Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬学) 報 告 番 号 甲第1761号 学 位 記 番 号 第358号 氏 名 堺 陽子 授 与 年 月 日 令和 2 年 3 月 25 日 学位論文の題名 In vitro 胆汁鬱滞肝毒性評価系の構築 論文審査担当者 主査: 肥田 重明 副査: 松永 民秀, 木村 和哲, 湯浅 博昭
名古屋市立大学学位論文
In vitro
胆汁鬱滞肝毒性評価系の構築
令和元年度 (2020 年 3 月)
名古屋市立大学大学院 薬学研究科
堺 陽子
一. 本論文は、2020 年 3 月名古屋市立大学大学院薬学研究科において審査さ れたものである。 主査 肥田 重明 教授 副査 木村 和哲 教授 湯浅 博昭 教授 松永 民秀 教授 二. 本論文は、学術情報雑誌に収載された次の報文を基礎とするものである。 【基礎となる報文】
1. Yoko Sakai, Takahiro Iwao, Takeshi Susukida, Akinori Takemura, Takumi Nukaga, Shuichi Sekine, Kousei Ito, Tamihide Matsunaga, In vitro bile acid-dependent hepatocyte toxicity assay system using human induced pluripotent stem cell-derived hepatocytes: Current status and disadvantages to overcome.,
Drug Metab. Pharmacokinet., 34; 264-271, 2019
2. Yoko Sakai, Hiroki Okumura, Takahiro Iwao, Koichi Watashi, Kousei Ito, Tamihide Matsunaga, Development of an in vitro cholestatic drug-induced liver injury evaluation system using HepG2-hNTCP-C4 cells in sandwich configuration.,
Toxicol. In Vitro, 61; 104619, 2019
三. 本論文の基礎となる研究は、松永 民秀 教授の指導の下に名古屋市立大学
略語一覧
BAs; bile acids
BCRP; breast cancer resistance protein BEI; biliary excretion index
BSEP; bile salt export pump
CDF; 5- (and-6) -carboxy-2',7'-dichlorofluorescein
CDFDA; 5- (and-6) -carboxy-2',7'-dichlorofluorescein diacetate CsA; cyclosporin A
DAPI; 4,6-diamidino-2-phenylindole DEX; dexamethasone
DILI; drug-induced liver injury DMSO; dimethyl sulfoxide ECM; extracellular matrix FBS; fetal bovine serum FXR; farnesoid X-receptor
GFR-Matrigel; Matrigel matrix growth factor reduced HBSS; Hank’s balanced salt solution
HGF; hepatocyte growth factor
HHs; cryopreserved human primary hepatocytes iPS cells; induced pluripotent stem cells
KO-DMEM; knockout Dulbecco’s modified Eagle’s medium KSR; knockout serum replacement
MDR1; multidrug resistance protein 1
MRP2; multidrug resistance-associated protein 2 NTCP; Na⁺-taurocholate cotransporting polypeptide OATP1B3; organic anion transporting polypeptide 1B3 OSM; oncostatin M
PBS; phosphate-buffered saline PVDF; polyvinylidene difluoride
SC-RHs; sandwich cultured rat hepatocytes SC-HHs; sandwich cultured human hepatocytes SDS; sodium dodecyl sulfate
tauro-nor-THCA-24-DBD; N- (24-[7- (4-N,N-dimethylaminosulfonyl-2,1,3-benzoxadiazole) ]amino-3α,7α,12α-trihydroxy-27-nor-5β-cholestan-26-oyl) -2'-aminoethanesulfonate
VPA; valproic acid
i 目次 第一章 序論... 1 第二章 HepG2-hNTCP-C4 細胞のサンドイッチ培養による胆汁鬱滞肝毒性評価 ... 4 2.1 緒言 ... 4 2.2 実験方法 ... 6 2.2.1 試薬及び細胞 ... 6 2.2.2 細胞培養 ... 9 2.2.3 RT-qPCR 解析 ... 9 2.2.4 蛍光免疫染色 ... 10 2.2.5 蛍光基質を用いた胆汁酸トランスポーターの機能活性 ... 11 2.2.6 Cyclosporin A、MK571 による胆汁酸濃度依存的な細胞毒性評価 .... 12 2.2.7 薬剤性胆汁鬱滞肝毒性評価 ... 14 2.2.8 統計学的解析 ... 14 2.3 結果 ... 15 2.3.1 胆汁酸トランスポーターに関与する遺伝子発現 ... 15 2.3.2 細胞の形態およびタンパク質の局在 ... 16 2.3.3 胆汁酸取り込みトランスポーターの機能評価 ... 18 2.3.4 胆汁酸トランスポーターの機能評価 ... 19 2.3.5 胆汁酸濃度依存的な細胞毒性評価 ... 19 2.3.6 薬剤性胆汁鬱滞肝毒性評価 ... 20 2.4 考察 ... 22 2.5 小括 ... 26 第三章 サンドイッチ培養したヒト iPS 細胞由来肝細胞を用いた胆汁鬱滞肝毒性評価
ii の検討 ... 27 3.1 緒言 ... 27 3.2 実験方法 ... 28 3.2.1 試薬及び細胞 ... 28 3.2.2 細胞培養 ... 28 3.2.3 ヒト iPS 細胞から肝細胞への分化誘導 ... 29 3.2.4 蛍光免疫染色 ... 30 3.2.5 RT-qPCR 解析 ... 30 3.2.6 ウェスタンブロッティング解析 ... 31 3.2.7 蛍光 NTCP、BSEP および MRP2 プローブを用いた胆汁酸トランスポ ーターの機能活性 ... 30 3.2.8 Cyclosporin A による胆汁酸濃度依存的な細胞毒性評価 ... 32 3.2.9 薬剤性胆汁鬱滞肝毒性評価 ... 33 3.2.10 統計学的解析 ... 33 3.3 結果 ... 33 3.3.1 経日的な形態変化と MRP2 および radixin タンパク質における蛍光免疫 染色 ... 34 3.3.2 胆汁酸トランスポーターの mRNA およびタンパク質発現 ... 36 3.3.3 蛍光基質を用いた胆汁酸トランスポーターの機能活性および毛細胆管形 成能 ... 38 3.3.4 胆汁酸濃度依存的な肝細胞毒性 ... 39 3.3.5 サンドイッチ培養したヒト iPS 細胞由来肝細胞と HepaRG 細胞および HepG2 細胞間における胆汁鬱滞性肝毒性の検出の相関分析 ... 40 3.4 考察 ... 42
iii
3.5 小括 ... 45
第四章 総括... 46
謝辞 ... 47
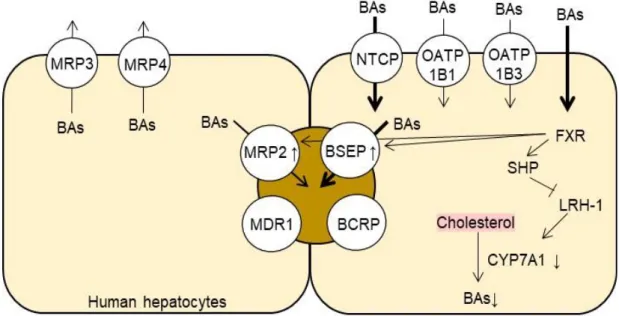
1 第一章 序論 非臨床試験における肝毒性試験は、必須項目であることから、肝臓における医薬 品候補化合物による毒性をより正確に予測することは非常に重要となる。そこで、薬 剤性肝障害 (DILI) を早期発見する技術を見出すことは、医薬品開発の臨床試験 の段階で起こるリスクやコスト削減の面から有用である。DILI の一つである胆汁鬱滞 肝障害を起こす原因の一つとして、薬剤によるトランスポーターの機能阻害があり、肝 細胞内または毛細胆管に胆汁酸が蓄積するために起こることが知られている[1]。また、 肝 細 胞 に 発 現 し て い る ト ラ ン ス ポ ー タ ー (Na⁺-taurocholate cotransporting polypeptide : NTCP 、 bile salt export pump : BSEP 、 multidrug resistance-associated protein 2,3,4:MRP2,3,4) は胆汁酸の取り込みと排出に密接に関わり、薬物や内因性 物質の体内動態に深く関与していることから、そのトランスポーターの活性変動や遺 伝子多型が薬理作用や副作用の発現に大きな影響を与えることも知られている (Fig. 1)[2]。したがって、トランスポーターに着目した胆汁鬱滞肝障害評価系は、肝毒性を 予測するためのモデルの一つとなる。 現在、肝毒性試験系としては実験動物を用いることが多いが、ヒトとの間には種差が あるため、結果をヒトへ外挿することが困難である。また、3R (Replacement、Reduction、 Refinement) の規制強化が進んでいる。そのため、動物を使用することなく、安全性、 毒性評価を行う方法を見出すことに注目が集まっている。そこで、in vitro 試験系として、 ATP 依存的な小胞内輸送の検証のために胆管側膜ベシクルを用いた方法があるが、 他のトランスポーターとの生体内相互作用を解析できない難点がある[3]。その欠点を 解決するために、ヒト初代肝細胞やヒト肝ガン由来のHepaRG 細胞や HepG2 細胞など が用いられている。しかし、ヒト初代肝細胞 (HHs) は、新鮮な肝細胞の入手が困難で あり、培養によって機能が著しく低下しやすい[4]。また、HepaRG 細胞は、高額かつライ センスを取得した人のみに使用が制限されるといった、問題点がある[5]。また、HepG2
2 細胞は胆汁酸のトランスポーター活性が低いなど、生理的状態を再現しているとは言 い難い[6]。このように、現在、医薬品の安全性評価系は、数多く報告されているが、さ らなる改善が必要であることが示唆されている。さらに、肝毒性の研究に関して、国内 外問わず、薬剤によるトランスポーターの機能に対する影響の検討は行われているが、 トランスポーターが関わる肝細胞毒性を評価する系に関する報告は少ない。したがっ て、in vitro 薬剤性胆汁鬱滞型肝障害モデルを構築することは、胆汁鬱滞肝毒性を起 こす候補化合物の早期発見を可能にし、非臨床試験における毒性試験の段階にお いて、必要不可欠なモデルを提供できると思われる。 サンドイッチ培養法とは、細胞をコラーゲンゲルやマトリゲルなどの細胞外マトリック ス (ECM) で挟んで培養する方法である。この方法により、三次元構造が形成され、 比較的長期間肝機能維持することが可能になり、細胞の極性が認められるとの報告が ある[7]。また、胆管腔が形成するとともに胆管側膜上に排出トランスポーターが局在化 することも知られている[8]。さらに、HHs を I 型コラーゲンがコートされているプレート上 に播種し、上層からマトリゲルで覆ったサンドイッチ培養法にて、細胞間に微小胆管の 形成促進が認められたとの報告もある[1]。したがって、サンドイッチ培養法は,薬物の 胆汁排泄を評価するための方法に適していると考えられる。ECM はシグナル分子や 足場として肝細胞を支持する役割があり、インテグリンを介して直接的、または間接的 に遺伝子発現を変動させる可能性がある[9]。肝細胞の機能向上に対する効果を期待 してよく用いられている ECM として、コラーゲンやマトリゲルが知られている。コラーゲ ンの中でも I 型コラーゲンは、特有の物理的・生物学的性質を有するため、様々な細 胞培養に汎用されている。また、マトリゲルはマウス Engelbreth-Holm-Swarm (EHS) 肉腫由来のゲルで、ECM の主要な成分であるラミニン-111 や IV 型コラーゲン、エン タクチンなどに加え、transforming growth factor (TGF)-β や FGF など様々な増殖因子 も含んでいる。肝細胞をマトリゲル上で培養を行なうことにより立体的な形態をもたらす
3 こと、またマトリゲルの主要成分であるラミニンが肝特異的タンパク質の発現を促進す るなど、肝機能向上には、マトリゲルの性質が大きく関与していることが知られている [10]。 したがって、本研究は、I 型コラーゲンとマトリゲルを用いたサンドイッチ培養法による in vitro 薬剤性胆汁鬱滞型肝障害モデル系の構築を行うことを目的とした。
Figure 1. Mechanism of hepatic of bile acids
Na⁺-taurocholate cotransporting polypeptide (NTCP), bile salt export pump (BSEP), and multidrug resistance-associated protein 2/3/4 (MRP2/3/4) play major roles in the uptake or excretion of bile acids (BAs) in hepatocytes. NTCP is modulates up to 80% of the uptake of conjugated BAs and 50% of the uptake of unconjugated BAs in hepatocytes. The farnesoid X-receptor (FXR), a known nuclear receptor of BSEP and MRP2. The CYP7A1 is inhibited by LRH-1, and then, BAs are not de novo synthesized from cholesterol.
4 第二章 HepG2-hNTCP-C4 細胞のサンドイッチ培養による胆汁鬱滞肝毒性評 価 2.1 緒言 肝臓は、薬物の代謝を担う重要な臓器であり、薬物による医薬品の毒性標的となる ことが多い。医薬品の非臨床安全性試験として実験動物を用いた反復投与毒性試験 が実施されるが、これらの毒性試験では肝毒性を示さなかった医薬品候補物質が臨 床試験段階や市販後にヒトで肝障害を引き起こし問題となることがある。そのため、近 年、肝臓に対して安全性の高い医薬品候補化合物を創製するために、動物での毒性 試験に加えて、培養ヒト肝細胞等のヒト由来材料を用いた毒性評価を非臨床試験の初 期段階において求められるケースがある。 Susukida らによって、サンドイッチ培養したラット肝細胞、HHs、HepaRG 細 胞あるいはHepG2 細胞を使用した胆汁鬱滞肝毒性評価系は、臨床的リスクを予 測するツールになり得ることが報告されている[11, 12]。このin vitro 胆汁鬱滞肝障
害評価系は、ヒト血清に含まれる12 種類の胆汁酸 (Cholic acid (CA)、Chenodeoxy
cholic acid (CDCA)、Glycochenodeoxycholic acid (GCDCA)、Deoxycholic acid (DCA)、 Lithocholic acid (LCA)、Ursodeoxycholic acid (UDCA)、Glycocholic acid (GCA)、 Glycodeoxycholic acid (GDCA)、Taurocholic acid (TCA)、Taurochenodeoxycholic acid (TCDCA)、Taurolithocholic acid (TLCA)、Tauroursodeoxycholic acid (TUDCA))と肝
障害を起こすことが知られている22 種類の化合物を添加することによって行わ れた。しかしながら、これらの細胞による胆汁鬱滞肝障害評価系は、それぞれに おいて種差、ロット差、使用の制限、および生理的状態を再現していない点が問題と なる。医薬品開発における胆汁鬱滞肝毒性に利用可能な肝細胞の条件として、ト ランスポーターなどの発現や機能が高いこと、再現性があること及び安定供給 が可能であることが挙げられる。したがって、このモデルを開発するにあたって、
5 容易に扱え、十分なトランスポーター発現や活性を有した細胞を用いる必要が あり、また、それを作出することも重要である。HepG2 細胞に代表される肝がん 細胞株は、安定した遺伝子発現、実験操作が簡易的である特徴があり、初代ヒト肝細 胞の代わりによく使用されている。しかし、このような細胞株は一部のトランスポーター の遺伝子発現が低いといった問題があることから、渡士博士らは、ヒト NTCP 遺伝子を 持続的に発現するように設計したHepG2 細胞 (HepG2- hNTCP-C4 細胞) を確立した [13]。NTCP は Na +依存的に胆汁酸を取り込むトランスポーターであり、肝細胞内に 80%の抱合胆汁酸と 50%の非抱合胆汁酸の取り込みを行うことが知られている[14]。 そこで本章では、NTCP を安定発現した HepG2-hNTCP-C4 細胞を用い、ECM に よるサンドイッチ培養法にてin vitro 胆汁鬱滞型肝毒性評価の検討を行い、HepG2 細 胞を用いた場合と比較した (Fig. 2)。
Figure 2. In vitro cholestatic drug-induced liver injury evaluation system using sandwich culture HepG2 or HepG2-hNTCP-C4 cells. Sandwich cultured HepG2 and HepG2-hNTCP-C4 (SC-HepG2 and SC-HepG2-hNTCP-C4) cells were exposed to 2 2 test compounds in the presence or absence of BAs for 24 h. All test compounds were evaluated at a concentration of 50 μM, except for CsA, which was tested at 10 μM, to avoid drug-induced cytotoxicity.
6
2.2 実験方法 2.2.1 試薬及び細胞
ヒト肝ガン由来細胞株HepG2 細胞は、愛知学院大学の中島健一博士によってご
供与いただいた。 国立感染症研究所の HepG2-hNTCP-C4 細胞は、渡士博士より
ご供与頂いたものであり、TransIT-LT1 (Mirus) を使用して HepG2 細胞に hNTCP
の発現プラスミドをトランスフェクションすることにより確立された。また、 HepG2-hNTCP-C4 細胞は、400 μg/mL G418 (Nacalai) 含有培地により培養することで セレクションされた[13]。凍結保存されたHHs (ロット番号 HPCH10 / 1310262;年齢 20〜71 歳のドナー10 人 (男性 5 人、女性 5 人) のプール) は、XenoTech (カン ザス州レネクサ、米国) から入手した。試薬類を以下に示す (Table 1.)。また、そ の他の試薬はすべて市販の特級品を用いた。
7
Table 1. Reagents and manufacturer
販売元 品名 場所
Abcam anti-BCRP polyclonal antibodies ケンブリッジ、英国
BD Biosciences Matrigel matrix Growth Factor Reduced (GFR-Matrigel) マサチューセッツ州ベッドフォード、米国 Beckman Coulter Agencort RNAdvance Tissue Kit カリフォルニア州ブレア、米国
Biological Industries
L-glutamine (L-Glu)
non-essential amino acids (NEAA) penicillin-streptomycin (PS)
ベイト・ヘメク、イスラエル
Biopredic
Thawing medium without additives basal hepatic cell medium
basal hepatic cell medium containing additives
レンヌ、フランス
Bioss anti-MRP2 polyclonal antibodies ボストン、マサチューセッツ州、英国
Biowest fetal bovine serum (FBS) ニュアイエ、仏国
COSMO BIO Co. Cosmedium 004 東京、日本
Kapa Biosystems KAPA SYBR FAST qPCR Kit Master mix ABI Prism マサチューセッツ州ボストン、米国
KOKEN atelocollagen (collagen type I) 東京、日本
Sigma-Aldrich Co. anti-NTCP monoclonal antibodies
dimethyl sulfoxide (DMSO) ミズーリ州セントルイス、米国
Thermo Fisher Scientific
Alexa Fluor 488 donkey anti-rabbit IgG Alexa Fluor 488 goat anti-mouse IgG rhodamine phalloidin
8
タカラバイオ Dulbecco’s phosphate buffered saline without calcium,
magnesium (PBS) 滋賀、日本
東洋紡 ReverTra Ace qPCR RT Master Mix 大阪、日本
富士フィルム和光純薬工業
Cyclosporin A (CsA)
Dulbecco's Modified Eagle Medium (DMEM) DMEM/Ham’s F-12 medium (DMEM/F12) mouse monoclonal anti-human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody
9
2.2.2 細胞培養
HepG2 細胞は、ゼラチンでコーティングされた培養皿で培養した後、trypsin ethylenediaminetetraacetic acid (トリプシン EDTA) で剥離し、アテロコラーゲンで
コーティングされた96 または 24 ウェルプレートまたは 8 ウェルチャンバース ライドに、それぞれ0.5、2.4、または 1.0×105細胞/ウェルの密度で継代した。播 種4 時間後、培地を吸引し、GFR-Matrigel (0.25 mg/mL) を含む氷冷 Cosmedium 004 を上部から覆いサンドイッチ培養を行った。培地交換は、毎日 Cosmedium 004 を使用して行った。HepG2-hNTCP-C4 細胞は、HepG2 細胞と同様の方法を使 用して培養した。またHHs は、添加物なしの解凍用培地にて解凍し、肝細胞播 種用培地を用いコラーゲンタイプI でコーティングされたプレートに播種した。 その後、36 時間培養した。 2.2.3 RT-qPCR 解析
サンドイッチ培養したHepG2 HepG2) 細胞および HepG2-hNTCP-C4
(SC-HepG2-hNTCP-C4) 細胞は、3 日間培養後に Agencort RNAdvance Tissue Kit の添
付マニュアルに従って総 RNA を抽出した。RNA 量は、超微量紫外可視分光光
度計NanoDrop One (Thermo Fisher Scientific、マサチューセッツ州ウォルサム、米
国) を 用 い て 測定し た 。 cDNA 合 成 は ReverTra Ace qPCR RT Master Mix (TOYOBO、大阪、日本) を使用し 0.5 μg の total RNA から添付マニュアルに従
い、サーマルサイクラーを用いて37°C にて 15 分間、50°C にて 5 分間、98°C に
て5 分間処理することで行った。リアルタイム RT-PCR は KAPA SYBR Fast qPCR
Kit (Genetics、東京、日本) を使用し、最終容量 10 μL とした。反応は、Eco リア
ルタイムPCR システム (Illumina Inc.、サンディエゴ、カリフォルニア、米国) を
10 て 31 秒間のサイクルを 40 サイクル行い検出した。結果は内在性コントロール としてグリセルアルデヒド 3-リン酸デヒドロゲナーゼを用いて補正し、算出し た。ポジティブコントロールとして、凍結ヒト肝細胞解凍直後 (HHs 0 h) 及び凍 結ヒト肝細胞 48 時間培養後 (HHs 48 h) を用いた。この実験で使用したプライ マーをTable 2.に示す。
Table 2. Primers used for qPCR
2.2.4 蛍光免疫染色
SC-HepG2 および SC-HepG2-hNTCP-C4 細胞をリン酸緩衝生理食塩水 (PBS)
で2 回洗浄後、4%paraformaldehyde を用いて室温 (24 ± 4℃) で 15 分間固定処理
し、0.1% Triton X-100 を用いて室温にて 10 分間膜透過処理を行った。その後、 1%ウシ血清アルブミンを用いて 30 分間室温 (24 ± 4℃) にてブロッキング処理 を行った。ブロッキング処理後、一次抗体は、anti-NTCP monoclonal antibodies (1:50) 、 anti-BCRP polyclonal antibodies (1:50) 、 ま た は anti-MRP2 polyclonal antibodies (1:50) を用いて、4°C 遮光下で一晩反応させた。次に、二次抗体は、
Gene Names
Forward primer sequences (5'-3')
Reverse primer sequences (5'-3')
BCRP AGATGGGTTTCCAAGCGTTCAT CCAGTCCCAGTACGACTGTGACA
BSEP TGAGCCTGGTCATCTTGTG TCCGTAAATATTGGCTTTCTG
CYP7A1 CAGAACTGAATGACCTGCCA GGTGCAAAGTGAAATCCTCC
FXR ACTGACCTGTGAGGGGTGTA TGCCCCCGTTTTTACACTTG
GAPDH GAGTCAACGGATTTGGTCGT GACAAGCTTCCCGTTCTCAG
MDR1 CCCATCATTGCAATAGCAGG TGTTCAAACTTCTGCTCCTGA
MRP2 ACAGAGGCTGGTGGCAACC ACCATTACCTTGTCACTGTCCATGA
MRP3 GTCCGCAGAATGGACTTGAT TCACCACTTGGGGATCATTT
MRP4 GGCGAATTGTTAGCTGTGGT CAGGGCTGCTGAGACACATA
11
Alexa Fluor 488 donkey anti-rabbit IgG (1:250) または Alexa Fluor 488 goat anti-mouse IgG (1:250) を用いて遮光下室温にて 60 分間反応させた。また、F-アクチン染色 時には二次抗体溶液にrhodamine phalloidin (1:500) を添加した。核染色は 1 µg/mL 4',6-diamidino-2-phenylindole (DAPI) を遮光下室温にて 15 分間処理することで 行った。細胞を共焦点レーザー走査顕微鏡 (LSM 510; Carl Zeiss、オーバーコッ ヘン、ドイツ) にて観察した。 2.2.5 蛍光基質を用いた胆汁酸トランスポーターの機能活性 タイトジャンクションを維持するために、Ca2 +とMg2 +を含むハンクス平衡塩 溶液 (HBSS) を使用した。SC-HepG2 および SC-HepG2-hNTCP-C4 細胞を HBSS で2 回洗浄し、温めた HBSS とともに 10 分間保存した。続いて、NTCP の基質 で あ る N- (24-[7- (4-N,Ndimethylaminosulfonyl-2,1,3-benzoxadiazole) ]amino-3α,7α,12α-trihydroxy-27-nor-5β-cholestan-26-oyl) -2ʹ-aminoethanesulfonate (tauro-nor-THCA-24-DBD, Genomembrane、ケンブリッジ、英国) または MRP2 の基質で ある5- (and-6) -carboxy-2',7'-dichlorofluorescein diacetate (CDFDA、Thermo Fisher Scientific、マサチューセッツ州ウォルサム、米国) をそれぞれ 10 µM ずつ加えた 後、37°C、5% CO2 に設定したIN Cell Analyzer 6000 (GE Healthcare、リトル・チ
ャルフォント、英国)
を使用して細胞を観察した。次に、培地を除去し、tauro-nor-THCA-24-DBD の細胞内蓄積率を調べるために、Ca2+、Mg2 を含む冷した
HBSS で細胞を 3 回洗浄した。その後、1% Triton X-100 を含む HBSS で細胞を溶
解した。さらに、NTCP 阻害剤である 10 µM cyclosporin A (CsA) は NTCP 阻害の
評価を行うために使用した。溶解物の蛍光強度は、Synergy 2 modular multi-mode reader (BioTek、バーモント州ウィヌースキー、米国) を使用して評価した。
12
2.2.6 Cyclosporin A、MK571 による胆汁酸濃度依存的な細胞毒性評価
添加する胆汁酸は、ヒト血清胆汁酸成分に従って 12 種類選択した (Table 3.)
[15]。トランスポーターを阻害する化合物として、10 µM CsA および MK571 を使
用した。CsA および MK571 濃度は、BSEP または MRP2/3/4 の 50%阻害濃度 (IC50) および阻害定数 (Ki) 値に基づいて決定した[16, 17]。10 mM CsA、10 mM
MK571 および胆汁酸は、ストック溶液として DMSO に溶解した。各分析では、
対照群と化合物群では同じ DMSO 濃度となるように添加し、最終濃度は常に
0.5%未満に設定した。SC-HepG2 および SC-HepG2-hNTCP-C4 細胞に、胆汁酸の
存在下または非存在下で24 時間 10 µM CsA または 10 µM MK571 を添加した後、
LDH-Cytotoxic Detection Kit を使用して、損傷細胞から放出された乳酸デヒドロ
ゲナーゼ (LDH) の活性 (LDHsample) を測定することにより細胞毒性を評価した。
細胞における LDH 活性のポジティブコントロールでは、0.25% TritonX-100
(LDHTritonX-100) を含む Cosmedium 004 を用いた。細胞毒性は、次の式を使用して
算出した。
Cell toxicity (%) = (LDHsample – LDHblank) / (LDHTritonX-100 – LDHblank) × 100.・・・(1)
LDHblank値は、未処理のSC-HepG2 および SC-HepG2-hNTCP-C4 細胞における値
13
Table 3. Standard BA constituents in human serum.
2.2.7 薬剤性胆汁鬱滞肝毒性評価 SC-HepG2 および SC-HepG2-hNTCP-C4 細胞に、胆汁酸の存在下または非存在 下で24 時間、22 種類の化合物をそれぞれ添加した[11, 17]。22 種類の化合物の選 択は、Susukida らの報告によると医 薬 品 医 療 機 器 総 合 機 構 (PMDA) の医薬 品副作用データベース(2004.4-2013.8)より、全 1,866,993 件の報告から第一被疑 薬のみをまず抽出し、医薬品 (一般名) ごとに肝細胞障害型 DILI および胆汁うっ 滞型DILI 関連有害事象件数を集計 (1,984 薬物) した。そして、全 DILI 件数が少な すぎる医薬品を除き (243 薬物)、【胆汁うっ滞型 DILI の割合(%)=胆汁うっ滞型 DILI
件数(( ALP、γ-GTP、ビリルビン値の上昇 etc.))/全 DILI 件数】の式に基づいて胆汁う
っ滞型 DILI のリスクの大小を算出した (38 化合物)。さらに、ラット肝細胞での胆汁鬱
滞肝毒性評価の結果を元に、22 種類の化合物まで絞り込んだ。薬物誘発性の細胞
毒性を避けるために、10 µM CsA を除き、全ての化合物を 50 µM の濃度にて評
価した。相対的な細胞毒性 (%) は、計算式 (1)の LDHblankをDMSO で処理した
Bile acids 1×standard concentration (µM)
Cholic acid (CA) 0.3
Chenodeoxy cholic acid (CDCA) 0.5
Glycochenodeoxycholic acid (GCDCA) 2.6
Deoxycholic acid (DCA) 1.1
Lithocholic acid (LCA) 0.045
Ursodeoxycholic acid (UDCA) 0.17
Glycocholic acid (GCA) 0.62
Glycodeoxycholic acid (GDCA) 0.57
Taurocholic acid (TCA) 0.07
Taurochenodeoxycholic acid (TCDCA) 0.32
Taurolithocholic acid (TLCA) 0.13
14
SC-HepG2 および SC-HepG2-hNTCP-C4 細胞の値である LDHvehicleに置き換える
ことで算出した。胆汁鬱滞肝毒性は、次の式を使用した。 Cholestatic cytotoxicity (%)
= relative cell toxicity (%) +BAs – relative cell toxicity (%) -BAs. ・・・・・・・・・(2)
ここで、胆汁鬱滞肝毒性 (%) は、胆汁酸存在下と非存在下での相対的な細胞毒 性を差し引いた値により算出した。
2.2.8 統計学的解析
2 群間の比較は Student's t-test により行った。多重比較は、分散分析を行った 後、Tukey’s HSD (honestly significant difference) test によって行った。その際の統 計分析は、SPSS Statistics software package, version 13.0J (IBM Japan、東京) を用い
て行った。データは、3 ウェルの平均±標準偏差として表した。p 値が 0.05 未満
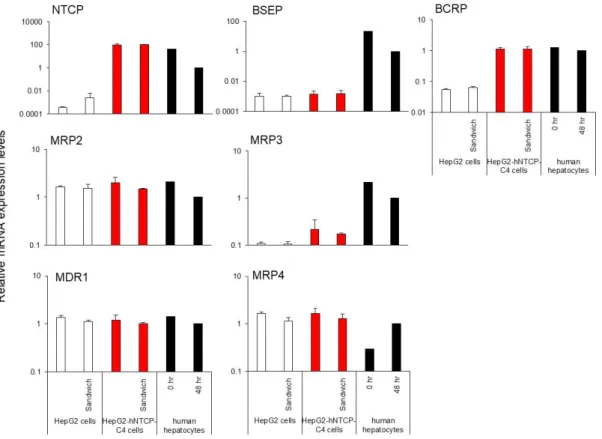
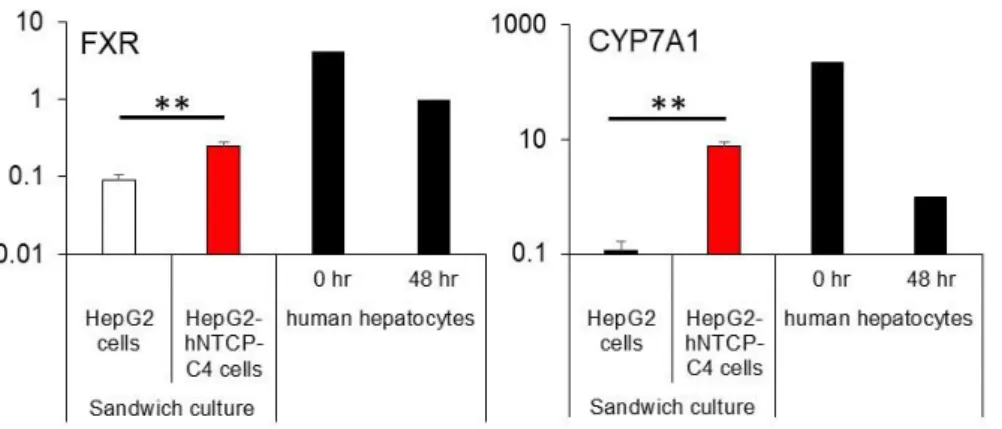
15 2.3 結果 2.3.1 胆汁酸トランスポーターに関与する遺伝子発現 基底膜側および頂端側に発現するトランスポーターのmRNA 発現量を測定し た。トランスポーターのmRNA 発現量は、単層培養とサンドイッチ培養された HepG2-hNTCP-C4 細胞に有意な差は認められなかった (Fig. 3)。ただし、SC-HepG2-hNTCP-C4 細胞における NTCP、BCRP、MRP3、FXR、および CYP7A1 の mRNA 発現レベルは、SC-HepG2 細胞の mRNA 発現量よりも有意に高値を示し た(Fig. 3, 4)。MRP2、MRP4、および多剤耐性タンパク質 1 (MDR1) の mRNA 発
現量は、SC-HepG2 細胞および SC-HepG2-hNTCP-C4 細胞において同等であった。
しかし、両細胞株において、BSEP の mRNA 発現レベルは低値を示した (Fig. 3) 。
Figure 3. Relative expression of mRNAs encoding the basal or apical transporters according to glyceraldehyde 3-phosphate dehydrogenase expression in HepG2 and HepG2-hNTCP-C4 cells cultured in sandwich configuration or monolayer configuration .
Each bar represents the mean ± standard deviation (n = 3). ** p < 0.01. The graph represents gene expression relative to that detected in HPHs at 48 h.
16
Figure 4. Relative expression of mRNAs encoding the FXR, and CYP7A1 according to glyceraldehyde 3-phosphate dehydrogenase expression in SC-HepG2 and SC-HepG2-hNTCP-C4 cells.
Each bar represents the mean ± standard deviation (n = 3). **p < 0.01. The graph represents gene expression relative to that detected in HPHs at 48 h.
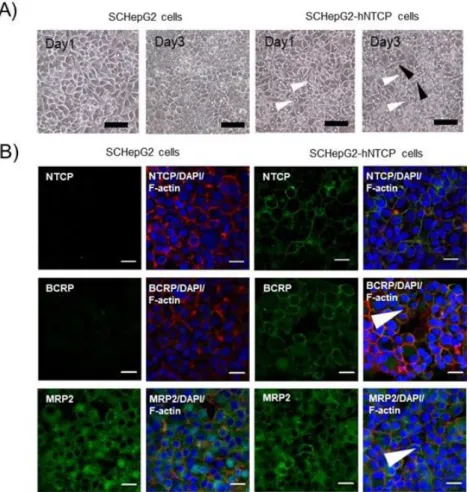
2.3.2 細胞の形態およびタンパク質の局在 SC-HepG2 細胞および SC-HepG2-hNTCP-C4 細胞において、基底膜側および頂 端側に発現するトランスポーターのタンパク質が発現しているのかを検討した。 SC-HepG2-hNTCP-C4 細胞内においては、空胞が認められたが、SC-HepG2 細胞 では認められなかった (Fig. 5A)。また、SC-HepG2-hNTCP-C4 細胞では NTCP、 BCRP、 MRP2 および細胞骨格 F-アクチンのタンパク質は細胞膜上で発現して いた。SC-HepG2 細胞において、F-アクチンのタンパク質は発現していたものの、 NTCP および BCRP の発現は認められなかった (Fig. 5B)。一方、BSEP のタンパ ク質発現量は認められなかった。また、両細胞において、MRP2 が細胞質に僅か に内在化していることが示唆された。これらタンパク質の発現量は、mRNA 発 現量と相関していた。
17
Figure 5. Cell morphology and transporter localization in SC -HepG2 and SC-HepG2-hNTCP-C4 cells.
(A) Morphological images of sandwich-cultured HepG2 and HepG2-hNTCP-C4 cells. Images are shown after 1 and 3 days of culture. White or black arrowheads show a part of the vacuoles or bile canaliculi, respectively. Scale bar, 100 µm. (B) Immunofluorescence staining for NTCP, BCRP, and MRP2 in SC-HepG2 and SC-HepG2-hNTCP-C4 cells. Cell nuclei were stained with DAPI (blue). Cytoskeletal F-actin was stained with rhodamine-phalloidin (red). NTCP, BCRP, and MRP2 were stained with Alexa Fluor 488 goat anti-mouse IgG (green). White arrowheads show a part of the bile canaliculi. Scale bar, 20 µm.
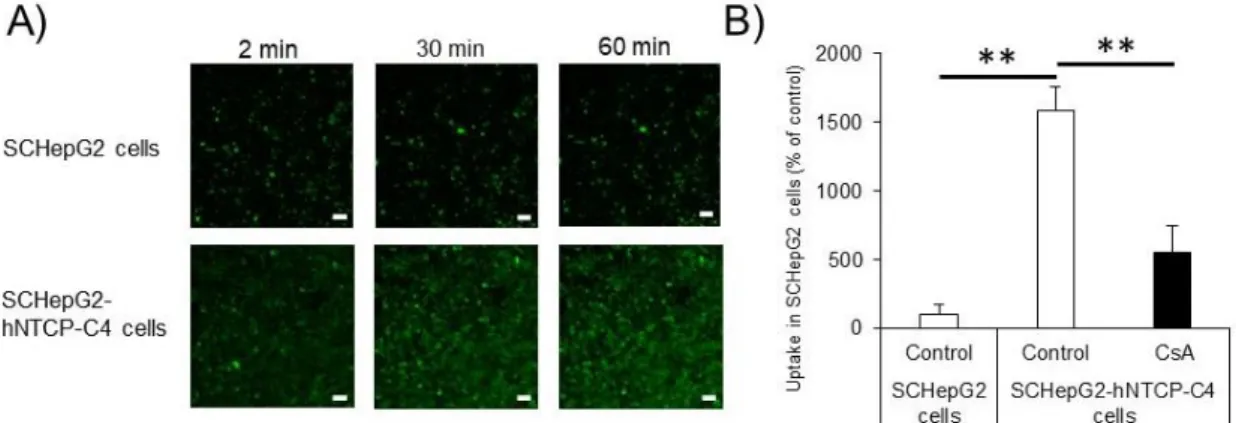
18 2.3.3 胆汁酸取り込みトランスポーターの機能評価 Tauro-nor-THCA-24-DBD を使用して、NTCP トランスポーターの機能を評価し た。SC-HepG2-hNTCP-C4 細胞において、細胞内の tauro-nor-THCA-24-DBD 蓄積 量は、時間依存的に増加した (Fig. 6A)。また、その 10 分後の細胞内取り込み量 はNTCP の阻害剤である CsA 処理により優位に減少した (Fig. 6B)。しかし、SC-HepG2 細胞における tauro-nor-THCA-24-DBD 蓄積量は時間依存的に変化しなか った (Fig. 6A)。したがって、SC-HepG2-hNTCP-C4 細胞は機能の備わった NTCP が発現していると考えられた。
Figure 6. Functional activities of NTCP in SC-HepG2 and SC-HepG2-hNTCP-C4 cells. (A) NTCP activity was determined using tauro-nor-THCA-24-DBD. Scale bar, 50 µm. (B) Intracellular accumulation of tauro-nor-THCA-24-DBD. Each bar represents the mean ± standard deviation (n = 3). **p < 0.01. 2.3.4 胆汁酸トランスポーターの機能評価 CDFDA を使用して、MRP2 トランスポーターの機能を評価した。SC-HepG2 細 胞におけるCDF の細胞内蓄積量は細胞間のドットが時間依存的に増加し、毛細 胆管に排出していることも確認できた (Fig. 7A)。一方、SC-HepG2-hNTCP-C4 細 胞ではCDFDA 処理 30 分後にその蓄積量は最も増加し、その後徐々に減少した
(Fig. 7A)。Fig. 7B は、SC-HepG2-hNTCP-C4 細胞の毛細胆管の拡大図を示した。 SC-HepG2-hNTCP-C4 細胞でも SC-HepG2 細胞と同様に、CDF が毛細胆管に蓄積
19
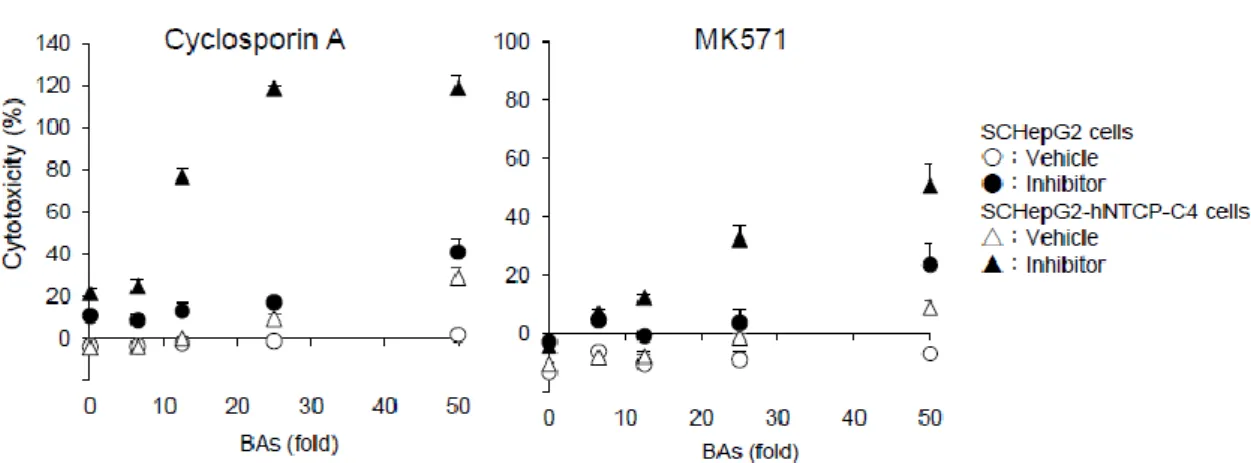
Figure 7. Functional activities of MRP2 in SC-HepG2 and SC-HepG2-hNTCP-C4 cells. (A) Excretion of CDF into the bile canaliculi via MRP2. Scale bar, 50 µm. (B) Enlarged view showing the bile canaliculi in the red frame in (A). White arrows show a part of the bile canaliculi. Scale bar, 50 µm. 2.3.5 胆汁酸濃度依存的な細胞毒性評価 SC-HepG2 細胞および SC-HepG2-hNTCP-C4 細胞において、胆汁鬱滞肝毒性の ヒト血清胆汁酸濃度を決定するために、CsA および MK571 を用いて細胞毒性評 価を行った。両細胞は、ヒト血清胆汁酸濃度依存的に細胞毒性を増強させた。 SC-HepG2-hNTCP-C4 細胞および SC-HepG2 細胞における胆汁鬱滞肝毒性は、25 倍お よび50 倍のヒト血清胆汁酸濃度において、vehicle 群と CsA 処理群間の差が最も大き く、これらの濃度の胆汁酸を用いることで細胞毒性を感度良く検出できる可能性が示 唆された。なお、この条件において、胆汁酸自身 (vehicle 群) の細胞毒性は認めら れなかった。さらに、50 倍濃度のヒト胆汁酸を加えて CsA で処理した群では、 SC-HepG2-hNTCP-C4 細胞において SC-HepG2 細胞よりも約 3 倍の細胞毒性が認
20
められた。これらの結果から、SC-HepG2-hNTCP-C4 および SC-HepG2 細胞に添
加するヒト血清胆汁酸濃度をそれぞれ25 倍および 50 倍の濃度に決定した (Fig.
8)。
Figure 8. Concentration-dependent BA cytotoxicity in SC-HepG2 and SC-HepG2-hNTCP-C4 cells. SCHepG2 and SCHepG2-hNTCP-C4 cells were exposed to 10 µM CsA or 20 µM MK571 in the presence or absence of BAs for 24 h. BA concentrations were changed by 6.25-, 12.5-, 25-, and 50-fold. Cell toxicity (%) = (LDHsample – LDHblank) / (LDHTritonX-100 – LDHblank) × 100. Cells treated with DMSO were used as the vehicle. Data are means ± standard deviations (n = 3).
2.3.6 薬剤性胆汁鬱滞肝毒性評価 両細胞における胆汁鬱滞肝毒性は、胆汁鬱滞肝毒性の相対値から肝細胞毒性 の相対値を除いた値にて算出した。また、22 種類の化合物は、トランスポータ ーを阻害し肝障害を起こすことが知られている。SC-HepG2-hNTCP-C4 細胞に おいて、Fexofenadine や Voriconazole を除いたすべての化合物は、胆汁鬱滞肝 毒性を示した。しかし、SC-HepG2 細胞では、いくつかの化合物において胆汁 酸依存性肝細胞毒性を示さなかった。SC-HepG2-hNTCP-C4 細胞は、全ての化 合物において、SC-HepG2 細胞の 1~50 倍ほどに検出感度が増加していること が示唆された。また、SC-HepG2-hNTCP-C4 細胞の Amiodarone や Atrvastatin は、薬の特異性による肝障害も生じている可能性が考えられた。したがって、
21
SC-HepG2-hNTCP-C4 細胞は薬剤性胆汁鬱滞型肝毒性をより多く検出することを可 能にした (Fig. 9)。
Figure 9. Drug-induced cholestatic cytotoxicity in SC-HepG2 and SC-HepG2-hNTCP-C4 cells. SC-HepG2 and SC-HepG2-hNTCP-C4 cells were exposed to 22 compounds for 24 h in the presence of BA at concentrations 50 or 25-fold higher than that in normal human serum. Data are means ± standard deviations (n = 3). Cholestatic cytotoxicity (%) = relative cell toxicity (%)+BAs – relative cell toxicity (%)-BAs. Relative cell toxicity (%) was obtained by replacing LDHblank with LDHvehicle
22 2.4 考察 胆汁鬱滞肝毒性評価系は、DILI のメカニズムを明らかにするために重要であ る。ヒトでは、NTCP、BSEP および MRP2/3/4 が主に肝臓の胆汁酸輸送に関与し ている[18]。また、サンドイッチ培養した肝細胞は、生体内の構造的および機能的 特性を維持し、胆汁酸取り込みと排出を同時に評価することが可能になること が報告されている[19]。したがって、本研究ではまずSC-HepG2 および SC-HepG2-hNTCP-C4 細胞における取り込みおよび排出トランスポーター遺伝子の発現を 検討した。SC-HepG2-hNTCP-C4 細胞の NTCP、MRP2 および MRP4 の mRNA 発
現量は、HHs (48 h) と同程度であった(Fig. 3)。しかし、BSEP の mRNA 発現量
は低値を示し、サンドイッチ培養をしていないHepG2 と HepG2-hNTCP-C4 細胞 と類似していた。この結果から、サンドイッチ培養法によって、in vivo の構造並 びに機能的特性を維持および向上させるとの報告がある[7]が、BSEP の mRNA 発 現量の増加には関与していないことが示唆された (Fig. 3)。さらに、胆汁酸をリガンド とするファルネソイドX 受容体 (FXR) [20] の mRNA 発現量は、HepG2 細胞と SC-HepG2-hNTCP-C4 細胞で発現していた (Fig. 4)。しかし、その発現量は HHs (48 h) よりも低値を示していたことから、BSEP の mRNA 発現を誘導するには十分 ではなかった可能性が考えられる。また、SC-HepG2 細胞と比較して、SC-HepG2-hNTCP-C4 細胞では BCRP、MRP3、および CYP7A1 の mRNA 発現量が有意に増 加していた(Fig. 3)。これは、一部の胆汁酸が NTCP によって SC-HepG2-hNTCP-C4 細胞内に取り込まれたため、CYP7A1、FXR および MRP3 の発現が誘導され た可能性も考えられる。SC-HepG2-hNTCP-C4 細胞におけるこれらのトランスポ ーターの誘導メカニズムを解明するには、さらなる研究が必要である。 高密度で細胞を培養することで、細胞同士が密接に接着した状態になるため機能 を高めるとの報告がある[21]。そこで、本研究では両者において、培養皿に均一に満遍
23 なく敷き詰められた状態であった3 日間培養したものを評価・解析に用いた (Fig. 5A)。 SC-HepG2-hNTCP-C4 細胞において、細胞骨格 F-アクチン、BCRP、および MRP2 タンパク質は発現が認められた (Fig. 5B)。以前の報告において、MRP2 と F-アクチン が共局在すること で機能を有することが 示されている[19]。そのため 、 SC-HepG2-hNTCP-C4 細胞では、BCRP および MRP2 が F-アクチンと共局在しており、機能を有 することが確認された。 SC-HepG2 および SC-HepG2-hNTCP-C4 細胞における NTCP および MRP2 の活 性を評価するために、tauro-nor-THCA-24-DBD および CDFDA を用いて検討を行っ た。SC-HepG2-hNTCP-C4 細胞における tauro-nor-THCA-24-DBD の細胞内蓄積量
は、時間依存的に増加し、10 μM CsA の処理により有意に抑制された (Fig. 6A, B)。 Tauro-nor-THCA-24-DBD は、NTCP や BSEP の基質である。また、CsA における NTCP の IC50は、NTCP-HEK293 細胞で約 10.3 μM であり、BSEP の IC50は、ヒト肝細 胞で 0.5 μM であることが報告されている[22, 23]。一方、SC-HepG2-hNTCP-C4 細胞の BSEP は、発現レベルが非常に低いことから、関与していないと考えられた。したがっ て、SC-HepG2-hNTCP-C4 細胞において、CsA は主に NTCP 阻害に大きく寄与して いる可能性が示唆され、NTCP を介した輸送活性を有することが認められた。また、 CDFDA は、受動輸送によって細胞内に取り込まれた後、エステラーゼによって加水 分解され、CDF になる。その後、MRP2 および MRP3 によって排出されることが知られ ている[24]。本研究において、SC-HepG2-hNTCP-C4 細胞内への CDF の蓄積量は急
速に増加し、その後すぐに減少した (Fig. 7A, B)。したがって、SC-HepG2-hNTCP-C4
細胞は、SC-HepG2 細胞と同様に MRP2 の機能が備わっていることが示唆された。ま
た、SC-HepG2-hNTCP-C4 細胞は SC-HepG2 細胞と比較して、より早く CDF に変換さ
れていることから、高いエステラーゼ活性を持つ可能性が考えられた。さらに、MRP3
24
トランスポーターを介した胆汁酸の輸送阻害によって、引き起こされる肝毒性の評価 を行ったところ、同じ胆汁酸濃度において、SC-HepG2-hNTCP-C4 細胞における CsA
および MK571 による細胞毒性は、SC-HepG2 細胞での細胞毒性と比較して著しく増
加した (Fig. 8)。用いた CsA には、BSEP や MRP2/3 以外に胆汁酸トランスポーター
である MRP4 を阻害することも知られている[25]。SC-HepG2-hNTCP-C4 細胞における MRP4 の機能解析は検討していないため、肝毒性の正確な予測のためにはさらなる 研究が必要である。 22 種類の肝毒性を引き起こす化合物を添加した SC-HepG2-hNTCP-C4 および SC-HepG2 細胞は、25 倍および 50 倍のヒト血清胆汁酸存在下にて検討された。 SC-HepG2-hNTCP-C4 細胞では、ほぼ全ての化合物において胆汁鬱滞肝毒性が検
出されたが、SC-HepG2 細胞では一部検出されない化合物が存在した (Fig. 9)。Ellis
らの報告によると、スタチン類の一部はMRP2 を阻害するため、胆汁酸が排泄でき
ず、肝毒性を示すことが知られている[26]。本研究において、スタチン類は
SC-HepG2-hNTCP-C4 細胞において胆汁酸存在下で細胞毒性を検出したが、SC-HepG2
細胞では検出されなかった (Fig. 9)。SC-HepG2 および SC-HepG2-hNTCP-C4 細胞に
おけるMRP2 の遺伝子発現量は HHs (48 h) と同等であり、MRP2 活性も認められた。
また、SC-HepG2-hNTCP-C4 細胞は、NTCP の機能を有している。したがって、in vitro
胆汁鬱滞肝毒性評価系の構築には、NTCP が十分に発現している必要があり NTCP を介して胆汁酸が肝細胞内に十分に取り込まれることで、MRP2 の機能評価が可能に なることが示唆された。また、CsA を除いた全ての化合物は 50 µM の濃度を使用 している。SC-HepG2-hNTCP-C4 細胞を用いた胆汁鬱滞肝毒性評価は、SC-HepG2 細胞と比較し、MRP2 の IC50が50 µM 以下の化合物をより感度良く検出可能に した。したがって、SC-HepG2-hNTCP-C4 細胞を用いた胆汁鬱滞肝毒性は、より MRP2 を反映させた評価が可能であることを示唆した。
25
しかしながら、SC-HepG2-hNTCP-C4 細胞は BSEP の発現が低いため、薬剤性胆
26
2.5 小括
本研究では、SC-HepG2-hNTCP-C4 細胞を使用した胆汁鬱滞肝毒性評価につい
て検討を行った。その結果、NTCP を高発現させることで、胆汁鬱滞肝毒性をよ
27 第三章 サンドイッチ培養したヒト iPS 細胞由来肝細胞を用いた胆汁鬱滞肝毒 性評価の検討 3.1 緒言 高橋らによって確立されたヒト人工多能性幹細胞 (iPS 細胞) は、生体を構成する ほとんどのタイプの細胞に分化可能であることが知られており、さまざまなヒト細胞から 樹立された場合、異なる遺伝的背景を持つ特徴を有している[27]。また、ヒト iPS 細胞は、 再生医療だけでなく、創薬研究にも適していると期待されている。
Kondo らは、バルプロ酸 (VPA) を用いて、薬物動態機能を持つヒト iPS 細胞由来
肝細胞に分化させる効率的な方法を確立している[28]。その他にも、Takayama らは発 生の過程で肝細胞への分化および成熟に関与する転写因子類をアデノウイルスベク ターによって導入し、ヒト iPS 細胞由来肝細胞を作製する方法を報告している[29]。この ように、近年さまざまな肝分化誘導法が報告されており、成熟化という課題は徐々に克 服されつつある。 ヒト iPS 細胞由来肝細胞は、薬物代謝に起因した肝細胞毒性あるいは相互作用に 用いられている報告はある[30]が、排出トランスポーターの阻害により生じる肝毒性に着 眼した報告はほとんどない。これらのトランスポーターの阻害によって、肝細胞内に胆 汁酸が蓄積し、胆汁鬱滞肝障害を起こすことが知られている。薬剤性胆汁鬱滞肝障 害は、薬剤性肝障害 (DILI) の一つであり、臨床的に非常に重要な疾患である。した がって、本研究では、Kondo らの肝分化プロトコールにより分化誘導したヒト iPS 細胞 由来肝細胞 (HiHs) にサンドイッチ培養を行うことで、胆汁鬱滞肝毒性評価が可能で あるのか検討を行った。
28
3.2 実験方法 3.2.1 試薬及び細胞
フィーダー細胞は mouse embryonic fibroblast (MEF) を使用した。ヒト iPS 細胞 (Windy) は、ヒト胎児肺線維芽細胞 MRC-5 に OCT3/4、SOX2、KLF4、c-MYC 遺伝子 を、パントロピックレトロウイルスベクターを用いて導入後、ヒト ES 細胞様コロニーをクロ ーン化したものであり、国立成育医療研究センター梅澤明弘博士よりご供与いただい た。凍結保存されたヒト初代肝細胞 (HHs ; lot. HPCH10/ 0910463; 32〜76 歳の 10 人 のドナー) は、XenoTech (Lenexa、カンザス、米国) から入手した。試薬類を以下に示 す (Table 4.)。その他の試薬はすべて市販の特級品を用いた。
Table 4. Reagents and manufacturer
品名 販売元 場所
Abcam anti-MRP2 polyclonal antibodies ケンブリッジ、英国 Abnova anti-radixin monoclonal antibodies タイペイ、台湾
MS Techno Systems Accutase 大阪、日本
PeproTech Activin A
hepatocyte growth factor (HGF) ニュージャージー州ロッキーヒル、米国 Santa Cruz Biotechnology donkey anti-mouse IgG-HRP テキサス州ダラス、米国
Thermo Fisher Scientific KnockOut Serum Replacement (KSR)
KnockOut DMEM (KO-DMEM) マサチューセッツ州ウォルサム、米国
富士フィルム和光純薬工業
oncostatin M (OSM) dexamethasone (DEX) valproic acid (VPA)
大阪、日本 3.2.2 細胞培養 HepG2 細胞をゼラチンでコーティングされた 12、24 ウェルプレートに 4.2、 2.4×105細胞/ウェルの密度で播種し、Cosmedium 004 で培養した。また、HHs は、 添加物なしの解凍用培地にて解凍し、肝細胞播種用培地を用いコラーゲンタイ プ I でコーティングされたプレートに播種した。その後、36 時間培養した。そ
29 して、ヒトiPS 細胞はマイトマイシン C 処理を施した MEF (6×105 細胞/100 mm ディッシュ) 上に播種し、37°C、5% CO2インキュベーター中にて培養した。ヒ トiPS 細胞の継代は、3~5 日培養後、1:2~1:3 のスプリット比で行った。ヒト iPS 細胞は解凍48 時間後に培地を交換し、それ以降は毎日交換した。 3.2.3 ヒト iPS 細胞から肝細胞への分化誘導 ヒト iPS 細胞の培養および肝分化は、当研究室で確立された方法を用いて行
った[28]。ヒトiPS 細胞を 0.5% FBS、100 ng/mL activin A を含む RPMI + GlutaMax
培地で3 日間培養後、2% KSR、100 ng/mL activin A を含む RPMI + GlutaMax 培
地でさらに 2 日間培養することで内胚葉へと分化させた。分化誘導 5 日目に、
細胞を Accutase にて 5 分間処理することによって剥離し、あらかじめヒト iPS
細胞用培地にて 30 倍に希釈した GFR-Matrigel でコーティングした細胞培養用
24 ウェルプレートあるいは 96 ウェルプレートに 1×105 細胞/cm2で播種した。
細胞播種後、20% KSR、1% GlutaMax、0.1 mM NEAA、0.1 mM 2-ME、1% DMSO
を含むKO-DMEM で 7 日間培養することにより肝芽細胞へと分化させた。最後
に、10 ng/mL HGF、20 ng/mL OSM、100 nM DEX、2 mM VPA を含む Cosmedium 004 で 7 日間、2 mM VPA を除いた 10 ng/mL HGF、20 ng/mL OSM、100 nM DEX、 2 mM VPA を含む Cosmedium 004 で 3 日間、Cosmedium 004 で 3 日間培養するこ
とにより肝細胞へ分化させた。サンドイッチ培養は分化 20 日目に開始された。 細胞を0.25%トリプシン EDTA で剥離後、4.2、2.4、0.5 または 1.0×105 細胞/well を、それぞれアテロコラーゲンでコーティングされた 12、24 または 96 ウェル プレートまたは 8 ウェルチャンバースライドに播種した。播種 4 時間後、培地 を吸引し、GFR-Matrigel (0.25 mg / mL) を含む氷冷 Cosmedium 004 で細胞を覆っ た。培地は一日おきに交換した。
30 3.2.4 蛍光免疫染色 サンドイッチ培養したヒトiPS 細胞由来肝細胞 (SC-HiHs) は、PBS で 2 回洗 浄後、4%paraformaldehyde を用いて室温にて 20 分間固定処理し、0.1% Triton X-100 を用いて室温にて 10 分間膜透過処理を行った。その後、1% ウシ血清アル ブミンを用いて室温にて30 分間ブロッキング処理を行った。ブロッキング処理
後、一次抗体は anti-MRP2 polyclonal antibodies (1:50)、anti-radixin monoclonal antibodies (1:50) で一晩、4°C にて染色した。二次抗体は、Alexa Fluor 488 donkey anti-rabbit IgG (1:250) を用いて遮光下室温にて 60 分間反応させた。また、F-ア
クチン染色時には二次抗体溶液にrhodamine phalloidin (1:200) を添加した。核染
色は、TO-PRO3 (1:750) を遮光下室温にて 15 分間処理することで行った。染色
後のSC-HiHs を共焦点レーザー走査顕微鏡 (LSM 510 Carl Zeiss、オーバーコッ
ヘン、ドイツ) で観察した。
3.2.5 RT-qPCR 解析
1、3 または 5 日間 SC-HiHs、HiHs、HepG2 細胞および HepaRG 細胞を Agencort RNAdvance Tissue Kit の添付マニュアルに従い抽出した。RNA 量は、NanoDrop One (Thermo Fisher Scientific、マサチューセッツ州ウォルサム、米国) を用いて 測定し、ReverTra Ace qPCR RT Master Mix (TOYOBO、大阪、日本) を使用し 0.5 μg の total RNA から添付マニュアルに従い、サーマルサイクラーを用いて 37°C
にて 15 分間、50°C にて 5 分間、98°C にて 5 分間処理することで行った。
THUNDERBIRDTMTM定量的ポリメラーゼ連鎖反応 (qPCR) ミックス (東洋紡
績株式会社、大阪、日本) を使用して 40 ng の全 RNA から cDNA を生成した。
反応はEco リアルタイム PCR システム (Illumina Inc.、サンディエゴ、カリフォ
31
トロールとして β-アクチンを用いて補正し算出した。この実験で使用したプラ
イマーをTable 5.に示した。また、HHs (0 h) 及び HHs (48 h) を用いた。
Table 5. Primers used for quantitative polymerase chain reaction (qPCR)
3.2.6 ウェスタンブロッティング解析
HiHs、SC-HiHs および HepG2 細胞は、氷上にて 0.1% sodium dodecyl sulfate (SDS)、 プロテアーゼ阻害剤カクテルは、Sigma-Aldrich (東京、日本) を含む PBS (pH 7.6)で
溶解し、回収した。このライセート (各レーン 40 µg) を 7% SDS-ポリアクリルアミドゲル
(PAGE) 電 気 泳 動 後 、 PVDF メ ン ブ レ ン に 転 写 し た 。 一 次 抗 体 は 、 anti-MRP2 polyclonal antibodies (1:50) を 用い、4℃で一 晩保存 し た 。その後、 二次抗体 は donkey anti-mouse IgG-HRP (1:50) を用いて室温 (24 ± 4℃) にて 60 分間反応させ
た。検出及び解析はImageQuant TL は、GE Healthcare (バッキンガムシャー、英国)
を使用してデンシトメトリーを実施した。
3.2.7 蛍光 NTCP、BSEP および MRP2 プローブを用いた胆汁酸トランスポー
ターの機能活性
以前の報告にあるように、胆汁排泄はBiliary Excretion Index (BEI) 法を使用し
て評価した[15]。Ca2 +とMg2 +の有無によるHBSS は、胆管ネットワークを維持ま
たは崩壊するために使用した。SC-HiHs を温めた HBSS で 2 回洗浄し、10 分間 Gene
Names
Forward primer sequences (5'-3')
Reverse primer sequences (5'-3')
β-actin TTCAACACCCCAGCCATGTACG GTGGTGGTGAAGCTGTAGCC
BSEP TGAGCCTGGTCATCTTGTG TCCGTAAATATTGGCTTTCTG
MRP2 ACAGAGGCTGGTGGCAACC ACCATTACCTTGTCACTGTCCATGA
MRP3 GTCCGCAGAATGGACTTGAT TCACCACTTGGGGATCATTT
MRP4 GGCGAATTGTTAGCTGTGGT CAGGGCTGCTGAGACACATA
32
保存した。次に、SC-HiHs を NTCP、BSEP の基質である tauro-nor-THCA-24-DBD
を加え、37℃、15 分間反応させた。そして、NTCP、BSEP、MRP2、MDR1 など の複数のトランスポーターの阻害剤である10 µM CsA (富士フィルム和光純薬株 式会社、大阪、日本) を必要に応じて添加した[31, 32]。その後、培地を除去し、 SC-HiHs を Ca2 +とMg2 +を含む氷冷したHBSS で 3 回洗浄し、共焦点レーザー走査 顕微鏡 (LSM 510 Carl Zeiss、オーバーコッヘン、ドイツ) にて観察した。続いて、 MRP2 の基質である 10 µM CDFDA を加えた後、37°C、5% CO2 に設定した IN
Cell Analyzer 6000 (GE Healthcare、リトル・チャルフォント、英国) を使用して SC-HiHs を観察した。 3.2.8 Cyclosporin A による胆汁酸濃度依存的な細胞毒性評価 添加する胆汁酸は、ヒト血清胆汁酸成分に従って12 種類選択した[17]。CsA は 一般的なトランスポーターを阻害する化合物として使用した。CsA 濃度は、BSEP の50%阻害濃度 (IC50) および阻害定数 (Ki) 値に基づいて決定した[32]。10 また は50 mM CsA および胆汁酸は、DMSO に溶解しストック溶液とした。この分析 で用いるDMSO の終濃度は 0.5% 未満に設定した。3 日目の SC-HiHs は、胆汁 酸の存在下または非存在下において、24 時間 10 または 50 µM CsA を添加した
後、LDH-Cytotoxic Detection Kit を使用して、損傷細胞から放出された乳酸デヒ
ドロゲナーゼ (LDH) の活性 (LDHsample) を測定することにより細胞毒性を評価
した。細胞におけるLDH 活性のポジティブコントロールでは、0.25% Triton
X-100 (LDHTritonX-100) を含む Cosmedium 004 で処理した。細胞毒性は、次の式を使
用して算出した。
Cytotoxicity (%) = (LDHsample – LDHblank) / (LDHTritonX-100 - LDHblank) × 100.・・・・(3)
33 3.2.9 薬剤性胆汁鬱滞肝毒性評価 SC-HiHs に、胆汁酸の存在下または非存在下で 24 時間、22 種類の試験化合物 を添加した[11, 17]。全ての化合物の濃度は、薬物誘発性の細胞毒性を避けるため に、10 µM CsA を除いて 50 µM に設定し、細胞毒性は以下の式を使用して算出 した。 Relative cytotoxicity (%)
= (LDHsample – LDHvehicle) / (LDHTritonX-100 – LDHvehicle) × 100. ・・・・・・・・・(4)
LDHvehicle値は、DMSO 処理した SC-HiHs の値を使用した。
3.2.10 統計学的解析
統計分析は、SPSS Statistics software package, version 13.0J (IBM Japan、東京) を 使用した。2 群間の比較は Student's t-test により行った。多重比較は、分散分析
を行った後、Dunnett test によって検定を行った。データは、3 ウェルの平均±標
準偏差として表し、p 値が 0.05 未満の場合に有意差ありと判断した。2~3 回繰
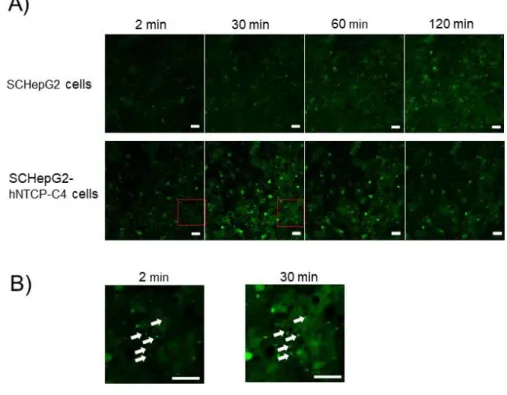
34 3.3 結果 3.3.1 経日的な形態変化と MRP2 および radixin タンパク質における蛍光免疫 染色 毛細胆管形成能を確認するために、ヒトiPS 細胞由来肝細胞の分化過程において、 サンドイッチ培養の有無による形態変化の検討を行った。 サンドイッチ培養1、3 および 5 日後 の SC-HiHs では、胆管様構造が形成されてい
る様子が観察された (Fig. 10A)。3 日目の SC-HiHs は、HiHs と比較して大きく平板化
していた。さらに、MRP2 および F-アクチンの発現は SC-HiHs で認められた (Fig. 10B)。 MRP2 は F-アクチンと共局在していたことから、毛細胆管構造が確認できた。さらに、5 日目のSC-HiHs では MRP2 および F-アクチンの共局在が認められなかった。そのた め、MRP2 が機能の備わったトランスポーターではないと考えられた。また、3 日目の SC-HiHs において、毛細胆管マーカーである radixin が染色されたため、毛細胆管が 形成できている可能性が示唆された (Fig. 10C)。
35
Figure 10. Morphology and polarity of transporters in sandwich-cultured human induced pluripotent stem cell (iPS cell)-derived hepatocytes (SC-HiHs).
(A) Morphological images of sandwich-cultured human iPS cell-derived hepatocytes (SC-HiHs) and conventional differentiation method (HiHs). Images are from day 1, 3, and 5 after sandwich culture. Black arrowhead, canalicular. Scale bar, 50 µm. (B) Immunofluorescence staining of multi-drug resistance-associated protein 2 (MRP2, green) in SC-HiHs on day 1, 3, and 5. Cell nuclei were stained with TO-PRO3 (blue). Cytoskeletal F-actin was stained with rhodamine-phalloidin (red). White arrow heads show canalicular. Scale bar, 100 µm. (C) Immunofluorescence staining of radixin (green) in SC-HiHs on day 3. Cell nuclei were stained with TO-PRO3 (blue). Scale bar, 20 µm.
36
3.3.2 胆汁酸トランスポーターの mRNA およびタンパク質発現
SC-HiHs における胆汁酸トランスポーターの発現を確認するために、mRNA およ
びタンパク質発現量を測定した。SC-HiHs における NTCP の mRNA 発現量は、
HepG2 細胞および HHs (48 h) よりも高値を示した (Fig.11A)。しかしながら、SC-HiHs
のBSEP の mRNA やタンパク質発現はほとんど認められず(Fig. 11A)、サンドイッチ培
養の有無にかかわらず、HepG2 細胞および HiHs と同様に検出されなかった。一方で、 SC-HiHs における MRP2 の mRNA 発現量は HiHs と同等であったが、HepG2 細胞お よび HHs (48 h) における発現量よりも低値を示した (Fig. 11A, 12A, B)。また、SC-HiHs の MRP2 タンパク発現量は HHs のタンパク発現量とほぼ同等であり、HepG2 細
胞およびHiHs のタンパク発現量よりも高値を示した (Fig. 12A, B)。これらの結果より、
サンドイッチ培養によって MRP2 のタンパク質発現量が増加することが示唆された。ま
た、5 日間培養した SC-HiHs では、培養日数依存的に MRP3 の mRNA 発現量は増
加することが示唆された。一方、MRP4 の mRNA 発現量は 3 日目に最も高値を示し
た。この値は、HepG2 細胞での値よりも高く、HepaRG 細胞での値と同等であった (Fig. 11B)。これらの結果から、F-アクチンと MRP2 は共局在し、機能が備わっていると考え られ (Fig. 10B)、また、MRP3 の mRNA 発現も高値を示すことから (Fig. 11B)、ヒト iPS
細胞由来肝細胞をサンドイッチ培養後 3 日目のものを機能評価かつ胆汁鬱滞肝毒性
37
Figure 11. mRNA expression levels of Na⁺-taurocholate cotransporting polypeptide (NTCP), bile salt export pump (BSEP), multidrug-resistance-associated protein 2 (MRP2), MRP3, and MRP4 in sandwich-cultured human induced pluripotent stem cell (iPS cell) -derived
hepatocytes (SC-HiHs).
(A) Samples were obtained from SC-HiHs, HepG2 cells, HiHs, human iPS cells, and cryopreserved human primary hepatocytes (HHs). Graph represents gene expression levels relative to that detected in HHs at 48 h. Data are expressed as means ± SD (n = 3, BSEP in SC-HiHs on day 1, 5; n = 2). (B) Samples were obtained from SC-HiHs, HepG2 and HepaRG cells. Graph represents gene expression levels relative to that detected in SCHiHs on day 1. Data are expressed as means ± SD (n = 3). **p < 0.01.
38 Figure 12. Protein expression of MRP2 and BSEP.
(A) Western blot analysis images. Samples were obtained from SC-HiHs, HepG2 cells, HiHs, and HPHs. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was the loading control. (B) Relative expression of MRP2 and BSEP. Data are expressed as means ± SD (n = 3, MRP2 and BSEP in SC-HiHs on day 1; n = 2). Graph shows protein expression relative to that of HPHs considered as 100.
**p < 0.01.
3.3.3 蛍光基質を用いた胆汁酸トランスポーターの機能活性および毛細胆管形 成能
CDFDA や tauro-nor-THCA-24-DBD を使用して、SC-HiHs における MRP2、 NTCP や BSEP トランスポーターの機能を評価した。CDF は時間依存的に毛細胆 管に蓄積した (Fig. 13A)。また、毛細胆管腔への tauro-nor-THCA-24-DBD の蓄積量 は、Ca2+およびMg2+を含まないHBSS 処理により減少した (Fig. 13B)。さらに、CsA の
39
Figure 13. Bile canaliculi imaging using fluorescent multidrug-resistance-associated protein 2 (MRP2) and bile salt export pump (BSEP) probe in sandwich-cultured human induced pluripotent stem cell (iPS cell) -derived hepatocytes (SC-HiHs).
(A) Excretion of 5- (and-6) -carboxy-2',7'-dichlorofluorescein (CDF) into the bile canaliculi via MRP2. Scale bar, 50 µm. (B) The fluorescence intensity of the substrate was determined using N- (24-[7- (4-N, N-dimethylaminosulfonyl-2,1,3-benzoxadiazole) ]amino-3α,7α,12α-trihydroxy-27-nor-5β-cholestan-26-oyl) -2'-aminoethanesulfonate (tauro-nor-THCA-24-DBD) in Hank’s balanced salt solution (HBSS) with or without Ca2+ and Mg2+, or 10 µM CsA. Scale bar, 50 µm.
3.3.4 胆汁酸濃度依存的な肝細胞毒性
SC-HiHs において、胆汁鬱滞肝毒性のヒト血清胆汁酸濃度を決定するために、 CsA を用いて細胞毒性評価を行った。SC-HiHs において、vehicle 群の細胞毒性は、 150 倍のヒト血清胆汁酸濃度で認められた。一方、細胞毒性は、CsA を処理することで 増強し、100 倍以上のヒト血清胆汁酸濃度において、プラトーに達した (Fig. 14)。
40
Figure 14. Concentration-dependent bile acid (BA) cytotoxicity in sandwich-cultured human induced pluripotent stem cell (iPS cell) -derived hepatocytes (SC-HiHs) .
SC-HiHs were exposed to CsA (10 and 50 µM, gray and black circles, respectively) or dimethyl sulfoxide (DMSO, vehicle, open circles) in the presence or absence of BAs for 24 h. BA concentrations were changed from 0- to 200-fold. Data are expressed as means ± SD (n = 3).
3.3.5 サンドイッチ培養したヒト iPS 細胞由来肝細胞と HepaRG 細胞および HepG2 細胞間における胆汁鬱滞性肝毒性の検出の相関分析 3.3.4 での結果に基づいて、100 倍のヒト血清胆汁酸濃度を使用して 22 化合物によ る胆汁鬱滞肝毒性評価を行った。その結果、アトルバスタチン、CsA、エベロリムス、フ ルタミド、ピオグリタゾン、シンバスタチン、およびタクロリムスによって胆汁酸存在下に て顕著な胆汁鬱滞肝毒性が引き起こされた (Fig. 15A)。一方、ラミブジンとチクロピジ ンは、胆汁酸を処理していない肝細胞毒性を有意に示した。次に、SC-HiHs と HepaRG または HepG2 細胞間における 22 化合物の胆汁鬱滞肝毒性の検出の相関 を検討した。HepaRG および HepG2 細胞の胆汁鬱滞肝毒性については、以前に報告 された値を使用した[11]。その結果、SC-HiHs の胆汁鬱滞肝毒性は HepG2 細胞より検 出できたが、HepaRG 細胞のそれよりも低い検出感度であった (Fig. 15B:y = 1.3461x-4.6876、r²= 0.6156; Fig. 15C:y = 1.2665x +5.1157、r²= 0.6722)。ピオグリタゾンの
41
毒性の値は HepG2 および HepaRG
細胞の値よりも高値を示した。これは、SC-HiHs に特異性があり、ヒト iPS 細胞から肝細胞への分化度が関与していると思 われた。
Figure 15. Drug-induced cholestatic cytotoxicity in sandwich-cultured human induced pluripotent stem cell (iPS cell) -derived hepatocytes (SC-HiHs)
(A) Relative cytotoxicity of 22 compounds with or without BA mixture. SC-HiHs were exposed to each compound for 24 h in the presence (closed bars) or absence (open bars) of 100-fold
concentration of bile acids (BAs). Data are expressed as means ± SD (n = 3). *p values < 0.05, **p values < 0.01. (B, C) Correlation of cytotoxicity of cholestatic drug-induced liver injury (DILI) compounds between SC-HiHs and HepaRG or HepG2 cells.
42
3.4 考察
トランスポーターの制御は、DILI の重要なメカニズムを知る手掛かりとなる。
ヒトでは、BSEP、MRP2 が胆管腔への胆汁酸排泄に関与していることが知られ
ている[33]。したがって、SC-HiHs、HepG2 細胞、および HiHs における BSEP お
よびMRP2 のタンパク発現量を調べた。SC-HiHs において、BSEP タンパク質は
発現しておらず、MRP2 タンパク質発現量は、以前に報告のある SC-HHs におけ
るタンパク質発現量と同様の傾向を示した[34]。また、SC-HiHs における細胞形態
は毛細胆管様構造が確認され、これは、サンドイッチ培養を行うことでのみ認められた (Fig. 10A)。したがって、サンドイッチ培養に用いる ECM は、SC-HiHs の毛細胆管形
成のために非常に重要であると考えられた。以前の研究で、ヒト iPS 細胞から肝細胞 への分化過程に用いるECM が、肝細胞および薬物代謝酵素マーカーの発現に影響 を与えることを報告した[35]。したがって、HiHs の発生学的特徴は、組織工学技術によ ってより改善される可能性が示唆された。また、SC-HiHs の MRP2 は F-アクチンと共局 在していた (Fig. 10B)。以前の報告において、HepaRG 細胞および SC-HHs における MRP2 は、F-アクチンと共局在することで、機能を有するトランスポーターであることが 知られている[19]。したがって、3 日目の SC-HiHs の MRP2 は、F-アクチン上にあり、 HepaRG 細胞および SC-HHs と類似した様子であり、MRP2 は機能を持ったトランスポ ーターであることが確認された。 毛細胆管への tauro-nor-THCA-24-DBD の蓄積量は、Ca2+および Mg2+を含まな いHBSS、または CsA の処理により減少した (Fig. 13B)。この結果により、胆汁 酸トランスポーターに輸送活性があることが示唆された。さらに、 tauro-nor-THCA-24-DBD は、胆汁酸の取り込みトランスポーターである NTCP の基質でも ある。また、CsA は NTCP の阻害剤であることが知られている[19]。したがって、 NTCP を介して肝細胞内へ取り込まれる tauro-nor-THCA-24-DBD は、CsA 処理に