表 題 AAVベクターによる呼吸器細胞への最適な遺伝子導入法の確立
論 文 の 区 分 博士課程
著 者 名 黒崎 史朗
担当指導教員氏名 萩原 弘一 教授
所 属 自治医科大学大学院医学研究科
地域医療学系
循環器・呼吸器疾患学
呼吸器内科学
2017年1月10日申請の学位論文
目次
1. はじめに ... 1
2. 実験方法と材料 ... 4
2.1 AAVベクタープラスミドの作製 ... 4
2.2 組み換えAAVベクターの作製 ... 5
2.3 細胞培養... 6
2.4 呼吸器系由来細胞株に対する血清型別AAVベクターの 遺伝子導入効果のスクリーニング ... 7
2.5 ルシフェラーゼ発現AAVベクターによる細胞株への遺伝子導入と発現 ... 8
2.6 呼吸器系由来細胞株に対するプロモーター別AAVベクターの 遺伝子導入効果の比較 ... 8
2.7 動物実験... 9
2.8 生体イメージング ... 10
2.9 組織中のAAVベクターの分布と遺伝子発現の測定 ... 10
2.10 組織学的検討 ... 12
2.11 統計学的分析 ... 14
3. 結果 ... 15
3.1 呼吸器系由来細胞株に最適なAAV血清型のスクリーニング ... 15
3.2 呼吸器系由来細胞株に対するプロモーターの効果 ... 18
3.3 投与経路についての評価 ... 20
3.4 マウスの肺に対する血清型別AAVベクターの遺伝子導入効果 ... 21
3.5 マウスの肺に対するプロモーターの影響 ... 24
3.6 肺組織および肝臓組織でのAAVベクターの生体分布 ... 26
3.7 肺組織および肝臓組織のmRNA発現解析 ... 28
3.8 肺組織および肝臓組織のルシフェラーゼ量の測定 ... 29
3.9 遺伝子導入された肺組織の評価 ... 32
4. 考察 ... 37
5. 終わりに ... 41
6. 引用文献 ... 42
7. 謝辞 ... 51
1
1. はじめに
嚢胞性肺線維症や特発性肺線維症などの慢性呼吸器疾患は、進行性で有効な 治療法がない。現在これらの疾患に対する治療は、薬物を用いた対症療法や感 染症などによる増悪の予防に限られている。予後は不良であり、嚢胞性肺線維
症の平均生存期間は39.3年で [1]、特発性肺線維症は診断後の平均生存期間が3 年から5年と言われている [2]。一方で、これらの疾患については関連遺伝子の 同定など機構解明が進んでいる。嚢胞性肺線維症では ATP依存性クロライドイ オ ン チ ャ ンネ ル 分 子を コ ー ドし て いる CFTR(cystic fibrosis transmembrane conductance regulator)遺伝子の異常が[3]、特発性肺線維症では粘液タンパク質 をコードする MUC5B 遺伝子の発現調節領域の関与が知られている[4]。このよ うな背景を踏まえ、遺伝子治療はこれらの病態を引き起こす細胞機能を修復し、
病勢を改善する根治療法として、遺伝子治療法の開発への期待が高まっている。
そこで、私は呼吸器細胞に治療遺伝子を導入するために、非病原性ウイルス由 来で安全性が高く、長期に渡って遺伝子発現ができ、広範な組織特異性をもつ アデノ随伴性ウイルス(AAV)ベクターに注目した[5, 6]。これまでに、AAVベ
クターを用いた遺伝子治療の応用では、血友病B[7]やパーキンソン病[8, 9]、先 天性黒内障[10, 11]の臨床試験が行われ、有効性が認められている。一方、呼吸 器領域では嚢胞性肺線維症に対して多くの臨床試験が行われてきたが、気道上
2
皮において一過性かつ低レベルの遺伝子発現が確認されたのみで、治療効果は 認められなかった [12-14]。その理由として、これまでの嚢胞性肺線維症に対す る遺伝子治療研究におけるいくつかの問題点が指摘されている。第一に、広く
用いられてきた血清型すなわち AAV2 型では呼吸器系の細胞に対する遺伝子導 入効率が低い可能性があった。また、これまでに用いられた治療ベクターのプ ロモーター活性が非常に弱く、遺伝子が導入されたとしてもその発現が十分で なかったと考えられる。さらにベクターの投与がネブライザーによるものであ ったため、下気道や肺へ十分到達していなかった可能性が高い。
その後AAVについては多くの血清型が報告され、血清型によって組織特異性 が異なることが知られている。たとえば、脳への遺伝子導入ではAAV2型が[8, 9]、
肝臓に対しては AAV8 型[7]が優れている。呼吸器細胞については、従来広く用 いられてきたAAV2型よりもAAV1型、AAV5型、AAV6型、AAV9型が優れて いるという報告もあるが、詳細には検討されていない[15-18]。呼吸器細胞にお けるプロモーター活性については、cytomegalovirus(CMV)プロモーター[19]、
Rous sarcoma virusプロモーター[20]、CMVエンハンサーにニワトリのβアクチ ンプロモーターを組み合わせた CAG プロモーター[15, 21]などが強い活性を有 し、これらの使用によって導入遺伝子の発現が高まるとも報告されている。投 与経路については、呼吸器上皮細胞における極性とベクターの相性についても
3
考慮に入れなければならないことがわかってきた。すなわち、管腔側(apical)
と側基底膜側(basolateral)からの遺伝子導入で効果が異なるため[22-24]、ベク ターによって投与経路を使い分けることも検討する必要がある。
そこで本研究では、AAV ベクターによる呼吸器細胞への遺伝子導入に関与す る因子を解明し、細胞株およびマウスを用いて最適な遺伝子導入法を確立する ことを目的として実験を行った。
4
2. 実験材料と方法
2.1 AAVベクタープラスミドの作製
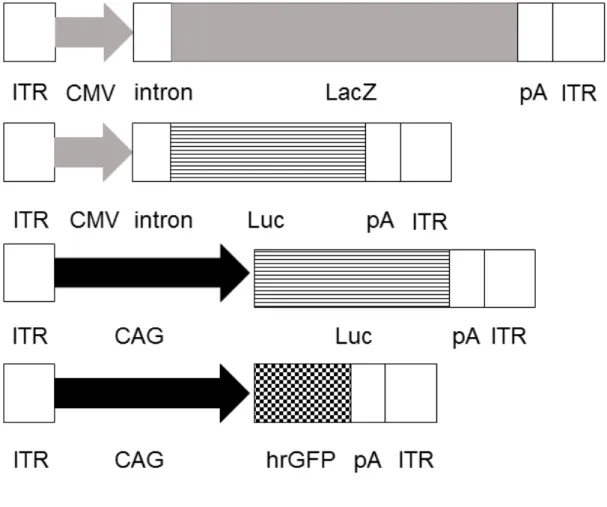
動物を用いた実験に入る前のin vitroスクリーニングのために、大腸菌由来の
β ガラクトシダーゼを発現する LacZ 遺伝子を搭載したプラスミドを構築した
(図 1)。プロモーター活性を比較するためには、ホタルルシフェラーゼ遺伝子
(Luc)を搭載したプラスミドを作製した。pGL3-Control Vector プラスミド
(Promega, Madison, WI, USA)からLuc遺伝子断片を切り出し、pAAV-MCSプ ラスミド(Agilent Technologies, Palo Alto, CA, USA)のマルチクローニングサイ
トに挿入し、これをpCMV-Lucとした。これと比較するため、pCMV-Luc のCMV プロモーターを pCAGGS[25]由来の CAGプロモーターに置き換えて pCAG-Luc を作製した(図 1)。組織学的検討を行う実験に用いるためには、ヒト化ウミシ イタケ由来の緑色蛍光タンパク質(hrGFP)遺伝子を搭載したプラスミドを構築 した。pAAV-hrGFP プラスミド(Agilent Technologies)のCMVプロモーターを CAGプロモーターに置き換えてpCAG-hrGFPを作製した(図1)。
5
図1 各プラスミドベクターの構造図
ITR: inverted terminal repeat、CMV: cytomegalovirus promoter、
pA: polyadenylation sequence、Luc: firefly luciferase、
CAG: chicken β-actin promoter with the CMV immediate-early enhancer、
hrGFP: humanized Renilla reniformis green fluorescent protein
2.2 組み換えAAVベクターの作製
組換えAAVベクターの作製は小倉らの方法[26]によった。2.1で作製したベク
6
タープラスミドとウイルスタンパク質(Rep, Cap)をコードするパッケージング
プラスミド、アデノウイルス由来の E2A、E4、VA を含むヘルパープラスミド
(pHelper; Agilent Technologies)をリン酸カルシウム共沈法でヒト胎児腎細胞293
(HEK293; Agilent Technologies)にトランスフェクションした。72時間培養後、
HEK293細胞を回収して4回凍結・融解を繰り返すことにより組み換えAAVベ
クターを含む粗抽出液を得た。これを 2 度の塩化セシウム密度勾配遠心法で精 製し、AAVベクターのウイルス力価はReal-Time Polymerase Chain Reaction(PCR)
法にて測定した。作製した組み換え AAV ベクターは AAV-CMV-LacZ (1 型か ら9型までの血清型[27-32])、AAV-CAG-Luc (1型、2型、6型、9型)、AAV-CMV-Luc
(6型)、AAV-CAG-hrGFP (6型)である。
2.3 細胞培養
ヒト気管支上皮細胞(BEAS-2B)、マウス肺胞上皮細胞(MLE12)、マウス肺
胞マクロファージ(MH-S)、ヒト胎児肺線維芽細胞(MRC-5)はAmerican Type
Culture Collection(ATCC; Manassas, VA, USA)より購入した。正常ヒト気管支上 皮細胞(NHBE)はLonza(Walkersville, MD, USA)より購入した。BEAS-2Bお よびNHBEは無血清の気管支上皮細胞培地(BEGM; Lonza)にBEGM添加因子 セット(Lonza)を加えて培養した。MLE12 は 10%の非働化牛胎仔血清(FBS;
7
Sigma-Aldrich, St. Louis, MO, USA)及び 1%のペニシリン/ストレプトマイシン
(Pen Strep; Life Technologies, Carlsbad, CA, USA)を含むDulbecco's Modified
Eagle's Medium(DMEM; Life Technologies)培地で培養した。MH-Sは10%のFBS、
1%の Pen Strep に 0.05 mM 2-mercaptoethanol を加えた RPMI1640 培地(Life Technologies)で培養した。MRC-5は10%のFBS、1%のPen Strepを加えたEagle's Minimum Essential Medium(EMEM; ATCC)培地で培養した。HEK293は10%の FBS、1%のPen Strepを加えたDMEM/F-12 medium(Life Technologies)培地で培 養を行った。全ての細胞は37℃、5%二酸化炭素下で培養を行った。
2.4 呼吸器系由来細胞株に対する血清型別 AAV ベクターの遺伝子導入効果の スクリーニング
BEAS-2B、MLE12、MRC-5、MH-S、NHBE をそれぞれ96 穴プレートに1穴 あたり 2×104、4×104、3×104、5×104、1×104個ずつ撒き、24 時間後に LacZ 遺伝 子 を 搭 載 し た AAV1 型 か ら AAV9 型 ま で の 9 種 類 の AAV ベ ク タ ー
(AAV1-9-CMV-LacZ)をそれぞれ1細胞あたり3×105 viral genome(vg)ずつ感 染させた(ただし、MH-Sは1細胞あたり1×106 vgにして感染させた)。72時 間後に(MH-Sは48時間後)、β-Gal Assay Kit(Thermo Fisher Scientific, Waltham,
MA, USA)を用いて β-ガラクトシダーゼの活性を測定した。総蛋白は Protein
8
Assay CBB Solution(Nacalai Tesque, Kyoto, Japan)を用いてBradford法にて測定 し、β-ガラクトシダーゼの活性を蛋白質量で補正した。さらに、72 時間後の細 胞量をCountess Automated Cell Counter(Thermo Fisher Scientific)を用いて測定 し、104個細胞あたりのβ-ガラクトシダーゼの活性量を求めた。本実験は3回繰 り返し、同様の結果を確認した。
2.5 ルシフェラーゼ発現AAVベクターによる細胞株への遺伝子導入と発現
HEK293とBEAS-2Bをそれぞれ96穴プレートに1穴あたり2×104個ずつ撒き、
24時間後にAAV-CAG-Luc(1型、2型、6型、9型)をそれぞれ1細胞あたり3×105 vg ずつ感染させた。48 時間後に発光強度をBright-Glo Luciferase Assay System
(Promega)を用いてFluoroskan Ascent FL(Thermo Fisher Scientific)で測定した。
本実験は3回繰り返し、同様の結果を確認した。
2.6 呼吸器系由来細胞株におけるプロモーターの発現効率の比較
呼吸器系由来細胞株に対するプロモーター別AAVベクターの遺伝子発現効率 を比較するため、5 種類の細胞(BEAS-2B、MLE12、MRC-5、MH-S、NHBE)
をそれぞれ96穴プレートに1穴あたり2×104、4×104、3×104、5×104、1×104個ず つ撒き、24時間後にAAV6-CAG-Lucと AAV6-CMV-Lucをそれぞれ3×105vgず
9
つ感染させた(ただし、MH-Sは1細胞あたり1×106vgにして感染させた)。48 時間後に発光強度を上述と同様に測定した。2.4 と同様に、総蛋白をBradford 法にて測定し、発光強度を蛋白質量で補正した。本実験を 3回繰り返し、結 果の信頼性を確認した。
2.7 動物実験
8~12週齢の C57BL/6Jマウス(Japan SLC, Shizuoka, Japan)を実験に供し た。動物実験は全て自治医科大学動物実験ガイドラインに従って施行した。
投 与 経 路 に よ る AAV ベ ク タ ー の 遺 伝 子 導 入 効 率 を 比 較 す る た め 、
AAV6-CAG-Luc を マ ウ ス に そ れ ぞ れ 静 脈 内 投 与 (1 個 体 あ た り 1×109 vg/150µL)、鼻腔内投与(1個体あたり1×109 vg/50µL)、気管内投与(1個体
あたり 1×109 vg/50µL)で行った。マウスにおける血清型やプロモーターの
違いによる AAV ベクターの遺伝子導入効果を評価するため、AAV-CAG-Luc
(1型、2型、6型、9型)、AAV-CMV-Luc(6型)を気管内投与(1個体あたり
1×109 vg/50µL)で行った。対照となるマウスは AAVベクターの代わりに HN
バッファー(50 mmol l-14-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, pH 7.4, 150 mmol l-1 NaCl)を投与した。
10
2.8 生体イメージング
マウスの腹腔内に100mg/mL D-ルシフェリン溶液(Ieda Chemical, Tokyo, Japan)
をマウス1個体あたり75 mg/kgの量で投与し、生体イメージング装置(IVIS100;
Xenogen, Hopkinton, MA, USA)でルシフェラーゼ由来の発光を測定した。胸部 の発光量を解析ソフトLiving Image software(Xenogen, Alameda, CA, USA)で計 測した。
2.9 組織中のAAVベクターの分布と遺伝子発現の測定
マウスに AAV-CAG-Luc(1 型、2型、6 型、9 型)および AAV6-CMV-Lucの 投与を行い、2ヶ月後に屠殺して肺と肝臓を摘出し、組織中の導入遺伝子のコピ ー数と遺伝子発現(mRNA, ルシフェラーゼ活性)を測定した。DNAはNucleoSpin
Tissue kit(Macherey-Nagel, Düren, Germany)を用いて抽出し、Thermal Cycler Dice Real Time System II(Takara Bio, Shiga, Japan)を用いてquantitative PCR(qPCR)
法によりルシフェラーゼ遺伝子(Luc)のコピー数を測定し、マウスゲノムハプ
ロイドあたり1コピー存在するmouse ribosomal protein large P1 (Rplp1)遺伝子 で 補 正 し た 。qPCR は 抽 出 し た DNA 300 ng を テ ン プ レ ー ト と し て QuantiTectSYBRGreen PCR Kit(Qiagen, Valencia, CA, USA)を用いて測定した。
使用したLucプライマー配列およびマウスRplp1プライマーの配列を以下に示す。
11
Luc forward: 5’- GAAAAAGTTGCGCGGAGGAG-3’
Luc reverse: 5’- CCGCCCTTCTTGGCCTTTAT-3’
Rplp1 forward: 5’- TACGATTTCCACCACCAGCCTTG-3’
Rplp1 reverse: 5’- CTCATTCTCAAGCCATGGACCGT-3’
mRNAの測定は、全RNAをRNeasy Plus Mini Kit(Qiagen)を用いて抽出した。
これを鋳型として逆転写酵素(PrimeScript RT reagent Kit with gDNA Eraser;
Takara Bio)を用いて相補的DNA(cDNA)を作製した。定量的qPCRはSYBR
Premix Ex Taq II(Tli RNaseH Plus; Takara Bio)とThermal Cycler Dice Real Time System II(Takara Bio)を用いて説明書に沿って行った。目的のmRNAはLucで あり、glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH)の発現量を内在 性コントロールとして使用した。Lucの発現量はマウスGAPDHの発現量で補正 した蛍光シグナル値との比で算出した。使用したLucおよびマウスGAPDHのプ ライマー配列を以下に示す。
Luc forward: 5’- CTATGAAGAGATACGCCCTG-3’
Luc reverse-: 5’- TTCGAAGTACTCAGCGTAAG-3’
12
GAPDHforward: 5’- GTTCCAGTATGACTCCACTC-3’
GAPDH reverse: 5’- CCTCACCCCATTTGATGTTA-3’
ルシフェラーゼ活性はPicaGene Luminescence Kit(Toyo Ink, Tokyo, Japan)を用 いて測定した。摘出した臓器の100mgを300-500µLの滅菌水で破砕し、10,000×g、
4℃で 10 分間遠心分離を行い、上清を抽出した。上清と同量の溶解バッファー
(Toyo Ink)を入れ、時々ボルテックスを行いながら、15 分間室温で反応させ
た。その後、混合溶液から96穴プレートに1穴あたり100µLを一対ずつ入れた。
発光基質(Toyo Ink)を100µL入れ、Fluoroskan Ascent FL(Thermo Fisher Scientific)
で発光強度を測定した。ルシフェラーゼ発現量を求めるために、同時にスタン
ダード酵素(Toyo Ink)を10 pg、100 pg、1 ng、10 ngの4点を測定し、検量線 を引いて測定した。なお、本酵素量は上清の総蛋白を Bradford 法にて測定し、
蛋白質量で補正した。
2.10 組織学的検討
AAV ベ ク タ ー の 肺 へ の 遺 伝 子 導 入 を 組 織 学 的 に 評 価 す る た め 、 AAV6-CAG-hrGFPをマウスの気管内に投与した(1個体あたり1×1011 vg/50 µL)。
AAV6-CAG-hrGFPの気管内投与を行った1ヶ月後に屠殺して肺を摘出した。肺
13
組織は気管より 4% パラホルムアルデヒド溶液1mL を注入し、翌日まで4℃で 固定した。その後、氷晶を防止する目的で30% sucrose in 0.1 mol l-1 phosphate
buffer(pH 7.4)に置換し、同様に翌日まで4℃の条件で静置した。そして、凍結
組織切片作製用包埋剤(Tissue-Tek O.C.T Compound; Sakura Finetek, Tokyo, Japan)
で肺を包埋し、クライオスタットで厚さ10µmの凍結切片を作製した。
肺細胞の評価は気管支上皮細胞、肺胞上皮細胞、肺胞マクロファージ、間葉
系細胞について行った。気管支上皮細胞のhrGFPシグナルは蛍光顕微鏡(BX63;
Olympus, Tokyo, Japan)で直接観察を行った。肺胞上皮細胞の免疫組織化学染色
(免疫染色)はマウスモノクローナル抗体の抗サイトケラチン抗体(AE1/AE3,
ab27988; Abcam, Cambridge, MA, USA)を一次抗体に、ビオチン化抗マウスIgG 抗体(M.O.M. Immunodetection Kit; Vector Laboratories, Burlingame, CA, USA)を
二次抗体に用いてRhodamine Avidin D(Vector Laboratories)で蛍光染色を行った。
肺胞マクロファージの免疫染色はラットモノクローナル抗体である抗 F4/80 抗 体(ab6640; Abcam)を一次抗体に、Alexa Fluor 568標識抗ラットヤギポリクロ ーナル抗体(ab175710; Abcam)を二次抗体として行った。間葉系細胞の免疫染
色はウサギポリクローナル抗体である抗α-平滑筋(α-SMA)抗体(ab5694; Abcam)
を一次抗体に、Alexa Fluor 568標識抗ウサギヤギポリクローナル抗体(ab175471;
Abcam)を二次抗体として行った。核は核染色用試薬4’,6-diamidino-2-phenylindole
14
(DAPI)を含む封入剤(VECTASHIELDMounting Medium with DAPI; Vector Laboratories)で封入した。それぞれの免疫染色について、一次抗体を用いない 二次抗体のみを使用した陰性対照では非特異的シグナルは認められなかった。
これらの免疫染色の観察も蛍光顕微鏡(BX63; Olympus)で観察を行った。結果
は1個体あたり3切片を無作為に抽出した複数視野で観察し、200倍の倍率で数 えた。その後、形態学的に評価するためにHematoxyline-Eosin(HE)染色し、脱 水・透徹・封入を行い、光学顕微鏡(BX63; Olympus)で観察した。
2.11統計学的分析
データ解析はStatMate V(ATMS, Tokyo, Japan)とSPSS(IBM Japan, Tokyo, Japan)
を用いて行い、平均値±標準誤差(standard error of the mean: SEM)で記した。独
立 2 群間の比較には Welch の t 検定を用い、多群間比較には一元配置分散分析
(one-way ANOVA)を行い、事後比較にはTukeyまたはGames-Howell法を用い た。P値が0.05未満を有意差ありと判断した。
15
3. 結果
3.1 呼吸器系由来細胞株に最適なAAV血清型のスクリーニング
AAV には様々な血清型があり、血清型によって組織特異性が異なることが知 られているが、気道や肺に最適な血清型は明らかにされてはいなかった。そこ
で、呼吸器細胞に適した血清型を探索するために、βガラクトシダーゼをコード する LacZ を搭 載した 代表的 な AAV の 血 清型 1 型 から 9 型のベ クター
(AAV1-9-CMV-LacZ)を呼吸器系由来細胞株の5種類の細胞(BEAS-2B、MLE12、
MRC-5、MH-S、NHBE)に遺伝子導入した。図 2 に示す通り、全ての呼吸器系
由来細胞株において 6 型の遺伝子導入効率が最も高く、他の血清型による発現 は低かった。また、細胞あたりの発現量に換算しても同様に AAV6 型の遺伝子 導入が最も優れていた(図3)。
16
図2 呼吸器系由来細胞株に対する血清型別AAVベクターの遺伝子導入効果
AAV1-9-CMV-LacZをBEAS-2B、MLE12、MRC-5、MH-S、NHBEにそれぞれ感 染させ(1細胞あたり3×105 vg、MH-Sは1×106 vg)、72時間後(MH-Sは48 時間後)のβガラクトシダーゼの活性量(各 n=4)を測定した。結果は平均値±
標準誤差(SEM)で示した。P <0.05であるものを*で示し, P <0.01は**、P <0.001 は***で示した。
17
図 3 呼吸器系由来細胞株に対する血清型別AAV ベクターの遺伝子導入効果を
細胞数で補正したもの
AAV1-9-CMV-LacZをBEAS-2B、MLE12、MRC-5、MH-S、NHBEにそれぞれ感 染させ(1細胞あたり3×105 vg、MH-Sは1×106 vg)、72時間後(MH-Sは48 時間後)の β ガラクトシダーゼの活性量を測定した。一方で 72 時間後(MH-S は48時間後)の細胞数をカウントし、104細胞あたりのβガラクトシダーゼの活 性量を示した(n=4)。結果は平均値±標準誤差(SEM)で示した。P <0.05であ るものを*で示し, P <0.01は**で示した。
18
3.2 呼吸器系由来細胞株に対するプロモーターの効果
次いで、呼吸器系由来細胞株についてプロモーターによる影響を比較した。
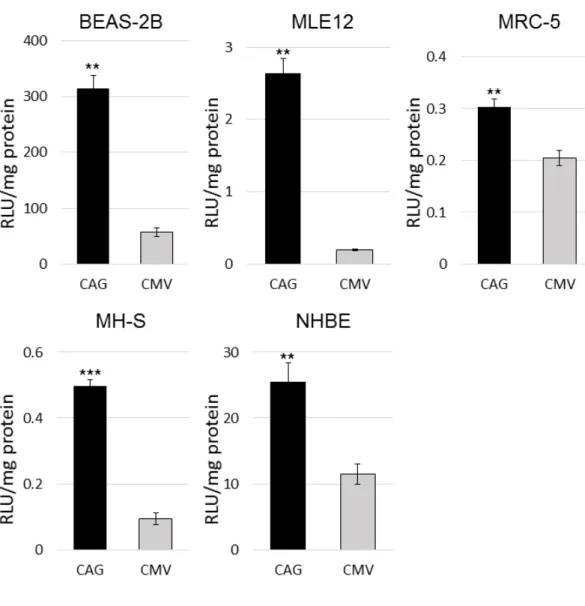
3.1の結果から、呼吸器系由来細胞株にはAAV6型が優れていることが分かった ため、CMV プロモーターとCAGプロモーターを有する AAV6 型のベクターを それぞれ作製し(AAV6-CMV-LucとAAV6-CAG-Luc)、3.1と同様に5種類の細 胞株(BEAS-2B、MLE12、MRC-5、MH-S、NHBE)にそれぞれ感染させた。5 種類の細胞株全てでCAGプロモーターがCMVプロモーターよりも遺伝子発現 を強く認めることが分かった(図4)。
19
図 4 呼吸器系由来細胞株に対するCMV プロモーターとCAG プロモーターの
比較
AAV6-CMV-Luc または AAV-CAG-Luc を BEAS-2B、MLE12、MRC-5、MH-S、
NHBEにそれぞれ感染させ(1細胞あたり3×105 vg、MH-Sは1×106 vg)、48 時間後の発光強度を測定し、蛋白質量で補正した(n=4)。結果は平均値±標準 誤差(SEM)で示した。P <0.01であるものを**で示し、P <0.001は***で示した。
20
3.1および3.2の結果から、次のマウスを用いた実験ではAAV6型およびCAG プロモーターを有するAAVベクターを用いて行った。
3.3 投与経路についての評価
投与経路の違いによる肺への遺伝子導入効果を評価するため、AAV6-CAG-Luc をマウスに静脈内投与、鼻腔内投与、気管内投与を行って比較した。遺伝子発 現を定量的に評価するために生体イメージング装置(IVIS 100)を使用した。
AAV6-CAG-Luc の投与 1 ヶ月後には遺伝子発現を明瞭に確認することができた
(図 5)。静脈内投与では肝臓に発現が限局し、鼻腔内投与では鼻腔に発現が限 局しており、肺での発現は殆ど認められなかった。一方で、気管内投与で行っ た場合は、肺に高い遺伝子発現を認め、気管内投与が肺への遺伝子導入に最適 な投与経路であると分かった。
21
図5 投与経路による比較
AAV6-CAG-Luc をマウスに 3 つの投与経路で投与して 1ヶ月後に生体イメージ
ング装置で評価を行った(1個体あたり1×109 vg)。測定は各 n=4-6で図はその 一部を示す。
3.4 マウスの肺に対する血清型別AAVベクターの遺伝子導入効果
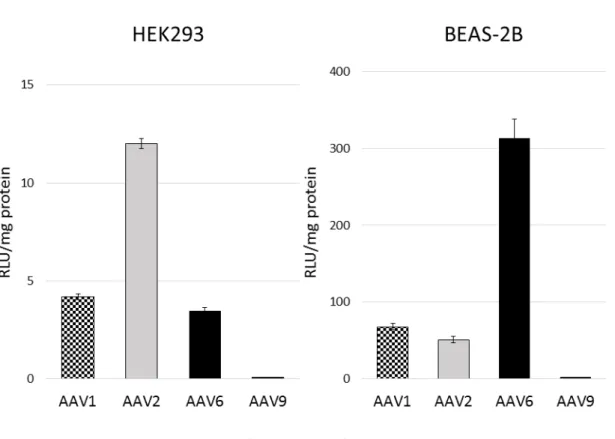
3.3よりAAV6型を用いた場合は肺への遺伝子導入に気管内投与が最適である と分かったが、これまでに広く用いられてきた AAV2 型と肺への遺伝子導入に 適していると報告されてきた AAV1 型および AAV9 型についても気管内投与を 行って検討した。AAV1, 2, 9-CAG-Lucを作製し、マウスに投与する前にHEK293、
BEAS-2B に遺伝子導入を行って発現を行う AAV ベクターであることを確認し
た(図6)。AAV9型のベクターは細胞株では弱い発現しか認めなかったが、AAV1 型およびAAV2型のベクターは発現を確認できた。
22
図6 血清型別AAVベクターの細胞株での遺伝子発現
AAV1, 2, 6, 9-CAG-LucをHEK293またはBEAS-2Bにそれぞれ感染させ、48時 間後の発光強度を測定し、蛋白質量で補正した(n=4)。結果は平均値±標準誤 差(SEM)で示した。
これらのベクター(AAV1, 2, 6, 9-CAG-Luc)をマウスの気管内に投与した。マ
ウスでの遺伝子発現は生体イメージングで評価した(図7a)。AAV1型は肺でわ ずかに発現を認めたが、AAV2型では発現を認めなかった。AAV9型は肝臓で中 程度の発現を認めたが、肺では弱い発現しか認めなかった。また、生体イメー ジング装置および解析ソフトで胸部の発光強度を測定し、経時的に評価した(図 7b)。発光強度のピークは AAV ベクターの投与 1 ヶ月後から 2ヶ月後であり、
23
以降は徐々に低下していった。AAV6型では少なくとも6ヶ月間は比較的高い遺 伝子発現が保たれ、マウスにおける血清型の比較においても AAV6 型が肺での 遺伝子導入に最も優れていることが分かった。
図7 血清型別AAVベクターによるマウスでの遺伝子発現
AAV1, 2, 6, 9-CAG-Luc をマウスの気管内に投与した(それぞれ 1 個体あたり
1×109 vg)。
(a)AAVベクターの投与1ヶ月後に生体イメージング装置で評価し、図はその 一部を示す。
(b)胸部での発光強度を定量化し、経時的に表した(AAV1, 2, 9-CAG-Luc: 各
24
n=4、AAV6-CAG-Luc: n = 6)。結果は平均値±標準誤差(SEM)で示した。P <0.05 であるものを*で示し, P <0.01は**で示した。
3.5 マウスの肺に対するプロモーターの影響
マウスの肺についても遺伝子発現がプロモーターによる影響を受けるか評価
するため、AAV6-CMV-Luc を作製し、マウスに気管内投与して AAV6-CAG-Luc を投与したマウスと比較した。生体イメージング装置で評価したところ、細胞
株の実験結果と同様にCAGプロモーターの方がCMVプロモーターより強い遺 伝子発現を認めた(図8a)。生体イメージング装置および解析ソフトで胸部の発 光強度を定量し経時的に評価したところ、CAGプロモーターは12ヶ月に渡って CMV プロモーターより高い発現を保ち、統計学的にも優位差が確認された。
CAGプロモーターの発現は頑健で、12ヶ月後でもベースラインよりも6.7倍高 い発現を認めていた。一方、CMVプロモーターは8ヶ月後にはベースラインに 戻っていた(図8b)
25
図8 マウスの肺に対するCMVプロモーターとCAGプロモーターの比較
マウスに AAV6-CAG(または CMV)-Luc を気管内投与で行った(それぞれ 1
個体あたり1×109 vg)。
(a)AAVベクターの投与1ヶ月後に生体イメージング装置で評価し、図はその 一部を示す。
(b)胸部での発光強度を定量化し、経時的に表した(AAV6-CAG-Luc: n=6、
AAV6-CMV-Luc: n = 6)。結果は平均値±標準誤差(SEM)で示した。P <0.05で あるものを*で示し, P <0.01は**で示した。
26
3.6 肺組織および肝臓組織でのAAVベクターの生体分布
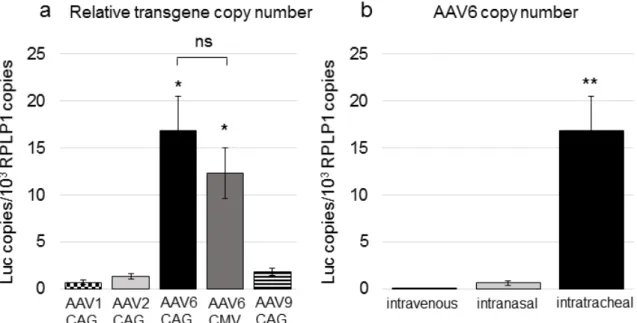
マウスに投与したAAVベクターの生体分布について調べるために、遺伝子発 現のピークにあたる投与 2 ヶ月後に肺と肝臓を摘出した。肺組織と肝臓組織か ら DNAを抽出し、qPCRで導入遺伝子のコピー数を求めた。肺組織における導 入遺伝子のコピー数は、気管内投与した様々な AAV ベクター(AAV1, 2, 6,
9-CAG-LucおよびAAV6-CMV-Luc)で比較すると、AAV6型はAAV1型、AAV2 型、AAV9 型と比較して最も多く認めた(図 9a)。また、AAV6 型ではプロモー ターによる差はなかった(図9a)。投与経路で比較すると、静脈内投与と鼻腔内 投与では肺組織中の導入遺伝子のコピー数は非常に少なく、これらの投与経路
では肺に到達する AAV ベクターの量は少ないことを示唆した(図9b)。なお、
肝臓での導入遺伝子のコピー数は血清型によっても、投与経路によるもので評 価しても、検出されなかった(データ省略)。
27
図9 マウス肺組織中の導入遺伝子のコピー数
(a)マウスにAAV1, 2, 6, 9-CAG-LucおよびAAV6-CMV-Lucの気管内投与を行 い(それぞれ1個体あたり1×109 vg)、投与2ヶ月後に肺組織を摘出し、DNAを 抽出した。ゲノムDNA 300ngあたりのLuc遺伝子のコピー数をqPCR法にて測 定し、マウス Rplp1 遺伝子で補正した。(AAV1, 2, 9-CAG-Luc: 各 n =4、
AAV6-CAG-Luc: n=7、AAV6-CMV-Luc: n = 7)。結果は平均値±標準誤差(SEM)
で示した。P <0.05であるものを*で示した。nsは統計学的に有意差がないこと を示した。
(b)マウスにAAV6-CAG-Lucを静脈内投与、鼻腔内投与、気管内投与を行い(そ れぞれ1個体あたり1×109 vg)、投与2ヶ月後に肺組織を摘出し、DNAを抽出し た。ゲノムDNA 300ngあたりのLuc遺伝子のコピー数をqPCR法にて測定し、
マウスRplp1遺伝子で補正した。(intravenous: n=4、intranasal: n=4、intratracheal:
28
n = 7)。結果は平均値±標準誤差(SEM)で示した。P <0.01であるものを**で
示した。
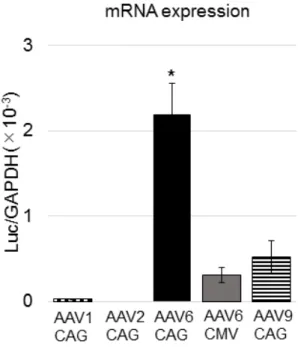
3.7 肺組織および肝臓組織のmRNA発現解析
肺組織および肝臓組織からRNAを抽出して、mRNAの発現解析を行った。気 管 内 投 与を 行 った 様 々な AAV ベ ク タ ー(AAV1, 2, 6, 9-CAG-Luc お よ び
AAV6-CMV-Luc)について肺組織での発現を比較すると、AAV6-CAG-Luc が最
も 強 か っ た ( 図 10)。 プ ロ モ ー タ ー の 比 較 で は 、CAG プ ロ モ ー タ ー
(AAV6-CAG-Luc)はCMVプロモーター(AAV6-CMV-Luc)の7倍強い遺伝子 発現を認めた(図 10)。3.6の結果ではプロモーターの違いによって遺伝子導入 効率自体に違いはなかった(導入遺伝子のコピー数に差はなかった)ことから、
CAG プロモーターは肺組織で強い転写能力をもつことが示された。また、静脈 内投与や鼻腔内投与後の導入遺伝子のコピー数が非常に少なかったことから予
想されたように、肺組織中で導入遺伝子のmRNA は検出されなかった(データ 省略)。なお、肝臓では、どの血清型をどの経路で投与した場合でも、導入遺伝 子のmRNAは検出されなかった(データ省略)。
29
図10 マウス肺組織中のmRNA発現量
マウスにAAV1, 2, 6, 9-CAG-LucおよびAAV6-CMV-Lucの気管内投与を行い(そ れぞれ1 個体あたり1×109 vg)、投与2ヶ月後に肺組織を摘出し、全RNAを抽 出した。mRNA発現量はReverse Transcription PCR(RT-PCR)法にてLucのmRNA 発現量を求めた。GAPDH mRNAを内在性コントロールに使用し、ΔΔCt 法で計 測した(AAV1, 2, 9-CAG-Luc: 各 n=4、AAV6-CAG-Luc: n=7、AAV6-CMV-Luc: n
= 7)。結果は平均値±標準誤差(SEM)で示した。P <0.05であるものを*で示し
た。
3.8 肺組織および肝臓組織のルシフェラーゼ発現量の測定
肺組織および肝臓組織中のルシフェラーゼ活性を測定した。肺組織では、気
30
管 内 投 与 し た 様 々 な AAV ベ ク タ ー (AAV1, 2, 6, 9-CAG-Luc お よ び
AAV6-CMV-Luc)で比較すると、mRNAの解析結果に一致して AAV6-CAG-Luc
が最も多い酵素量を示した(図11a)。プロモーターで比較すると、CAGプロモ ーター(AAV6-CAG-Luc)はCMVプロモーター(AAV6-CMV-Luc)の14.4倍多 い酵素量を示した(図 11a)。静脈内投与や鼻腔内投与による肺組織中の酵素活 性はmRNA解析の結果と同様に検出されなかった(データ省略)。肝臓組織のル シフェラーゼの発現についてはAAV6-CAG-Lucの静脈内投与とAAV9-CAG-Luc の気管内投与の場合にて非常に弱い酵素活性を認めたが、他の条件では検出さ
れなかった(図 11b)。ルシフェラーゼ発現については、生体イメージングによ る遺伝子発現(図 5、図 7)は酵素活性を反映しているのに対し、図 11 は酵素 蛋白量を表している。そのため、両者の間に必ずしも比例関係が成り立つとは 限らない。さらに、肝臓では全体の蛋白量が多いため、補正によりルシフェラ
ーゼ発現量が過小評価されている可能性もある。この点も含め、AAV6-CAG-Luc の静脈内投与とAAV9-CAG-Lucの気管内投与後の体内動態については今後の検 討課題である。
31
図11 マウス肺組織および肝臓組織のルシフェラーゼ発現量
(a)マウスにAAV1, 2, 6, 9-CAG-LucおよびAAV6-CMV-Lucの気管内投与を行 い(それぞれ1個体あたり1×109 vg)、投与2ヶ月後に肺組織を摘出し、ルシフ ェラーゼ発現量を測定し、蛋白質量で補正した(AAV1, 2, 9-CAG-Luc: 各 n=4、
AAV6-CAG-Luc: n=7、AAV6-CMV-Luc: n = 7)。結果は平均値±標準誤差(SEM)
で示した。P <0.001を***で示した。
(b)マウスに AAV6-CAG-Luc の静脈内投与、AAV9-CAG-Luc の気管内投与を 行ったもので(それぞれ1個体あたり1×109 vg)、投与2ヶ月後の肝臓組織のル シフェラーゼ発現量を示す(AAV6-CAG-Luc: n=4、AAV9-CAG-Luc: n=4)。結 果は平均値±標準誤差(SEM)で示した。
32
3.9 遺伝子導入された肺組織の評価
上記の結果から、肺への遺伝子導入にはAAV6-CAGベクターの気管内投与が 最適であることが明らかになった。そこで、AAV6-CAG ベクターが肺のどの細 胞に遺伝子導入しているかを評価するため、AAV6-CAG-hrGFP をマウスに気管
内投与で遺伝子導入を行った。AAV ベクターの投与 1ヶ月後に屠殺して肺の凍 結切片を作製し、免疫染色などを行って蛍光顕微鏡で評価した(図 12a-d)。肺 全体で評価すると、遺伝子導入された細胞は 14.5%あった(図 12e)。蛍光顕微
鏡で直接 hrGFP シグナルを観察すると、気管支上皮細胞の hrGFP 陽性割合は
12.8%であった(図12a, f)。サイトケラチン、F4/80、α-SMAの抗体を用いて免 疫染色を行うと、サイトケラチン陽性の肺胞上皮細胞に hrGFP シグナルを伴う 割合は6.3%(図12b, f)、F4/80陽性マクロファージにhrGFPシグナルを伴う割
合は1.5%(図12c, f)と非常に少なく、これに対し、α-SMA陽性細胞にhrGFP
シグナルを伴う割合は 67.3%(図12d, f)と非常に高かった。HE 染色による形 態学的評価を行ったところ、炎症所見はなく、hrGFP陽性のα-SMA陽性細胞は 肺胞中隔の毛細血管周囲に殆ど存在し(図 13)、AAV6-CAG ベクターは周皮細 胞を主に遺伝子導入することが明らかになった。
33
図12 AAVベクターにより遺伝子導入された肺組織の評価
マウスに AAV6-CAG-hrGFP の気管内投与を行い(1 個体あたり 1×1011 vg)、投 与1ヶ月後に肺の凍結切片を作製した(n=4)。1個体あたり3切片を評価し、図 はその一部を示す。
34
(a)気管支上皮細胞を蛍光顕微鏡で直接観察した。
(b-d)免疫染色を行った。(b)抗サイトケラチン抗体、(c)抗F4/80抗体、(d)
抗α-SMA抗体。
矢尻は遺伝子導入された細胞を示す。
(e)肺組織全体のhrGFP陽性細胞の割合を示す。(f)気管支上皮細胞および各 免疫染色陽性細胞に対する hrGFP 陽性細胞の割合を定量した。結果は平均値±
標準誤差(SEM)で示した。
35
36
図13 α-SMA陽性細胞の評価
マウスに AAV6-CAG-hrGFP の気管内投与を行い(1 個体あたり 1×1011 vg)、投 与1ヶ月後に肺の凍結切片を作製し(n = 4)、抗α-SMA抗体による免疫染色な らびにHE染色を行った。図は同一切片のもので、代表的な組織像を示す。
(a, c)蛍光顕微鏡での観察像。
(b)HE染色を行い、光学顕微鏡で観察を行った。
矢尻はα-SMA陽性細胞を示す。
37
4. 考察
私は、本研究においてAAVベクターによる呼吸器細胞への最適な遺伝子導入 法を確立した。呼吸器細胞への遺伝子導入に最も期待できる AAV ベクターは
AAV6とCAGプロモーターの組み合わせであり、投与方法としては気管内投与 法が呼吸器細胞への遺伝子導入に最も優れていた。さらに、AAV6-CAG ベクタ ーの気管内投与はマウスの肺で長期にかつ高い発現を維持し、組織学的には周 皮細胞を主に遺伝子導入することが明らかになった。
これまでにも、呼吸器細胞への遺伝子導入は AAV1 型、AAV5 型、AAV6 型、
AAV9型が効果的であると言われてきた[15-18]。私は呼吸器細胞の遺伝子導入に 最も有効な血清型を探索するために、呼吸器系由来細胞株やマウス実験により
代表的なAAVの血清型を用いて評価した。呼吸器系由来細胞株を用いた実験で は、AAV6型の遺伝子導入効果が最も優れていた。マウスを用いた実験でも、Luc 遺伝子を搭載した数種類のAAVベクターをマウスに気管内投与して生体イメー ジングで評価したところ、AAV6 型が肺の遺伝子導入に最も優れていた。AAV6 型が肺への遺伝子導入において優れている機序については、明らかにされては
いない部分が多い。AAV6型は細胞表面のN結合型糖鎖であるα-2, 3結合または
α-2, 6結合シアル酸、およびヘパラン硫酸プロテオグリカンを遺伝子導入の際に
用いていると言われている[33, 34]。これらのシアル酸は呼吸器上皮細胞の管腔
38
側 (apical) に 多 く 存 在 し 、 ヘ パ ラ ン 硫 酸 プ ロ テ オ グ リ カ ン は 側 基 底 膜 側
(basolateral)に多く存在する。このように、レセプター分子が呼吸器上皮細胞 に多く発現しているため、AAV6 型は呼吸器細胞に親和性が高いと推測される。
また、遺伝子導入後の発現はプロモーター活性の強さに影響される[15, 19-21]。
CAG プロモーターは動物実験で長期の遺伝子発現を導く強い活性をもつプロモ ーターの一つであるが[15]、本研究のマウス肺においても長期かつ強い遺伝子発
現を認めた。CAGプロモーターは比較的サイズが大きく(約 1.7kb)、パッケー ジできる遺伝子サイズが限られるAAVベクター(~4.5kb)には使用しづらいと いう問題点はあるが、機能ドメインを残して治療用遺伝子を切り詰めるなど、
工夫の余地はある。例えば、CFTR のようにサイズの大きな遺伝子を短くした
cDNAを合成する方法が試みられ、細胞株でも[35]動物においても有効な機能を 示した研究もある[36, 37]。
実際の臨床応用を考えた時、少ない副作用で十分な効果を発揮することが望 まれ、標的臓器に特異的な投与を行うことが重要である。マウスに対する気管 内投与の侵襲性は比較的高いという議論もあるが[38]、気管支鏡は実臨床におい てしばしば用いられる検査手技であり、容易かつ確実に気管支上皮細胞や肺胞 上皮細胞にベクターを投与することができる。また、投与方法を検討する上で は、細胞極性についても考慮しなければならない。同じAAVベクターでも管腔
39
側(apical)から投与されるのと、側基底膜側(basolateral)から投与されるので
は遺伝子導入効果が異なる。嚢胞性肺線維症の臨床試験で使用された AAV2 型 は気道上皮細胞の側基底膜側(basolateral)からの遺伝子導入に優れてはいるが
[23]、ネブライザーを用いて管腔側から投与された場合の効果については疑問が ある。一方、AAV6型は管腔側(apical)からの遺伝子導入に優れている[24]と報
告されており、これを裏付けるように、本研究でも、AAV6型は気管内投与によ って気道上皮細胞に優れた遺伝子導入を示した。細胞極性とAAV血清型の特異 的な関係を考慮して、今後も適切な投与方法を検討する必要がある。
さらに、AAV6 型は再投与が可能であるという点で有利と考えられる。AAV2
型の再投与は成功していないが[14]、AAV5型やAAV6型ではin vivoで遺伝子発 現が再発現しており[39, 40]、初回投与後の発現低下にも対応できる可能性があ り期待される。
最後に、AAV ベクターが肺のどの細胞に遺伝子導入を行っているか、殆ど組 織学的に検討された例がなく、検討されたものでも主に評価されてきたのは気 管支上皮細胞だった[21, 24]。本研究では、蛍光蛋白マーカー(hrGFP)と免疫染 色を併用することで気管支上皮細胞と末梢の肺細胞について詳細に検討し、気 管支上皮細胞だけでなく、α-SMA 陽性細胞に多く遺伝子導入されていることを 明らかにした。成熟マウスの肺では、α-SMA は血管や気管支の平滑筋細胞だけ
40
でなく、alveolar ring cellと呼ばれる肺胞の平滑筋細胞(肺胞筋線維芽細胞とも 呼ばれる[41])と周皮細胞にも発現している。Alveolar ring cellは肺胞管の入口 部に多く存在し、周皮細胞は肺胞中隔の毛細血管周囲に存在している。今回の
マウス肺の検討では、炎症所見はなく、hrGFP と αSMA 陽性の共局在は肺胞管 入口部ではなく肺胞中隔の毛細血管周囲に認めたことから、AAV6により遺伝子 導入された肺実質の細胞は主に周皮細胞と考えられた。周皮細胞は毛細血管や 血管に接するある種の間葉系細胞由来の壁細胞で[42]、微小な血流を変化させる ことで肺実質の形態や機能を維持している[43]。AAV6型が周皮細胞へ遺伝子導 入する機序については今後の検討事項であるが、肺胞上皮細胞内を通過してい
る (transcellular transport)、 ま た は 上 皮 間 の 密 結 合 の 間 を 通 過 し て い る
(paracellular transport)などの可能性が考えられる[44, 45]。興味深いことに、周 皮細胞は筋線維芽細胞の前駆細胞とも考えられている[46]。筋線維芽細胞は持続 活性化と増殖によって過剰な線維性細胞外基質を産生することから、肺線維症 の発症に重要な因子となっている[46]。従って、周皮細胞への遺伝子導入は肺線 維症などの治療標的になるとも考えられる。
以上、本研究結果からAAV6-CAGベクターはin vitro(細胞株)でもin vivo(マ ウス)でも呼吸器細胞に優れた遺伝子導入を図ることが明らかになった。これ を気管内投与すると、気管支上皮細胞や周皮細胞が標的となるため、嚢胞性肺
41
線維症や特発性肺線維症など難治性肺疾患に対して大きな治療効果を発揮でき ると期待される。
5. 終わりに
本研究により、AAV ベクターによる呼吸器細胞への最適な遺伝子導入法を確 立できた。特に、マウス肺で長期に渡って遺伝子発現を確認できたこと、そし て、AAV6-CAG ベクターの気管内投与は気管支上皮細胞だけでなく、多くの周 皮細胞に遺伝子導入されることを明らかにしたことは今後の呼吸器の遺伝子治 療に大いに貢献できるものと考えられる。
今後は、今回確立した手法を用いて、疾患モデル動物に治療遺伝子を搭載し
たAAVベクターを投与して、有効な治療を示すか検討していきたいと考えてい る。AAV6-CAG ベクターは周皮細胞を主に遺伝子導入し、この周皮細胞は筋線 維芽細胞の前駆細胞と考えられていることから、現在肺線維症モデルに対する 治療実験を計画している。AAV6-CAG ベクターを利用した治療戦略が難治性呼 吸器疾患への遺伝子治療の発展につながることを期待している。
42
6. 引用文献
1.Cystic Fibrosis Foundation. Cystic Fibrosis Foundation Patient Registry 2014 Annual Data Report; 2015 [Bethesda, Maryland]
2. Bjoraker JA, Ryu JH, Edwin MK, Myers JL, Tazelaar HD, Schroeder DR, Offord KP. Prognostic significance of histopathologic subsets in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 157: 199-203, 1998.
3. Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL, Drumm ML, Iannuzzi MC, Collins FS, Tsui
LC.Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science. 245: 1066-1073, 1989.
4.Seibold MA, Wise AL, Speer MC, Steele MP, Brown KK, Loyd JE, Fingerlin TE, Zhang W, Gudmundsson G, Groshong SD, Evans CM, Garantziotis S, Adler KB, Dickey BF, du Bois RM, Yang IV, Herron A, Kervitsky D, Talbert JL, Markin C, Park J, Crews AL, Slifer SH, Auerbach S, Roy MG, Lin J, Hennessy CE, Schwarz MI, Schwartz DA. A common MUC5B promoter polymorphism and pulmonary fibrosis.
N Engl J Med. 364: 1503-1512, 2011.
5. Mueller C, Flotte TR. Clinical gene therapy using recombinant adeno-associated virus vectors. Gene Ther 15: 858–863, 2008.
43
6. Asokan A, Schaffer DV, Samulski RJ. The AAV vector toolkit: poised at the clinical crossroads. Mol Ther .20: 699-708, 2012.
7. Nathwani AC, Reiss UM, Tuddenham EG, Rosales C, Chowdary P, McIntosh J, Della Peruta M, Lheriteau E, Patel N, Raj D, Riddell A, Pie J, Rangarajan S, Bevan D, Recht M, Shen YM, Halka KG, Basner-Tschakarjan E, Mingozzi F, High KA, Allay J, Kay MA, Ng CY, Zhou J, Cancio M, Morton CL, Gray JT, Srivastava D, Nienhuis AW, Davidoff AM. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N Engl J Med. 371: 1994-2004, 2014.
8. Muramatsu S, Fujimoto K, Kato S, Mizukami H, Asari S, Ikeguchi K, Kawakami T, Urabe M, Kume A, Sato T, Watanabe E, Ozawa K, Nakano I. A phase I study of aromatic L-amino acid decarboxylase gene therapy for Parkinson's disease. Mol Ther.
18: 1731-1735, 2010.
9. LeWitt PA, Rezai AR, Leehey MA, Ojemann SG, Flaherty AW, Eskandar EN, Kostyk SK, Thomas K, Sarkar A, Siddiqui MS, Tatter SB, Schwalb JM, Poston KL, Henderson JM, Kurlan RM, Richard IH, Van Meter L, Sapan CV, During MJ, Kaplitt MG, Feigin A.AAV2-GAD gene therapy for advanced Parkinson's disease: a double-blind, sham-surgery controlled, randomized trial.Lancet Neurol. 10: 309-319, 2011.
44
10. Ashtari M, Cyckowski LL, Monroe JF, Marshall KA, Chung DC, Auricchio A Simonelli F, Leroy BP, Maguire AM, Shindler KS, Bennett J. The human visual cortex responds to gene therapy-mediated recovery of retinal function. J Clin Invest. 121:
2160-2168, 2011.
11. Testa F, Maguire AM, Rossi S, Pierce EA, Melillo P, Marshall K Banfi S, Surace EM, Sun J, Acerra C, Wright JF, Wellman J, High KA, Auricchio A, Bennett J, Simonelli F.Three-year follow-up after unilateral subretinal delivery of adeno-associated virus in patients with Leber congenital Amaurosis type 2.
Ophthalmology. 120: 1283-1291, 2013.
12. Aitken ML, Moss RB, Waltz DA, Dovey ME, Tonelli MR, McNamara SC,Gibson RL, Ramsey BW, Carter BJ, Reynolds TC.A phase I study of aerosolized administration of tgAAVCF to cystic fibrosis subjects with mild lung disease. Hum Gene Ther. 12:
1907-1916, 2001.
13. Moss RB, Rodman D, Spencer LT, Aitken ML, Zeitlin PL, Waltz D, Milla C, Brody AS, Clancy JP, Ramsey B, Hamblett N, Heald AE.Repeated adeno-associated virus serotype 2 aerosol-mediated cystic fibrosis transmembrane regulator gene transfer to the lungs of patients with cystic fibrosis: a multicenter, double-blind, placebo-controlled trial. Chest. 125: 509-521, 2004.
45
14. Moss RB, Milla C, Colombo J, Accurso F, Zeitlin PL, Clancy JP, Spencer LT, Pilewski J, Waltz DA, Dorkin HL, Ferkol T, Pian M, Ramsey B, Carter BJ, Martin DB, Heald AE.Repeated aerosolized AAV-CFTR for treatment of cystic fibrosis: a randomized placebo-controlled phase 2B trial. Hum Gene Ther. 18: 726–732, 2007.
15. Virella-Lowell I, Zusman B, Foust K, Loiler S, Conlon T, Song S, Chesnut KA, Ferkol T, Flotte TR. Enhancing rAAV vector expression in the lung. J Gene Med. 7:
842-850, 2005.
16. Gao G, Vandenberghe LH, Alvira MR, Lu Y, Calcedo R, Zhou X, Wilson JM.
Clades of Adeno-associated viruses are widely disseminated in human tissues. J Virol.
78: 6381-6388, 2004.
17. Halbert CL, Allen JM, Miller AD. Adeno-associated virus type 6 (AAV6) vectors mediate efficient transduction of airway epithelial cells in mouse lungs compared to that of AAV2 vectors. J Virol. 75: 6615-6624, 2001.
18. Flotte TR, Fischer AC, Goetzmann J, Mueller C, Cebotaru L, Yan Z, Wang L, Wilson JM, Guggino WB, Engelhardt JF. Dual reporter comparative indexing of rAAV pseudotyped vectors in chimpanzee airway. Mol Ther. 18: 594-600, 2010.
19. Song S, Morgan M, Ellis T, Poirier A, Chesnut K, Wang J, Brantly M, Muzyczka N, Byrne BJ, Atkinson M, Flotte TR.Sustained secretion of human alpha-1-antitrypsin
46
from murine muscle transduced with adeno-associated virus vectors. Proc Natl Acad Sci USA. 95: 14384-14388, 1998.
20.Sirninger J, Muller C, Braag S, Tang Q, Yue H, Detrisac C, Ferkol T, Guggino WB, Flotte TR. Functional characterization of a recombinant adeno-associated virus 5-pseudotyped cystic fibrosis transmembrane conductance regulator vector. Hum Gene Ther. 15: 832-841, 2004.
21. Halbert CL, Lam SL, Miller AD. High-efficiency promoter-dependent transduction by adeno-associated virus type 6 vectors in mouse lung. Hum Gene Ther.
18: 344-354, 2007.
22. Yan Z, Lei-Butters DC, Liu X, Zhang Y, Zhang L, Luo M, Zak R, Engelhardt JF.
Unique biologic properties of recombinant AAV1 transduction in polarized human airway epithelia. J Biol Chem. 281: 29684-29692, 2006.
23. Duan D, Yue Y, Yan Z, McCray PB Jr, Engelhardt JF. Polarity influences the efficiency of recombinant adenoassociated virus infection in differentiated airway epithelia. Hum Gene Ther. 9: 2761-2776, 1998.
24. Limberis MP, Vandenberghe LH, Zhang L, Pickles RJ, Wilson JM. Transduction efficiencies of novel AAV vectors in mouse airway epithelium in vivo and human ciliated airway epithelium in vitro. Mol Ther. 17: 294-301, 2009.
47
25. Niwa H, Yamamura K, Miyazaki J.Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene. 108: 193-199, 1991.
26. Ogura M, Urabe M, Akimoto T, Onishi A, Ito C, Ito T, Tsukahara T, Mizukami H, Kume A, Muto S, Kusano E, Ozawa K.et al. Interleukin-10 expression induced by adeno-associated virus vector suppresses proteinuria in Zucker obese rats. Gene Ther.
19: 476-482, 2012.
27. Mochizuki S, Mizukami H, Kume A, Muramatsu S, Takeuchi K, Matsushita T, Okada T, Kobayashi E, Hoshika A, Ozawa K. Adeno-associated virus (AAV)
vector-mediated liver- and muscle-directed transgene expression using various kinds of promoters and serotypes. Gene Ther Mol Biol. 8: 9-18, 2004.
28. Muramatsu S, Mizukami H, Young NS, Brown KE. Nucleotide sequencing and generation of an infectious clone of adeno-associated virus 3. Virology. 221: 208-217, 1996.
29. Chiorini JA, Yang L, Liu Y, Safer B, Kotin RM. Cloning of adeno-associated virus type 4 (AAV4) and generation of recombinant AAV4 particles. J Virol. 71:
6823-6833, 1997.
30. Chiorini JA, Kim F, Yang L, Kotin RM. Cloning and characterization of adeno-associated virus type 5. J Virol. 73: 1309-1319, 1999.
48
31. Rutledge EA, Halbert CL, Russell DW. Infectious clones and vectors derived from adeno-associated virus (AAV) serotypes other than AAV type 2. J Virol. 72:
309-319, 1998.
32. Gao GP, Alvira MR, Wang L, Calcedo R, Johnston J, Wilson JM. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy.
Proc Natl Acad Sci USA. 99: 11854-11859, 2002.
33. Wu Z, Miller E, Agbandje-McKenna M, Samulski RJ. Alpha2,3 and alpha2,6 N-linked sialic acids facilitate efficient binding and transduction by adeno-associated virus types 1 and 6. J Virol. 80: 9093-9103, 2006.
34. Ng R, Govindasamy L, Gurda BL, McKenna R, Kozyreva OG, Samulski RJ, Parent KN, Baker TS, Agbandje-McKenna M. Structural characterization of the dual glycan binding adeno-associated virus serotype 6. J Virol. 84: 12945-12957, 2010.
35. Ostedgaard LS, Rokhlina T, Karp PH, Lashmit P, Afione S, Schmidt M, Zabner J, Stinski MF, Chiorini JA, Welsh MJ. A shortened adeno-associated virus expression cassette for CFTR gene transfer to cystic fibrosis airway epithelia. Proc Natl Acad Sci USA. 102: 2952-2957, 2005.
36. Fischer AC, Smith CI, Cebotaru L, Zhang X, Askin FB, Wright J, Guggino SE, Adams RJ, Flotte T, Guggino WB. Expression of a truncated cystic fibrosis
49
transmembrane conductance regulator with an AAV5-pseudotyped vector in primates.
Mol Ther. 15: 756-763, 2007.
37. Mueller C, Torrez D, Braag S, Martino A, Clarke T, Campbell-Thompson M, Flotte TR. Partial correction of the CFTR-dependent ABPA mouse model with recombinant adeno-associated virus gene transfer of truncated CFTR gene. J Gene Med. 10: 51-60, 2008.
38. Liqun Wang R, McLaughlin T, Cossette T, Tang Q, Foust K, Campbell-Thompson M, Martino A, Cruz P, Loiler S, Mueller C, Flotte TR. Recombinant AAV serotype and capsid mutant comparison for pulmonary gene transfer of alpha-1-antitrypsin using invasive and noninvasive delivery. Mol Ther. 17: 81-87, 2009.
39. Vidović D, Gijsbers R, Quiles Jimenez A, Dooley J, Van den Haute C, Van derPerren A, Liston A, Baekelandt V, Debyser Z, Carlon MS. Noninvasive Imaging Reveals Stable Transgene Expression in Mouse Airways After Delivery of a Nonintegrating Recombinant Adeno-Associated Viral Vector. Hum Gene Ther. 27:
60-71, 2016.
40. Halbert CL, Rutledge EA, Allen JM, Russell DW, Miller AD. Repeat transduction in the mouse lung by using adeno-associated virus vectors with different serotypes. J Virol. 74: 1524-1532, 2000.
50
41. 妹尾春樹, 佐藤充, 今井克幸.肝臓星細胞と肺胞中隔細胞. 呼吸. 16:
604-615, 1997.
42. Armulik A, Genové G, Betsholtz C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell. 21: 193-215, 2011.
43. Dickie R, Wang YT, Butler JP, Schulz H, Tsuda A. Distribution and quantity of contractile tissue in postnatal development of rat alveolar interstitium.Anat Rec (Hoboken). 291: 83-93, 2008.
44. Di Pasquale G, Chiorini JA. AAV transcytosis through barrier epithelia and endothelium. Mol Ther. 13: 506-516, 2006.
45. Patton JS. Mechanisms of macromolecule absorption by the lungs. Adv Drug Deliv Rev. 19: 3-36, 1996.
46. Hung C, Linn G, Chow YH, Kobayashi A, Mittelsteadt K, Altemeier WA, Gharib SA, Schnapp LM, Duffield JS. Role of lung pericytes and resident fibroblasts in the pathogenesis of pulmonary fibrosis. Am J Respir Crit Care Med. 188: 820-830, 2013.
47. Hutchison N, Fligny C, Duffield JS. Resident mesenchymal cells and fibrosis.
Biochim Biophys Acta. 1832: 962-971, 2013.
51
7. 謝辞
本研究を遂行するにあたり、自治医科大学免疫遺伝子細胞治療学(タカラバ イオ)講座の内堀亮介講師、臨床研究支援センターの久米晃啓教授、前呼吸器 内科教授の杉山幸比古先生には多大なるご指導ご弁鞭撻を頂き、心より感謝い たします。
また、本研究は自治医科大学分子病態治療研究センター遺伝子治療研究部なら びに内科学講座呼吸器内科学講座で遂行させて頂き、皆様からたくさんのご支 援、ご協力を頂きました。厚くお礼申し上げます。