した環境試料タイムカプセル化事業(環境省委託事業)により様々な絶滅危 惧動物の精子や卵子、受精卵、体細胞等の凍結保存を行っている(桑名, 2006)。 しかしながら、将来の個体復元や増殖には前述の技術の他に様々な周辺技術の 開発が必要である。例えば、作製した生殖系列キメラから効率よくドナーPGC
4
由来の後代を得るために、ホスト胚に内在的に存在するPGCを除去あるいは減 少させる方法が検討されている。すなわち、胚盤葉明域中央部の細胞除去 (Kagami et al., 1997)や2.5日胚の血液除去(Naito et al., 1994; Tajima et al., 1998)と いった外科的除去法、軟X線(Nakamichi et al., 2007; Atsumi et al., 2008)や紫外線 (Reynaud, 1976)、ガンマー線(Carsience et al., 1993; Kagami et al., 1995)を放卵直後 の胚や2日胚に照射し、内在するPGCsを不活化する物理的除去法、そしてコナ カバリンA(Lee et al.,1978)、ブスルファン(Aige-Gil and Simkiss, 1991; Hallett and Wentworth, 1991; Song et al., 2005)や乳化ブスルファン(Nakamura et al., 2008)を放 卵直後の胚に投与し、生殖細胞を不活化する化学的除去法などである。また、 筆者はホストに生殖能力を持たない種間雑種を用いることを提案している。 このように、鳥類の発生工学的な増殖法が数多く開発改良されているが、操 作後の胚は、培養するための卵殻が必要となってくる。 鳥類の卵は卵殻を有しており、受精から孵化までの過程の大部分が卵殻内で 進行することから、卵殻に異常のある卵(軟卵・破卵)や、卵管内から取り出した 卵殻が形成される前の卵を孵化させることは困難である。鳥類の卵殻は胚発生 において主に3つの重要な役割を果たしている。すなわち、卵殻を構成する主 要なミネラルは炭酸カルシウム 98.43%、炭酸マグネシウム 0.84%、リン酸カル シウム 0.73%であり(Romanoff, 1949a)、胚は発生に必要なミネラルの 80%を卵 殻から吸収している(Johnston and Comar, 1955; 藤田ら, 2007a)。また、卵殻は卵 殻孔を通じて卵殻内外のガス交換の調節機能も有している(Romanoff, 1949b)。さ らに、胚発生を安全に行うための器としての役割がある。このように鳥類胚に とって、卵殻はミネラル供給源、ガス調整機能および培養容器としての機能を 有している。
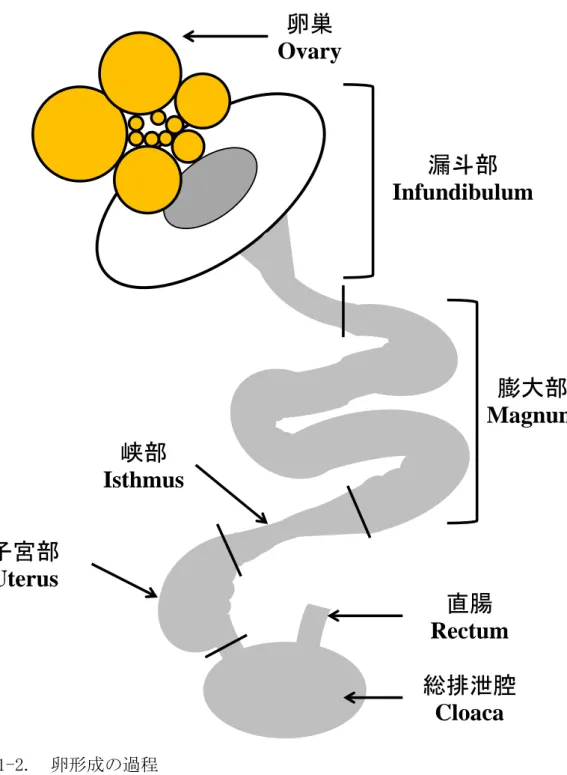
家禽における排卵から孵化までの過程(図 1-2)は、受精が排卵後 15 分以内に漏 斗部で行われる(Olsen and Neher, 1948)。また、漏斗部での受精後 4.5 時間後に最
5
初の卵割が始まり、続く膨大部、峡部および卵殻腺部でそれぞれ卵白、卵殻膜 および卵殻が形成される(Perry, 1987)。胚は、母鳥の体外に放出(放卵)される時点 では、細胞数が 50,000 から 60,000 までに成長している(Spratt and Haas, 1961)。 このため、鳥類胚への遺伝子導入を確実に行う際には 1 細胞期の受精卵を使用 する必要がある。また、胚操作後に、卵白や卵殻膜、卵殻を形成させる必要が あることから、他個体の輸卵管へ操作した胚の移植が試みられたが、操作は困 難であり、成功率も低かった(Olsen and Neher, 1948)。
6
図1-1. キメラ作出の過程
Fig.1-1. Production of chimeric chicken
ドナーニワトリ Donor chicken
胚盤葉キメラ blastogerm cells chimera
始原生殖細胞(PGCs)キメラ PGC chimera ドナー細胞の採集方法 Collecting of donor cells ホストニワトリ Host chicken 胚盤葉明域中央部を採取 Removal of blastogerm cells
血流中のPGCsを採取 Collection of the PGCs
from circulating blood
作出されたキメラ Production of germline chimera 羽装および生殖巣キメラ Somatic and germline chimera 生殖巣キメラ Germline chimera ホスト胚への移植方法 Injection to host embryo 胚盤葉明域中央部に移植 Injection of cells into blastogerm
血流中に移植
Injection of cells into vasculature
得られる配偶子 Gamete from chimera
ドナー由来の配偶子形成 Donor-derived gamete
卵子 Ovum
精子
7
図 1-2. 卵形成の過程
Fig. 1-2. Process of egg formation
膨大部
Magnum
漏斗部
Infundibulum
卵巣
Ovary
峡部
Isthmus
子宮部
Uterus
総排泄腔
Cloaca
直腸
Rectum
8
3節 鳥類胚の卵殻外培養の歴史
鳥類胚の卵殻外培養は、孵卵 60 時間後のニホンウズラ胚(Ono and Wakasugi, 1984)や孵卵 72 時間後のニワトリ胚(Rowlett and Simkiss, 1985; 1987)をそれぞれ 代理卵殻で培養し孵化したことが報告されている。しかし、いずれの報告も孵 卵後 2~3 日経過した胚からの培養であり、鳥類胚の卵殻外培養としては完全な ものとはいえなかった。 この後、Perry(1988)の開発した代理卵殻法と呼ばれる培養法がその後の卵殻外 培養の基本となった。この代理卵殻法は、ニワトリの体内から取り出した 1 細 胞期の受精卵を人工容器と代理卵殻を用いて培養する方法である。すなわち、 ニワトリ胚における受精から孵化までのステージを System I(受精から胚盤葉形 成: 受精後 24 時間)、System II (胚形成:胚盤葉期から 3 日間)および System III (胚 成長から孵化:3 日目以降)の 3 つの段階に分け、System I はガラス容器、System II とそれに続く System III は代理卵殻を用いて培養する方法である。
開発当初は、System IからSystem IIIを通じて培養を行い孵化率7%であった (Perry, 1988)が、Naito et al.(1990)によって改良を加えられ、50%近い孵化率に至 っている。その後、代理卵殻法は、ニホンウズラ(Ono et al., 1994)、ヒメウズラ (Ono et al., 2005)およびボブホワイト(Kato et al., 2013)への応用が検討され孵化 が報告されている。 しかしながら、上記の報告は主にニワトリやウズラといった家禽を対象とし ており、代理卵殻も家禽由来の卵殻を使用している。 希少野生鳥類保全の現場である動物園などの施設では、食癖、踏みつけ、巣 からの排除等の行動により貴重な受精卵が破損することがしばしば見受けられ る。Perry(1988)の代理卵殻法は、受精卵に外来遺伝子を導入したトランスジェニ ックニワトリを作出する際に必要な技術として開発されたが、この技術は破卵 や卵殻異常、死亡個体の卵管内受精卵の発生、さらには胚盤葉キメラや生殖系
9 列キメラ等の発生工学的手法による増殖法に用いることで希少鳥類の保護・保 全へ応用することができる。また、卵子(卵黄)を代理卵殻で培養し孵化まで至ら なくとも、発生初期にPGCsを回収するといった活用が期待される。しかしなが ら、代理卵殻法を希少鳥類の保護に適用しようとした場合、鳥類種によって様々 な卵殻サイズがあり(最小はハチドリの卵重量0.5gから最大はダチョウの卵重量 1.3~1.6kg)、完全に対応する事が難しい。また、細菌等の混入を防ぐために卵殻 を完全滅菌する事はほぼ不可能であることや卵殻はもろく壊れやすいため取扱 が困難であるという問題点が挙げられる。 そのため、代理卵殻法における問題点を解決し発展させうる方法として人工 容器を用いた胚培養法が検討されてきた。表1-1および表1-2は、先行研究および 本研究における人工容器での培養条件および培養開始時期を示している。
Ono et al. (1994)は、ニホンウズラの卵管内受精卵をPerry(1988)のSystem Iに準 じた方法で培養を行い、約80%の胚が1細胞期から胚盤葉期への発生に至ったと 報告している。この方法では培養容器にプラスチックカップ、培養液としてニ ワトリの水様性卵白を用いているが、この時期の卵子は体内に留まっているこ とと卵割期にあることから、成長に必要なミネラル供給源、ガス調整機能とし ての卵殻の機能は付与していない。
Kawashima et al. (2005) は、続くSystem IIの培養においてニワトリおよびニホン ウズラを対象として、人工容器にプラスチックケースを、人工卵殻膜としてポ リテトラフルオロエチレン (PTFE) 膜を、培養液としてニワトリの水様性卵白 をそれぞれ用いることで胚の83%が胚盤葉期から胚形成までの発生に至ってい る 。この時期の胚は胚盤葉期からStage 20(Hamburger and Hamilton, 1951)程度ま で発生が進行する。しかしこの時期は、胚のサイズやミネラル含有量(Ono and Wakasugi, 1984)が急激な増加を示す時期よりも前の段階であることから、ミネラ ル供給源としての卵殻の機能は付与していない。
10
Kamihira et al. (1998)は、System IIIの培養においてニホンウズラを対象として、 人工容器に空気を流通させる穴をあけたプラスチックカップを、人工卵殻膜と してPTFE膜を、上蓋としてポリ塩化ビニリデン製のサランラップを、培養液と して乳酸カルシウムと卵殻粉末を添加したニホンウズラの水様性卵白をそれぞ れ用いて、43%が胚成長から孵化に至っている。この方法では卵殻が有するミネ ラル供給源、ガス調整機能および培養容器のすべての機能を付与している。 以上のように、ニホンウズラにおいては1細胞期からの各段階に対して人工容 器を用いて胚発生を行わせる事ができるようになった。しかし、これらの方法 を用いて1細胞期または胚盤葉期から孵化予定日まで培養した報告は見当たら ず、孵化の例もない。 これまでの知見として、ニホンウズラ胚をSystem IIIから人工容器で培養した 場合、孵化に必要なミネラルを添加することで孵化率が向上するものの、得ら れた胚の重量は、通常孵卵した胚に比べ軽いこと(Kamihira et al., 1998)、また、 使用する人工容器のサイズは供試する胚が本来持っている卵殻サイズとほぼ同 じ大きさの場合に生存率が高いこと(Kawashima et al., 2005)などが報告されてい る。 そこで本研究ではニホンウズラの受精卵を用い、卵殻としてミネラル供給源、 ガス調整機能および培養容器の三つの機能を付与した Kamihira et al. (1998)の方 法を参考にして胚盤葉期から培養を行い、人工容器による胚培養が胚に与える 影響を調べることで、人工容器培養技術の問題点や胚盤葉期からの人工容器培 養における好適な培養条件を明らかにしようとした。
11
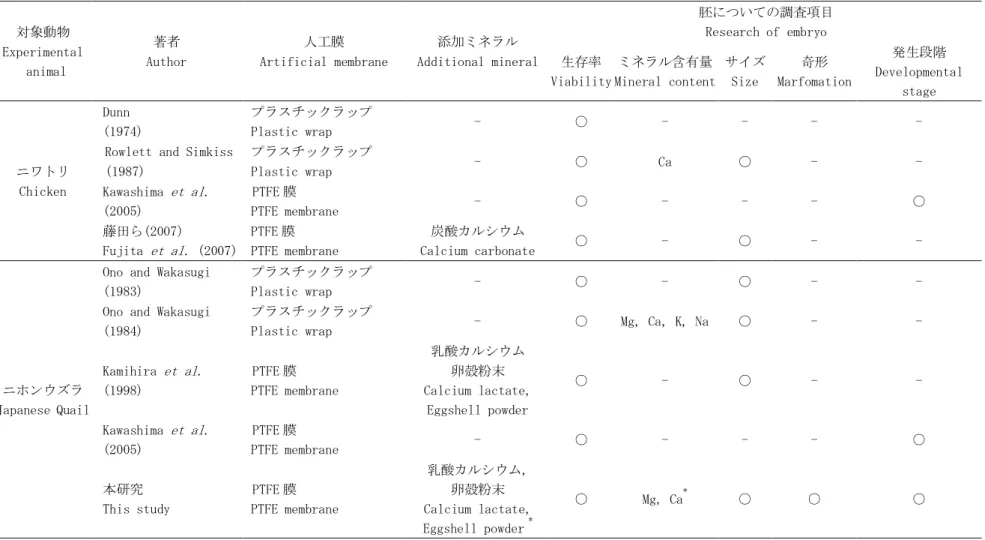
表 1-1. 鳥類胚の人工容器による培養方法と調査項目
Table. 1-1. Methods and Research Items in Avian Embryo Culture System using Artificial Vessel
対象動物 Experimental animal 著者 Author 人工膜 Artificial membrane 添加ミネラル Additional mineral 胚についての調査項目 Research of embryo 生存率 Viability ミネラル含有量 Mineral content サイズ Size 奇形 Marfomation 発生段階 Developmental stage ニワトリ Chicken Dunn (1974) プラスチックラップ Plastic wrap - ○ - - - -
Rowlett and Simkiss (1987) プラスチックラップ Plastic wrap - ○ Ca ○ - - Kawashima et al. (2005) PTFE 膜 PTFE membrane - ○ - - - ○ 藤田ら(2007) Fujita et al. (2007) PTFE 膜 PTFE membrane 炭酸カルシウム Calcium carbonate ○ - ○ - - ニホンウズラ Japanese Quail
Ono and Wakasugi (1983)
プラスチックラップ
Plastic wrap - ○ - ○ - -
Ono and Wakasugi (1984)
プラスチックラップ
Plastic wrap - ○ Mg, Ca, K, Na ○ - -
Kamihira et al. (1998) PTFE 膜 PTFE membrane 乳酸カルシウム 卵殻粉末 Calcium lactate, Eggshell powder ○ - ○ - - Kawashima et al. (2005) PTFE 膜 PTFE membrane - ○ - - - ○ 本研究 This study PTFE 膜 PTFE membrane 乳酸カルシウム, 卵殻粉末 Calcium lactate, Eggshell powder * ○ Mg, Ca* ○ ○ ○ -: 試行なし, -: No experiment
12
表1-2.先行研究および本研究における人工容器培養の開始時期と培養期間 Table.1-2. Onset of artificial vessel culture and culture period
対象動物 Experimental animal
著者 Author
人工容器での培養開始時期 Onset of artificial vessel culture
孵化予定日 Expected hatching day 胚盤葉期 Blastodermal stage 前孵卵の時間(h) Prior incubation (h) 27 48 55 60 72 ニワトリ Chicken Dunn (1974)
Rowlett and Simkiss (1987)
Kawashima et al. (2005)
藤田ら (2007)
Fujita et al. (2007)
ニホンウズラ Japanese quail
Ono and Wakasugi (1983) Ono and Wakasugi (1984)
Kamihira et al. (1998)
Kawashima et al. (2005)
本研究 This study
13
第2章 培養開始時期の違いと培養容器の空気層の有無が胚に与える影響
1. 緒言
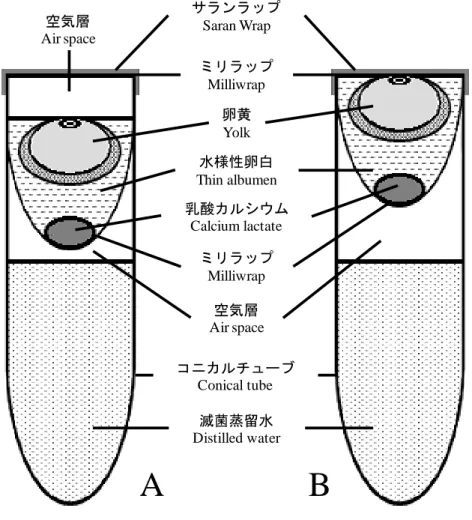
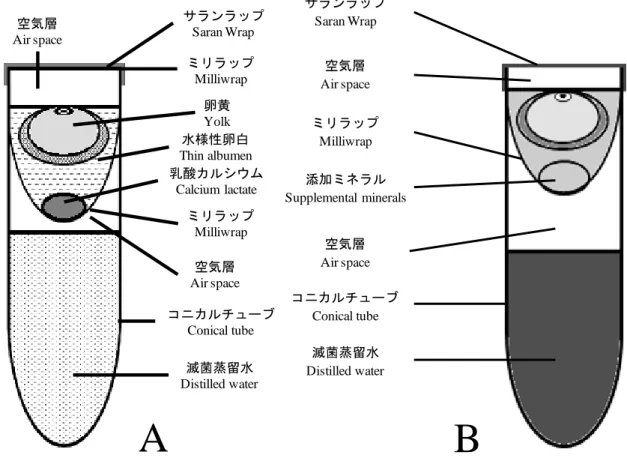
Perry(1988)の代理卵殻培養法を発展させた人工容器による培養法がニホンウ ズラにおいて検討され(System I: Ono et al.,1994; System II: Kawashima et al.,2005; System III: Kamihira et al.,1998)、各段階終了時の生存率が 80%、83%、43%とな っている。しかし、これらの研究は各 System に限定したものであり1細胞期ま たは胚盤葉期から一貫して人工容器で培養を行い、孵化まで至ったという報告 は見当たらず、また、胚盤葉期から培養した胚に関する報告も見当たらない。 そこで本実験では、ニホンウズラを対象として胚盤葉期および孵卵 60 時間後 (ニワトリの 72 時間後に相当)から人工容器を用いて培養を行い、培養環境が胚 にどのような影響を与えるのかを明らかにしようとした。 2. 材料および方法 (1) 供試卵 東海有機株式会社(Aichi, Japan)より購入したニホンウズラの受精卵 (2) 培養容器 実験で使用した培養容器を図 2-1 に示した。50 ml ポリプロピレン製チューブ (BD Falcon Conical Tube; Nihon BD Co, Tokyo, Japan. 以降コニカルチューブと表 記.)に培養器内の乾燥を防ぐため滅菌蒸留水を 35 ml 入れた後、多孔性でガス透 過性のあるポリテトラフルオロエチレン(PTFE)製の膜(Milliwrap; Nihon Millipore Co, Tokyo, Japan. 以降ミリラップと表記.)を袋状に取り付けたものを培養容器と した。培養容器にウズラ卵の内容物を移し替えたのち、空気層の有るものと無 いものを用意し上蓋をした。上蓋としてミリラップの上にポリ塩化ビニリデン
14
製フィルム(Saran Wrap; Asahi Kasei Life & Living Co, Tokyo, Japan. 以降サランラ ップと表記.)を重ねたものを用いた。なお、空気層は容器の開口部から 5~7 ㎜ 設けた。培養容器の材料はオートクレーブにより滅菌処理(121℃、20 分)を施し た。
(3) 胚培養
人工容器による培養開始時期は、胚盤葉期または孵卵 60 時間後とした。培養 液は Kamihira et al.(1998)を参考に、乳酸カルシウム(Sigma-Aldrich Co, Tokyo, Japan)35 mg をニワトリの水様性卵白(1.5 ml)に添加したものを使用した。また、 対照群として操作を加えず通常孵卵した胚を用いた。胚は、孵卵器(Type P-008; Showa Furanki Co, Tokyo, Japan) を用いて 38.0℃、湿度 60%、転卵 60°/時間の条 件下で培養した。孵化予定日の 2 日前には転卵を停止し、孵化予定日前日には 上蓋に穴をあけた。培養中の胚は、24 時間毎に生存の確認を行い、死亡した胚 を取り除いた。なお、血流、心臓の拍動または胎動が停止したものを死亡胚と した。また、死亡胚は尿漿膜と卵黄を取り除き、胚重量と第 3 趾長を計測した。 胚の発生段階は、Hamburger and Hamilton (1951)の報告を参考に決定した。
(4) 統計処理
空気層の有無による生存率の違いは、χ二乗検定を用いて有意水準5%として 検討した。また、Stage45で死亡した胚の胚重量と第3趾長はScheffé’s multiple comparison test を用いて有意水準5%として検討した。
15
図 2-1. 培養に用いた人工容器
A: 空気層を設けた場合, B: 空気層を設けなかった場合 人工容器はサ ランラップとミリラップにより蓋をしてある
Fig. 2-1. Quail embryo culture vessels
A: presence of airspace, B: absence of airspace. The vessels were sealed with Saran Wrap and Milliwrap.
サランラップ Saran Wrap ミリラップ Milliwrap 乳酸カルシウム Calcium lactate 滅菌蒸留水 Distilled water コニカルチューブ Conical tube 卵黄 Yolk 水様性卵白 Thin albumen
A
B
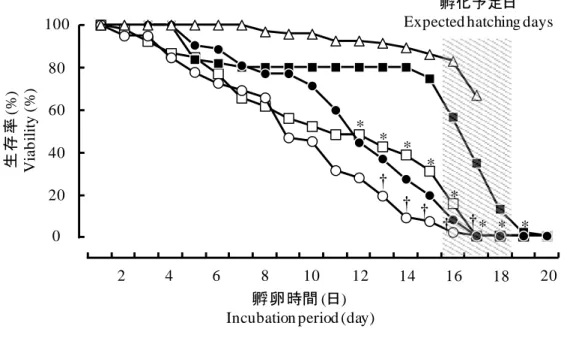
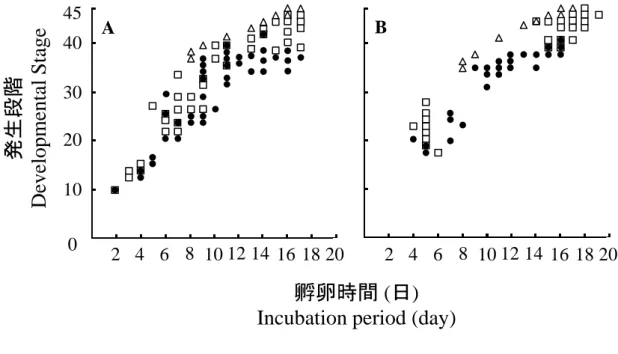
空気層 Air space ミリラップ Milliwrap 空気層 Air space16 3. 結果 (1) 胚盤葉期から人工容器で培養した胚の生存率と発生段階 図2-2はウズラ胚の生存率と孵化率を示している。空気層を設けた場合、胚の 生存率は1日目(100%, 52/52)から15日目(19.2%, 10/52)にかけて徐々に低下した。 全ての胚は17日目までに死亡し、孵化直前であるStage 45まで到達した胚は8体 (15.4%)であった。一方で、空気層を設けない場合、胚の生存率は1日目(100%、 58/58)から8日目(65.5%, 38/58)にかけて徐々に低下し、9日目(46.6%, 27/58)には急 激に生存率が低下した。全ての胚は17日目までに死亡し、孵化直前であるStage 45まで到達した胚は得られなかった。図2-3Aは死亡した胚の発生段階の分布を 示している。空気層を設けて培養した場合、16日目と17日目(15.4%, 8/52)におい て孵化直前のStage 45まで到達したが、空気層を設けないで培養した場合、14日 目(1.7%, 1/58)においてStage 42まで到達した。 (2) 孵卵 60 時間後から人工容器で培養した胚の生存率と発生段階 空気層を設けた場合、胚の生存率は 4 日目まで 100%であった(図 2-2)。6 日目 から 14 日目にかけて生存率は 80%以上であった。15 日目(74.5%, 41/55)から 17 日目(34.5%, 19/55)にかけて急激に生存率が低下した。全ての胚は 20 日までに死 亡し、孵化直前である Stage 45 は 10 体(18.2%, 10/55)得られた。一方で、空気層 を設けない場合、胚の生存率は 4 日目まで 100%であったが、10 日目(71.2%, 37/52) から 12 日目(44.2%, 23/52)にかけて急激に生存率を下げた。全ての胚は 17 日目 までに死亡し、Stage 45 まで到達した胚は得られなかった。図 2-3B は死亡した 胚の発生段階の分布を示している。空気層を設けて培養した場合、16 日目(1.8%, 1/55)において Stage 46 まで到達したが、空気層を設けないで培養した場合、16 日目(1.9%, 1/52)において Stage 40 まで到達した。
17 図 2-2. ウズラ胚の生存率 胚盤葉期から人工容器で培養した胚であり、□: 空気層を設けた場合(n = 52)または、○: 空気層を設けない場合(n = 58)、孵卵 60 時間後から 人工容器で培養した胚であり、■: 空気層を設けた場合 (n = 55) ま たは、●: 空気層を設けない場合 (n = 52)、△: 通常孵卵した胚 (n = 93)。 空気層の有無による生存率の違いは、χ 二乗検定を用いて有意水 準 5%として統計処理を行った。* 胚盤葉期から人工容器培養した胚、ま たは、†孵卵 60 時間後から人工容器で培養した胚
Fig. 2-2. Viability of quail embryos
In vitro culture was set up from the blastoderm stage in the □: presence (n = 52)
or ○: absence (n = 58) of airspace, and after 60 hours preincubation in the ■: presence (n = 55) or ●: absence (n = 52) of airspace; Intact fertilized eggs as Δ: controls (n = 93).
A significant difference (p <0.05) in viability was observed between the presence and absence of airspace in cultures from the *blastoderm stage or †60
hours preincubation, after analyses using the chi-square test.
2 4 6 8 10 12 14 16 18 20 0 20 40 60 80 100 孵卵時間 (日) Incubation period (day)
生存率 (% ) V ia b il it y ( % ) 孵化予定日 Expected hatching days
† * * * * * * * * † † † †
18
図 2-3. 死亡した胚の発生段階
A: 胚盤葉期から人工容器で培養した胚、B: 孵卵 60 時間後から人工容 器で培養した胚、□: 空気層を設けた場合、●: 空気層を設けなかった 場合、△: 通常孵卵した胚
Fig. 2-3. Developmental stages attained by dead quail embryos
Embryonic culture set up from the A: blastoderm stage or B: 60 hours preincubation. In vitro culture in the □: presence or ●: absence of airspace; Intact fertilized eggs as Δ: controls.
0 10 20 30 40
発生段階
Deve
lopment
al
Stag
e
45 A孵卵時間 (日)
Incubation period (day)
B
2 4 6 8 10 12 14 16 18 20 2 4 6 8 10 12 14 16 18 20
19 (3) 通常孵卵した胚の生存率と発生段階 胚の生存率は 7 日目まで 100%(93/93)であった(図 2-2)。8 日目(96.8%, 90/93) から 15 日目(86.7%, 80/93)にかけて徐々に生存率が低下した。胚は 16 日目から 17 日目にかけて孵化し(66.7%, 62/93)、Stage 45 で死亡した胚は 4 体(4.3%, 4/93) であった。図 2-3 は死亡した胚の発生段階の分布を示している。通常孵卵した 場合、16 日と 17 日目(11.8%, 11/93)に Stage 46 まで到達した。 (4) Stage 45 で死亡した胚の重量および第 3 趾長 図 2-4 は Stage 45 で死亡した胚の重量と第 3 趾長を示している。人工容器で 培養した胚の重量と第 3 趾長は、通常孵卵した胚に比べ低い値を示した(p < 0.05)。 また、胚盤葉期から人工容器で培養した胚の重量と第 3 趾長は、孵卵 60 時間後 から人工容器で培養した胚に比べ低い値を示した(p < 0.05)。 (5) 奇形 表 2-1 は、奇形の発生率を示している。奇形は、胚盤葉期から人工容器で培 養した胚にのみ発生し、空気層を設けた場合 17.3%(9/52)であり、空気層を設 けなかった場合 22.4%(13/58)であった。一方で、孵卵 60 時間後から人工容器で 培養した胚と通常孵卵した胚に奇形の発生は認められなかった。 表 2-2 と 図 2-5 は発生した奇形の種類と奇形の状態を示している。発生し た奇形の大部分は、 無眼球症や小眼球症といった眼の形成に関するもので、空 気 層 を 設 け た 場 合 は 69.2%(9/13) で あ り 、 空 気 層 を 設 け な か っ た 場 合 は 64.3%(9/14)であった。
20
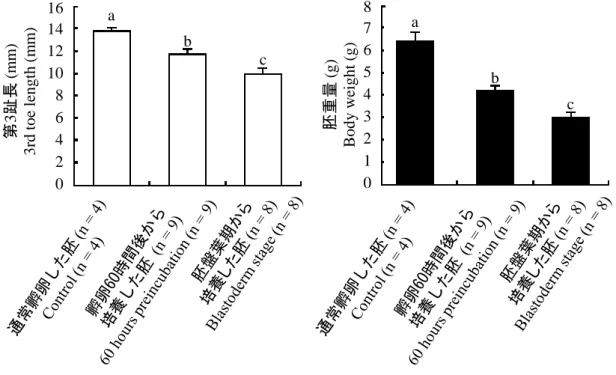
図 2-4. Stage 45 で死亡した胚の胚重量および第 3 趾長
カラムおよびエラーバーは平均値と標準誤差を示している。 Scheffé’s test を用いて統計処理を行った。 異文字間(A-B-C または a-b-c)に有意 な差が認められた(p < 0.05)。
Fig. 2-4. Average body weight and 3rd toe length of dead embryos at Stage 45
Open columns: 3rd toe length, Solid columns: body weight. Column and vertical lines represent the mean and standard error of mean, different letters represent significant differences (p < 0.05) as assessed by Scheffé’s test.
8 7 6 5 4 3 2 1 0 胚重量 (g ) Bo d y w ei g h t (g ) 16 14 12 10 8 6 4 2 0 第 3 趾長 (m m ) 3 rd t o e len g th (m m ) a b c a b c
21
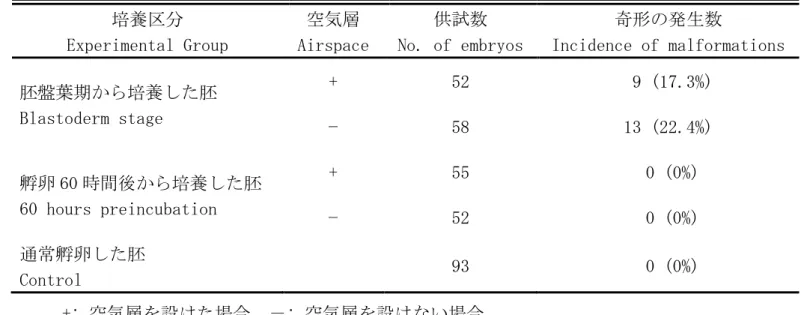
表 2-1. 培養した胚の奇形発生
Table. 2-1. Incidence of embryonic malformations 培養区分 Experimental Group 空気層 Airspace 供試数 No. of embryos 奇形の発生数 Incidence of malformations 胚盤葉期から培養した胚 Blastoderm stage + 52 9 (17.3%) − 58 13 (22.4%) 孵卵 60 時間後から培養した胚 60 hours preincubation + 55 0 (0%) − 52 0 (0%) 通常孵卵した胚 Control 93 0 (0%) +: 空気層を設けた場合, -: 空気層を設けない場合 +: Presence of airspace, −: Absence of airspace
22
表 2-2. 胚盤葉期から人工容器で培養した胚に認められた奇形の内訳
Table. 2-2. Types of malformations observed in cultures using blastoderm stage embryos 奇形の種類 Type of malformation 空気層 Airspace + − 無眼球症 および/または 小眼球症 9 (69.2%) 9 (64.3%) Anophthalmia and/or Microphthalmos
無頭蓋症 2 (15.4%) 3 (21.4%) Acrania 胸骨の形成不全 2 (15.4%) 1 (7.1%) Hypoplastic breastbone 脚の形成不全 0 (0%) 1 (7.1%) Digit hypoplasia 合計 Total 13 14 +: 空気層を設けた場合, -: 空気層を設けない場合 +: Presence of airspace, −: Absence of airspace
23
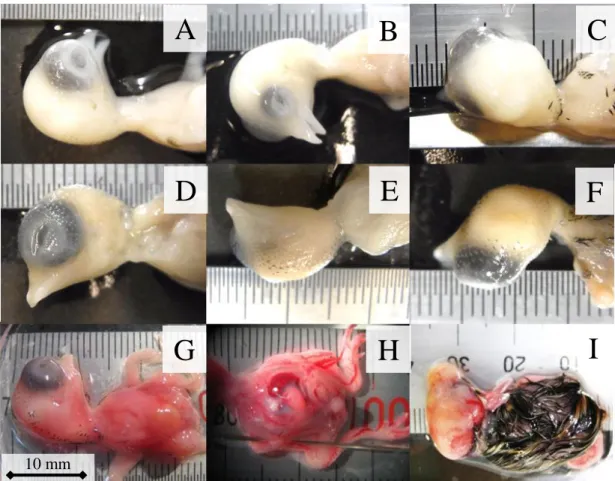
図 2-5. 発生した奇形の種類
A、 B、 C: 左眼が小眼球症(同一胚); D、E、F: 右眼が無眼球症(同一 胚); G: 正常な胚; H: 胸骨の形成不全による内臓露出; I: 頭蓋骨の形 成不全による脳の露出。
Fig. 2-5. Types of observed embryonic malformation
A, B and C: microphthalmos of left eye (same embryo); D, E and F: anophthalmia of right eye; G: normal embryo; H: evisceration caused by hypoplastic breastbone; I: brain exposure caused by acrania.
C
F
I
B
E
H
A
D
G
10 mm24 4. 考察 本実験において胚盤葉期から人工容器で培養した胚は空気層の有無に関係 なく、55 時間後の生存率が 90%以上であった。この結果はプラスチックの容器 と PTFE 膜を用い、空気層を排除した人工容器で、胚盤葉期からニホンウズラを 培養した Kawashima et al.(2005)の 55 時間後の生存率(55~83%)よりも高い値を 示した。また、韮澤ら(1992)はウズラ胚の代理卵殻培養において、System II に相 当する期間の培養時に空気層を設けると、空気層を排除した場合に比べ孵卵 3 日目の生存率が有意に低くなることを報告している。Perry(1988)の開発した代理 卵殻法では System II の培養において空気層を排除した条件で培養している。し かし、人工容器で培養した本実験では、空気層の有無によって胚盤葉期から孵 卵 60 時間後までの生存率に差は見られなかった。このことから、人工容器を用 いた培養において孵卵 60 時間後までは空気層の有無と胚の生存率は直接関係し ないことが考えられた。 孵卵 10 日目以降の生存率において有意な差は見られなかったが人工容器で の培養開始時期に関係なく空気層を設けなかった場合、空気層を設けた場合に 比べ低い値を示し、胚盤葉期から培養を開始した胚では 13 日目以降、孵卵 60 時間後から培養を開始した胚では 12 日目以降に空気層を設けなかった場合の生 存率が有意に(p < 0.05)低い値を示した。Perry(1988)は、System III の培養には空 気層の存在が必要であり、8 日目以降も胚が培養液中に没していると生存率が下 がることを報告している。このことから、人工容器を用いた培養において培養 3 日目以降は空気層の有無が胚の生存率に影響を及ぼすことが考えられた。 また、空気層を設けた場合、孵卵 60 時間後から人工容器で培養した胚は、15~16 日目にかけて生存率の急激な低下が認められた。ニホンウズラ胚の培養におけ る後期の生存率の低下は、人工容器(Ono et al.,1994; Kamihira et al., 1998)や代 理卵殻(韮澤ら, 1992)を用いた培養において報告されている。また、ニワトリに
25
おいても培養後期の生存率の低下は、人工容器(Dunn, 1974、 藤田ら, 2007b)や代 理卵殻(Kawashima et al.,2002、 藤田ら, 2007c) を用いた培養において報告されて いる。一方で、高い二酸化炭素濃度や低い酸素濃度の環境下で孵卵すると孵化 率が低下するとの報告もある(Taylor et al., 1956; 1971; Taylor and Kreutziger, 1965; 1966; 1969)。
Kamihira et al. (1998) は、孵卵 48 時間後からのウズラ胚の代理卵殻培養にお いて、通常の気相で培養した胚は孵化個体が得られなかったのに対し、孵卵 3 ~12 日目を 0.25L/min、12~15 日目を 0.5L/min および 15~17 日目を 0.7L/min と 3 段階に分けて酸素を孵卵器内に通気し、孵卵器内の酸素量を増加させることで 34%の孵化個体を得たことを報告している。ニワトリの胚発生においては、孵 卵から 19 日目に尿膜呼吸から肺呼吸へと移行するとされている(Kawashima et
al., 2002; Romanoff, 1960b)が、この時期はニホンウズラにおいては孵卵 15 日目に
相当する(Zacchei, 1961)。一方、Ruijtenbeek et al.(2000)は、ニワトリ胚の通常孵 卵において 6 日目から 21 日目まで低酸素下(15% O2)で孵卵された胚は、通常の 酸素濃度下(21% O2)で孵卵された胚に比べ胚重量が軽くなることを報告してい る。本実験で認められた発生の遅延や胚のサイズが小さくなるといった現象は、 人工容器での培養開始時期に関係なく空気層が排除された場合により顕著であ った。 これらのことから、人工容器における培養では酸素を含む空気の供給が十分 に行われずその結果培養後期における生存率の低下や胚のサイズが小さくなる といった現象が認められたのではないかと考えられることから、人工容器のガ ス調整機能が不十分である可能性が示された。 本実験において、同じ孵卵日数で死亡した胚の発生段階を比較した結果、人 工容器で培養した胚は通常孵卵した胚に比べ発生が遅れていた。また、人工容 器培養において、胚盤葉期から培養した胚は孵卵 60 時間後から培養した胚に比
26 べ発生が遅れていた。Kawashima et al. (2005)は、人工容器を用いてニワトリ胚と ウズラ胚を胚盤葉期からそれぞれ 72 時間と 55 時間培養し、人工容器で培養し た胚の発生段階が遅れることを報告している。また、藤田ら(2007b)は、ニワト リ胚を孵卵 72 時間後から人工容器で培養した場合、通常孵卵した胚に比べ発生 段階が遅れることを報告している。これらのことから、人工容器を用いて培養 すると胚の発生段階が遅れること、さらに、人工容器での培養開始時期が早い ほど遅れるものと考えられた。しかしながら、本実験では死亡胚を対象として いるため、発生段階の結果のみから人工容器が発生遅延の原因であると結論付 けることはできない。 本実験では、Kamihira et al. (1998)の培養法を参考にして胚発生に必要なミネ ラルを水様性卵白に添加した乳酸カルシウムで補っている。しかし、Stage 45 で 死亡した胚の胚重量と第 3 趾長の値は、人工容器で培養した胚が通常孵卵した 胚に比べ低かった。また、胚盤葉期から人工容器で培養した胚は、孵卵 60 時間 後から人工容器で培養した胚に比べてサイズが小さかった。 鳥類の胚発生において、胚発生に必要なカルシウムの約 80%を卵殻から吸収 している(Johnston and Comar, 1955; 藤田ら 2007a)ことから、卵殻は重要なミネ ラル供給源といえる。ニホンウズラ胚のミネラル含有量や胚重量、第 3 趾長の 値が孵卵 10 日目から 15 日目にかけて急激に増加すること(Ono and Wakasugi, 1984)や、ニホンウズラにおいて第 3 趾長の骨化が孵卵 9 日目以降であること (Nakane and Tsudzuki, 1999)が報告されている。このことから、ニホンウズラにお いては孵卵 9 日目以降が骨形成を伴う胚の急激な成長によりミネラルの要求量 が高くなる時期であると考えられる。Ono and Wakasugi(1984)は、孵卵 60 時間後 からウズラ胚をニワトリ卵殻や人工容器で培養し、孵化直前の胚が通常孵卵し た胚に比べ第 3 趾長や胚重量の値が低くなることを報告している。Kamihira et al. (1998)は、孵卵 48 時間後からウズラ胚を人工容器で培養し、培養液に乳酸カル
27 シウムや卵殻粉末を添加することで孵化個体を得られること、培養液に乳酸カ ルシウムや卵殻粉末を添加したにもかかわらず 17 日目に死亡した胚の重量が、 人工容器で培養した胚は通常孵卵胚よりも軽いことを報告している。このこと から、人工容器培養においては、骨形成を伴う胚の急成長によりミネラルの要 求量が高くなる時期に、培養液に添加したミネラルを十分に利用できていない と推測できる。 本実験において、空気層の有無に関係なく胚盤葉期から人工容器で培養した 胚にのみ奇形が認められた。 韮澤ら(1992)は、ニホンウズラ胚を胚盤葉期からニワトリ卵殻を用いて培養 し、 空気層を設けた場合に奇形が 24%認められたのに対し、代理卵殻中を卵白 で満たし空気層を設けなかった場合に奇形が認められなかったことを報告して いる。また、Kawashima et al.(2002) は、ニワトリ胚の代理卵殻を用いた System III の培養において、発生初期の胚は羊膜が形成されておらず、そのため羊膜で保 護されていない胚が露出し、空気に触れることで発生に悪影響が出ると考察し ている。羊膜の形成は、ニワトリ(Kawashima et al., 2002)で 65~69 時間、ニホン ウズラ(Zaccchci, 1961)で 55~63 時間後に完了するとされている。 奇形を含む先天異常は、発生の原因として、遺伝子・染色体の異常による影 響、胎児期における環境の影響が挙げられる。また、眼の奇形には、片方もし くは両方の眼が形成されないもの(無眼球症: anophthalmia)や小さいもの(小眼球 症: microphthalmia)が挙げられる。眼の形成不全に関する遺伝子・染色体の原因 として、Sox2、Pax6、Otx2 そして Rax などを含む遺伝子群が同定されている。 また、環境による原因として、低酸素環境下での孵卵が眼の形成不全をはじ め と す る 奇 形 の 発 生 を 引 き 起 こ す こ と が 報 告 さ れ て い る (Naujoks, 1953; Grabowski and Paar, 1958)。
28 や染色体の変異が原因ではないと考えられるとともに、空気層の有無にかかわ らず奇形が発生したことから、発生初期の胚が PTFE 膜や空気層と接触している 培養環境による影響が考えられた。 本実験で発生した奇形の約 67%が眼に関するものであり、この結果は胚盤葉 期から代理卵殻を用いて培養し、発生した奇形の大部分(82%, 18/22; 88%, 22/25) が眼に関するものであるという韮澤ら(1992)の報告と一致していた。眼の形成は 培養 6 日頃までにはある程度終了することから(韮澤ら, 1992)、発生初期におけ る人工容器での環境(空気層や PTFE 膜)が胚に何らかのダメージを与え、その結 果、眼の奇形が発生したのではないかと考えられた。 以上の結果から、胚盤葉期から人工容器を用いて培養する際に、容器が胚の 死亡率増加や奇形発生率の増加に影響を与えていると考えられた。このため、 培養胚の死亡率や奇形発生率を低下させるためには、培養初期の培養環境の改 善をする必要があると考えられた。
29 5. 小括 本実験では、ポリテトラフルオロエチレン(PTFE)製の膜とポリプロピレ ン製のコニカルチューブを用いて人工容器を作製し、ニホンウズラ胚を培養 した。 ニホンウズラ胚は、胚盤葉期または孵卵 60 時間後から人工容器に移 し、空気層の設置区・未設置区を設けた後 PTFE 膜とポリ塩化ビニリデン製 のサランラップで封をした。 また、人工容器に移す際には乳酸カルシウム を 35mg 添加した。人工容器を用いて培養をした結果、胚盤葉期から培養し た胚は空気層の有無にかかわらず孵化直前である Stage 45 まで徐々に生存 率を下げていった。 孵卵 60 時間後から培養した胚は高い生存率を維持して いたが、9 日目以降に空気層の未設置区では生存率が下がったのに対し設置 区では高い生存率を維持していた。 空気層を設置した区では、人工容器で の培養開始時期に関係なく孵化直前である Stage 45 まで到達し羊膜を破る 胚が見られた。空気層の未設置区では、人工容器での培養開始時期に関係な く全ての胚が Stage 45 に到達する前に死亡した。人工容器で培養し、死亡し た胚を同じ培養日数または同じ発生段階で通常孵卵した胚と比較した結果、 人工容器で培養した胚には、発生の遅れや体重、第三趾長が小さい事が分か った。 また、胚盤葉期から人工容器で培養し、死亡した胚を同じ培養日数 または同じ発生段階で孵卵 60 時間後から培養した胚を比較した結果、 発生 の遅れや体重、第三趾長が小さい事が分かった。 また、胚盤葉期から人工 容器で培養した胚にのみ眼に関する奇形が見られた。 そして、このことか ら培養初期の胚へのダメージが奇形発生と培養初期の死亡率に関係がある と推察された。
30
第3 章 人工容器での培養開始時期が胚の外部計測値とカルシウムおよびマ グネシウムの含有量に与える影響
1. 緒言
これまで、卵殻の役割を人工容器に代替させた培養法が検討されてきたが孵 化個体は得られていない(Ono and Wakasugi, 1983; 1984)。これらの研究で用 いられた人工容器は、培養容器としての役割は果たしているものの、ミネラル 供給源としての役割が果たせていなかったものと考えられる。 Kamihira et al. (1998)は、ニホンウズラ胚を孵卵 48 時間後から人工容器で培 養する際に、卵殻の主成分である炭酸カルシウムの代替として、乳酸カルシウ ムや卵殻粉末を卵白に添加することで 43%の孵化率を得ている。Kamihira et al. (1998)の報告以前も、ニホンウズラにおいて前孵卵後から人工容器を用いて 培養を行った報告はあるが孵化個体は得られておらず、また、ミネラルの添加 も行われていない。このため、人工容器培養においては、乳酸カルシウムや卵 殻粉末等のミネラルを添加することが培養胚を孵化させるために必要であると 考えられた。その一方で、ミネラルを添加しても通常孵卵した胚に比べ体が小 さいことがウズラ(乳酸カルシウムと卵殻粉末を添加: Kamihira et al., 1998)や ニワトリ(炭酸カルシウムを添加:藤田ら 2007b)で報告されている。また、第 2 章の結果から、人工容器で培養した胚と通常孵卵した胚の大きさを同じ発生段 階(Stage 45)で死亡した胚において比較し、乳酸カルシウムを添加したにもかか わらず人工容器で培養した胚は通常孵卵した胚よりも小さいことや胚盤葉期か ら人工容器で培養した胚は孵卵60 時間後から培養した胚よりも小さくなること が明らかとなり、添加した乳酸カルシウムが吸収されていない可能性が示唆さ れた。 そこで本実験では、培養液に乳酸カルシウムまたは乳酸カルシウムと卵殻粉
31 末を添加し、人工容器での培養開始時期が胚の成長やカルシウム(Ca)およびマ グネシウム(Mg)含有量にどのような影響を与えるのかを明らかにしようとした。 2. 材料および方法 (1) 供試卵 ニホンウズラの受精卵は、株式会社モトキ(Saitama, Japan)より購入したも のを実験に供試した。 (2) 培養容器 本実験で使用した培養容器を図3-1(B)に示した。50 ml ポリプロピレン製チュ ーブ(BD Falcon Conical Tube; Nihon BD Co, Tokyo, Japan. 以降コニカルチュ ーブと表記.)に培養器内の乾燥を防ぐため滅菌蒸留水を 35 ml 入れた後、多孔性 でガス透過性のあるポリテトラフルオロエチレン(PTFE)製の膜(Milliwrap; Nihon Millipore Co, Tokyo, Japan. 以降ミリラップと表記.)を袋状に取り付け たものを培養容器とした。
上蓋としてポリ塩化ビニリデン製フィルム(Saran Wrap; Asahi Kasei Life & Living Co, Tokyo, Japan. 以降サランラップと表記.)を用いた。なお、第 2 章で は上蓋としてミリラップの上にサランラップを重ねたものを用いたが、予備実 験の結果サランラップのみの場合と生存率に差が見られなかったことから視認 性向上のため上蓋をサランラップのみとした。空気層は容器の開口部から 5~7 ㎜設けた。培養容器の材料はオートクレーブにより滅菌処理(121℃、20 分)を施 した。 (3) 胚培養 人工容器による培養開始時期は、胚盤葉期(放卵直後)または孵卵 60 時間後と した。培養液はKamihira et al. (1998)を参考に、乳酸カルシウム(Sigma-Aldrich Co, Tokyo, Japan)35 mg または乳酸カルシウム 25 mg+ウズラの卵殻粉末 10
32
mg をニワトリの水様性卵白に添加したものを使用した。また、対照群として操 作を加えず通常孵卵した胚を用いた。なお、ウズラの卵殻粉末は、卵殻膜を除 去し乾熱処理(121℃、30 分)を施した後、乳鉢ですりつぶしたものを使用した。 胚は、孵卵器(Type P-008; Showa Furanki Co, Tokyo, Japan) を用いて 38.0℃、 湿度60%、転卵 60°/時間の条件下で培養した。また、 培養開始 60、 120、240 および360 時間後に生存しておりかつ奇形のない胚を回収しサンプルとした。 (4) 胚の外部計測 採取した胚は、卵黄を除去したのち発生段階、全長、第 3 趾長、眼直径、嘴 峯長および 乾重量 を測定し た。胚 の発生 段階(Stage)は、Hamburger and Hamilton(1951)の発生段階表を参考にして決定した。 (5) 胚のカルシウムおよびマグネシウム含有量の測定法 供試した胚は、凍結乾燥機にて重量変化がなくなるまで乾燥したのち有機物 を分解する湿式灰化を施した。すなわち、胚を灰化用試験管に移した後、濃硝 酸を4 ml 入れ、ドライバスで内容物が乾固するまで加熱(100℃、24 時間)した。 冷却後、同じ灰化用試験管に過酸化水素水を2 ml 入れ再び内容物をドライバス で加熱し乾固させた。湿式灰化後、0.8 M の塩酸を用いて分析部位ごとに適した 希釈率にサンプルを希釈調整した。その後サンプルの全量に対し、10%量の 1% 塩化ランタンを加え夾雑物を除く処理を施し、原子吸光光度計(AA-640-13; Shimadzu Co, Tokyo, Japan)により胚の Ca および Mg 量を測定した。
(6) 統計処理
外部計測値、発生段階、Ca と Mg の含有量および濃度は、同一時間内または 同一実験区内でOne-way ANOVA 後、Scheffe’s multiple comparison test を用 いて統計処理を行い、p < 0.05 を統計的に有意とした。
33
図 3-1. 培養に用いた人工容器
A:第 2 章で使用した培養容器、B:本実験で使用した培養容器 Fig. 3-1. Quail embryo culture vessels
A: the second chapter, B: this experiment
サランラップ Saran Wrap 空気層 Air space ミリラップ Milliwrap 添加ミネラル Supplemental minerals コニカルチューブ Conical tube 滅菌蒸留水 Distilled water 空気層 Air space
A
サランラップ Saran Wrap ミリラップ Milliwrap 乳酸カルシウム Calcium lactate 滅菌蒸留水 Distilled water コニカルチューブ Conical tube 卵黄 Yolk 水様性卵白 Thin albumen 空気層 Air space ミリラップ Milliwrap 空気層 Air spaceB
34 3. 結果 (1) 発生段階 胚の発生段階を図 3-2(A)に示した。同一培養実験区内ではいずれの区分にお いても 60 から 120 時間、120 から 240 時間および 240 から 360 時間にかけて 有意な成長が認められた(p < 0.05)。同一培養時間内では 60 時間と 120 時間に おいて各実験区間に差は認められなかったが、240 時間と 360 時間では、通常 孵卵した胚に比べ人工容器で培養した胚は発生段階が遅れていた(p < 0.05)。ま た、240 時間と 360 時間において孵卵 60 時間後から培養した胚に比べ、胚盤葉 期から培養した胚は発生段階が遅れていた (p < 0.05)。一方で、乳酸カルシウム や卵殻粉末の添加による実験区間での差は認められなかった。 (2) 乾重量 胚の乾燥重量を図 3-2(B)に示した。同一培養実験区内ではいずれの区分にお いても 60 から 120 時間にかけて増加は認められなかった。しかし、120 から 240 時間および 240 から 360 時間にかけては有意な増加が認められた(p < 0.05)。 同一培養時間内では60 時間と 120 時間において各実験区間に差は認められなか ったが、 240 時間と 360 時間では、通常孵卵した胚に比べ人工容器で培養した 胚は有意に軽かった(p < 0.05)。また、240 時間および 360 時間においては孵卵 60 時間後から培養した胚に比べ、胚盤葉期から培養した胚は有意に軽かった(p < 0.05)。一方で、乳酸カルシウムや卵殻粉末の添加による実験区間での差は認 められなかった。 (3) 全長 胚の全長を図 3-2(C)に示した。同一培養実験区内ではいずれの区分において も 60 から 120 時間、120 から 240 時間および 240 から 360 時間にかけて有意 な増加が認められた(p < 0.05)。同一培養時間内では 60 時間と 120 時間におい
35 て各実験区間に差は認められなかったが、240 時間と 360 時間では、通常孵卵 した胚に比べ人工容器で培養した胚は有意に短かった(p < 0.05)。また、240 時 間と360 時間においては孵卵 60 時間後から培養した胚に比べ、胚盤葉期から培 養した胚は有意に短かった (p < 0.05)。一方で、乳酸カルシウムや卵殻粉末の添 加による実験区間での差は認められなかった。 (4) 第 3 趾長 胚の第3 趾長を図 3-2(D)に示した。第 3 趾長については 60 時間での計測が出 来なかったため 120 時間以降の測定とした。同一培養実験区内ではいずれの区 分においても120 から 240 時間および 240 から 360 時間にかけて有意な増加が 認められた(p < 0.05)。同一培養時間内では 120 時間において各実験区間に差は 認められなかったが、240 時間と 360 時間では、通常孵卵した胚に比べ人工容 器で培養した胚は有意に短かった (p < 0.05)。また、全ての時間において人工容 器での培養開始時期による差は認められなかった。一方で、乳酸カルシウムや 卵殻粉末の添加による実験区間での差は認められなかった。 (5) 嘴峯長 胚の嘴峯長を図3-2(E)に示した。嘴峯長については 60 時間での計測が出来な かったため 120 時間以降の測定とした。同一培養実験区内ではいずれの区分に おいても120 から 240 時間および 240 から 360 時間にかけて有意な増加が認め られた(p < 0.05)。同一培養時間内では 120 時間において各実験区間に差は認め られなかったが、240 時間と 360 時間において通常孵卵した胚に比べ人工容器 で培養した胚は有意に短かった(p < 0.05)。また、全ての時間において人工容器 での培養開始時期による差は認められなかった。一方で、乳酸カルシウムや卵 殻粉末の添加による実験区間での差は認められなかった。
36 (6) 眼直径 胚の眼直径を図 3-2(F)に示した。同一培養実験区内ではいずれの区分におい ても 60 から 120 時間、120 から 240 時間および 240 から 360 時間にかけて有 意な増加が認められた(p < 0.05)。同一培養時間内では 60 時間と 120 時間にお いて各実験区間に差は認められなかったが、240 時間と 360 時間では、通常孵 卵した胚や孵卵60 時間後から人工容器で培養した胚に比べ胚盤葉期から人工容 器で培養した胚は有意に小さかった(p < 0.05)。一方で、乳酸カルシウムや卵殻 粉末の添加による実験区間での差は認められなかった。 (7) カルシウム濃度・含有量 胚のカルシウム濃度を図 3-3(A)に示した。同一培養実験区内ではいずれの区 分においても60 から 120 時間にかけて増加は認められなかった。しかし、120 から240 時間および 240 から 360 時間にかけて有意な濃度の上昇が認められた (p < 0.05)。同一培養時間内では 60、120 および 240 時間において各実験区間に 差は認められなかったが、360 時間では通常孵卵した胚に比べ人工容器で培養し た胚は有意に低かった(p < 0.05)。また、全ての時間において人工容器での培養 開始時期による濃度の差は認められなかった。一方で、乳酸カルシウムや卵殻 粉末の添加による実験区間での差は認められなかった。 胚のカルシウム含有量を図 3-3(B)に示した。同一培養実験区内ではいずれの区 分においても60 から 120 時間にかけて増加は認められなかった。しかし、120 から240 時間および 240 から 360 時間にかけて含有量の上昇が認められた(p < 0.05)。同一培養時間内では 60、120 および 240 時間において各実験区間に差は 認められなかったが、360 時間では通常孵卵した胚に比べ人工容器で培養した胚 は有意に低かった(p < 0.05)。また、全ての時間において人工容器での培養開始 時期による差は認められなかった。一方で、乳酸カルシウムや卵殻粉末の添加
37 による実験区間での差は認められなかった。 (8) マグネシウム濃度・含有量 胚のマグネシウム濃度を図 3-3(C)に示した。同一培養実験区内ではいずれの 区分においても 60 から 120 時間および 240 から 360 時間にかけて濃度の減少 は認められなかったが、120 から 240 時間にかけて有意な濃度の減少が認めら れた(p < 0.05)。同一培養時間内では全ての時間において各実験区間に差は認め られなかった。一方で、乳酸カルシウムや卵殻粉末の添加による実験区間での 差は認められなかった。 胚のマグネシウム含有量を図 3-3(D)に示した。同一培養実験区内ではいずれ の区分においても60 から 120 時間にかけて増加は認められなかったが、120 か ら240 時間および 240 から 360 時間にかけて有意な増加が認められた(p < 0.05)。 同一培養時間内では60 時間と 120 時間において各実験区間に差は認められなか ったが、240 時間と 360 時間では、通常孵卵した胚に比べ人工容器で培養した 胚は有意に含有量が低かった(p < 0.05)。また、240 時間において孵卵 60 時間後 から培養した胚に比べ、胚盤葉期から培養した胚は有意に低かった(p < 0.05)。 しかし、120 時間と 360 時間においては人工容器での培養開始時期による差は 認められなかった。一方で、乳酸カルシウムや卵殻粉末の添加による実験区間 での差は認められなかった。
38 図 3-2. 培養したウズラ胚の外部計測値 A: 発生段階、B: 乾燥重量、C: 全長、D: 第 3 趾長、E:嘴峯長お よび F: 眼直径 ◆:胚盤葉期、乳酸カルシウム 35 mg(n = 5); ■:胚盤葉期、乳酸カルシウム 25 mg+卵殻粉末 10 mg(n = 5); ◇:孵卵 60 時間後、乳酸カルシウム 35 mg(n = 5); □:孵卵 60 時間後、乳酸カルシウム 25 mg+卵殻粉末 10 mg(n = 5); ▲: 通常孵卵した胚(n = 5)。各点およびバーは平均値±標準誤差。 同一時間内において異文字間(a-d)に有意差あり(p < 0.05)。 *:同一実験区の前測定値に対して有意差あり(p < 0.05)。 Ey e diamete r (mm) 10 8 6 0 4 2 0 2 4 6 8 10 60 120 240 360 b * a * a a a a * a * a* a* a * a * a * a * a * a * b * b * b * 60 120 240 360
Time of incubation (hours)
0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 2 4 6 8 10 60 120 240 360 10 8 6 0 Beak le ngth (mm) 4 2 b * a * a a a a a a * b * b * b * b * *b b * *b 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 4 8 12 16 60 120 240 360 3r d toe le ngth (mm ) 16 12 8 0 4 b * a * a a a a a a * a*a* a * a * a * a * b * 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 80 60 40 0 T otal le ngth (mm) 20 0 20 40 60 80 60 120 240 360 b * a a a a a a a a c * c* b * b* a* a * b * c * c * 60 120 240 360
Time of incubation (hours)
0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 45 30 0 Deve lopme ntal stage 15 0 5 10 15 20 25 30 35 40 45 60 120 240 360 b * a * a a a a * a*a**a a * a * b * b * b * c * c * c * c * 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 Dry w ei ght (g) 1.2 0.8 0.4 00.0 0.4 0.8 1.2 60 120 240 360 b * a a * c * a a a a a a a a * b * b * b * c * c * c * 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 A B C D E F 孵卵時間 (時間) Incubation period (hour) 孵卵時間 (時間)
Incubation period (hour)
発生段階 D ev el o p me n ta l st ag e 乾 重量 (g ) D ry w ei g h t (g ) 全長 (mm ) T o ta l le n g th ( mm ) 眼直径 (mm ) Ey e d ia me te r (mm ) 嘴峯長 (mm ) B ea k l en g th t ( mm ) 第 3 趾長 (mm ) 3 rd t o e le n g th ( mm )
39
Fig. 3-2. Quail embryonic external measurement
A: developmental stage, B: Dry weight, C: Total length, D: The 3rd toe length, E: Beak length and F: Eye diameter. In vitro culture was set up from the blastoderm stage and additional mineral was ◆:Calcium lactate 35 mg (n = 5) or ■:Calcium lactate 25 mg+eggshell powder 10 mg (n = 5). In
vitro culture was set up after 60 hours preincubation and additional mineral
was ◇:Calcium lactate 35 mg (n = 5) or □:Calcium lactate 25 mg+ eggshell powder 10 mg (n = 5). Intact fertilized eggs as ▲: controls (n = 5). Each points and vertical bars represent the mean and standard error of mean values, respectively. Different letters (a-d) represent significant differences (p < 0.05) as assessed by Scheffé’s test. *:Significant differences (p < 0.05) in same experimental group.
40 図 3-3. 培養したウズラ胚のミネラル濃度および含有量 A: カルシウム濃度、B: カルシウム含有量、C: マグネシウム濃度およ び D:マグネシウム含有量 ◆:胚盤葉期、乳酸カルシウム 35 mg(n = 5); ■:胚盤葉期、乳酸カルシウム 25 mg+卵殻粉末 10 mg(n = 5); ◇: 孵卵 60 時間後、乳酸カルシウム 35 mg(n = 5); □:孵卵 60 時間後、 乳酸カルシウム 25 mg+卵殻粉末 10 mg(n = 5); ▲:通常孵卵した胚(n = 5)。各点およびバーは平均値±標準誤差。同一時間内において異文字 間(a-d)に有意差あり(p < 0.05)。 *:同一実験区の前測定値に対して 有意差あり(p < 0.05)。
Fig. 3-3. Quail embryonic mineral concentration and content
A: Calcium concentration, B: Calcium content, C: Magnesium concentration and D: Magnesium content. In vitro culture was set up from the blastoderm stage and additional mineral was ◆:Calcium lactate 35 mg (n = 5) or ■: Calcium lactate 25 mg+eggshell powder 10 mg (n = 5). In vitro culture was
1.2 0.8 0.4 0 M g co nte nt (m g) 0.0 0.4 0.8 1.2 60 120 240 360 b * a a a a a a a a a * b * c * c * a * b * b * c **c 60 120 240 360 Time of incubation (hours)
0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 2.5 2.0 1.5 1.0 0.5 0 Mg conce ntration 0.0 0.5 1.0 1.5 2.0 2.5 60 120 240 360 (mg/g Dry Embryo) a * a a a a a a a a a *a*a * a* a aa a a 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 C D 0 5 10 15 20 60 120 240 360 Ca conte nt (mg) 20 15 10 5 0 b * a a* c * a a a a a a a a*a* a*a* b * c * a * 60 120 240 360 Time of incubation (hours)
0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 60 120 240 360 20 15 10 5 0 Ca conc entr at ion (mg/g Dry Embryo) a * a a a a a a a a a *a* a*a* b * b * a * c * c * 0 5 10 15 20 25 30 35 40 45 60 120 240 360 0 5 10 15 20 25 30 35 40 45 60 120 240 360 A B 孵卵時間 (時間) Incubation period (hour)
(mg/g 乾燥重量) (mg/g Dry Embryo) (mg/g 乾燥重量)
(mg/g Dry Embryo)
孵卵時間 (時間) Incubation period (hour)
Ca 濃度 C a co n ce n tr at io n M g 濃度 M g c o n ce n tr at io n Ca 含有量 C a co n te n t (mg ) M g 含有量 M g c o n te n t (mg )
41
set up after 60 hours preincubation and additional mineral was ◇:Calcium lactate 35 mg (n = 5) or □:Calcium lactate 25 mg+eggshell powder 10 mg (n = 5). Intact fertilized eggs as ▲: controls (n = 5). Each points and vertical bars represent the mean and standard error of mean values, respectively. Different letters (a-d) represent significant differences (p < 0.05) as assessed by Scheffé’s test. *:Significant differences (p < 0.05) in same experimental group.
42
4. 考察
Kamihira et al. (1998)はウズラ胚を、藤田ら(2007b)はニワトリ胚を、 Perry(1988)の代理卵殻法における System III に相当する段階から人工容器で 培養し、乳酸カルシウムや炭酸カルシウムを添加した場合でも通常孵卵した胚 に比べて胚の重量が軽いことを報告している。本実験では乳酸カルシウムと卵 殻粉末の添加を行い、時間毎に培養胚の外部計測を行った。その結果(図 2A~ F)、人工容器での培養開始時期に関係なく、発生開始 120 から 360 時間にかけ て胚の急激な成長が認められた。この胚の急激な成長は、通常孵卵した胚、人 工容器や代理卵殻で培養されたウズラ胚(Ono and Wakasugi, 1984)における体 重や第 3 趾長の急激な成長と同様の推移であった。一方、眼直径を除く全ての 計測部位において、120 時間までは人工容器培養胚と通常孵卵した胚の間に差は 認められなかったが、発生開始 240 時間以降では人工容器培養胚は通常孵卵し た胚よりも小さく発生が遅れていた。また、胚盤葉期から人工容器で培養した 胚は、発生開始 240 時間以降に乾燥重量、全長、眼直径および発生段階の値が 孵卵60 時間後から培養した胚よりも有意に低い値を示した。 Ruijtenbeek et al. (2000)は、通常孵卵において孵卵 6 日目から 21 日目まで 低酸素(15%)の気相条件下で培養した胚は 21 日目の体重および内臓重量が低い ことを報告している。本実験では、胚の上面に空気層を設けており、蓋として 用いているサランラップは通気性を有している。また、胚を収納しているミリ ラップは通気性を有していることから下面の空気層からもガス交換をすること ができる。しかし、培養容器として穴のない人工容器を用いたことから、ミリ ラップを通じてガス交換を行っても培養用容器中の二酸化炭素濃度の上昇また は、酸素濃度の低下が抑えられない可能性が考えられた。一方で、代理卵殻を 用いた培養においても通常孵卵した胚に比べ培養胚は重量が軽いことが報告さ れており(Ono and Wakasugi,1984; Kamihira et al., 1998; 藤田ら,2007c) 、酸