実験てんかん(Kind1ing)の生理心理学的研究ー扁桃
核Kindlingの永続性に及ぼす海馬損傷効果(注)ー
著者
吉田 和典
雑誌名
福井医科大学一般教育紀要
巻
3
ページ
25-44
発行年
1983-12
URL
http://hdl.handle.net/10098/5319
福井医科大学一般教育紀要 第
3
号(
1
9
8
3
)
実験てんかん
(
K
i
n
d
l
i
n
g
)
の生理心理学的研究
一 肩 桃 核
Kindling
の永続性に及ぼす海馬損傷効果(注)一
土 口田
和
典 心理学教室 (昭和58年10月14日 受 理 )序および目的
Kindling(燃え上がり)現象とは,脳のある部位(特に大脳辺縁系の中で肩桃核)を一定の 刺激条件のもとに低強度で毎日一回刺激を続けると,最初はその刺激に対して無反応であった ものが徐々にてんかん様反応(顔面措揚や点頭など)が増強し,最終的に全身けいれん発作が 出現してくるようになることを言う(8)四(34)0 この言葉は19
6
9
年Goddardら(8)によって初めて文 献上に記載され,それ以来Kindling現象を用いて,生理心理学的には脳神経の可塑性(例えば 記憶や学習など)の実験モデル(7)(8)(1抑 0)(21)仰として,また神経精神医学的にはてんかんの実験モ デル四四倒)としてそのメカニズムを探る数多くの研究が行なわれているO本現象の主な研究観点、 はその発現過程に置かれており,今まで得られた成果をまとめると,神経機序として大脳辺縁 系聞の神経結合の増強回)や辺縁系一脳幹部位聞の結合増強(14)(32)(33)などが指摘され,それらが運動 系を興奮させると考えられている口すなわち刺激による経シナプス性変化として本現象をとら えることができる(190一方,神経薬理学や神経化学的立場から脳内モノアミン系の変化,特に カテコラミンがKindlingの発展過程で徐々に減少する(印刷ことから本現象に対してこの系は抑制 的に僧Iいていることが言われてきた(5四
九
Kindling現象は-j!形成されると永久的に持続し,もはや電気刺激を与えなくても発作が自 発的に生Uるようになる側(31)ことが報告されている口従って最近では完成したKindling現象の 永続性に関する研究に注意が向けられている。臨床的には種々の抗てんかん薬の作用効果が評 価されている(18)が,基礎的には電気生理学及び解剖学的手法を用いて脳の様々の部位の刺激あ るいは破壊による本現象の動態をより厳密にとらえる方向に関心が集められている倒。 WadaとSato(33)はKindlingの完成したネコを用いて中脳網様体 (Midbrain Reticular Formation ,.
MRF) を破壊すると,本現象の再出現が全般的に抑制されることを報告し,このMRFが専ら Kindling現象の永続性のメカニズムに係っていることを示唆した口一方,以前からMRFは 大 脳辺縁系の一部である海馬 (Hippocampus)と密接に結合していることが形態学的(4)(間にある いは機能的(1)(19)にも言われてきた。すなわち, MRFは視床下部(1)や中隔野(19)を通じて海馬に影 F h d つ ﹄
響を及ぼしており,海馬がむしろ行動統制(2)(3)(9)や感覚情報処理(羽(36)に対して重要な役制を果し ているごとが示唆されているoRacine側と森分ら(16)によって海馬破壊実験からKindling現象の 発展過程を統制しているのが海馬であることを示唆するデータがいくつか報告されているo し かしながら今だ統ーした見解には達しておらず,またKindling現象の永続性に対する海馬の働 きに関する報告は全くな¥,)0 従って本実験では, Kindling現象完成後,両側の海馬を破壊しそ の後の本現象の動態を行動的,脳波的に厳密に観察し, Kindling現象の永続性に及ぼす海馬の 働きについて検討した口特に今回は海馬を背側と腹側部位に分けて別々に破壊し,海馬内の部 位的差異を吟昧した。
方法
(1)被 験 体
Wistar-lmamichi系雄ラット10匹を用い、うち7匹において海馬破壊を行なった。手術時の 体重は300~490g で、実験前及び実験中餌水は ad libに与え飼育された。 (2)手郁
I
ネンブタール (Pentobarbital Sodium, 35mg / kg,腹腔内投与)麻酔下で脳定位固定装置に 頭部を固定し電極植込み手術を行なった。扇桃核刺激用電極は先端約O.5mmを 除 い て ガ ラ ス で 絶縁されたステンレス線(直径約O.2mm)を2本貼り合わせたもの(電極間距離約1mm)を用い, 右肩桃核群の外側基底核 (basolateralnucleus)に刺入した。海馬脳波導出用及び破壊用電 極は同ビステンレス線を左右の背側海馬 (Dorsal Hippocampus,.DHPC)と腹側海馬 (Ve-ntral Hippocampus,.VHPC)にそれぞれ1本ずつ刺入した。また同じ電極をそれぞれの海馬 上の大脳皮質に刺入(皮質上から約1mm)し、コントロール破壊用として用いた。電極刺入部位はKδnigとKlippelの脳図譜(11)に従い,扇桃核はLambdaよりAnterior (A)ニ4.89mm,正中
線より右Lateral (L) =4.8mm, Horizontal zero planeよりVertical (V) = -3 .7mm, DH PC はA=4 .23mm, L =::!:2 .5mm, V =+2 .4mm, V H P C はA=2 .97mm, L = 土4.4mm,V = -1.8mmにそれぞれ刺入したO 他に皮質脳波記録用として前頭部 (Bregmaより前方1mm,外側 に3mm).後頭部 (Lambdaより外側3mm)上の頭蓋及び鼻骨(基準電極用)にステンレスネジ を植込み,すべての電極を10ピンミニチュアソケットに半田付けし歯科用セメントで頭蓋に固 定した。
(
3
)
手続
子術から l週間の回復期の後,脳波記録及び刺激用ケージに慣化させると同時に予備的脳波 記録を行なった口その際,扇桃核Kindling刺激強度を決定した。 RacineによりKindling現象の 完成には後発射 (Afterdischarge・,AD)の出現が必要である凹)と言われており,本実験でも 刺激強度をA D闘値とした。まず図1に示した様に2台の電気刺激装置により2相性矩形波パ ルスを発生させ,アイソレーターを通じてラ y卜の右肩桃核の刺激を行なった。刺激パラメー実験てんかん (Kindling) の生理心理学的研究
9
ch. Data Recorder ATAC-450 Computer V A =o
=
ロ u ¥、
1 1 ﹂ 両 日 ﹃ ' -n u F f t、
=
i
;
a T 目 、 d 咽 制 G / ﹂ v v V -J -VE--p'lh 完 ゾ 国 戸 ρ 、 │ 1 ﹄ har-戸 ﹄ s d h ﹂ U 晶 、 ハ U Etectric Stimulator1
JL
Electric Sti mutator2
u
sw
図1
実験ブロックダイアグラムターは60Hz,パルス持続時聞は4.5msec. 刺激時間は 1秒間で, 50μA より始め 50μAステッ
プで刺激強度を増し,初めて刺激部位にのみ約数秒間 2~3 Cfs椋ー徐波すなわち AD が出現し た強度をA D闇値とした。それを以後 Kindling用刺激強度として用いた。すべての被験体で A D闇値は 120----350μAの範囲であったO 翌日より毎日一定時刻に1回この A D閣値で、扇#搬 を刺激し,全身けいれん発作が数回連続して出現するまで続けた。発作の発展段階は Racine の分類聞に従い表lに示した通りであるo まず発作段階
o
(Stage 0 )は刺激に対して無反応 表 Kindling発展過程における各発作段階 (Stage) の分類 Stage 0 ---No Response,
Arrest or 1mmobility Stage 1 --- Mouth and Facial TwitchingStage 2 ---The above as well as Head Nodding Stage 3
-
ー
-
Forelimb Clonus in addition七0Head
N
.
oddingStage 4 ---Forelimb Clonus and Rearing onto the hindlegs
S七age5 ---Rearing onto七he hindlegs and
then Falling over
アンダーラインが各Stageでの分類基準
(No Response) か不動状態(Immobility) になることで,数回刺激を続けると口や顔面の 播樹 (Twitching, Stage 1 )が起こり,次に点頭 (HeadNodding, Stage 2) ,さらに筋緊 張した状態で前肢を持ち上げ (Forelimb Clonus, Stage 3 ,) Stage 4になるとその状態か
27-ら後肢で立ち上がり (Rearing) ,最終的に転倒(Falling )する Stage5に達するo これは てんかんの分類で言うと 2次性全般化けいれん発作あるいは大発作 (Grandmal) に相当し, との時点、で Kindlingが完成したと見倣す。この Stage5が数回連続して出現した日の翌日から 刺激強度を下げていき,もはや全身けいれん発作が出現しなくなる直前の刺激強度(Gener-alized Seizure triggering Threshol d; G S T) を調べた。 脳波は前頭部,後頭部皮質,両側の背側と腹側海馬,さらに刺激の対側及び同側扇桃核から 導出し,刺激前l分から刺激後 5分間のすべての脳波はソニー製 8チャンネルエルカセットデ
ータレコーダ (FE -39A) に磁気記録し,日本光電製AT AC-450を用いてパワースベク
トルを求めた(図 1)。個々のパワースベクトルの分析時間は
2
.56
秒,サンプリング時間は10m secで主に海馬脳波を中心に分析した。 その後ネンブタールで再び麻酔(10mg/ kg,腹腔内投与)し, Kindlingの完成したすべての 被験体の両側海馬を電気凝固法 (2m A, 20~30秒間)により破壊した。 DHPC (背側海馬) 破壊群は2匹, VHPC (腹側海馬)破壊群は 5匹であった。他にコントロール群としてそれ ぞれの海馬の上に位置する大脳皮質に刺入した電極を通じて同様の手続きで皮質を両側に破壊 した。少なくとも1
週間の回復期の後,破壊前と同じ刺激パラメータで扇桃核を刺激し行動及 び脳波を観察したo行動は破壊前と同様に各発作段階の出現状況や Kindlingの再出現の有無を 調べた。脳波的にはA Dの持続時間 (duration) と潜時(Iatency) 及びそれらの部位的差異, さらに刺激前 1分間の発作間散時発作性放電 (Interictal discharge,・ 11 D)や刺激後 5分 分間に出現した発作後発作性放電 (Postictal spike,・ P 1 S) の回数を測定したO すべての実験終了後,ネンブタール深麻酔下で頭部を濯流固定し脳を取り出した後,従来の 組織学的検索法倒により肩桃核刺激電極部位と海馬破壊範囲を確認した。結果
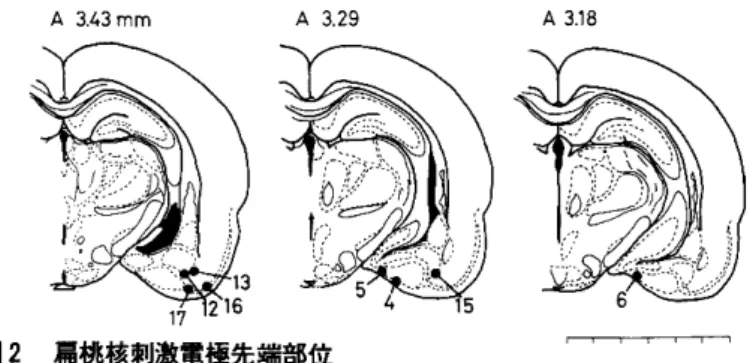
(1)肩桃核刺激部位
5timulating Electrode Tips in Right Amygdala A 3.43mm A 3.29 A 3.18 図2 肩桃核刺激電極先端部位 ・,-, 黒丸が電極先端部位,数字は被験体番号。図の上の数字は Kohigと Klippel(l1)の脳図譜でのLambdaより前(A)への距離。 図右下の1目盛は 1mm。
実験てんかん (Kindling)の生理心理学的研究 図2の黒点で示したものはすべての被験体の刺激電極先端部位で 2本の刺激電極のうちど ちらか一方あるいは両方がすべて右肩桃核群に属し,主に皮質核 (YK-4, 5, 6) と基底 核 (yK -12, 13) に位置していた。一部梨状葉皮質や嘆皮質に電極が刺入されていた (YK -16, 17)。これらの部位の刺激によりすべての被験体で Kindlingの発展が見られた。またこれ らの部位には組織学的に顕著な損傷(例えば電極や絶縁塗料のもつ毒性,金属イオン沈着によ るてんかん原性変化)は認められなかったことから, Kindling現象がニューロンの電気的な賦 活による経シナプス性変化であることを裏づけていた。
(
2
)
海馬破壊前の
K
i
n
d
l
i
n
g
現象の発展過程
図3は海馬破壊前の Kindling現象の発展段階を示したものであるO 図から明らかな様に, Kindlingが完成する (Stage5) までの刺激回数(日数)は 7----10日前後の群 (yK - 4, 12, 16, 17) と 17~20 日前後の群 (yK - 5, 6, 13, 15) に分けられた。全体の平均値は. 12.6 :i::5.1日で従来の報告(8)とほぼ一致していた。速くKindlingが完成した群は刺激に対して無反応 であるStage0から翌日直ちに Stage1となり,日毎に発作段階が発展していくのに対して,Kindling完成の遅い群は Stage0や Stage1の発作が比較的長く続き, Stage 2に達しでも日
数のかかる例が多く見られたD さらにこの群はStage3から 2ヘ,あるいは Stage-4から 3
へ
の発作の減弱(退行現象)が見られた。脳波上では海馬脳波のパワースベクトル分折結果を図 4 と 5 に示した口扇桃核刺激前と刺激後を比較してみると,刺激前は 7~8 Cfs 以下の海馬シー 。一一-0YK-45
φ ロvl
t
l
4
+-' (/)~
3
EU 2
O
D
e
v
e
l
o
p
m
e
n
t
P
r
o
c
e
s
s
e
s
o
f
K
i
n
d
[
i
n
g
0---0 YK-6 0---0 YK-13・
-
-
_
YK-15 0一一-0YK-16・
一
一
・
YK-175
1
0
1
5
2
0
S
t
i
m
.
s
図3 海馬破壊前の Kindling発展過程 横軸は毎日 1回の刺激回数 (Stim.s)。縦軸は表lに従った 各発作段階 (ClinicalStage)o Kindling完成が10日前後と 20日前後の二群に分けられる。 - 29ー8th Pre-Stim 8Hz
一一一一、「一一一一
o 5 10 15 20 2'j1) 5 10 15 20 2,
i IIL :) 101~ -;口 Y日 ld lS 70 ".)1.)l L' 国4 Kindling刺激前後の海馬脳派のパワースペクトル (YK -6) 各国の左端は刺激前後の海馬脳波のパワー刺激回数 (8th~ 19th) . 右は各stage (1~ 4 ) .縦線は8Hzo タj皮が優勢に出現していたのに対して刺激後は明らかに発作波を反映する 2~3 Cfs の徐波と 15 ~20 Cfs 以上の速波成分(これは椋波成分を含む)とが混在する複雑なスベクトル構造を呈する ようになった口図5は3匹のラットでの図右からそれぞれ発作段階(S tage) 3, 4, 5の例 を示したものであるが,発作段階が進むにつれてスペクトル構造が徐波を中心としたパターン から速波の多いより複雑なパターンへと移行していくのが明らかとなった。さらに一旦発作が 起きるとこのl次的発作は約2分間程度続き,数分後2次的発作が生じてくることが認められ た(図5. YK-4)。従って 1 回の発作は約 4~5 分間持続することになり,すべての被験体 でほぼ同じ傾向を示した。次に各発作段階での後発射 (AD)の持続時間,潜時及び発作後の 発作性放電 (p1 S )について分析してみると, A D持続時間とPISの数とは発作段階が進 むにつれて共に増大し,これら2つの測度聞には有意な相聞が認められた(図6,rニ0.24, t =2.09, dfニ72,pく.05)。しかしながら図7で、各Stage毎に相関係数をみてみると,これら 2つの聞で相関が高いのはStage1から4の 間 (r =0 .5~0 .6)で、Stage5になるとむしろ負 の相関となり, A D持続時間(これは1次性に全身けいれん発作が生じた時間を含む)が長く なるとむしろPISの出現数は少なくなり抑制されたことを示しているO 他の測度聞の相関も 同様の傾向を示した(図7
)
0
A D潜時に関しては約1秒前後でStage3でやや減少する傾向に はあるが有意なStage聞での差は認められなかった(図6口)実験てんかん
(
ki
n
d
l
i
n
g
)の生理心理学的研究
長
;主主主三三龍三
思足三三三奪 三
二コセ二二二二 ヨ 恒 三 叫 こ こ こ 二 二 三注三三三 縦 三 :A.-"t" ー ピ 二 二 二 二 点 む こ Jト一一一一 塁三三三 N _ご亡山 州 一 一 一
一 一 号 室 長
一 匡 一 一 一
一 一 一 一 一
一 一 一 一 一
一 一 一 一 一
一 一 一 一 喜
一 一 一 定
K
i
n
d
l
i
n
g
刺激時のパワースベクトル(YK-4
,5
,6
)
。 各被験体の8,14, 13回目の全記録時間内のパワースペクトルの変化。 右図からSt
a
g
e
3
, 4,5
0 (3)海馬破壊範囲
図5
9
,1
0
は各々K
ぬi
g
とK
l
i
p
p
e
l
の脳図譜(11)に従って破壊範囲を示したものであるO まず 図8はDHPC破 壊 群(
y
K-4,
部大脳皮質,脳梁,視床背側部に破壊が及んで、いた。VHPC
破壊群(図9
,Y
K-5
, 13とy
K-12
,1
7
)では主に海馬腹側部の海馬支脚
CA1
が破壊され 一 部 脳 幹 部 位 の 黒 質(
S
u
b
s
t
a
n
t
i
an
i
g
r
a
)
にも破壊範囲が広がっていた。これらの破壊後すべての被験体においCA4
, 歯 状 回 が 中 心 で ー6
)で,海馬背側部のCA1
, 図8
, 図10,
て一般活動性,摂食行動,体重等に著明な変化は認められなかった。 (4)破壊後のKi
ndling現象の再出現状況
図11
は海馬破壊を行なった被験体すべての破壊前後のK
i
n
d
l
i
n
g
の発展及び再出現過程を示し たものであるO 図 の 上 段 はDHPC破 壊 例 で , 明 ら か に2
例ともK
i
n
d
l
i
n
g
の 再 出 現 が み ら れ6 ~8 回目の刺激で、再び、 Stage5
に達した。脳波的にも図12
に示した通り破壊前のS
t
a
g
e5
での ' E i q J(sec) 60 50 40 30 J. イ (sec] 0.5 (/10 1';p.c 2.5 lη ム L AD Duration MEAN + SE N • 8 f ._--,-5 Stage
ιL
イ
5 5tage Postictal Spike 1一一一一一ーー← " St,)C[l図6 破壊前の後発射 (AD) と発作後発作性敢電 (Postictal Spike; PIS)の変化
M E A N士SE:平均値と標準誤差・横軸は各発作 Stage。 ( I-! ~ E ω .rl U 'rl 斗4 斗4 ω O u ロ 0.6 0.5 0.4 0.3 0.2 0.1 O .~ -0.1 ~ rcl aj-0.2 M M
8
-0.3 -0.4 Duration 後発射持続時間, Latency 後発射出現までの潜時。 国7、
、
、
、
N 8t
>
AD 1atency:PIS λD duration:AD 1atency AD dura七ion:P工S 1 2 3 4 5 Stage A DとP I S聞の各 Stageにおける相関係数(r 。)図B 図9 実験てんかん (Kindling)の生理心理学的研究 YK・4 !
/
i
1究弐
;
i
(
子 背側海馬破壊範囲 (YK -4, 黒で塗りつぶしたところが破壊された範囲。図の上下の数字は 図2と同様. Lambdaより前 (A)への距離。 YK-5 腹側海馬破壊範囲 (YK-5,13) これらの被験体はKindling再出現が抑制された(図13参照)。 - 33 VK・6YK-l1 典 和 口 YK-12 図10
Dorsal Hippocampal Lesions
After
。一ー一-0YK-4 o---<lト6 -<l 0・t.--o Q。
Before
0 0・・d Q Q ・ p・.O--<l 0 b-.-O .0-..0-.-0-・4 ヰヰ 5 4 1 J ヲ -1 ι ニ ユ ﹂ 内 心 ﹃ 宅 二 二 両 日 υT
l
ペf.¥
~,:.\,..r-l\
14i
Ri
l
、
i
M
M
j
i
i
RJ-l
/
i
i
八
/jMili
l
i
/
,
i
¥
¥
1
1
¥
/
\~!
¥
!
¥
/
¥
1
¥
f
¥
i
~ ~tLii
Stim.s 10 20 15 10Lesions
Vent r
a
l
Hippocampal
Stエロ.s 海 馬 破 壊 前 後 のKindling発展及び出現経過破壊 i請の結~はは]:1 とIt iJ じ。凶のl:が 1rf!!lJi毎!己破壊 (Dorsal Hippocampal
L山 ions) 群 (YK-4,6),ドが版{WJ (Vcntral) 群 (YK-:>,12, 13, 16,17)】,
実験てんかん
(
K
i
n
d
l
i
n
g
)
の生理心理学的研究発作波と全く変らないパターンを示した。それに対して図
1
1
下段に示したVHPC
破壊では5
例中
3
例(
y
K
-5
,1
3
,1
6
)においてK
i
n
d
l
i
n
g
の再出現が全く認められず,刺激に対して無反応か不動状態で脳波的にも明僚な
AD
は出現しなかった(図1
3
)
。またこのうち1
列(YK-1
3
)
は破壊後1
9
回の刺激を行なったがその間全くAD
の出現はなく,また図1
3
の下2
例(
1
3
t
h
t
r
i
a
l
と1
7
t
h
trial) で示した様に刺激強度を最初の AD 闇値である 150μA から 250~450μAに上げでもわずかに
AD
様のスパイクと約3
0
秒からl分後に全誘導に1mV
程度のPIS
が認 められたにすぎず,行動的にも明僚な臨床的発作はみられず不動状態であった。なおこの例で は刺激強度を最大1mV
まで高めたが同様の結果であった。これらの結果を示した例は図9に 示した範囲の海馬が破壊されていた。VHPC
破壊群の残りの2
例 (yK
-12
,1
7
)
において は,一旦K
i
n
d
l
i
n
g
の再出現をみるがその翌日の刺激ではS
t
a
g
e1
,2
ないしOになり発作の減 弱ないし消失(退行現象)がみられた。それがほぼ 2日から 5日の周期でリズミックに生じて いた(図1
1
)
。脳波像からみると(図1
4
)
,VHPC
破壊後4
回目の刺激で全身けいれん発作(
S
-t
a
g
e
5
)の再出現がみられ,2
次的発作波も海馬(この場合は背側海馬)を中心にt
o
n
i
c
に出 し PIS も破壊前と同程度に生じていた。しかし翌日の 5 白目の東u~>>t では AD が約10秒間出現 し行動的にはひげのわずかなけいれん(
St
a
g
e
1
)のみで明らかな発作は生じなくなった。そ れが 6回目になると再び全身けいれんに及び 7回目でまた同様の退行現象がみられた。但し刺 激から数分後に出現するPIS
に関しては破壊による顕著な影響は認められず,破壊前と同程 8efore lesjon 7lh Atterlesion 6 th SlIm. ,,;:-'''''''''''、 "',,,~~.r"、~叩吋内向山、,↓ ;::{t.m守'...-...:-Jヘ.,"'.,vへ.1'"ザポd作-W.JV岬 " ^ N ;<A吋¥ノ山戸、んヘムr¥p吋f,^岬'NYV'めいずd 司 oV小J寸品、Ivr...~'.t~.戸峠'Ý.~州内 LO 、..A.."-J' u,mgV孔,....J"'.ずY"¥fNN'"吋仏内内J 図12 背側海馬破壊前後の全身けいれん発作時の脳波パターン 凶の上が破壊前7
白J[l
目(
B
e
f
o
r
el
e
s
i
o
n
7
t
h
)
,下が破壊後6
[nl日(
A
f
t
e
r
l
e
s
i
o
n
6
t
h
)
の刺i
%
i
前後の記録。S
t
i
m
:
11肘'mÍ桃核車刺11ì~í股仰t矧H ,y},LO : I陀d筒4是~I}(i坑(iじ出叶i行i部11布1;, R A mq:イ1f日桃枝 ( 1
l
iJ]側),L A mg : fr:扇桃核 (吋fftlJ)〈
。
ゲ コ。
泊
、
以
ハ
コ
一
心
け
ニ
叩
F
J
い γ ﹂ ぜ 凶m
C
4
川 4刊
一
(
普
与
S E )∞
-L d J 山 間 剃 輔 副 同 之 副 市G
E
-2
2
県三副革川一や 4 二 日 沖 司 令 ( 一 日 と ︻ { 与 ) 一 一 一 三 廿 一 定 都 世 入 l H 明 文 相 席 謹G
灘揺軒揮蛇崎県軍盟 主園 月f
f
r
i
t
T
主
i
I
t
-T
s
i
l
i
c
i
-三 ヰ ョ.
3
8
3
J
J
J
L
j
f
!
i
1
i
l
l
l
ギF
L
E
i
-
毛
よ
j
a
,
r
i
喜F
F
l
i
l
-i
i
z
-5
2
L
E
t
-'
j
'
i
z
I
一
? 工t t
ミ ﹁
- i
T T
T t
1 f
T 7
t T
E τ
} l
I 1
l l
i t
i
-芋 之z
τ
:
ι
J
5
1
2
L
i
l
l
i
-1
i
i
i
3
2
1
f
f
F
干t
f
H
}
t
f
f
J
4
4
4
l
F
f
3
1
1
ミi
i
i
J
ミ
王
、
与
一
三
J
-e
一
i
l
i
t
?
耳 目 川L ?
と1 S
帥 E O
一 凶 釦
﹄ ι w
Z 4 一
咽 亡 ﹂
F Z
-トf
T
i
i
!
?
1
1
1
3
5
1
j
三
一
三
幸
一
書
号
一
一
言
司
.
3
.
3
ゴLifili--4
1
ム ミi
T
︿
一
三
五
一
幸
-喜
4 1主
主
主
二
重
喜
一
一
}
一
量
一
霊
主
主
主
主
¥
一
4
7
2
L
i
l
i
i
j
i
d
s
寸(
7
4
1
毛
主
主
主
主
主
主
ヲ
,
一
三
重
主
主
主
主
主
﹃
- z v
↑?とijjiii
一
1
1
よi
l
i
-3
i
3
a
r
.
-4
一
三
五
亘
書
主
主
亘
書
z
z
j
ξ
l
i
-l
i
z
F L
凶 C
O 一
帥 也
﹂ ﹄
@
-h d
司 一
帽 に
﹂ ﹁
F 古田レ
l
t
f
トT
T
l
i
i
j
j
i
l
-i
i
i
f
主
主
ミ
ミ
寸
L
i
l
i
-i
f
E
f i
a t
ふ すJ f
3 t
t
、
t t
E i
z
-- 2
4 V
E T
t f
I i
i I
t i
s t
1 1
F 2
4 ヰ
主 4
3 c
- f
t
-t
↑4 4
1 1
t i
-' i
i J
F E
d
-[
!
?
十
T
I
t
i
i
i
i
i
i
l
i
l
t
-i
l
t
E
E
午 杢 4 J h一
j
i
l
-!
i
v
i
l
l
i
-l
l
-t
i
f
-,
f
l
i
?
r
)
1
i
i
t
t
f
4
・
2
3
4
L
Z
4
手F
1
1
1
1
1
t
e
-i
;
-1 F 目 ﹄ E ミ司王五回lief
土
!
;
i
!
3
1
i
き
,
章
、
三
﹁
芸
.
歪
.
で
一
↓
主
妻
a
ミa
3
.
3
L
I
l
-t
A
t
i
t
i
i
l
i
l
i
-t
i
k
i
t
t
i
-よ
主
主
主
主
一
幸
子
一
主
主
主
a
d
z
-T
4
5
f
i
-i
l
j
j
;
i
d
i
l
f
T
r
T
十g
i
i
i
i
i
三
毛
主
主
主
一
委
主
主
主
E
t
と
・
5
J
r
l
i
l
i
-I
{
t
j
i
J
1
2
j
i
i
j
ミ
一
J
主
主
主
J 3、
主
主
J
d
h
i
z
-z
r
L
i
i
f
E
J
M
F
F
4
-2
↑5
1
1
4
i
i
i
γ
F
i
j
i
j
一
重
一
三
一
三
一
一
一
一
三
二
蓮
寺
三
三
j
i
l
l
f
主 主
主 主
主 主
主
i
?
とJ E
主
主
主
主
主
主
主
主
主
-1
宅
百
三
- F
、
k
j
j
i
i
l
a
i
d
-2
i
1
1
3
E
A
平ι
E
E
'
f
r
1
主
主
主
主
主
主
一
三
一
二
重
喜
章
主
主
主
主
工
、
1
4
i
l
i
-i
‘
I
i
s
i
i
J
f
f
i
i
i
主
主
是
正
玉
三
宮
F書
官
主
主
主
君
主
︿
-3
2
F
l
j
話
3
Z
E
E
-N
F
z r h
F
︼F 8 2 2
3 E E
的 ﹁ ﹄ O 一 山 由 ﹂ 司 a E M W U o a a 一 工 一 E v c h w ﹀ 。 ( 二 仰 二 ド J同 刊
一 石
) 川 平 一 ﹄ ー や 制 ︿ EH 什一包立唱イ﹃叫 c m ず や っ 一 に 同 ) ロ 田 口 ' 士 ぃ { 川 一 ︿ 司 会 的 N f p f ︿ ﹃6
2
s
h ( 王 己 ) 白E
2
o
u
J
持
制
一
一
世
主
的
榊
冨
G 耳坦昨 ω z一
司
E出
一
泡
勝
握
入 l H 明 文 相 屑 謹 Q事
握
騨
橿
瞳
崎
一
軍
睡
竺図、 E E ‘ , , E 4 ・it
J ' E , E M
' a E
1
1
4
i
・
3
1
1
1
1
2
i
0
・
E ﹀ 1
寸4
T
$
7
4
6
1
イ
十
子
主
主
F
-2
7
i
5
7
t
l
s
主
2
1
l
i
l
i
-?
E
t
=
担 ︻1
2
Z
帥
E
O
B
白. 4 ﹄ @
Z 4 一 一
団 ﹄ ﹂ F
L H
トF
;
;
i
I
j
l
?
?
j
i
l
i
-7
i
i
i
j
- f
t '
f i
L 4
T p
i t
i t
i J
﹄ h ザ3
圭 てT 1
4
﹄t T
I l
l i
-g t
j J
4 1
t F
S F
F h
﹄I i J
1 4 }
t z a
d
- f '
L Y E
- 3 3
t s t
t s f
f - F
2 g 主
t t h
f J L
J f t
p t J
t f t
J B I
t - }
2 4 h
t z J
3 5
え てf p k
、 ¥ E
t i
-- l i
l ' l
' I S
5 1 1
r
官 官 e; o h i
, ﹀ t t
・ 5 a t
l B t f
J t s d
B S , 、
ィ t 3 f
、 P
﹀ τ, J 、 ィ J
B I t t
-‘ , f h p
b L 4 1 ‘
I ' P 4
もE
S
£4 5 1 t S
も3 5 p d
t a t 、
i 、 ‘ k
r 1 4 、
4 p t r
h t t f
J t t t
、 , t I
t l E
;
t
l
l
i
f
t
i
曾f
i
I
f
-J
a
l
l
-i
i
J
E
v
-t
i
s
-引
主
主
主
-Z
Z
H
円 pi
l
l
i
-1
1
1
t
i
l
i
l
i
-h
i
l
}
f
i
r
-L
i
l
l
i
-l
l
J
f
!
;
i
'
l
i
l
i
-一 ‘ 7 ' t
g
をf t s e
﹄l f ' 4
t 3 4 L
4 1 1 t
t t S 5
9 h p p
孔J - 4
f E 4
i 1 J
L i t
, s 6
4 4 L
t t 1
5 1 f
t t i
b
-. t J
i l t
E
等i b i
- - t
1 E b
-i
i
i
i
l
i
l
i
-i
i
l
i
-i
I
l
l
u
-1
1
1
i
i
J
i
l
i
l
t
j
、
三
ゆ
J
F
i
-E
1
4
h
Z
4
-Z
£ 聞い 1
1 s
i l
i
-- l
i I
l i
-} i
t
-- I
l l
i
-- 1
4 T
l '
ムi s
-i t
E a
s
-- a
s f
E
一 ' h e
- - b E
F F P 2
v h 4 4
I L e
-- 4 3 ・
f f 3 4
B S 1 t
e t f 3
1 2 2 1
1 1 1 4
f ' i 、
, ‘ . 2
・ 1 Z
﹂ ; ' ﹃4 . , 、 4
・ a u
-2 9 -2 1 1
' : I t
-- , 、 町 J
u - a g
i
l
i
-?
f
i
f
t
i
-i
I f
-- t i
l l
-- P 1
h f l
i t
-- -- f
t 1 1
t i l
J 1 9
g - h
t i h
- - g
l ' t
f t !
t i s
- i t
もE
-i
l
l
i
t
-j
i
i
i
g
-a
凶
r﹄
。
一
凶
ω﹂ ﹄ @ Z
4 - 咽 =
﹂ ﹁ Z H
a 司
- r
ー?
4
?
i
}
1
i
l
i
-、
主
主
一
去
三
一
三
塁
委
主
主
主
三
重
ミ
J
f
i
l
l
i
i
J
l
J
J
1
i
t
-、
寸
斗
、
コ
ミ
ミ
コ
そ
t
t
豆 室 ミ 1主
主
主
主
主
主
幸
喜
i
I
4
5
3
f
i
t
-t
i
l
l
-、
b
i
d
i
-i
f
i
!
土
と
主
主
主
と
E
t
4
三
三
一
ミ
﹃
1 2 4 2 号 吾 妻 童 手f
j
、
、
-T
i
l
i
l
i
-F
i
l
i
-i
i
l
i
f
-f
i
j
j
三
主
主
主
主
主
・
5
3
3
j
7
電E
f
-i
i
i
l
i
-I
i
L
1
1
i
J
f
j
l
J
,
i
J
J
豆E z
・
-吾
妻
宅
Z
4
5
3
2
3
1
・
立
日
目
有
日
刊
i
p
d
剖創立ふ占EEE
凶 C O B & ﹂ 可 a E g o a a 王 一 E V C 釦 ﹀ 剖( 再 三 田 引E実験てんかん (Kindling)の生理心理学的研究 Ventral Hippocampal Lesions 圏内腹側海馬破壊後の発作波パターン 短時間(約8秒間)で発作 (Stage4, Rear ing )が終了していることに注意。 この被験体 (YK-l7)も Kindlingの退行現象を示した(図11参照)口 度の
1mV
以上高電位椋波が出現していたO 図15
は退行現象の生じた別の被験体(YK-17)
の脳波例を示したもので,破壊後12回目の刺激後約1.5秒 た っ て か ら 右 肩 桃 核 (RAmg)優 勢 にADの出現と同時に点頭 (Headnodding) が生じ, 7~8 秒後に急速に後肢で立ち上がる ( Rearing) S tage 4に達しその後突然発作が終了し,もはやスパイクの出現もなく正常と思 AD Duration ft-12: 川川
(ne) , 。:
:
:
.
0
O-~ 4rtn ,P'" , , , , , , , , , , , , , , ~..._.c ... U P田Z 10 $ta. ,e Stqe (110・
00)1 Postictal Spikes 図16 腹側海馬破壊前後の A DとPISの変化 退行f見幕を示した被!強体 (yK-12) の11間投び唱存Stage聞での変化。 詳細は本文を参照口 Sta., 月 i 円 ふVentral Hippocampal lesions g B E N -吋1 1 1 吋 } l l J ﹃ l l l﹂ 寸 l l l J I l -c e 5 4 3 2 L ' o l / AD Duration
-
・
:
1
!
J
E
::rOR.E: ,?0-・-<1'円四円~勺日
- - 、 ,、 loi a- .,,o-....-d 5 0 StAqeu l
-叫 l l 1 1 1 ー ー AD latencyム二十一〉噌
. stage Postictal Spike 17'._..4、、、 ,,-P - 、 、 ' /- ?"-¥ 可プ . d / Stl9'e 図17 腹側海馬破壊前後の A DとPISの変化 図16
と同様にもうl
例(YK-17)での変化
われる脳波パターンを示した。これはKindlingの発展過程では全く見られない出現様相であっ た。すなわち通常のKindling現象は一旦発作が出現し主身に及んだ場合,それは約数分間持続 する(図5)ことが多い。従ってこのタイプの退行現象も明らかに海馬破壊効果と考えられた。 図2
各被験体の A D持続時間.潜時及び PIS聞の相関保数Sub. n AD duration:1atency AD duration:PIS AD 1atency:PIS
YK-4 6 0.0469 巴 0.8107 0 0.0223 ( 5) (0.7878) (0.8688)ロ (0.8454) 0 YK-5 11 -0.3671 ー0.2418 0.1142 YK-6 12 -0.2991 -0.2977 -0.2229 (8 ) (0.2037) (0.0261) (0.2958) YK-12 10 -0.8178*合 肯 0.8279官 官 -0.0732 ( 15) (-0.4415)。 (0.5209)* (-0.2500) YK-13 12 0.5341 -0.1934 0.2631 YK-15 13 -0.84l3*** 0.8978ま** -0.6873まt YK-16 8 一0.1214 0.8741宮 古 0.0071 YK-17 12 0.2011 -0.1587 0.1047 (16) (-0.6407)** (0.4853)白 (ー0.4594)。 ***: p<.001, 官 官 pく.01, *: pぐ.05,0:' ".1>p).05 カッコ内は破壊後の値を示す。
実験てんかん (Kindling) の生理心理学的研究 次にA DとPISの破壊前後の出現状況をより詳しく比較してみると図16,17の通りであったD これらはVHPC破壊後発作の退行現象を示した例をそれぞれ個別に図示したものであるO ま ず図16の例では図左の刺激回数(日数)による変化からみて明らかに発作の退行現象が見られ, それに応じてA D持続時聞がほぼ
1
対1
で変化していた口しかし潜時やPISとは明白な対応 関係は認められなかった。図右の各発作段階毎にみると, A D持続時間は破壊後減少する傾向 であった。しかしStageが進むにつれてその差はほとんどなくなった。潜時についても同様に Stage 1, 2の早い発作段階ではA Dの遅い潜時のものが多いがStage4や5になると破壊前 とほぼ同じ値を示した。 PISに関しては個体内変動が大きく一定した傾向は認められなかっ た。破壊後わずかにPISの増加する傾向がみられたが,破壊による効果というよりもむしろ 肩桃械を刺激した回数に比例して山ると考えられるO 図17の別の例でも同様の結果を示して台 り,海馬(持にVHPC)破壊により刺激直後に出現する A Dに対しではある程度抑制的効果 を及ぼしたが,発作後数分たってから出現するPISに対してはそれほど効果を持たないこと がうかがえた。すべての被験体で破壊前後のA D持続時間,潜時, P 1 Sとの聞の相関係数を 求め表2
に示したD 特にVHPCを破壊したYK-12
のラットに注目すると, A D持続時間と PISとの聞の正の相関が破壊後有意で、はあるが低くなっていた。またA D持続時間と潜時ど の負の相関が有意でなくなるのが目立った。しかしながら退行現象を示したもう1
例 (YK-17) では逆に VHPC破壊後A D持続時間と潜時との聞の負の相関が有意な方向に変化したD またDHPC破壊群でも個体差が大きく一定した傾向とは考えられなかったD コントロール群としてそれぞれの海馬の背側に位置する大脳皮質を破壊した結果については, ここで示していないが, Kindling再出現に対する明らかな破壊による影響はなく,ほぼDHP C破壊群と同様の結果であった。考察
本実験で得られた結果をまとめると,海馬破壊によりKindlingの再出現が抑制された。つま り海馬(持に腹側海馬,・ VHPC) はKindlingの再出現に対して促進的あるいは Kindlingの永 続性を維持する働きを持っていると考えられた。この結果は従来示唆されてきた報告(16)聞とは 矛盾するようであるが,海馬はKindling発展過程に対しては抑制効果を持ち側, Kindlin~ の再 出現あるいはKindlingの永続性に対しては促進的に{動いていると考えたほうが妥当であろう口 すなわち.Kindlingが形成されていく過程で働く脳内メカニズムと形成された後の永続性に関 与するメカニズムとは全く異なることを示唆しているかも知れない。しかしながら本実験と同 様にKindling完成後海馬を破壊した実騨16)で逆の結果が報告されているD この実験はネコを用 いてKindling完成後海馬(特に海馬後部から腹側部住)を両側に破壊した口その直後から扇桃 核刺激を行ない1回目の刺激で全身けいれん発作が再出現し徐々に発作の程度が増強し数日目 で死に至った口このことから海馬はKindling現象の再出現を抑制していたと彼らは考えた。本 -3S-実験との相違は海喝破壊から再び扇桃核を刺激するまでの時間経過である口海馬を破壊すると その最申からKindlingに似たけいれん発作が誘発され,その後数分から数 10分 発 作 後 抑 制 (postictal depression) があり回復するoこれは通常のKindling形成過程で見られる現象と 全く同じで,海馬破壊そのものがKind
1
i
ng刺激効果であったことを示しているo従って海馬の Kindling再出現に対する働きを検討する場合海馬破壊による上述の直接的効果をできるだけ少 なくする(本実験では破壊から 1週間後に再刺激した)ことが望ましいと考えられるo この1 週間の回復期間中に海馬のてんかん原性聞による自発けいれんの出現等は観察しなかったが, 少なくともKindlingの再刺激直前の脳波や行動を見る限り発作間歌時発作性放電 (1 1 D )や 発作後抑制など発作を反映した脳波及び行動パターンは全く示していなかったO このことから も海馬破壊の直接的影響はほとんど消失していたものと考えられた。 海馬破壊によりいかにしてKindlingの再出現が抑制されたかを示唆するいくつかの報告(6)側 聞があるo まず通常のKindling形成過程では脳内モノアミンのうちドーパミンの濃度が大脳辺 縁系あるいは前脳 (forebrain)領域で有意に減少する(6)凶ことが知られているO 一方, Sprin-gerとIsaacson聞はラットの海馬(背側から腹側のほぼ全域)を両側に破壊し 1週間後に脳内 (特に側坐核)ドーパミンの濃度を測定したところ統制群と比較して有意に高くなっているこ とを報告しているO これらのことから本実験で、のKindling再出現に対して海馬破壊は脳内モノ アミン,特にドーパミン系の2次的変化をもたらしたのではないかと考えられる。すなわち脳 内ドーパミンの線維経路は最終的に扇桃核群に投射しており側,そこから海馬支脚(subiculum) の腹側部位への線維結合も報告されている(12)。従って海馬特に腹側海馬がKindlingにより賦活 された神経回路と脳内ドーパミン線維系両方に対して積極的に働きかけていたことが推測され るo 本実験結果を別の観点からとらえることもできるO 以前から海馬の記憶メカニズムに対する 役割が注目されている(10)側一方で, Kindling手続も同様に記憶や学習の背景となる脳内過程の 変化と同義に考えられる点が多い(7)(13)。このことから本実験結果を脳内記憶過程の障害として 考えることもできるo しかしながらその詳細については今後の研究に委ねなければならないが, 少なくとも海馬特に腹側海馬は,Kindlingされた神経回路を再賦活するのに対して持続的ないし 一過性に促進したりそれを維持する働きを持っており,その背景に脳内カテコラミン系(特に ドーパミン系)あるいは脳内記憶メカニズムが重要な位置を占めていることが示唆されるO 本実験ではKindlingに対する海馬内の部位的差異が指摘された口このことは刺激した肩桃核 と腹側海馬とは直接線維結合を持ち背側海馬はそうではない(4)(12)という形態学的差異がある程 度反映されたものかも知れないが,本来これらの海馬の部位は機能的に全く異なっていること も考えられるo そこで今後は海馬の入出力系を部位的に別々に考慮した上で海馬と密接に線維 連絡を持つ他の大脳辺縁系や脳幹部位の働きについても吟味し, Kindlingに対する海馬本来の 役割をより詳細に検討していく必要があろう白実験てんかん (Kindling)の生理心理学的研究
要約
本実験はてんかんの実験的モデルである Kindling現象の永続性に対する海馬の働きを検討す るために扇桃核Kindlingの完成したラットの海馬を破壊しその後の Kindling再出現様相を脳波 的及び行動的に観察した。得られた結果をまとめると,まず背側海馬破壊により Kindlingの再 出現が見られ,何んら著明な変化は認められなかった。それに対して腹側海馬を破壊した数例 において 10数回再刺激を行なっても後発射 (AD) 等の発作を反映する脳波パターンを示さず 行動的にも不動状態でKindling再出現が全般的に抑制されたo また残りの腹側海馬破壊例では ~s
.
Kindlingの再出現を示したが約数日周期で発作の退行現象が見られたoこれらの海馬破壊 効果の背景には脳内カテコラミン系や記憶メカニズムの変容が考えられたoReferences
1) Anchel, H., and Lindsley, D. B. Differentiation of two reticulo-hypothalamic systems regulating hippocampal activity. Electroenceph. clin. Nevrophysiol, 1972, 32, 209・226. 2) Bennett, T.L., and Gottfr恒d, J. Hippocampal theta acti叫tyand response inhibition. Electroenceph. clin. Neurophysiol., 1970, 29, 1部-2∞
.
3) Douglas, R.
J
.
Thehippocampus and behavior. Psychol. Bulletin, 1967, 67( 6 ,)416-442. 4) Elul, R. Regional d.ifferences in the hippocampus of the cat. : Il Projection of the dorsal and ventral hippocampus. Electroenceph. clin. Neurophysio 1,.l 964, 16, 489-502. 5) Engel, J., and Katzman, R. Facilitation of amygdaloid kindling by lesions of the stria terminalis. Brain Res., 1977, 122, 137-142. 6) Engel, J., and Sharpless, N. S. Long-Iasting depletion of dopamine in the rat amygdala induced by kindling stimulation. Brain Res., 1977, 136, 381-386.7) Goddar,d G. V., and Douglas, R. M. Does the engram of kind.ling model the engram of normal long term memory? In Wada, J.A. (Ed) : Kindling. New Y ork Raven Press,
1976, 1-18.
8) Goddar,d G. V., Mclntyre, D. C., and Leech, C. K. A permanent change in brain function resulting from daily electrical stimulation. Exp. Neuro,.l 1969, 25, 295-330. 9)岩崎庸男,反応抑制に関する海馬の役割 東京教育大学教育学部紀要 1976,77-87 10) Klemm, W. R., Landfiel,d P. W., McGaugh,
J
.
L, and Tusa, R.J
.
Theta rhythmand momory. Science司 1972,176. 1449
ll) Konig, J. F. R., and KlippeI, R. A. The Rat Brain. The Williams and Wilkins, Baltimore, 1963.
12) Krettek, J. E, and Price, J. L Projection from the amygdaloid complex and adjacent olfactory structures to the entorhinal cortex and to the subiculum in the rat and cat.J.
Comp. Neurol., 1977, 172,723-752.
13)Leech, C. K., and Mclntyre, D. C. Kindling rates in inbred mice : An analog to learning?
Behav. Biol,. 1976, 16, 439-452.
14)McC.aughram, J.A., Corooran, M. E., and Wada, J.A. Role of the forebrain
com-missures in amygdaloi,d kin占ingin rats. Epilepsia.1978, 19, 19-33.
15)Mclntyre, D. C., and Goddard, G. V. Transfer, interference and spontaneous recovery
of convulsions kindled from the rat amygdala. Electroenceph. clin. Neurophysiol., 1973,
35, 533-543.
16)森 分 隆 , 森 分 清,佐藤光源,ネコの扇桃核キンドリングに対する海馬の役割 第12回日本脳
j皮筋電図学会抄録集 1782, 131.
17)Nauta. W. J.H. Hippocampal projections and related neural pathways to the mid-brain
the cat. Brai n.1958, 8,1 319-340.
18)奥田 宏,和国有司,三辺義雄,山口成良,吉田和典, 外側膝状体キンドリング描に対する閃
光一Thiosemicarbazide 賦活の効果第17回日本てんかん学会抄録集1983
19)Petsche, H., Stumpf, Ch., and Gogolak, G. The significance of the rabbit's septum as
~..relay station between the midbrain and the hippocampus : 1. The control of hippocampus
', aro~sal activity by the septum cells. Electroenceph. clin. Neurophysiol.1962, 14, 202-211.
20)Racine, R. Kindling: The first decade. Neurosurg., 1978, 3, 234-252.
21)Racine, R..J. Modicacation of seizure activity by electrical stimulation : 1.After
-discharge threshold. Electroenceph. clin. Neurophysio,.l 1972, 32, 269-279.
22)Racine, R.J.Modification of seizure activity by electrical stimulation : 11.Motor seizure
Electroenceph. clin. Neurophysiol,. 1972, 32, 281-294.
23)Racine, K, J., Gartner, J.G., and Burnham, W. H. Epileptiform activity and neuronal
plasticity in,limbic structures. Brain Res., 1972, 47, 262-268.
24)Sato, M., and Nakashima, T. Kindling: Secondary epileptogenesis, sleep and catechol
-amines. In.Wada, J. A.(Ed) : Kindling. New York Raven Press, 1976, 103-116.
25)佐藤光源, Wada, J.A. 新しい実験てんかんモデルとしてのと Kindling//preparation
:展望,脳と神経, 1975,27,257-276
26)Scovil1e, ,W. B., and Milner, B. Loss of recent memory after bilateral lesions.
J
.
Neurol. Neurosurg. psychiat., 1957, 20, 11-21.
27)Springer, J.E., and Isaacson, R.L. Catecholamine altらrationsin basal ganglia after
hippocampal lesions. Brain Res., 1982, 252, 185-188.
28)Ungerstedt, U.
.
.
s
tereotaxic mapping of the monoamine pathways in the rat brain. Acta.実験てんかん (Kindling) の生理心理学的研究
29)Wada, J.A Kindling. New York Raven Press, 1976.
30)Wa也, J. A., and Osawa, T. Generalized convulsive (or spontaneous recurrent)
seizure state induced by daily electrical amygdaloid stimulation in Senegalese baboons
(pa-pio papio) . Neurol., 1976, 26, 273-286.
31)Wada, J. A., and Sato, M. Generalized convulsive seizures induced by daily electrical
stimulation of the amygdala in cats : Correlative electrographic and behavioral features.
Neurol., 1974, 24, 565-574.
32)Wa也, J.A., and Sato, M. The generalized convulsive seizure state induced by daily
electrical stimulation of the amygdala in split brain cats. Epilepsia, 1975, 16, 417-430.
33)Wada, J.A., and Sato, M. Effects of unilateral lesion in the midbrain reticular formation
on kindled amygdaloid convulsion in cats. Epilepsia, 1975, 16, 693-697. 34)Wada, J.A.,佐藤光源. 燃え上がり現象一てんかんと精神病への新しいアプローチー創造 出版, 1981. 35)吉田和典,海号シータ波と視覚誘発電位との関連.福井医科大学一般教育紀要, 1981,第1号, 31-42. 36)吉田和典,海馬ニューロン活動と視覚情報処理について.福井医科大学一般教育紀要, 1982, 第2号, 9 -23. 脚注 (注)本研究は昭和57年, 58年度文部省科学研究(奨励研兜A})の一環として行なったもので, 一部はすでに第29回国際生理科学会議(シドニー,オーストラリア)で発表し, Physiology & Behavior(1984)に印刷中である。 -
43-Bulletin ofLiberal Arts, Fukui Medical School No. 3 (1983)
The Effect of Bilateral Hippocampal Lesions on Kin
-dled Amygdaloid Convulsion in Rats.
Kazunori Y oshida
Department of Psychology
Abstracts
In the present study, rat hippocampus was lesioned bi1aterally after completion of amygdaloid
kind1ing, in order to examine how hippocampus affects reappearance of kindling phenomenon.
The obtained results were summarized as follows; (I)The kindled rats with dorsal hippocampal
lesions again reached the genera1ized convulsion (kindling) within several days after the recovery
period from lesions. Afterdischarge (A D) duration and latency were somewhat changed
in direction to the inhibition of rekindling, but no remarkable change was observed in the feature
of convulsive behavior as compared with that before lesions. (2)The ventral hippocampal lesions
of some kindled rats inhibited the reappearence of any kind1ed seizures. No electrographical
discharge was provoked by restimulation of the original focus (amygdala) for a maximum of
several weeks, even when stimulus intensity was heightened over AD threshold (to1 mA).
And the other rats of this group showed the regression of generalized convulsion at two to five
day cycle which was not seen in the usual process of amygdaloid kindling. (3) These results
suggest that hippocampus, especially ventral parts of hippocampus, would have rather faci1itatory