Conservation
law
and
Stability in
Competitive
Systems:
Restoration
phenomena
from external perturbation
*,1 Lisa Uechi and *,2 TatsuyaAkutsu
*Bioinformatics
Center, Institutefor
Chemical Research,Kyoto University, Gokasho, Uji, Kyoto 611-0011, Japan
[email protected] [email protected] 非線形競合現象における保存則と安定性: システムの外部摂動からの回復現象 * 上地理沙 * 阿久津達也 * 京都大学化学研究所バイオインフォマティクスセンター 数理生物情報研究領域
A conservation law and stability, recovering phenomenaand characteristic patterns of a nonlinear
dynamical systemhave been studied and applied to physical, biological and ecological systems. Inour
previous study,weinvestigateaconservation law ofasystem of symmetric$2n$-dimensional nonlinear
differentialequations. WeuseLagrangianapproachandNoether’stheoremto analyze Lotka-Volterra
typeofcompetitivesystem. We observe that the coefficients of the 2$n$-dimensional nonlinear
differ-ential equations arestrictly restricted when the system has aconserved quantity, and the relation betweenaconserved systemandLyapunovfunctionis shown in termsof Noether’s theorem. Wefind
that asystem of the$2n$-dimensional first-ordernonlinear differentialequationsin asymmetric form
should appear in abinary-coupled form ($BCF$), and a$BCF$ hasa conserved quantity if parameters satisfy certain conditions. In this paper, competitive systems describedby 2-dimensional nonlinear
dynamical ($ND$) model with external perturbations areappliedto population cyclesand recovering
phenomena of systems from microbes to mammals. Thefamous10-year cycleofpopulationdensity
of Canadianlynx and snowshoe hare is numerically analyzed. We find thata nonlinear dynamical system with a conservation law is stable and generates a characteristic rhythm (cycle) of
popula-tiondensity, which we call the standardrhythmofa nonlineardynamical system. The stability and
restoration phenomena arestronglyrelated toaconservation law and thebalance ofasystem. The
standardrhythmofpopulation densityis amanifestationof the survival of the fittestto the balance ofanonlinear dynamical system.
保存則をもつシステムの安定性、回復現象や非線形現象における固有パターンの出現は、 物理現象のみ ならず生命現象や生態系でも基礎研究やその特質についての応用がなされてきた。本研究の先行研究で は、 我々は対照的な相互作用を持つ $2n$ 次元の非線形常微分方程式に従うシステムの保存則を導出した。 非線形競合システムで、主に Lotka-Volterra型の競合項を含むシステムの保存則の導出にあたり、我々 はラグランジャンの手法やネーターの定理を用いた。システムが保存則を持つ場合には、$2n$次元の非線 形常微分方程式に現れる係数の関係は、保存則からの制限を持つことが明らかになりシステムの保存則と Lyapunov関数と関連や、ネーターの定理からも古典的Lotka-Volterraシステムの保存量が導出できるこ とが示された。 また、対照的な$2n$次元一階非線形常微分方程式で記述されるシステムはbinary-coupled form $(BCF)$ で出現し、$BCF$はパラメタが保存則から導出される条件を満たす場合は保存則を持つこと が分かった。本論文では、$2n$次元の非線形競合システムを外部摂動項を含めた形に拡張し、 2次元の非線 形競合モデルと外部摂動を含む保存則モデルを個体数の周期変動や外部摂動からの回復現象に応用した。 そして、Lotka-Volterra型非線形競合モデルの応用例として用いられるオオヤマネコと白ウサギの10年 周期で観測される個体密度変動に対して数値的解析を行った。我々は、保存則を持つ非線形システムは安 定性を持ち、個体数の変動に対し固有のリズムまたはサイクルを持つことが明らかとなった。我々は非線 形競合システムにおける固有のリズムを standard rhythmと呼び、安定性の一つの尺度と定義した。そし て、安定性と回復現象は保存則とシステムのバランスと強い相関があることを明らかにし、個体密度の時 間的変化に対する standardrhythmは非線形動的システムのバランスを保つための一つの適応戦略である 可能性を示した。
1
Introduction
The conceptof stability isimportantinordertounderstandnatural phenomenainphysical,biological and engineering systems. In ourpreviousstudy, we studied the relation between aconservation law and
stability of
a
$2n$-dimensional competitive systemthat contains competitive interactions, self-interactionsfrom
Noether’s
theorem [1]. The$2n$-dimensional
nonlinear ordinarydifferential
equations fora
compet-itive systemconstructed to satisfy the conservation law have properties suchas
the addition law,whichis empirically interpreted
as
recovery frominjuriesof skin and tissues in biologicalbodies.It hasbeen shown by many researchers that
a
relatively simple set of interactionscan
explaincom-plex phenomena in biologicalsystems [2, 3]. Forexample, in 1952, Turing suggested chemical molecular mechanism called the
reaction-diffusion
system [4] which isdefined
as
semi-linear parabolic partial dif-ferential equations. Thisreaction-diffusion
system is well applied for explaining stripe patterns of themarine angelfish, Pomacanthus, and restoration phenomena in its stripe pattems from injuries
was
ob-served [5, 6, 7]. Prigogine also proposedBrusselator modelwith nonlinear ordinarydifferentialequationsto illustrate spatial oscillations and Turing patterns [8]. It is also
an
interestingproblem to investigate in ecological systems ifa
large complex systemshould
bestable or
not, andmany researchers
havedis-cussed the criteria concerningthestabilityof
a
systemfor $n$dimensional
ordinarydifferential
equationsand statistical framework [9, 10, 11, 12, 13]. What would be
a
reason
whya
simple set of interactionscan
explain complex phenomena? Wediscussed a
systemof interactions generalizingLotka-Volterra
type nonlinear competitive interactions and suggested thata
conservation law could be akey to understandcomplex phenomena
even
in biological and ecologicalsystems.We investigated the system of $2n$
-dimensional
coupled first-order differential equations by usingNoether’s theorem, which led to the following results. (i) The form of differential equations and
co-efficients of nonlinearinteractions
are
strictly confined when thesystem has a conservation lawwhich isconstructed by interacting speciesof
a
particularexperimental system. (ii) The conserved quantityofa
system produces
a
Lyapunov function whichis usually employed to studysolutions of nonlinear differen-tial equations. The conserved quantity isconstructed
byNoether’stheorem,but the analysis of Lyapunov function would be used tocheck
solutions todifferential
equations including those fornon-conservative
and dissipative systems. The system ofdifferential equations with conservation law is different in this
respect. (iii) $A$ systemof interactions could be analyzed
as an
assemblyofa
basic binary-coupledform($BCF$). In other words,
a
complex interacting systemcan
be decomposed intoan
assembly ofbinary-coupled systems. The $BCF$ system is
a
simple basic set to explain complex phenomena defined by Noether’s theorem. (iv) The$BCF$ systemwith conservation law indicatesan
addition law whichmay
be interpretedas
the restorationor
rehabilitation phenomena; thoseare
known ina
large system of neuralnetwork
or
computer network whena
small disordered deviceor
a
partofnetworksystem is replaced bya
normaldevice. Theseproperties could be applied to stabilityandrestoration phenomena of biologicalsystems. (v) The conservation law is also useful to check accuracy of numerical solutions to nonlinear differential equations. As
summarized
above,we
discussed that the basic nonlinear system in $BCF$ isstable.
The binary-coupled systemand
addition law supported bya
conservation lawcan
lead
toa
large,stablecomplex system. Thisisanimportantconclusion on the
conserved
binary-coupledmodel. Becausethe$BCF$systemhassuchseveralinteresting properties,
we
willapplythe modelinorder tostudy stability and interaction mechanisms ofbiologicalsystems.In this paper,
we
will explain the propertiesofsolutions witha
conservation law and applications to biological systems. InSection2,we
extend the$BCF$model to simulate externalperturbations numerically.There
are
various prey-predator type competitive modelswith perturbations, however, most ofthemare
with small, stochastic perturbations. The behaviors ofconservation laws with external perturbations
have been seldom considered. We will explicitly discuss properties of the conserved, stable, 2-variable nonlinear interacting system with external perturbations and the conservation law, its indications and
possible applications to nonlinear interacting system. We will show that 2-variable $ND$ model has the
propertiesofrestoration andrecovery from externalperturbations. InSection3,stabilityandpopulation cycles of biological systems
are
examined in terms ofa
conservation law of the system. We will alsoexamine specific examplesof the Canadian lynxand snowshoe hare [14, 15, 16, 17, 18, 19, 20] and the
questionofpopulation cycles, andfood chainofmicrobes in thelake [21, 22]. Conclusionsandsummary ofresults
are
given inSection 4.2
The
model of
binary-coupled
form
(
$BCF$)
2.1
2n-
$ND$system with perturbations
We discussed $BCF$ system and the conservation law of $2n$-nonlinear dynamical (2n-$ND$) model in detail in the previous work [1]. In this study,
we
add external perturbations in $2n$-variable nonlineardifferential equations in order to examine characteristic behaviors of conserved nonlinear interacting systems. It should benoticed that the2n-$ND$model isextended by addingexternal perturbationterms which maintain a conservation law given by Noether’s theorem. The odd variable terms for $x_{i}(i=$
$1,$
$\ldots,$$2n)$
are
$d_{2k,2k-1^{\dot{X}}2k-1}= \sum_{\prime,\iota=1}^{n}\{(\alpha_{\{2ni+2k\}}+\alpha_{\{2n^{2}+2nk+2i-1\}})x_{2i-1}+(\alpha_{\{2n^{2}+2ni+2k\}}+\alpha_{\{2n^{2}+2nk+2i\}})x_{2i}$
(1)
$+\alpha_{\{4n^{2}+2ni+2k\}}x_{2i-1}x_{2i}\}\alpha_{\{4n^{2}+2nk+j\}^{X}j^{X}2k-1},$
where$k=1,$$\ldots,$$n$
.
Theeven variable
terms for$x_{i}(i=1, \ldots, 2n)$are
$d_{2k-1,2k} \dot{x}_{2k}=\sum_{i=1}^{n}\{(\alpha_{\{2ni+2k-1\}}+\alpha_{\{2nk+2i-1\}})x_{2i-1}+(\alpha_{\{2n^{2}+2ni+2k-1\}}+\alpha_{\{2nk+2i\}})x_{2i}$
(2)
$+\alpha_{\{4n^{2}+2ni+2k-1\}^{X}2i-1^{X}2i\}+\sum_{j=1}^{2n}\alpha_{\{4n^{2}+2nk+j\}}x_{j}x_{2k}+c_{2k}},$
where $\dot{x}=dx/dt$, coefficients, $d_{i,j}$ express $d_{2k,2k-1}=\alpha_{2k}-\alpha_{2k-1},$ $d_{2k-1,2k}=\alpha_{2k-1}-\alpha_{2k}$
.
The linear$co$efficients and nonlinear $co$efficients $\alpha_{i},$ $(i=1, \ldots, 8n^{2}+2n)$
are
arbitrary constantvalues. The lastterms$c_{2k-1},$ $c_{2k},$ $(k=1, \ldots, n)$of(1) and (2)
are
constants orpiecewisecontinuous constants, whichare interpretedas
external perturbations (temperature,seasons
and other temporal, external inputs). Oneshould note that constant terms have dimension ofvelocity,
so
theyare
different from actual external perturbations whichare
considered to effectively express external perturbations. Because extemal per-turbations (inputs) changepopulationdensities as$\dot{x}=dx/dt$, wesimulate numerically thoseeffectswith$c_{2k-1},$$c_{2k}$ asexternal inputs. The system has
a
conservation lawderived from Noether’s theoremwhich is proved inthe paper [1]:$\Psi\equiv\sum_{i=1}^{n}\sum_{j=1}^{2n}\{\alpha_{\{2ni+j\}}x_{2i-1}x_{j}+\alpha_{\{2n^{2}+2ni+j\}}x_{2i}x_{j}+\alpha_{\{4n^{2}+2ni+j\}^{X}2i-1^{X}2i^{X}j\}}$
(3)
$+ \sum_{i=1}^{n}\{c_{2i}x_{2i-1}+c_{2i-1}x_{2i}\}.$
Therefore, with theequations from (1) to (3),
we
are
abletoconsider the conservednonlinear dynamical system with external perturbations by employing piecewise continuousconstantterms, $c_{2k-1},$ $c_{2k}.$The physical meaning ofconserved quantities in
a
biological system is difficult to define contrary to classical mechanics in physics, andso
we
would like to explain differences between $\Psi$-function andHamiltonian. The $\Psi$-function in this study is derived from Noether’s theorem with Euler-Lagrange
equations of motion applied to the 2n-$ND$ system. We discussed the binary-coupled form to generalize
The binary-coupled system has the
conserved
quantity ($\Psi$-function) and the $\Psi$-function may havesimilar physical meanings
as
theHamiltonian ofa
system. However, theHamiltonian is definedas
thetotal energy of
a
system, and the energy has the dimension of the work, which is definedas
force $\cross$ displacement [23, 24]. Theconserved
quantity$\Psi$ is constantalong with time,but it isconstructed
frominteractions of 2n-$ND$ system, not fromthe force, kinetic energy and potentialswhich are, inprinciple,
$co$nverted to theworkproducedbythesystem. Hence,the$\Psi$-function may wellbecalled
as the‘conserved
quantity’, but not
as
the Hamiltonian ofthe system. The $\Psi$-function may correspond to (generalized)kineticand potential energies of
a
system, but it isnot possibletoprove that the$\Psi$-functionisequivalentto
the Hamiltonian in termsof
physics. Inthe 2n-
$ND$ model,variables
denote population densitiesof a
systemof extended
Lotka-Volterra
typedifferential
equations, anditis inappropriate todirectly interpretthe $\Psi$-function
as
the total energyor
biomass of the system. However, it is important to comprehendthat the conserved $\Psi$-function controls behaviors andproperties of the system.
Weshowed that theconserved quantity, $\Psi$-function,
can
reproducetheLyapunov function of classicalLotka-Volterra equationsintheprevious$wo$rk [1]. It is essential to understand that Lyapunov functions
for certain systems of differentialequations
can
be derived from Noether’s theorem whena
system hasconserved quantities. Hence, in
conserved
systems suchas
2n-$ND$ systems, the conservation law andNoether’s theorem are fundamental to study properties of the system. The system with Lyapunov function haslimit cyclesand attractors, which designateenergydissipations of thesystem. Thesystems with $\Psi$-functionsarestrictlyconserved systems, whichshouldcorrespond tolimit cycles atagiventime.
The dynamics of the system of$\Psi$-function evolvesaccordingto the conservation law $\Psi$,whichis equivalent
to Lagrangian dynamics in physical systems.
2.2
Properties of
2-variable
$ND$model
The equationsof2-variable$ND$ model
are
produced by setting$n=1(k=1)$ in equations (1)to (3),resulting in
$\dot{x}_{1}=\frac{1}{d_{21}}\{(\alpha_{4}+\alpha_{5})x_{1}+2\alpha_{6}x_{2}+2\alpha_{8}x_{1}x_{2}+\alpha_{7}x_{1}^{2}\}+\frac{c_{1}}{d_{21}}$, (4)
$\dot{x}_{2}=\frac{1}{d_{i2}}\{2\alpha_{3}x_{1}+(\alpha_{4}+\alpha_{5})x_{2}+2\alpha_{7}x_{1}x_{2}+\alpha_{8}x_{2}^{2}\}+\frac{c_{2}}{d_{12}}$, (5)
and the
2-variable
$ND$model has
the followingconservation
law,$\Psi\equiv\alpha_{3}x_{i}^{2}+(\alpha_{4}+\alpha_{5})x_{1}x_{2}+\alpha_{6}x_{2}^{2}+\alpha_{7}x_{1}^{2}x_{2}+\alpha_{8}x_{1}x_{2}^{2}+c_{2}x_{1}+c_{1}x_{2}$
.
(6)The nonlinear interactions can generally represent, for example, Lotka-Volterra type prey-predator, competitive interactions, food-chain relations by adjusting nonlinear parameters$\alpha_{1},$$\ldots,$$\alpha_{8}$
.
Thepiece-wise continuous constants, $c_{1}$ and$c_{2}$
are
usedas
externalperturbations in computer simulations,suchas
environmental conditions which increase
or
decreaseinteracting species in questions. The equations (4)$\sim(6)$ form 2-variable$BCF$ nonlineardifferentialequations with
a
conservation law.Byemploying eqs. (4) $\sim(6)$,
we
will show:(1) solutions tothe binary-couplednonlinear equations maintain acharacteristic $(x_{1}, x_{2})$ phase-spaceof
solutionsandrecovery fromexternal perturbations. Theexternal perturbations
can
numerically reproduce environmental conditions suchas temperature, climate and chemicalsubstances which affect interactingspecies. The nonlinear binary-coupled model
can
be applied to examine responses ofa
system whether theyare
induced from internal interactionsor
externalperturbations.(2) The binary-coupled nonlinear equations with conservation law exhibitstable phase-space solutions,
which
are
interpretedasstability andrecovery ofpopulation-change inabiological system. The properties of the binary-coupled nonlinear interactions will be shown explicitly in numerical simulations.(3) Byemploying the 2-variable binary-coupled model, it is possible to simulate cycles ofmaximaand
minima in population-change, delaysofperiodic times of population cycles for competitive species. Hence, cyclesof population-changewillbediscussed intermsof theconservation law and nonlinear interactions.
Time
(a) 2-variable $ND$ solutions. Solid and dashed
lines represent $x_{1}$ (prey) and $x_{2}$ (predator),
re-spectively. One shouldnotethat theunit of time
shouldbedefined withrespecttoasystemin
con-sideration.
$x_{1}$
Time
(b) Phase-spaceof 2-variable$ND$solutions. (c) Conservation law$\Psi$of 2-variable$ND$. Itis
con-stant with respect totime.
図 1: $A$ 2-variable $ND$solution and Conservationlaw $\Psi.$
Figure l(a) shows the nonlinear interactions between specieswithout external perturbations $(c_{1}=0$
and$c_{2}=0)$,whose coefficients of nonlinearequationsaresetasinTable 1 (Condition 1). Inaview of the classical Lotka-Volterra competitive system, it
can
be interpretedas
that $x_{1}$ and $x_{2}$ represent prey andare
periodic with respect to time, the maximum and minimumof
$(x_{1}, x_{2})$appear
witha
time-delay.Figure 1 (c) shows the numerical value of the conserved function $\Psi$definedby (6),which is constant with
respect to time.
Thesolutions $(x_{1}, x_{2})$ in Figure l(a) showexplicitly
a
time-delayof the peakfor interacting species.The timings ofpeak and delayedpeak
are
determinedby nonlinear interactions and strength ofcouplingconstant.
The solutions $(x_{1}, x_{2})$ in Figure l(b)show
phase-space solutions, whichare
stable
in themeaningthat the
conserved
quantity$\Psi$ ismaintained constant
and phase-spacesolutions
are
inthesame
trajectory for alltime. The unitoftimeshould beconsidered toadjust to the time scale of
a
system inconsideration, because biological unit times
are
generally different from microbes to mammals.Thephase-space diagram 1 (b) and thestraight lineof Figure 1(c)show that the solutionisexact and stable [1]. Thethree figures exhibit important propertiesofsolutions to thesystemof prey-predator type
ofcompetitive nonlinear interactions.
One of theimportantproperties shown by thestable,conserved nonlinear systemisthat theinteracting
species repeattherhythm of maxima and minima of thepopulation. The periods of the rhythm
are
the result ofcomplicated nonlinear interactions, but the systemkeepsthe constant quantity $\Psi$ with respecttotime. The interesting applications of the$BCF$ model
are
shown by employing in thepaper ‘Mysis in the Okanagan Lake food web ‘[21], Canadian Lynx and snowshoe hare [15], which will be explained inSection 3.
2.3
Recovering
and restoration
from
perturbations
In order to investigate
responses
ofa
system toexternal perturbations,we
introduce piecewisecon-tinuousconstants, $c_{1}$ and $c_{2}$, by using
$\theta$-functions such that
$c_{i}=f_{i}\{\theta(t-t_{start})-\theta(t-t_{end})\}, (i=1,2)$, (7)
where$\theta(t-t’)$ representsastep function:
$\theta(t-t’)=\{\begin{array}{l}1, (t\geq t’) ,0, (t<t’) ,\end{array}$ (8)
and coefficients $f_{i}(i=1,2)$
are
positiveor
negativeconstants to express strength of externalperturba-tions. The constants
are
adjusted to produce reasonable maxima and minimainnumerical simulations.Figures 2(a), 2(b) and 2(c) show the reaction and recovery ofthe nonlinear interacting system from an external perturbation. One ofthe typical recoveryof
a
system froma perturbed state is shown. InFigure 2(a), an external perturbation starts at $t=700$ (Sp. 1), and thecoefficient $f_{1}$ equals to $-1260.0$
and$f_{2}$ equals to
zero
inthis example. Theblackarrow
isthe starting point of perturbation, and thegrayarrow
istheend ofperturbation in Figures 2(a) and 2(c). The nonlinear coefficientsare
listed inTable 1(Condition 1). Thesolutions $(x_{1},x_{2})$
are
deformedbytheperturbation(Figure 2(a) and $2(b)$). However,the system does not disintegrate butfinds
a new
stablephase-spaceclose totheoriginal$phasrightarrow$space andmaintains
a new
conserved relation. The perturbation endsat$t=1200$ (Ep. 1), and the systemrecovers
the original state $(x_{1}, x_{2})$
.
Thetimingofnegativeperturbation
which
reduces the population number$x_{1}$or
$x_{2}$produces differentresults. Whenanegative perturbation isexertedintheincreasing phaseof$x_{1}$
or
$x_{2}$, thesystemwill finda new conserved stable solution
near
the original solution, but whena
negative strong perturbation is exerted before$x_{1}$ or $x_{2}$ gets to itsminimum, the system may collapse: the system exhibits nosolutionsTime
(a)2-variable$ND$solutions witha negative
pertur-bationonprey$X1$. The perturbationis introduced
from$t=700$ to$t=1200$which is representedas
graybackground.
$x_{l}$
Time
(b)The$(x_{1},x_{2})$phase-spacetransitionwith the (c) Conservation law$\Psi$of 2-variable$ND$withone negativeperturbationasin(a). Solid line(St. 1) perturbation. $\Psi$changed$\Psi\simeq 60000$to$\Psi\simeq 30000$
is initialstate, andSt. 2is therecovered state by introducing perturbation. It recovers after
after the end of perturbation. Dashed line is Ep. 1.
phase-space during Sp. 1-Ep. 1.
図 2: An external perturbation anda recovery.
The conserved nonlinear system naturallyexhibits maxima and minima without external perturba-tions, and
so
we
callthese maxima and minimaas
endogenous maximumand minimum. It is needed to distinguishthemfrom enhanced maxima andminima byexternal perturbations.In Figure 3, the response of a strong negative perturbation to prey after the peak of endogenous maximum is shown. The values ofcoefficients are listed in Table 1 (Condition 1). The starting point of this perturbation is at $t=800$ and the end point of the perturbation is at $t=950$. The negative
constant ofperturbationis $f_{1}=-3175.3879$
.
The prey, $x_{1}$, rapidlydeclines with negative perturbation,and $(x_{1}, x_{2})$ converges to
zero
for$t>1000\sim$.
Thesecomputersimulations may becompatiblewith knownempirical results, for example, in pest control. $A$pest controlis not
so
effective ifit is performed intheseason
when harmfulinsectsare
in peak and active, because speciesare
energetic enough to finda newstablelife to live. It iseffectivewhenapestcontrol is performed inthe
season
whenharmful insectsare
not
so
activeor inadecliningstateafter endogenousmaximum.In thenonlinearinteracting system, positive perturbations which will increase$x_{1}$ or$x_{2}$ donot always
mean a
positiveeffecton
stabilityofthe system. Thereisa
limitto the value ofa
positiveperturbation,Time
Time
(a) 2-variable$ND$solutions with acritical neg- (b) Conservation law $\Psi$ with acritical negative
ativeperturbationon prey, $X1$. Solutions con- perturbation. $\Psi$convergestozero afterthecritical
vergetozeroafter the perturbation. perturbation.
図3: Critical negative perturbation andextinction.
system hasinternallyallowed maximum and minimum populations.
Figures 4 shows the behaviors of$(x_{1}, x_{2})$ at normal and criticalvalues of positive perturbations, $c_{1},$
for $x_{1}$
.
The values ofcoefficientsare
listed in Table 1 (Condition 2). Figures 4(a) and 4(b) show thatthe normal positive perturbation which increases interacting species will increase the peak of $(x_{1}, x_{2})$
populations. However, at certain critical values of coupling constants, the prey-predator interaction cannot keep andsupportthe rhythm of maxima and minima, and the systemdiverges. Figures4(c) and
4(d) show that thesystem cannot maintain
a
stable, interactingsystem when the positiveperturbationsurpassesthe critical value ($c_{1}=1599.924999$ inthe current simulation). The unstablesolutionsbranch
out at$t\simeq 1100$ when thevalue of perturbationchanges from $f_{1}=1160.0$ to $f_{1}=1599.924999.$
Hence,in
a
conserved stablesystem, speciesseem
to strictly control each other by seekinga new
stable solution so that they can survive together. The competitive interacting system suchas
the conserved prey-predator relationsmay
beconsidered
to bea
cooperative systemfor
speciesto survive. Itshould
be noted that ifa
dynamicalprey-predator system is active,therhythmsof maxima and minimaare
clearlyrepeated, whichis known inreal prey-predator systems. However,ifan external perturbation (exogenous
interaction)exceeds a certain critical value of thecompetitive system,therhythmsof maxima andminima will disappear first and then afteratime, the system will diverge (disintegrate). Therefore, the rhythm
ofwild-life indicates that the dynamical interactions between species
are
active and stable. When the rhythm of changedisappearor
does notcome
back, it may indicate that related speciesare
in dangerof extinction. The rhythm is important toexamine if the wild life is normal and active,
or
harmed by human activities and external perturbations.On the other hand, by adding another perturbation,
we can
show that it is possible tosave
speciesfrom extinction. Figure 5(a) is
a
result of a positive perturbation tosave
species $(x_{1}, x_{2})$ in a dangerofextinction in Figure 3(a). We exerted
a
positive perturbation after Sp. 1 - Ep. 1 in Figure5.
Thepositive perturbationsstart at $t=1000$ (Sp. 2) and end at $t=1300$ (Ep. 2), the strengths of$c_{1}$ and $c_{2}$
are
$f_{1}=200,$ $f_{2}=-1000$. Figure 5(a) showsthat speciesare
indangerofextinction,however, ifpositiveexternal perturbationsare properly inserted, the systemwill
come
backto lifeagain.2.4
Comments on “atto-fox problem”
It should be noticed that
a
problem knownas
“atto-fox problem” [25, 26] ina
system of differential equations will notoccur
in a conserved system ofdifferential equations, because theproblem is related$\tilde{\approx\dot{vg}}h$
$\frac{.\overline{\Leftrightarrow}}{\underline {}a}$
$a^{e}\approx\Rightarrow$
Time
Time
(a) 2-variable$ND$solutions with a positiveper- (b)TheConserved quantity$\Psi$withapositive
per-turbation. Theperturbation starts at $t=500$ turbation. Itrecoversafterperturbationbutfinds
and ends at $t=900$ represented as gray back- anotherequilibriumstate.
ground. The amplitudes of$x_{1}$ and $x_{2}$ become
larger than before.
Time
Time
(c) 2-variable$ND$ solutionswith acritica] per- (d) Conservation law of 2-variable$ND$witha
crit-turbation. $x_{1}$ and$x_{2}$ convergeto zero after a ical perturbation. The$\Psi$convergestozeroaftera
critical perturbation. criticalperturbation.
図 4: Critical perturbations and divergenceof solutions.
to properties of the conserved
or
non-conserved system ofdifferential equations. The 2n-$ND$ system has the conservation law and the $\Psi$-function characterizes behaviors of solutionsand systems. If $\Psi-$
functionis conserved andnotequal to zero, Ae solutionwillconvergeand the system will be stable. The
nonlinear ordinary differentialequations with a conservation law
can
have a stable solution controlledby the $\Psi$-function, andsolutions consist of
a
closed hyper-surface of$(x_{1}, x_{2}, \ldots, x_{2n})$for $2n$-dimensional
case.
It should be noted that the admissible coefficients of nonlinear interactionsare
strictly confined by$\Psi$-function of the system.
The $\Psi$-function will not be constant when there
are no
solutions
or
unphysical solutions, and the property to maintain $\Psi$-functionas
constant will confine admissiblesolutions [1]. For example, if the
2-variable nonlinear interacting system has solutions which areextremely different
as
$10^{-18}$ orders hke“atto-foxproblem”, it isnot possible that thesystem
can
maintain $\Psi$-functionas
constant intime. Thephenomenon like “atto-foxproblem” would appearin dissipative
or
non-conservedsystems, becausenon-conserved and dissipative systems do not have the conservation law to control admissible solutions, and
alarge class of (unphysical) solutions
can
beallowedcomparedtothe system of$\Psi$-function, which istheTime
Time
(a) 2-variable $ND$solutions with perturbations (b)Conservation law $\Psi$withtwoperturbations in
toavoid convergingtozero afteracritical per- (a). $\Psi$recoversfrom the perturbationafterSp. 2
turbation. $x_{1}$and$x_{2}$comebackto lifeafterthe -Ep. 2.
second perturbations.
図5: Thecriticalbehavior and restoration.
conserved systemwith $\Psi$-functionwillproduce physicalsolutions controlledbythe conservationlaw,and
the phenomenon like the
“atto-fox
problem” will not be allowed ina
conserved systemwith $\Psi$-function.3
Conservation law and population
cycles
3.1
The
food-web
of
Microbes in
Okanagan
Lake
One of interesting data of the ecological interactions is the interaction described in ‘Mysis in the OkanaganLake food
web: a
time-series analysisof
interaction strengths inan
invaded plankton commu-nity ‘[21]. Althoughthe food-webin OkanaganLakeis notclarified
definitely,mysisintroduction
tolakes
isknown
as
an
effective methodtoenhance ecological interactions and its strengthsamong
microbes and other creaturesso as
to increase fisheriesproductions.The time-series of dominant crustacean zooplankton densities in Okanagan lake has been measured
monthlyand suggestedthatmysisandzooplankton populations
are
synchronousandcharacterized by thecycleof thepeakand bottom population densities. Thecyclesofpopulation densities
are
primarilydue tocyclesof
season
and climate and then to mutual interaction of microbes. The analysis of microbes suggeststhat the density-dependent and delayed population regulationof microbesisevident. In addition to the seasonalfactors, the regular cycles and the delayed peak and bottom populations densities of microbes
are
the results of strong nonlinear interactions ofspecies. Wenumericallyexaminedchangesofpopulation densities of microbesby employingthe2-variable conserved $ND$ model.Thecurrent
conserved
nonlinearmodel
shows that the interacting species designatea standard
rhythm of the peak and bottom population densities. Thereare some
fluctuations at the peak and bottomdensities, buttheyshow the stabledynamiclife
as
demonstratedinFigure6 $(a)\sim(c)$.
Although, normalpeak and bottom densities
can
be readily explained by adjusting coupling strength of model’s internal interactions,a
suddenchange ofmaximawhich isoften encountered inabiological datacannot be easily simulated by onlyadjustinginternal coupling constantsinthe 2-variable nonlinear interactingmodel.InFigure 6, several perturbations
are
exertedon
theinteracting2-variablesystem. The first extemalperturbation starts at $t=500$ (Sp. 1) and ends at $t=1000$ (Ep. 1). The strength ofperturbations in Sp. 1-Ep. 1
are
$f_{1}=-800,$ $f_{2}=-100$.
The second external perturbationstarts at $t=1400$ (Sp. 2) andTime
(a) 2-variable $ND$ solutions with three external
perturbations. The rhythm of$x_{1}$ and $x_{2}$
recov-ersfrom severalperturbations. Graybackgrounds
represent periodsofperturbations.
$x_{1}$
Time
(b) Phase-space transitions of $x_{1}$ and $x_{2}$. (c) Conservationlaw$\Psi$with three perturbations.
Dashed lines represent solutions, $x_{1}$ and $x_{2}$, Itrecovers from three perturbations. $\Psi\simeq 60000$
during perturbations. Solid line represent so- in the St. 1 andSt. 2.
lutionswithout perturbations.
図6: Severalexternal perturbationsandrecoveries.
third external perturbation starts at $t=2200$ (Sp. 3) and ends at $t=2600$ (Ep. 3). The strength of
perturbations in Sp. 3-Ep.3is set
as
$f_{1}=-500,$ $f_{2}=-50$.
Thelines$(x_{1}, x_{2})$ mayrepresentforinstance,theprey-predator interactions, speciesof food-chain,andspeciesinteractingwith its environmentalfactors
(temperature
or
some
environmentaleffects). Blackarrows are
starting point ofperturbations, and grayarrows are
the end ofperturbations; parametersare
listed inTable 1 (Condition 1). The timeperiodiswithin$t=4000$, initial values
are
$x_{1}=500,$ $x_{2}=300.$The significant propertiesof the stable nonlinear conserved system arethat if external perturbations
arenot large enough to disintegrate the system, the system will find astable conserved solution nearthe original system and continuea stable cycle (maximaand minima). Itis clearlyseen from $(x_{1}, x_{2})$-phase
space solutions inFigure 6(b). Thesystem
recovers
from several externalperturbations.The numerical analysis
can
be applied to examine the change ofpopulation densities of microbes. For example, the time-series data of dominant crustacean zooplankton densities in the Figure 2 of thepaper ‘Mysis in the Okanagan Lakefood-web ‘, show that the sudden maxima of dominant zooplankton
densitiesare seen in the period $99\sim 02$
.
The sudden increaseofthe peak is readily adjusted when ancouplingconstants in the
2-variable nonlinear model.
Hence,it isconcluded
inthe 2-variable model that
there would have
been
certain positiveexternal
perturbation to thesystemofmicrobesinOkangan Lakeduring $98\sim 01$ considering atime-delayofextemal perturbations.
It is interesting tocheckwhat kindof
external or
internalperturbations is affectingthe peak of popu-lationdensity during theperiod $98\sim 01$.
If thereare
no
explicitchanges in externalor
internal factorsduringtheperiod,
a
suddenincrease of thepeakcouldbea
resultofmore
complexinternal
interactions. Forexample,the rhythm of the peakand
bottom population densitiesshould
be explained by4-variable
or 6-variablenonlinear interactions of microbes. Theunusualrhythm indicates howexogenous (environ-mental) and endogenous (internal interactions) variables
are
affecting the dynamicsof each componentand environmental nature related tothe species. The analysis of nonlinear modelsuggests thatthe sud-den peak and bottom sud-densities have important information
on
the dynamics of the systemof
speciesand
environment. Hence, itis
importantto understand the standard
rhythmofthe
peakand bottom
populationdensities inorder to distinguish them from unusual maxima andminima.
$O$
ne
should becareful
thata
positive perturbationon
one
of interacting species not only enhancesthe peak ofmaximabut also decreases minima intherhythmofspecies. Itisoftentrue that the effect of
enhancement is usually emphasized without taking
care
after negativeeffects. Hence, the enhancement ofthe numberof population ofa
specific species may be harmful to other species in the food-web andconsequently it endangers itself. Our analyses in Figure 4 and 5 show that if
we
carefully control the increase or decrease ofthe population ofcertain species after introduction of a positive effect,we can
keep normal andstabledynamics of speciessuitable for theenvironment. For thispurpose, it isessential to explicitly understand the standard rhythm from real observed data.
3.2
Population regulation in Canadian
lynx
and snowshoe hare
It is difficult to identify population regulation mechanisms about prey-predator patterns of large
mammals because the large mammal’s life span is relatively long compared with microbes. The
prey-predator cycle such
as
wolves and caribous takessome
decades of years to observe, their interacting relation and behaviors havebeen recently revealed withmoderntechnology (GPS-colored animals) [14]. However, the food-web configuration between snowshoe hare and Canadian lynx is well-knownprey-predatortypephenomena,and
a
ten-year cycle of Canadianlynxwas
examined from the data ofCanada
lynxfur-trades return ofthe NorthernDepartment of theHudson’s Bay Company (thedata
are
fromC. Elton and M. Nicholson [15]$)$.
The Canadian lynx and snowshoe hare have a synchronous ten-year cycle in population numbers
[14, 16]. The fundamental mechanisms for thesecycles
are
maintainedbythe importantfactors suchas
nutrient,predationand social interactions [17]. Inadditiontotheimportantfactors,the nonlinear model
with conservation law suggests that species ofasystem consequently find astrategyor a mechanism to
survive for long-time periods. In other words, the cycleofpopulation density is a manifestation of the
strategy or mechanismtosurvive,whichissuggestedby stabilityofphase-space solutions determined by
conservation law ofasystem.
Thenonlinear interactions with conservation law show
a
standard rhythm andstabilityfrom externalperturbations
as
shown in Figure 6. The feeding and nutrient experiments in [17]are
consideredas
Time
Time
(a) The2-variable $ND$simulation of Canadian (b) The estimated population of Canadian lynx
lynx population. Thesolidline representsCana- and snowshoe hare. The dashed line represents
dian lynx population [15], and thedashed line Canadian lynx population simulated by 2-variable
represents a theoretical solution of 2-variable $ND$ model with perturbations, and the solid line
$ND$with severalperturbations. represents approximate population of snowshoe
hare.
Time
(c)Transition of conservationlaw $\Psi$with respect
to time. Severalperturbationsareintroduced.
図 7: Simulation of Canadianlynx and snowshoe hare.
external perturbations to thesystem. As shown in Figure 6,the perturbations
cause
certain effectson
the system, but the system will find
a
rhythm to maintain the dynamics of species, which is not so different from the original standard rhythm. Our numericalresults agree with conclusions derived from feedingexperimentsand nutrient-additionexperiments. Therefore, we propose that theproperties of thesystem which has a conservation law should be akey to understand the unanswered question: why do these cycles exist?.
Theresults of computersimulations showthat the timing ofperturbation leads to different results.
This isalso confirmed by the feedingexperiment of snowshoe hare: “
$\cdots$ duringthe peakofthe cycle in
1989 and 1990 had no impact
on
reproductive output $\cdots$ however, during the decline phase in 1991 and1992, thepredator exposureplus food treatment caused a dramaticincreasein reproductive output $\cdots$
”
[17]. This fact canbe examined inourmodel calculations. The perturbation in the peak phase does not
cause large effects on standard rhythm, but negative and positive perturbations during a decreasing
or
increasing phase induce dramatic effects.
The cycle of standard rhythm forCanadianlynx and snowshoe hareindicates that the stable
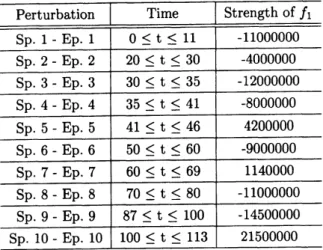
Table 3:
The list ofexternal
perturbations in Figure7.
The periods of positive and negative perturbationsto numerically simulateCanadianlynx population. Note that the values of$f_{1}$ have the meaningofvelocity
(number/time).
conditions. However,
as
we have shown in Figure 4(c) and 4(d), ifa
strong negative perturbation is applied persistently fora
long period, the system would fall intoa
dangerofextinction. The importantresults of
our
simulation tellthatbeforea
system getsin dangerofextinction,the standard rhythm of the system willtend to
become ambiguousor
disappear. Hence,ifwe
carefullyobserve the standard rhythm ofa
specific system of species,we
could
help the dynamical systemsave
and preserve related natural
environment.
In Figure 7,
we
simulated the Canadian lynx data of the Hudson’s Bay Company from 1821 to1910, which isapproximatelythought
as
thelynx-populationdensity. The interpolatedElton’s datawas
downloaded from [27]. The solid-line in Figure 7(a), is lynx-population dataandthe
dashed-line
is theresults of
our
numerical simulation using 2-variable nonlinear interactions between lynx and snowshoehare (Figure $7(b)$). The
conserved
binary-coupled model tells that thereshould
havebeensome
external
perturbations, although we cannot make
sure
atthe present what kinds of extemal perturbationswere
exerted. The actual population densityof snowshoe hare isnot known,and
so
we
assumeda
reasonablepopulation density and several externalperturbations for numerical simulations inorder to fit the lynx
population data (see,Table 2 and Table3).
The
snowshoe
hare getsseveral
positive and negative perturbations, but theoverall
rhythms of lynxand hare
are
not altered. As suggested by in Figure 6(b), the phase-spaceof
lynx andsnowshoe
hareis stable against several external perturbations. This is also compatible with the empirical fact that
the ten-year cycle in snowshoe hare is resilient to a variety of natural disturbances from forest fires to short-term climaticfluctuations. However,
as
shown inour
modelcalculationin Figure4(c), along-term(morethanten years) negativeperturbations and a vast environmentalchangethat humans could
cause
would definitely endangerthe standardrhythmofsnowshoehare, lynx and related species.
4
Conclusions
Inthis paper,
we
examined characteristic propertiesof severalecological systemsbasedon
conserved nonlinear interactions which include generalizedLotka-Volterra
type prey-predator, competitiveinterac-tions. In Section2.1,
we
extendedour
$2n$-variable$ND$ modelby includingexternal perturbations inorderto apply themodel to
more
realistic biological phenomenaand to study responses ofa
biologicalsystemWe simulated extemal positive and negative perturbations by employing piecewise constant terms
in
our
nonlinear equations. As it is discussed in the analysis, the results of simplified perturbations agreed with the experimentsand empirical data reasonably well. The numerical simulationsshowed the existence of the standard rhythm which ischaracteristic toa
nonlinear conserved system. It is essentialto understand standard rhythm by observing and taking dataof a system
so
thatwe
can distinguishunusual maxima and minima fr$om$ standard rhythm This gives
a
possibility to examine signaturesthat
distinguish internal effects
from
externalones.
Theten-year cycleoflynx and hare is a very interesting biological phenomenon. Though
a
cycle ofabiological system should be aphenomenon composedofcomplex and multi-biological interactions, the 2-variable $BCF$ analysis has revealed the interesting results
on
properties ofthe biological phenomena.The ten-year cycle of lynx and hare is stable and resilient to extemal perturbations, which is
repro-duced in
our
model calculations. The system with conservation law shows stablecycles and recovering phenomena, whichare
displayed numerically in phase-space solutions. The stability and conservation laware
constructed at least by binary-coupled species in biological and ecological systems, and theyare
maintained in
a more
complicated multi-coupled system,as we
proved ina
generalform [1].The coupling constants of interacting species expressed in nonlinear differential equations
are
con-sidered to havebeen determined in
a
long time by complicated environmental and internal factors ofa
specificsystem, such
as
the landforms, seasons, climate and temperature. Oncemembers
andstructures
ofdynamical systems
were
constructed, appropriate dynamical systems $wo$uld be maintained for long-time periodswith intemal factors suchas
nutrient, predationand social interactions. Thepredationand social interactionsare
expressed as complicated nonlinear relations in mathematical terms. This maybe explained by thefact that membersof
a
systemhavea well-conserved rhythm respectively andtheserhythms also have a well-determined slight delay to each other, which indicates that certain nonhnear
interactionsamong membersexist.
Theimportantfactors(nutrient, predationand socialinteractions)
are
needed for allspeciestosurvivein nature, but they easily change by natural conditions. In addition, an unusual increase of population
numbers of
a
species would endanger the survival of a species itselfas
wellas
other species (see thenumerical simulations in Figure 4). The important property of the nonlinear model with conservation
lawisthat the binary-coupledsystemcan havethe persistent stability and recovering strengthtoexternal perturbations. As
a
predatorneedsa
prey for its food,a
prey needs a predator for the conservation of theirown
species. The conservation law and rhythmofspeciesare
consideredtobeconstructedby speciesand natural conditions in
a
system fora
long time, and hence, the cycle (rhythm) ofspecieswould beinterpreted
as a
manifestation of the survival of the fittest to the balance ofa
biological system.Weconclude that stability and conservation law
are
constructed by species in mutual dependencyor
cooperationto survive for long-time periods in
severe
nature. The standard rhythmshould be regardedas
the resultof strategy for speciesto live in nature. Whatever roles theyhave to play, the species thatcan
fit and balance with othercreatures
can
survive innature. $A$strong predatorcannoteven
surviveifit ignores the law of thestandardrhythm and conservationlawof
a
systemconstructedbyother members and the environment. Wehopethat this study willhelpunderstandbothactivitiesofanimals and humans参考文献
[1] L.
Uechi
and T. Akutsu.Conservation
laws and symmetries in competitive systems. Progressof
TheoreticalPhysics Supplement, 194:210-222, 2012.
[2] H. Meinhardt. Models
of
biologicalpattem formation, volume6. Academic PressLondon,1982.
[3] A. Gierer and H. Meinhardt. $A$ theory of biological pattern formation. Biological Cybemetics,
12:30-39,
1972.
[4] A.M. Turing. The chemical basis ofmorphogenesis. Philosophical Transactions
of
the RoyalSocietyof
London. Series$B$, BiologicalSciences,237:37-72,1952.
[5] S. Kondo andR. Asai. $A$
reaction-diffusion wave on
the skin of themarine angelfish pomacanthus.Nature, 376:765-768,
1995.
[6] I. Lengyel and I.R. Epstein. Modeling
of
turingstructures in thechlorite-iodide-malonic acid-starch
reaction system. Science, $251:650\triangleleft 52$, 1991.
[7] N. Kumar and W. Horsthemke. Effects of
cross
diffusionon
turing bifurcations in two-species reaction-transport systems. Physical Review$E$, 83:036105, 2011.[8] T. Biancalani, T. Galla,
and
A.J. McKane. Stochastic
waves
ina
brusselator model with nonlocal
interaction. Physical Review $E$, 84:026201,
2011.
[9] R.M. May. Will
a
large complexsystem be stable? Nature, 238:413-414,1972.
[10] J.E. Cohen and C.M. Newman. When willalarge complex system be stable? Joumal
of
theoreticalBiology, 113:153-156,
1985.
[11] K. Tokita. Species
abundance
patterns in complex evolutionary dynamics. Physical review letters,93:178102, 2004.
[12] J. Daniels and A.L. Mackay. The stabilityofconnected linear systems. Nature, 251:49-50,
1974.
[13] A.R. Ives and S.R.Carpenter. Stabilityand diversity ofecosystems. science,$317(5834):58-62$,
2007.
[14] N.C. Stenseth, W. Falck,
K.S.
Chan,O.N.
Bjornstad, M. $O$’Donoghue, H. Tong, R. Boonstra,S. Boutin,C.J.Krebs, and N.G.Yoccoz.Frompatterns toprocesses: phaseanddensity dependencies inthe canadianlynx cycle. Proceedings
of
the NationalAcademyof
Sciences, 95:15430-15435,1998.
$[15]$ C. Eltonand M. Nicholson. The ten-year cycle in numbers ofthe lynx in canada. The Joumal
of
AnimalEcology, 11:215-244, 1942.
[16] N.C. Stenseth,W. Falck, O.N. Bjrnstad, and C.J. Krebs. Populationregulation insnowshoe hare
and canadian lynx: asymmetric foodweb configurations between hare andlynx. Proceedings
of
the NationalAcademyof
Sciences, 94:5147,1997.
[17] C.J.Krebs, R. Boonstra,S. Boutin,and A.R.E. Sinclair. What drives the 10-yearcycleof snowshoe hares? BioScience, 51:25-35, 2001.
[18] M. Basille, D. Fortin, C. Dussault, J.P. Ouellet, and R. Courtois. Ecologically based definition
of
seasons
clarifies predator-prey interactions. Ecography,35:
1-10,2011.
[19] B. Blasius, A. Huppert, and L. Stone. Complex dynamics and phase synchronization in spatially
[20] J. Maquet, C. Letellier, and L.A. Aguirre. Global modelsfrom the canadian lynx cycles
as
a direct evidence forchaosin real ecosystems. Journalof
MathematicalBiology, 55:21-39, 2007.$[21]$ D.E. Schindler, J.L. Carter, T.B. Francis, P.J. Lisi, P.J. Askey, and D.C. Sebastian. Mysis in the
okanagan lake food web:
a
time-series analysis of interaction strengths inan
invaded planktoncommunity. AquaticEcology, 46:1-13, 2012.
[22] N.H. Gazi. Dynamics of a marine plankton system: Diffusive instability and pattern formation.
AppliedMathematics and Computation, 218:8895-8905, 2012.
[23] J.D. Logan. Invariant variationalpnnciples. Academic Press,
1977.
[24] H. Goldsteinand V. Twersky. Classical mechanics. Physics Today, 5:19, 1952.
[25] D. Mollison. Dependence of epidemic and population velocities on basicparameters. Mathematical
biosciences, $107(2):255-287$, 1991.
[26] P.E. Kloedenand C. $P6$tzsche. Dynamics of modified predator-prey models. International Joumal