底生生物から考察する大洗サンビーチ沖域における
鹿島灘はまぐり生息の適否
半澤浩美・山崎幸夫
Study of habitats of hard-clam by macrobenthic community, in offshore at Oarai-beach
Hiromi HANZAWA・Yukio YAMAZAKI
Key Word:大洗サンビーチ,チョウセンハマグリ,生息環境,底生生物
目 的
鹿島灘の最北端に位置する大洗サンビーチは,大洗 港の南端に隣接する約 2km の砂浜海岸である。大洗港 の南防波堤と沖防波堤の遮蔽域にあたり,北向きの沿 岸漂砂によって大規模な堆積域が形成されたとされる (三村ら 1991)。汀線域には鹿島灘はまぐり(標準和 名:チョウセンハマグリ,以下「ハマグリ」とする。) の殻長 30mm に満たない稚貝の分布がほぼ毎年確認さ れる(根本ら 2009)。2000 年以降は毎年,発生から 1 年後とみられる殻長 1∼10mm ほどのハマグリが,大洗 サンビーチ全体で 100 万個体以上生息したと推定され ている。とくに 2005 年・2007 年・2008 年級群は約 800 万個体以上が生息していたと推定されている(杉浦 2009)。また,発生から 2 年後以降とみられる殻長 30mm ほどのハマグリの分布も確認されるが,これらは潮干 狩りの対象となっている。 大洗サンビーチの沖域におけるハマグリの分布につ いては,鹿島灘の二枚貝類の資源量を把握するために 茨城県水産試験場が毎年 6∼8 月頃に行う,貝桁網を用 いた調査で把握されてきた。ハマグリは,生後 3 年で 殻長 45∼60mm に達すると汀線付近から沖合へ移動す ることが知られているが(茂野 1955,藤本 1959a), 大洗サンビーチの沖合では,鹿島灘の他の水域と比較 して採集されるハマグリの数が少ないことから,大洗 サンビーチの汀線域に分布するハマグリは沖合には移 動せず,漁獲加入しない可能性が指摘されてきた(所 2006※)。しかし,この指摘は 1 年に 1 度行われた調査 結果に基づく可能性の提示であり,大洗サンビーチの 沖域におけるハマグリの分布の実態については詳細に は調べられていない。さらに,防波堤によって波が遮 蔽される水域である大洗サンビーチ沖域では,ハマグ リの分布は開放的な砂浜海岸域とは異なることが予想 される。 本研究では,大洗サンビーチ沖域におけるハマグリ の分布実態を明確にするとともに,同所に生息する特 徴的な大型底生生物の分布と底質の粒度組成分析結果 から,ハマグリの分布と関連する要因を検討した。方 法
調査は,2010 年 4,9,12 月および 2011 年 2 月にそ れぞれ 1 日,漁業調査指導船「あさなぎ」を用いて行 った。調査地点は,大洗サンビーチ北端にあたる大洗 港西防砂堤から南側に約 100m の汀線から沖に向けて 垂直に調査ラインを設定し,そのライン上の距岸 100m から 100m おきに 500m までの 5 点を基点とした(図 1)。 離岸距離は,調査時に「あさなぎ」に搭載されたレー ダーで計測したが,調査日によって潮位が異なるため, 同時に測位した GPS ナビゲータ(GARMIN 社製,eTrex Venture HC)の座標と地図ソフトウェア(GARMIN 社製, MapSource)を用い,7 月の大潮時(推算潮位:−5cm) に計測した最干時汀線からの距離に補正した。調査時 の水深から推定潮位を減じ,標準水深とした。調査時 の推算潮位(大洗;水戸地方気象台発表)は,4 月が +22∼30cm,9,12 月と 2 月は+98∼120cm の範囲であ った。 底生生物は,小型貝桁網(桁幅 53cm,爪隔 15mm,網 目合5mm)を基点からそれぞれ岸と平行に 50m 曳網し て採集した。小型貝桁網の曳網時に,水深と水温を計 測した。採集物は生体のまま実験室に持ち帰り,査定 等に供した。貝類の同定は奥谷編(2001)に,そのほ ※所 高利(2006)鹿島灘はまぐり幼貝の移植放流効果の検討.第 20 回波崎海洋研究施設研究成果報告会要旨集,4 −1∼3.図 1 調査地点の概要 矢印は基点を示す かの生物の同定は西村編(1995)に従った。ハマグリに ついては,デジタルノギスを用いて殻長(mm)を計測し た。粒度組成分析用の砂は,各調査基点で,スミス・マ ッキンタイヤ型採泥器を用いて採集した。分析は,水質 汚濁調査指針(日本水産資源保護協会 1980)に従って 乾式で行い,中央粒径値と泥分率(63μm 以下の組成分 の重量比)を計算した。砂の粒径は Wentworth の粒度区 分(須田・早川訳 2002)に従った。
結 果
生物 採集された底生生物は,5 動物門 41 種以上,7,614 個 体以上であった(表 1)。最も多く採集されたのはヒメバ カガイ(2,996 個体)で,ついでハスノハカシパン(1,136 個体),チヨノハナガイ(942 個体)と続いた。 ハマグリは 33 個体採集された。すべての調査日に採 集されたが,採集された地点は距岸 154∼336m(図 2), 標準水深 1.5∼3.4m であった(表 1)。貝桁の爪で破損し た 5 個体を除き,殻長を測定できた 28 個体の大きさは, 15 個体が殻長 90∼99mm,10 個体が殻長 100∼110mm の範 囲にあった(図 3)。調査日および離岸距離と殻長の間に 関係はみられず,常に同程度の大きさの個体が採集され た(図 4,図 5)。また,12 月には距岸 202m で,2001 年 9 月に大洗町成田地先(大洗サンビーチから約 4km 南) に放流したハマグリが 1 個体再捕された(刻印標識放 流;放流時殻長 37mm,再捕時殻長 97mm)。 採集された底生生物のうち,ハマグリと同様の分布を 示したのはハスノハカシパンで(表 1),距岸 154∼336m までの範囲で 99%以上の個体が採集された(図 6)。ハ マグリが採集されなかった距岸 384m 以遠でのみ採集さ 図 2 離岸距離と採集された鹿島灘はまぐりの個体数の関係 ○は鹿島灘はまぐりが採集されなかったことを示す 図 3 大洗サンビーチで採集された鹿島灘はまぐりの殻長組成 図 4 調査日と採集された鹿島灘はまぐりの殻長との関係 図 5 離岸距離と採集された鹿島灘はまぐりの殻長との関係 れたのは,ゲンロクソデガイとニッコウガイ科(モモノ ハナガイ,サクラガイ,オオモモノハナ),チヨノハナ 200m 大洗港 鹿島灘 大洗サンビーチ 南防波堤 沖防波堤 200m 大洗港 鹿島灘 大洗サンビーチ 南防波堤 沖防波堤 N = 28 0 5 10 15 20 0 10 20 30 40 50 60 70 80 90 100 110 個 体 数 殻長(mm) N = 28 0 20 40 60 80 100 120 3月1日 6月1日 9月1日 12月2日 3月4日 殻 長 ( m m ) N = 28 0 20 40 60 80 100 120 100 150 200 250 300 350 殻 長 ( m m ) 離岸距離(m) 0 1 2 3 4 5 6 7 0 200 400 600 800 離岸距離(m) 個 体 数合 計 2 3 3 3 0 8 4 5 6 54 9 6 5 0 1 5 5 2 98 4 3 4 53 5 6 2 5 2 0 2 2 80 3 8 4 52 8 5 9 8 1 5 4 2 40 3 3 6 48 2 5 8 9 2 .0 2 .9 4 .2 5 .2 6. 1 1 .6 2 .9 4 .2 5 .3 5. 9 1 .5 2 .3 3 .5 5 .1 5. 5 1 .5 2 .4 3. 4 4 .8 5 .6 ツ メ タ ガ イ C lo ss au la x di dy m a 1 74 2 9 7 1 1 4 2 3 2 3 8 7 1 6 24 4 1 4 7 32 6 4 ム シ ロ ガ イ 科 の 一 種 N io th a sp . 16 1 5 1 ホ タ ル ガ イ O liv el la j ap on ic a 1 57 2 9 3 9 2 3 3 1 0 4 1 8 3 10 7 5 5 19 ト カ シ オ リ イ レ C an ce lla ri a no du lif er a 5 3 2 タ ケ ノ コ ガ イ 科 の 一 種 T er eb ra s p. 1 1 ク チ キ レ ガ イ O ri ne lla p ul ch el lia 1 1 ウ ミ フ ク ロ ウ P le ur ob ra nc ha ea j ap on ic a 1 1 ゲ ン ロ ク ソ デ ガ イ J up it er ia c on fu sa 55 1 4 9 3 4 1 4 1 7 8 1 3 ア カ ガ イ S cp ha rc a br ou gh to ni i 16 1 1 6 1 3 2 1 1 ウ メ ノ ハ ナ ガ イ P ill uc in a pi si di um 1 1 ヒ メ バ カ ガ イ M ac tr a cr os se i 2 9 96 3 1 1 9 5 3 0 14 4 21 8 9 4 8 40 36 2 0 1 5 1 6 8 1 16 3 1 6 3 9 1 0 1 3 1 1 63 3 2 2 ウ バ ガ イ P se ud oc ar di um s ac ha lie ns e 3 49 8 1 9 2 7 19 4 1 6 6 1 2 2 7 1 6 2 3 5 9 7 1 1 5 チ ヨ ノ ハ ナ ガ イ R ae te llo ps p ul ch el lu s 9 4 2< 8 7 0 2 3 7 3 3 フ ジ ノ ハ ナ カ ゙イ C hi on s em ig ra no sa 2 2 キ ュ ウ シ ュ ウ ナ ミ ノ コ ガ イ L at on a ki us iu en si s 8 8 ワ ダ ツ ミ ザ ク ラ B at hy te lli na c it ro ca rn ea 2 2 ウ ズ ザ ク ラ N it id ot el lin a m in ut a 2 2 モ モ ノ ハ ナ ガ イ M o er el ia j ed oe ns is サ ク ラ ガ イ N it id ot el lin a ho kk ai do en si s 1 59 6 41 2 9 3 6 3 0 2 6 3 6 オ オ モ モ ノ ハ ナ M o co m a pr ae te xt a マ テ ガ イ S ol en s tr ic tu s 11 2 3 1 3 2 コ タ マ ガ イ G om ph in a m el an eg is 1 1 ワ ス レ ガ イ C yc lo su ne tt a m en st ru cl is 94 1 9 3 1 3 2 7 4 1 6 5 5 2 チ ョ ウ セ ン ハ マ グ リ M er et ri x la m ar ck ii 33 6 5 4 1 6 3 2 5 1 マ ダ コ 類 O c to p ad id ae s p p . 11 3 2 1 2 2 1 エ ビ ジ ャ コ C ra ng on sp . 17 3 4 1 0 長 尾 類 P e n ae id ea s p p . 42 1 1 2 4 2 3 5 3 6 6 2 1 5 1 ヤ ド カ リ 科 P ag ur o id e a sp p . 26 3 3 9 1 1 ヒ ラ コ ブ シ G ae ti ce d ep re ss us 8 50 5 2 6 3 8 20 3 52 4 4 2 4 6 1 0 5 1 6 6 1 6 5 6 3 1 5 6 6 0 8 1 3 4 サ メ ハ ダ ヘ イ ケ ガ ニ P ar ad o ri pp e gr an ul at a 6 1 2 1 1 1 ク ダ ヒ ゲ ガ ニ 科 A lb u n e id ae s p p . 2 1 1 ク モ ガ ニ 科 M o jo id ae s p p. 2 1 1 カ ク レ ガ ニ 科 の 一 種 P in no th er es s p . 2 2 短 尾 類 の 一 種 B ra h c yu ra s p . 12 2 4 1 2 2 1 ヒ ラ モ ミ ジ ガ イ A st ro pe ct en la te sp in os us 2 1 1 キ ヒ ト デ A st er ia s am ur en si s 2 1 1 ク モ ヒ ト デ 目 O p h iu ri da s p p . 5 1 4 ハ ス ノ ハ カ シ パ ン S ca ph ec hi nu s m ir ab ili s 1 1 36 9 1 3 4 5 8 1 72 1 3 7 1 57 5 2 1 4 6 3 1 5 1 オ カ メ ブ ン ブ ク E ch in oc ar di um c or da tu m 22 2 1 2 9 4 3 1 多 毛 類 P o ly c h ae ta sp p . 2 1 4< 1 1 1 1 9 3 6 9 2 7 9 1 8 12 2 8 3 0 5 5 11 1 8 2 0 ユ ム シ 類 E c h iu ra s p p . 2 1 1 ウ ミ サ ボ テ ン C av er nu la ri a o be sa 37 2 3 2 2 6 3 5 3 1 2 1 3 3 1 表 1 調 査 地 点 の 離 岸 距 離 と 水 深 お よ び 採 集 さ れ た 生 物 4月 1 5 日 9月 1 4 日 水 深 ( m ) 1 2 月 1 3日 2 月 1 0 日 調 査 日 離 岸 距 離 ( m )
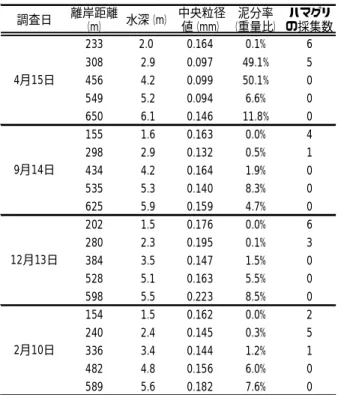
図 6 離岸距離と採集されたハスノハカシパンの個体数との関係 ○はハスノハカシパンが採集されなかったことを示す ガイであった(表 1)。 環境 調査地点の標準水深は,距岸 154m 地点で約 1.5m であ り,離岸距離が 100m 離れるごとに約 1m 深くなり,距岸 650m 地点の水深は 6.1m に達した(図 7)。 底質の中央粒径値は,0.094∼0.233mm の範囲であり, 極細砂ないし細砂に区分された。ハマグリが採集された 地点の中央粒径値は 0.097∼0.195 であった(図 8)。泥 分率は,4 月に距岸 308m と 456m で約 50%に達したほか は,距岸 434m までは 1.9%以下,距岸 482m 以遠では 4.7 ∼50.1%を示した(表 2)。ハマグリが採集された地点の 図 7 調査地点の離岸距離と水深の関係 図 8 調査地点の中央粒径値と採集された鹿島灘はまぐりの個 体数の関係 ○は鹿島灘はまぐりが採集されなかったことを示す 表 2 調査日ごとの調査地点の離岸距離と水深および底質の中 央粒径値と泥分率,ハマグリの採集数 調査日 離岸距離 (m) 水深(m) 中央粒径 値(mm) 泥分率 (重量比) ハマグリ の採集数 233 2.0 0.164 0.1% 6 308 2.9 0.097 49.1% 5 456 4.2 0.099 50.1% 0 549 5.2 0.094 6.6% 0 650 6.1 0.146 11.8% 0 155 1.6 0.163 0.0% 4 298 2.9 0.132 0.5% 1 434 4.2 0.164 1.9% 0 535 5.3 0.140 8.3% 0 625 5.9 0.159 4.7% 0 202 1.5 0.176 0.0% 6 280 2.3 0.195 0.1% 3 384 3.5 0.147 1.5% 0 528 5.1 0.163 5.5% 0 598 5.5 0.223 8.5% 0 154 1.5 0.162 0.0% 2 240 2.4 0.145 0.3% 5 336 3.4 0.144 1.2% 1 482 4.8 0.156 6.0% 0 589 5.6 0.182 7.6% 0 2月10日 4月15日 9月14日 12月13日 泥分率は 1 回を除き 0∼1.2%であり,採集されなかった 地点は 1.5∼50.1%以上であった(表 2)。
考 察
鹿島灘はまぐりの分布と底質および底生生物の分布と の関係 大洗サンビーチ北側の沖域では,ハマグリは距岸 154 ∼336m,標準水深 3.4m 以浅にのみ分布することが明ら かになった。分布するハマグリの大きさは 1 個体を除い て殻長 80∼110mm であったが,調査日および離岸距離と の間に関係がみられなかったことから,この水域に分布 するハマグリは一様にこの大きさであると考えられる。 過去の調査においても,主に距岸 200∼300m で殻長 80 ∼100mm のハマグリが採集されており,大洗サンビーチ の沖域におけるハマグリの分布を反映していると考え られる。 ここで,ハマグリの分布を規定する要因を考えてみる。 大洗サンビーチとよく似た汀線変化で形成された鹿島 港北部に隣接する平井海岸(木村ら 2006,勝山ら 2007) では,距岸 600m,水深 6.6m まで確認されている(半澤 未 発表)。また,茨城県水産試験場による過去の調査では, 距岸 400∼600m に分布の中心があり(根本 2009),複数 の地先で距岸 1300m,水深 11m まで分布することが確認 されている(茨城県 未発表)。大洗サンビーチの沖域に おけるハマグリの分布はこれらとは異なっており,距岸 0 1 2 3 4 5 6 7 0.000 0.050 0.100 0.150 0.200 0.250 中央粒径値(mm) 個 体 数 -7 -6 -5 -4 -3 -2 -1 0 0 100 200 300 400 500 600 700 4月 9月 12月 2月 離岸距離(m) 水 深 ( m ) 0 100 200 300 400 500 0 200 400 600 800 個 体 数 離岸距離(m)300m 以内,水深 3.4m 以浅に分布を制限する何らかの要 因があると考えられる。 まず底質の中央粒径値をみると,ハマグリの分布の有 無に関わらず全ての地点が極細砂から細砂に区分され た。ハマグリ漁場の底質の特徴は,鹿島灘では「よく操 業する場所」の中央粒径値が 0.15mm であり(原田ら 1957),他県の事例も含めると,細砂の卓越と平均粒径 が 0.17∼0.47mm,淘汰係数 0.3∼0.4 とされている(村 上 1969)。しかしながら本研究において,距岸 336m 以 遠,水深 3.4m 以深では中央粒径値がこの範囲にあって も,ハマグリは分布していなかった。このことは,中央 粒径値のみではハマグリの分布を説明することができ ないことを示唆する。 ハマグリは泥分に対して嫌忌性を示すことが知られ ている(安永・日向野 1985)。泥分のハマグリに対する 影響については,静置条件下では泥分自体が短期的な斃 死要因にはならないものの(安永・日向野 1985),懸濁 した泥に対する耐性が非常に弱く,その理由として泥の 粒子が鰓の目詰まりを生じ呼吸を阻害すると推測され ている(日向野ら 1993)。本研究でハマグリが採集され なかった地点の泥分率をみると 1.9%以上であり,距岸 456m 以遠では常に 5%以上を示した。殻長 15mm 以下の 個体では 5%から忌避性が高まるとされており(安永・ 日向野 1985),少なくとも殻長 15mm 以下のハマグリは 大洗サンビーチの距岸 456m 以遠の底質環境を忌避する と考えられるが,泥分率 5%に達しない距岸 456m までの 水域におけるハマグリの分布を説明する要因として,泥 分率のみを示すことは困難である。 最後に,ハマグリと他の底生生物の分布の関係から, 底質について再検討する。ハマグリと分布域が重ならな い底生生物として,ゲンロクソデガイやニッコウガイ科 (モモノハナガイ,サクラガイ,オオモモノハナ),チ ヨノハナガイといった小型の二枚貝類が距岸 384m 以遠 に分布していた。これらの貝類は深所に多く生息し,シ ルト質の穏やかな環境に適応した種であるという(道津 ら 1992,福田ら 2001)。とくにチヨノハナガイは有機汚 染域の指標種として扱われる生物である(横山 1985)。 中央粒径値には反映されなかったものの,これらの二枚 貝類が生息する距岸 384m 以遠の底質は,波浪の影響が 小さく,常に泥分が多く含まれている可能性がある。一 方,ハマグリと分布が重なったハスノハカシパンをみる と,採集数は距岸 308m から沖側では著しく減少し,分 布がみられないこともあった。ハスノハカシパンは浅海 の砂底に生息し(金沢 2009),鹿島灘で過去に行われた 調査からは,分布域がハマグリの分布域より沖側である こと(原田ら 1957,福田 1976),有機汚染の少ない砂 浜に生息すること(高橋・石川 1976)が明らかになっ ている。このことは,大洗サンビーチの距岸 308m 以遠 の環境は泥分もしくは有機物が多く生息に適さないた め,結果的にハマグリと生息域が重複したことを示唆す る。 以上のことから,大洗サンビーチの沖域でハマグリの 分布を距岸約 300m までに制限する要因は,泥分あるい は泥分が堆積しやすい極静穏な海域環境による可能性 が高いと考えられる。 本研究では,粒度組成分析で得られた数値のみではハ マグリの分布を説明することができなかったため,同時 に採集された底生生物の特徴を指標とすることで底質 環境の推定を試みた。ハマグリの生息に適した水域を把 握するためには,中央粒径値のみではなく,底生生物の 分布や泥分率も把握することが有効であると考えられ る。 大洗サンビーチ沖域に分布する鹿島灘はまぐりの由来 本研究によって,大洗サンビーチ沖域には,通年にわ たり主に殻長 80∼110mm のハマグリが生息すると考えら れた。過去に同じ水域で茨城県水産試験場が行った調査 でも,殻長 80mm 未満のハマグリがほとんど採集されて いないことを考慮すると(茨城県 未発表),この水域に 分布するハマグリは別の水域で殻長 80mm に達してから 移動してきたと考えられる。一例ではあるが,本研究で 約 4km 南の水域である大洗町成田地先に放流したハマグ リが大洗サンビーチで再捕されたことは,南の水域から の移動があることを示唆する。一方で沖域へのハマグリ の分散に関しては,鹿島灘の他の水域では,距岸 200m 付近までに殻長 20∼60mm 程度のハマグリが分布し,殻 長 80mm 以上の個体は距岸 300m 以遠に分布するといった 沖灘方向での体サイズ差がみられる傾向にあるが(半 澤・山崎 未発表),大洗サンビーチの沖域ではそのよう な傾向をみることはできなかった。大洗サンビーチでは, 本研究で対象とすることができなかった距岸 150m より も岸側の極浅海域に殻長 80mm 未満のハマグリが分布す る可能性があるが,砕波帯を含む極浅海域におけるハマ グリの分布に関する研究については,ようやく調査が行 われ始めたに過ぎない(半澤・山崎 2011)。 大洗サンビーチは,汀線域に発生後 1 年のハマグリが 高密度に分布する貴重な水域である。しかしその沖域で は,ハマグリの分布は距岸 150∼300m までに制限され, さらに分布するほぼ全てのハマグリが殻長 80mm 以上で あることを踏まえると,汀線域に分布するハマグリの移 動分散と沖域に分布するハマグリの由来を明らかにす
ることが重要な課題といえる。これらの課題を解決する ため,今後は大洗サンビーチ全体の極浅海域から沖域を 網羅した研究を行う必要があるだろう。