1)東海大学海洋研究所 〒424−8610 静岡市清水区折戸3−20−1
Institute of Oceanic Research and Development, Tokai University, 3−20−1 Orido, Shimizu-ku, Shizuoka 424−8610, Japan
(2019年8月31受付/2019年10月5日受理)
ナマズの行動と地震との関係に関する研究
野田 洋一
1)Relationship between the behavior of catfish and earthquakes.
Yoichi Noda
Abstract
There have been many reports of abnormal animal behavior that preceded some sizable earthquakes. However, scientific researches have not progressed. Therefore, it is needed for researches on the relationship between physical and chemical changes that precede earthquakes and the sensory organs and instincts of animals have been pointed out. However, there are not enough researches, which based on comprehensive zoological aspects such as animal behavior, ecology, physiology, psychology and so on. Taking into this background, we started research focusing on catfish, which has been researched by some scientists. In this study, we developed a method to quantify catfish behavior. Catfish behavior was classified as a behavior pattern composed of four elements: swimming trajectory, swimming time, swimming distance, and average swimming speed. As a result, we succeeded in comparing changes between detailed behavioral information and physical and chemical factors.
1. 緒 言 地震に先行する動物異常行動の報告は古くから 数多く残されているものの実在性を問う研究は進 んでいない.それゆえ旧来から地震に先行する物 理・化学的な変化と動物の感覚器官や本能などと の関係に関する研究の必要性が指摘されている (例えば力武,1979).しかし,動物行動学,生態 学,生理学,心理学などの総合的な動物学的見地 を踏まえた十分な研究は著者の知る限りこれまで 存在しない.このような背景から,比較的研究が 進んでいるナマズに着目して研究を開始した(野 田ほか,2002).特に動物の行動を定量化する手法 の開発は,動物学的見地において必要な過程のひ とつとして考えられる.本稿では,ビデオ観察に よるナマズの行動を定量化する手法を開発すると ともに,定量化された行動パターンと外的刺激と なる可能性がある雷活動との比較結果を報告する. なお,本研究は千葉大学海洋バイオシステム研究 センター「共同研究」において進められた成果を 引き継ぐものである(長尾,2006,2007,2008). 2. 研究方法 千葉大学海洋バイオシステム研究センターの水
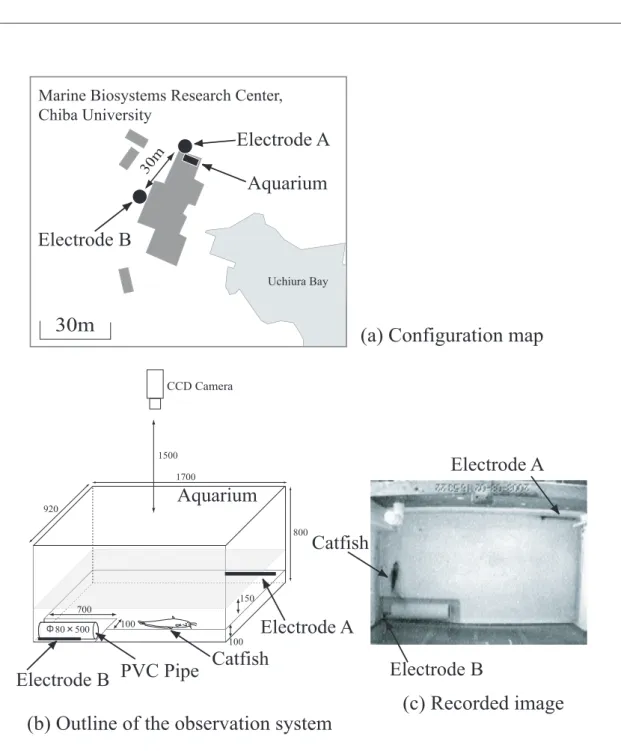
族館内の展示用水槽にナマズを飼育して,暗視ビ デオカメラを用いての24 時間連続記録を行う観 察システムを構築した.観察システムは,東京都 水産試験場において行われたナマズと地震の関係 に関する研究(東京都防災会議地震部会, 1980-1992;野田ほか,2004)を,画像解析と行動分類 の方法に関しては,野田ほか(2002)と若井(2004) を参考にした. 2.1 観察システム 観察システムの構成は次のとおりである. 1)観察場所 千葉県鴨川市内浦,千葉大学海洋バイオシステ ム研究センター内 水族館(35.1208N,140.1821E) 2)供試魚 埼玉県水産流通センターから養殖ナマズ(学 名: ),体長約30 cm を購入して供 試魚とした.餌は底棲肉食魚用のペレット状の飼 料「ひかりクレストキャット」(キョーリンフード 工業株式会社)を週2 回程度与えた. 3)観察システム 水族館の展示用水槽(170 cm 92 cm 80 cm)を 利用して観察水槽を作製した.天井部にCCD カメ ラ(Victor TK-C1360)を設置して,PC(Win2000, DELL PowerEdge 200sc)を用いて画像ファイル (JPEG 形式)を毎秒で保存するシステムを構築し た. システムの時計はNTP 同期により毎正時に 自動補正を行った.画像処理の過程で二値化処理 を施すために,水槽底面を白色として,黒色のナ マズと区別を可能とした.ナマズは暗くて狭い場 所を好むため,水槽の一部に高低差10 cm の低位 部を設けて塩ビ管を設置した.さらに地電流変化 を水槽内に導くために,建物外部に埋設した2 本 の電極と水槽内に設置した電極とを接続させた. Fig. 1 に観察システムを示す. 主な改良点の変遷は次のとおりである. 2005 年 9 月:暗視カメラを使用した毎秒 24 時間 連続撮影システムを構築した. 2007 年 9 月:地電流を水槽内に導くための電極を 屋外と水槽内に設置して両者を接続させた. 2008 年 7 月:水槽低位部に塩ビ管を設置して,行 動していない場合は塩ビ管内に極力停滞するよ うに改良した. 2.2 画像解析 1)対象期間 水槽低位部に塩ビ管を設置してから観察終了ま での期間とした. 2008 年 7 月 28 日∼ 2009 年 1 月 13 日(170 日間) 2)対象行動 低位部の塩ビ管から高位部へ遊泳して再び塩ビ 管へ戻る行動を一行動とした. 3)画像処理 Linux サーバにおいてフリーウエアである Im-ageMagick を利用してナマズの輪郭の抽出を行 い,その一行動の複数画像を合成して一枚の軌跡 図を作成した.さらに,OpenCV(こちらもフリー ウエア)を利用して重心座標を数値データとして 求めて,軌跡の描画,遊泳時間,遊泳距離,平均 遊泳速度を求めた. 3. 結 果 3.1 欠測について 全期間内において,3 ∼ 7 秒程度の欠測が毎分 2 回発生するなどして欠測が多く確認された.欠測 の原因のひとつとして,パソコンのスペックに対 してビデオキャプチャーの負荷が大きかったこと と,ISDN 回線において WEB サーバへの画像アッ プロードの負荷が大きかったことによると考えて いる. 欠測回数は全期間内で347,505 回生じた.周期 的な欠測の例をFig. 2 に示す.欠測の継続時間ご との頻度についてTable 1 に示す.3600 秒の欠測 が1 回発生しているが,これは 2008 年 10 月 19 日の8 時台の 1 時間分である.原因は PC の HDD への記録の障害だったと考えられる. 3.2 行動分類 1)行動分類表の作成 記録したビデオ画像を画像解析することにより ナマズの行動パターンを分類・記号化した.この ことにより物理・化学的要因との比較,平常や異 常という行動の定義に関する研究へ進めることが 可能となる.まず,行動を記号化するために各種 行動要素の抽出を行う.行動要素としては,遊泳 軌跡,遊泳時間,停滞時間,遊泳距離,最大遊泳 速度などの様々な要素が考えられるが,本研究で
Fig. 1. Observation system 00:00:00 00:01:00 00:02:00 00:03:00 00:04:00 00:05:00 (Time) Fl ag o f m iss in g re co rd in g Aug. 11, 2008 < Fig. 2 Noda > Fig. 2. An example of periodic missing
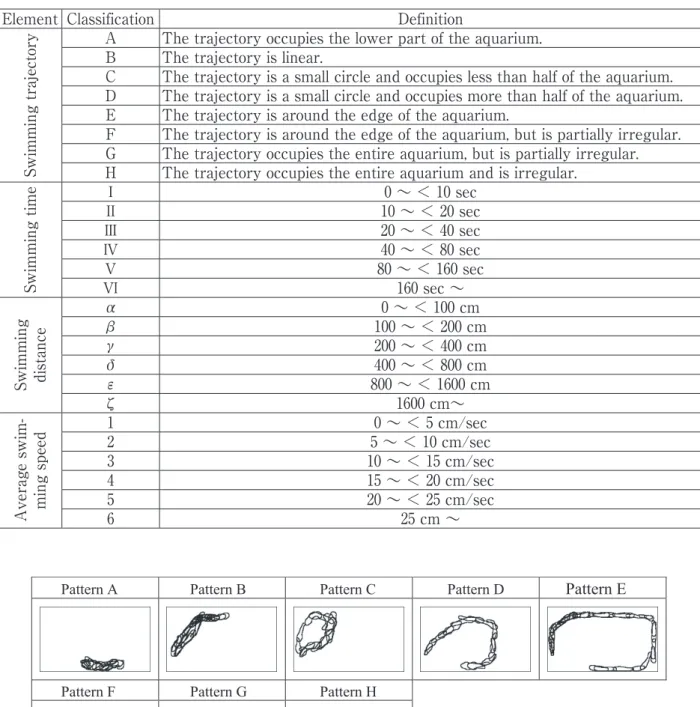
は,欠測が多いため停滞時間と最大遊泳速度は除 外し,遊泳軌跡,遊泳時間,遊泳距離,平均遊泳 速度の4 つとした.平均遊泳速度は遊泳距離と遊 泳時間から求めた.遊泳軌跡は画像処理によりナ マズの輪郭を抽出して合成した軌跡図とナマズの 重心座標値を求めて描画した軌跡図から目視によ り分類した.この4 つの要素で構成される行動を 行動パターン と定義する.行動分類の定義を Table 2 に示す.遊泳軌跡の分類例を Fig. 3 に示 す. 2)行動分類の結果 画像処理の結果,2008 年 7 月 28 日∼ 2009 年 1 月13 日(170 日間)において,2,151 行動が抽出さ れた.一日ごとの行動回数をFig. 4 に示す.なお, 欠測部分の行動は不明であるために分類作業には 少なからず支障を来した.安定した観察システム と高い精度の画像解析ならびに自動化は今後の課 題である. 解析期間内の2,151 回の行動を分類した結果, 224 種類の行動パターンに分類された(Table 3). 行動要素ごとの回数をTable 4 に示す. 本来ならば夜行性のナマズは夜間の活動が多い が,本供魚は昼夜問わず活動していたことが認め られた(Fig. 5). 月単位の上位10 つの行動パターンを Table 5 に 示す.月によって行動パターンの順位に変化が認 められる. 2008 年 8 月∼ 12 月までの月単位における各種 要素の割合についてFig. 6 に示す.季節的な傾向 を捉えるにはデータが不足しているが,9 月の遊泳 速度は若干速い傾向が認められることや,12 月は 遊泳軌跡が複雑で遊泳時間が長くなる傾向が認め られる. 全期間内における時間帯別の各行動要素の割合 をFig. 7 に示す.この図から昼夜の行動の傾向が 掴めると考えたが,顕著な違いは認められない. つまり,昼夜問わず,同様で同程度の行動であっ たことを示している. 3.3 雷活動との比較 分類された行動パターンと物理・化学的要因と の同時性を探ることを目的として,落雷とナマズ の行動変化について比較を行った.雷活動は落雷 だけではなく,雷雲の接近にともなう大気電場の 変化,雷鳴や降雨による音や振動などの影響も想 定される.ここでは,落雷前後のナマズの行動変 化の有無について確認を行った. 株式会社フランクリン・ジャパンの全国雷観測 ネットワークにより観測されたデータを使用した. データについては次のとおりである. 中 心 地 点: 千葉大学海洋バイオシステム研究 センター (北緯 35 7′14″/ 東経 140 11′3″) 調 査 範 囲:20 km 20 km 調 査 期 間: 2008 年 07 月 28 日 ∼ 2009 年 01 月 13 日 デ ー タ 元:株式会社フランクリン・ジャパン 落雷発生数:合計106 回(雲放電含む) 落雷地点についてはFig. 8 に示すとおりである. 落雷前後15 分間以内のナマズの行動を抽出し たものをTable 6 に示す.複数の落雷が認められ Missing measure-ment duration (sec) 1 2 3 4 5 6 7 8 conut 414 4 70,939 1,750 22,944 149,240 73,020 5,820 Missing measure-ment duration (sec) 9 10∼<30 30∼<60 60∼<120 120∼<300 300∼< 1800 1800∼<3600 3600 conut 3,725 13,349 4,878 1,361 53 7 − 1 Total count 347,505
Element Classification Definition Sw im m in g tr aj ec to
ry A The trajectory occupies the lower part of the aquarium. B The trajectory is linear.
C The trajectory is a small circle and occupies less than half of the aquarium. D The trajectory is a small circle and occupies more than half of the aquarium. E The trajectory is around the edge of the aquarium.
F The trajectory is around the edge of the aquarium, but is partially irregular. G The trajectory occupies the entire aquarium, but is partially irregular. H The trajectory occupies the entire aquarium and is irregular.
Sw im m in g tim e Ⅰ 0 ∼ < 10 sec Ⅱ 10 ∼ < 20 sec Ⅲ 20 ∼ < 40 sec Ⅳ 40 ∼ < 80 sec Ⅴ 80 ∼ < 160 sec Ⅵ 160 sec ∼ Sw im m in g di st an ce α 0 ∼ < 100 cm β 100 ∼ < 200 cm γ 200 ∼ < 400 cm δ 400 ∼ < 800 cm ε 800 ∼ < 1600 cm ζ 1600 cm∼ A ve ra ge s w im -m in g sp ee d 12 5 ∼ < 10 cm/sec0 ∼ < 5 cm/sec 3 10 ∼ < 15 cm/sec 4 15 ∼ < 20 cm/sec 5 20 ∼ < 25 cm/sec 6 25 cm ∼
Table 2 Behavior classification table
Pattern A Pattern B Pattern C Pattern D Pattern E
Pattern F Pattern G Pattern H
< Fig. 3 Noda >
Fig. 3. Classification of the trajectory pattern (sample)る場合はナマズが行動した時刻に一番近い時刻の 落雷を選出した. 7 月 29 日 13 時 19 分 48 秒の 2 つの落 雷の 2 秒 後にEⅡγ5(頻度順位 20 位)の行動を示した.同 時性という観点においては落雷が行動を誘発した とする仮定が成立する.また8 月 20 日 22 時 12 分 54 秒の落雷(雲放電)46 秒前には DⅡγ5(頻度順 位19 位)の行動を示した.8 月 21 日 19 時 16 分 30 秒,19 時 30 分 27 秒,20 時 20 分 06 秒 の 落 雷 (雲放電含む)に対してはそれぞれ約190 秒後に BⅠβ4(頻度順位 6 位),DⅢδ6(頻度順位 153 位), BⅠβ4(頻度順位 6 位)の行動を示している.8 月 29 日,12 月 5 日,12 月 16 日の 落 雷 に 対 し ては, 落雷時から 約600 秒の幅があるが,GⅡγ4(頻 度順位105 位),HⅤδ2(頻度順位 188 位),HⅣδ3 (頻度順位186 位)の行動が認められた.雷活動は 雷雲の接近に伴う大気電場の変化や激しい降雨の 気象変化なども伴うため,落雷時だけではなく前 後の物理・化学的要因との変化の比較が重要とな る. 4. まとめ 本研究ではナマズの行動を定量化するための手 法を開発した.ナマズの行動を遊泳軌跡,遊泳時 間,遊泳距離,平均遊泳速度の4 つの要素から構 成される行動パターンとして分類した.既往の研 究では,定量化のために振動計,通過センサー, 筋電位センサーなどが使用され頻度値が得られて いたが,行動分析が難しいことが課題として残さ れていた(東京都防災会議地震部会,1980-1992; 野田ほか,2002).本研究において開発した定量化 の手法は,行動分析と物理・化学的要因との比較 をより詳細に行うことを可能にするものである. 次に外的刺激の要因の可能性がある物理・化学的 変化との比較を目的として落雷との比較を行った. この結果,同時性の観点から7 月 29 日 13 時 19 分48 秒の 2 つの落雷の 2 秒後に生じた行動「EⅡ γ5」との関係性が疑われた.今後はさらにデータ を増やした解析を行う必要がある.システムのス ペック上の問題から多くの欠測を生じた.欠測部 分の行動が不明であるために画像解析と分類作業 には少なからず支障を来した.安定した観察シス テムと高い精度の画像解析ならびに自動化につい ては今後の課題である. ナマズに限らず行動分類表は,対象動物の飼育 や生息環境などを考慮して,鳴き声なども含む行 動の特徴を反映する要素を決定することが重要で ある.そして,計測機器の性能を超える動物の検 知・察知能力や本能的な行動,個体や群での場合, 多様な生息環境下における相互作用などを考慮し て,動物学的見地から本研究を進める.
100
80
60
40
20
0
Aug.
Sep.
Oct.
Nov.
Dec.
Jan., 2009
Jun., 2008
N
um
be
r o
f a
ct
iv
iti
es
(c
ou
nt
/d
ay
)
< Fig. 4 Noda >
Rank-ing Behavior pattern Total
num-ber % Rank-ing Behavior
pattern Total
num-ber % Rank-ing
Behavior pattern
Total
num-ber % Rank-ing
Behavior pattern
Total
num-ber % Rank-ing Behavior pattern Total num-ber % 1 AⅡβ3 79 3.67 51 AⅠα4 14 0.65 101 BⅥβ1 7 0.33 151 CⅣδ2 3 0.14 201 BⅤα1 1 0.05 2 EⅢγ3 67 3.11 52 CⅢγ3 14 0.65 102 EⅡδ6 7 0.33 152 CⅥγ1 3 0.14 202 CⅠγ5 1 0.05 3 AⅠα3 59 2.74 53 DⅣγ2 14 0.65 103 EⅢγ2 7 0.33 153 DⅢδ6 3 0.14 203 CⅢβ1 1 0.05 4 BⅡβ3 39 1.81 54 FⅢδ5 14 0.65 104 EⅤγ1 7 0.33 154 DⅤε2 3 0.14 204 CⅢδ5 1 0.05 5 CⅡβ3 38 1.77 55 DⅡγ6 13 0.6 105 GⅡγ4 7 0.33 155 EⅡδ5 3 0.14 205 DⅠγ5 1 0.05 6 BⅠβ4 35 1.63 56 EⅤδ2 13 0.6 106 BⅤγ1 6 0.28 156 EⅢδ6 3 0.14 206 DⅠδ6 1 0.05 7 GⅡδ6 34 1.58 57 FⅤδ2 13 0.6 107 CⅠβ4 6 0.28 157 EⅥγ1 3 0.14 207 DⅡδ5 1 0.05 8 GⅢδ4 33 1.53 58 GⅣγ2 13 0.6 108 EⅣδ4 6 0.28 158 FⅡδ5 3 0.14 208 DⅢβ2 1 0.05 9 DⅡγ4 32 1.49 59 HⅥε1 13 0.6 109 FⅡδ6 6 0.28 159 GⅡδ5 3 0.14 209 DⅣδ4 1 0.05 10 EⅢγ4 31 1.44 60 BⅠβ3 12 0.56 110 FⅣε4 6 0.28 160 GⅢε6 3 0.14 210 DⅣε3 1 0.05 11 GⅣδ3 31 1.44 61 BⅤβ1 12 0.56 111 GⅣγ1 6 0.28 161 GⅣε3 3 0.14 211 DⅥε1 1 0.05 12 HⅥζ2 31 1.44 62 CⅢβ2 12 0.56 112 GⅥδ1 6 0.28 162 GⅤδ1 3 0.14 212 EⅣγ1 1 0.05 13 GⅣδ2 30 1.39 63 EⅢδ3 12 0.56 113 AⅤγ1 5 0.23 163 GⅥε2 3 0.14 213 EⅤε3 1 0.05 14 AⅡβ2 29 1.35 64 EⅢδ5 12 0.56 114 BⅠα2 5 0.23 164 HⅣδ2 3 0.14 214 FⅢδ6 1 0.05 15 DⅢγ3 29 1.35 65 FⅤε2 12 0.56 115 BⅡγ4 5 0.23 165 AⅡα1 2 0.09 215 FⅥζ2 1 0.05 16 AⅠβ4 28 1.3 66 AⅢγ2 11 0.51 116 CⅠβ6 5 0.23 166 AⅢγ4 2 0.09 216 FⅥζ3 1 0.05 17 HⅥε2 27 1.26 67 CⅡγ3 11 0.51 117 CⅠγ6 5 0.23 167 AⅣδ2 2 0.09 217 GⅡβ4 1 0.05 18 AⅠα2 26 1.21 68 DⅠγ6 11 0.51 118 DⅡγ3 5 0.23 168 BⅠα6 2 0.09 218 GⅡγ3 1 0.05 19 DⅡγ5 26 1.21 69 GⅢγ4 11 0.51 119 DⅣγ1 5 0.23 169 BⅢα1 2 0.09 219 GⅢε5 1 0.05 20 EⅡγ5 26 1.21 70 AⅠβ5 10 0.46 120 DⅤδ2 5 0.23 170 CⅡδ6 2 0.09 220 GⅥγ1 1 0.05 21 EⅡγ6 26 1.21 71 BⅠβ6 10 0.46 121 DⅥγ1 5 0.23 171 CⅢδ4 2 0.09 221 HⅣδ4 1 0.05 22 EⅡγ4 25 1.16 72 BⅡβ4 10 0.46 122 FⅥδ1 5 0.23 172 CⅤβ1 2 0.09 222 HⅤδ3 1 0.05 23 GⅤδ2 25 1.16 73 CⅣγ1 10 0.46 123 FⅥε2 5 0.23 173 CⅤδ1 2 0.09 223 HⅤζ3 1 0.05 24 DⅣδ3 24 1.12 74 CⅣγ2 10 0.46 124 GⅣδ4 5 0.23 174 CⅥβ1 2 0.09 224 HⅤζ4 1 0.05 25 GⅢδ3 24 1.12 75 DⅤδ1 10 0.46 125 GⅣε4 5 0.23 175 CⅥδ1 2 0.09 26 DⅣδ2 23 1.07 76 FⅢδ4 10 0.46 126 HⅥζ1 5 0.23 176 DⅡβ3 2 0.09 27 EⅣδ2 23 1.07 77 GⅤε2 10 0.46 127 AⅡγ5 4 0.19 177 DⅡβ4 2 0.09 28 CⅡγ4 22 1.02 78 BⅡβ2 9 0.42 128 BⅠα4 4 0.19 178 EⅠγ6 2 0.09 29 EⅣδ3 22 1.02 79 DⅢγ4 9 0.42 129 BⅡα2 4 0.19 179 EⅣε3 2 0.09 30 FⅣδ3 20 0.93 80 DⅥδ1 9 0.42 130 BⅣγ2 4 0.19 180 EⅣε4 2 0.09 31 GⅢγ3 20 0.93 81 FⅣδ4 9 0.42 131 BⅥγ1 4 0.19 181 EⅥε1 2 0.09 32 AⅡβ4 19 0.88 82 BⅠα3 8 0.37 132 CⅢγ4 4 0.19 182 FⅢδ3 2 0.09 33 CⅡγ5 19 0.88 83 BⅢβ1 8 0.37 133 EⅤε2 4 0.19 183 FⅤδ1 2 0.09 34 FⅣδ2 18 0.84 84 BⅢβ2 8 0.37 134 EⅥδ1 4 0.19 184 GⅠγ6 2 0.09 35 CⅡβ4 17 0.79 85 BⅢγ2 8 0.37 135 FⅥε1 4 0.19 185 GⅤε3 2 0.09 36 CⅢγ2 17 0.79 86 CⅠβ5 8 0.37 136 GⅢγ2 4 0.19 186 HⅣδ3 2 0.09 37 DⅢγ2 17 0.79 87 CⅡβ2 8 0.37 137 GⅤγ1 4 0.19 187 HⅣε4 2 0.09 38 EⅣγ2 17 0.79 88 CⅤγ1 8 0.37 138 GⅥε1 4 0.19 188 HⅤδ2 2 0.09 39 GⅢδ5 17 0.79 89 DⅢδ3 8 0.37 139 HⅣε3 4 0.19 189 HⅥζ3 2 0.09 40 HⅤε2 16 0.74 90 DⅢδ4 8 0.37 140 AⅠα5 3 0.14 190 AⅠα1 1 0.05 41 AⅠβ3 15 0.7 91 DⅢδ5 8 0.37 141 AⅠα6 3 0.14 191 AⅢα1 1 0.05 42 AⅡα2 15 0.7 92 DⅤγ1 8 0.37 142 AⅠβ6 3 0.14 192 AⅢβ1 1 0.05 43 AⅡγ3 15 0.7 93 EⅤδ1 8 0.37 143 AⅣβ1 3 0.14 193 AⅣδ3 1 0.05 44 AⅢβ2 15 0.7 94 FⅣε3 8 0.37 144 AⅣγ1 3 0.14 194 AⅤβ1 1 0.05 45 BⅠβ5 15 0.7 95 FⅤε3 8 0.37 145 AⅣγ2 3 0.14 195 AⅤδ1 1 0.05 46 DⅡδ6 15 0.7 96 GⅢδ6 8 0.37 146 BⅠγ6 3 0.14 196 AⅥγ1 1 0.05 47 EⅢδ4 15 0.7 97 AⅡγ4 7 0.33 147 BⅡγ3 3 0.14 197 BⅠα5 1 0.05 48 GⅡγ5 15 0.7 98 AⅢγ3 7 0.33 148 CⅡγ6 3 0.14 198 BⅣα1 1 0.05 49 GⅡγ6 15 0.7 99 BⅢγ3 7 0.33 149 CⅢδ3 3 0.14 199 BⅣγ1 1 0.05 50 HⅤε3 15 0.7 100 BⅣβ1 7 0.33 150 CⅣβ1 3 0.14 200 BⅣδ2 1 0.05
Simming
trajectory Number Swimmingtime Number Swimmingdistance Number
Average swimming
speed (cm/sec)Number
A 389 I 299 α 152 1 217 B 242 II 628 β 483 2 530 C 244 III 502 γ 712 3 629 D 301 IV 359 δ 595 4 397 E 351 V 212 ε 167 5 192 F 148 VI 151 ζ 42 6 186 G 350 H 126
Table 4 Number of classifications of various elements
Time of day (24h) (N=2,151) 200 150 0 50 100 To ta l A ct iv ity (N ) 0 1 2 3 4 5 6 7 8 9 1011121314151617181920212223
< Fig. 5 Noda >
Fig. 5. The total number of hourly activities
Rank-ing
Aug., 2008 Sep. Oct. Nov. Dec.
Behavior
pattern Num-ber Behavior pattern Num-ber Behavior pattern Num-ber Behavior pattern Num-ber Behavior pattern Num-ber
1 AⅠα3 35 AⅠα3 15 EⅢγ3 6 EⅢγ3 10 AⅡβ3 20
2 CⅡβ3 31 GⅡδ6 14 AⅠα3 5 AⅡβ4 9 GⅣδ3 9
3 EⅢγ3 31 BⅡβ3 13 FⅢδ4 5 AⅡβ2 7 BⅤβ1 8
4 AⅡβ3 30 BⅠβ4 9 GⅡγ5 5 EⅣδ2 7 EⅢγ3 7
5 HⅥζ2 27 EⅡγ6 9 EⅡγ5 4 AⅡβ3 6 AⅡβ2 6
6 EⅡγ4 25 DⅠγ6 8 GⅢδ4 4 GⅣδ3 6 GⅢδ3 6
7 DⅡγ4 24 GⅡγ6 8 AⅡβ3 3 BⅢγ3 5 GⅢδ4 6
8 BⅠβ4 21 GⅢδ4 8 BⅡβ3 3 DⅢγ3 5 GⅣδ2 6
9 EⅢγ4 21 AⅠα2 6 CⅡγ4 3 FⅣδ2 5 DⅣδ3 5
10 AⅠβ4 17 CⅡβ4 6 CⅢγ3 3 GⅢδ3 5 GⅢγ3 5
Fig. 6. The monthly ratio of various elements
A-H : Swimming trajectory, I-VI : swimming time, α-ζ: Swimming distance, 1-6 : Average swimming speed
Fig. 7. The hourly ratio of various elements
A-H : Swimming trajectory, I-VI : swimming time, α-ζ: Swimming distance, 1-6 : Average swimming speed
lightning strike No./
Catfish DATE TIME
Elapsed
time(sec)Catfish behav-ior pattern Ampere(kA) Distance(km) dischargeCloud
7 2008/7/29 13:19:48 26 11 0 8 2008/7/29 13:19:48 11 9 0 Catfish 2008/7/29 13:19:50 2 EIIγ5 27 2008/8/20 22:12:54 7 7 1 Catfish 2008/8/20 22:12:08 46 DIIγ5 30 2008/8/21 19:16:30 11 12 0 Catfish 2008/8/21 19:19:44 194 BIβ4 31 2008/8/21 19:30:27 8 5 1 Catfish 2008/8/21 19:33:38 191 DIIIδ6 35 2008/8/21 20:20:06 6 12 0 Catfish 2008/8/21 20:23:17 191 BIβ4 60 2008/8/29 5:39:59 10 2.5 0 Catfish 2008/8/29 5:46:43 404 GIIγ4 97 2008/12/5 16:11:38 143 10.5 0 Catfish 2008/12/5 16:00:21 677 HVδ2 104 2008/12/16 15:52:13 8 6 0 Catfish 2008/12/16 16:02:57 644 HIVδ3
Table 6 Comparison of catfish behavior and lightning strike time Fig. 8. Lightning strike location map
●: Lightning strike location, Number : Please refer to attached.
140.1 140.1 140.2 140.2 140.3E 140.3 35.1 35.1 35.2N 35.2 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 5km 10km K amogawa C ity C atfish < Fig. 8 Noda >
謝 辞 本研究は東海大学海洋研究所の長尾年恭教授, 千葉大学大学院理学研究院の服部克巳教授による 千葉大学海洋バイオシステム研究センターの「共 同研究」の成果(2006,2007,2008)を使用させて 頂き,研究を遂行するにあたり有益なご指導とご 助言を賜り深く感謝いたします.東海大学海洋研 究所の織原義明博士,静岡県立大学グローバル地 域センターの鴨川仁博士,一般社団法人防災減災 技術開発機構の藤縄幸雄博士には,研究方法での 有益なご助言を頂きました.千葉大学海洋バイオ システム研究センター技術専門職員の瀧口謙一 氏,千葉大学大学院理学研究院技術補佐員の吉野 千恵氏には,観察水槽の構築と飼育,観察システ ムの保守などにおいて多大なる協力を得ました. 深謝の意を表します. 引用文献 長尾年恭(2006-2008)ナマズの行動パターンと地震の関係に関す る研究,千葉大学海洋バイオシステム研究センター「共同 研究」報告書. 野田洋一・細井敦子・高島千代里・浅野昌充・江川紳一郎・長 尾年恭(2002)ナマズの行動と刺激要素に関する研究(序報) ―地震とナマズの関係解明にむけて―.東海大学紀要海洋 学部,54,49-68. 野田洋一・江川紳一郎・長尾年恭(2004)東京都水産試験場のナ マズと地震の関係に関する研究の変遷,東海大学海洋研究 所研究報告,25,91-110. 力武常次(1979)犬が悲しく泣く−動物達の異常行動−.工業調 査会,東京,215pp. 東京都防災会議地震部会(1980-1992)地震予知に関する調査報告 書,昭和54∼平成3年度,魚類の異常生態に関する調査研 究(その1∼その13),東京都総務局災害対策企画課(研究担 当東京都水産試験場). 若井謙治(2004)ナマズと地震の関係に関する研究−画像解析ソ フトを利用したナマズの行動解析の試み−,東海大学海洋 学部海洋資源学科2004年度卒業論文. 付録資料(別添) 資料1:ナマズの行動分類結果 資料2:ナマズの活動プロット図 資料3-1:ナマズの遊泳軌跡(輪郭抽出処理による) 資料3-2:ナマズの遊泳軌跡(重心座標計算による) 資料4:ナマズの行動と落雷活動との比較図 資料5:雷活動一覧表