矢 内 玲 子 ・長 澤
弘
国 立 が ん セ ン ター 研 究 所 薬 効 試 験 部
(東 京都 中央 区 築 地5-1-1)
(1977年3月28日
受 付)
は
じ
め
に
1.繁
殖
性
2.発
が
ん
性
3.乳
腺 腫 瘍 の ホ ル モ ン 依 存 性
4.乳
腺 腫 瘍 の 発 生,発
育 に お け るgenotype
とmammary
tumor
virusの
関 係
お
わ
り に
は
じ め
に
現 在 使 用 さ れ て い る 近 交 系 マ ウ ス の 大 部 分 は ア メ リ カ
で 作 られ,さ
か の ぼ れ ば 同 じgenetic
backgroundを
持 つ も の が ほ と ん ど で あ る[11,25]。1965年
頃,ヨ
ー
ロ ッパ 由 来 の 新 し い 近 交 系 と し てGR系
マ ウ ス が 作 ら
れ た[14]。
こ の 系 統 の 特 徴 は,(1)経
産 雌 でmammary
carcinomaが,早
期,高
頻 度 に 発 生 す る 。(2)こ
のcar-cinomaはmammary
tumor
virus(MTV)に
よ り
発 生 し,こ
のMTVは
乳 汁 経 由[13]以
外 に 雌 の 卵[33]
お よ び 雄 の 精 子[4,15]か
ら も,子
に 受 け つ が れ る 。
(3)経 産 中 に 発 生 し た 乳 腺 腫 瘍*2の
発 育 は ホ ル モ ン依 存 性
で あ り,妊
娠 中 に 発 育 し,分
娩 後 急 速 に 退 行 す る 。 一 方,
未 経 産 個 体 や 繁 殖 終 了 後 に 発 生 し た も の は ホ ル モ ン依 存
性 が な く,悪 性 度 も高 い[14]な どで あ るが,現 在,が ん
の研 究 に用 い られ て い る マ ウ スの 乳 が ん は 大 部 分 が そ の
発 育 に お い て ホ ル モ ン依 存 性 が な く[12],ホ ル モ ン依 存
型 の腫 瘍 は少 ない 。 しか も,こ のGRの
妊 娠 依 存 性 腫
瘍 は,次 第 に ホ ル モ ン依 存 性 を 失 い,悪 性 度 を 増 して ゆ
くの で,正
常 な 組 織 か ら悪 性 腫 瘍へ の “progression”
の過 程 を知 る のに 貴 重 で あ り,乳 が ん の 研 究 に 広 く利 用
され 得 る と考 え る 。(そ の他,妊 娠 依 存 性 乳 が ん を 高 発
す る系 統 とし ては,や
は り ヨー ロ ッパ 系 のRIII[24],
BR6[10]な
どが あ る。)
我 々の 研 究 室 で は1974年 に この 系 統 を 入手 し,種 々 の
特 性 に つ い て 検 討 中 で あ るが,本 稿 で は我 々 の得 た結 果
を 中 心 に,GR系
マ ウス の 概 略 に つ い て述 べ る 。
1.繁
殖
性
1974年1月,The
Netherlands
Cancer Institute,
Amsterdam,The
NetherlandsのDrs.L.M.Boot
andG.R
アpckeよ
り恵 与 され たGR/A(F67:♀7,
〓5)を
基 に,交 配 を 開 始 した 。飼 育 室 は温 度24±1℃,
湿 度60―70%,光
条 件 は5:00―19:00明,19:00―5:
00暗 で あ る。(15×30×12)cmの
テ フ ロ ンケ ー ジ中 で
飼 育 し,飼 料 はCA-1(日
本 ク レア),飲
料 水 は 動 物 用
オ ー レオ マ イ シ ンを0.02%添
加 した 水 を 与 え た 。交 配 方
法 は 兄妹 交 配 で,60―70日
令 で 交 配 を 開 始 し,3産
ま で
追 っか け 妊 娠 させ て交 配 を 終 了 した 。1腹 子 が7匹 以 上
の場 合 は6匹 まで 減 ら し,6匹
以 下 の 場 合 は と くに 子数
の調 整 は行 なわ な か った 。F67-F78ま
で の 繁 殖 成 績 を
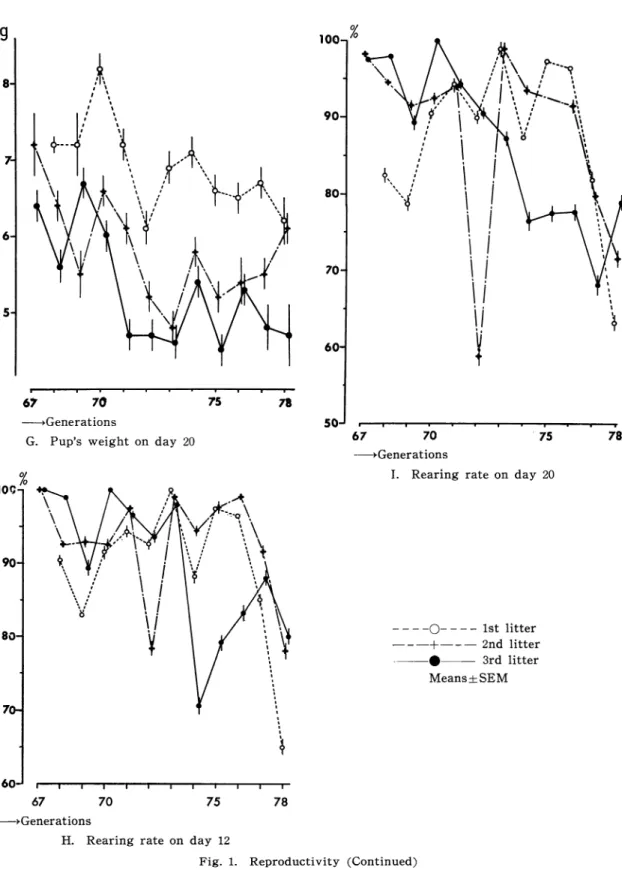
Fig.1に
示す 。交 配分 娩 間隔(Fig.1A)は
平 均25日
前 後,死
産 率(Fig.1B)は5%以
下 で,い ず れ も産 次
お よび 世代 に よる差 は 顕著 で はな か った 。 分 娩 時 乳 子 数
(Fig.1C),お
よび 分 娩 時 母 体 重(Fig.1D)は
各 世 代
と も産 次 がす す む に つれ て 明 らか に増 加 した 。 分 娩 時 子
体 重(Fig.1E)は
他 系 統 に比 べ て と くに違 いは な く,
Review : Studies on Reproduction
and
Pregnancy-Dependent
Mammary
Tumors
of GR/A
Mice.
Reiko YANAI
and Hiroshi
NAGASAWA,
Phar-macology
Division,
National
Cancer Center
Re-search Institute
*1 Standard
nomenclature
for inbred
strains
of
mice. Sixth listing : (Cancer
Res. 36, 4333, 1976)
で は,GRS/Aと
改 め られ て い る 。
*2詳
細 は 省 くが
,悪
性 度 の 低 い も の(Plaque,tumor
typepな
ど)を
「乳 腺 腫 瘍 」,悪 性 度 の 高 い も
の(ade-nocarcinoma)を
「
乳 が ん 」 と した 。
336
また 産 次 や 世 代 に よ る差 異 もな か った 。12日 令,20日
令
と も,子
体 重(Fig.1F,G)お
よび 増 体 率 は1産 か ら
3産 と経 るに つ れ て 次 第 に 低下 した 。 ま た20日 令 に おけ
る子 体 重 お よび 増 体 率 は 世 代 が進 む につ れ て低 下 す る傾
向 を 示 した 。 育 成 率(Fig・1H,1)は
産 次 お よび 世 代 に
よる 変化 が 大 き いが,ほ ぼ80%以
上 で あ った 。
Fig・2に
離 乳 後 の発 育 曲線 を示 した 。 雌 は お よそ80
日,雄 は70日 で ほ ぼ 一 定 の 体 重 に達 し,そ れ ぞれ 約209,
249,前
後 で あ った 。 雌 の腟 開 口 日令 は 生 後 約30日 で,
開 口時 体 重 は129前
後 で あ った 。
Fig.1Reproclucibility
338
Fig.2.Growth
curve
2―3カ
月令 の発 情 周 期 の 典 型 をFig.3に
示 した 。
他 の 多 くの 系統 の マ ウ ス と 同様,周 期 は 不 規 則 で,発 情
間 期 が 比 較 的長 い傾 向を 示 した 。 な お,こ れ らの マ ウ ス
は雌,雄 別 ケー ジで,原 則 と し て1ケ ー ジ6匹 で 飼 育 し
た 。
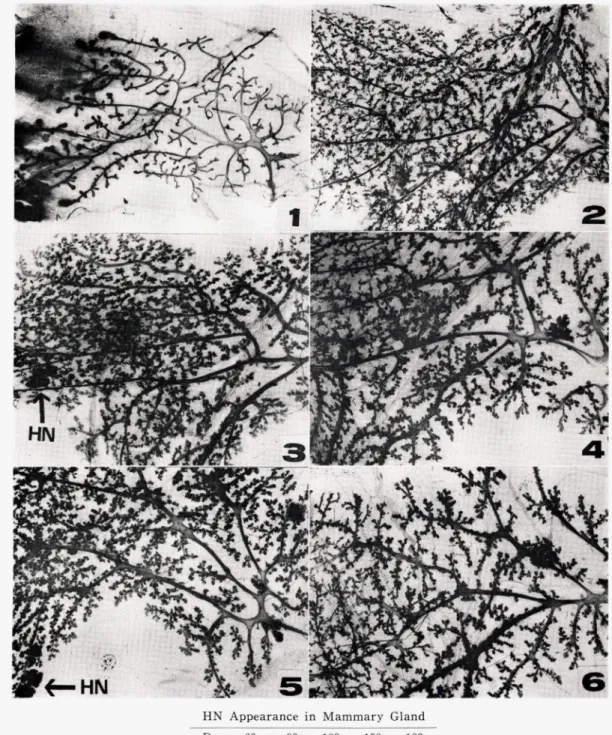
Fig.4は
生 後30日
令 か ら180日 令 の雌 の乳 腺 の ホー
ル マ ウ ン ト標 本 を 示 した もので あ る 。乳 腺 の発 育 は 非 常
に よ く,乳 がん の前 が ん症 状 で あ るhyperplastic
no-dules(HN)は90日
令 の雌 で7匹 中5匹 に 存 在 し,120
日令 以 降 で は全 例 に認 め られ た 。
2.発
が ん 性
先 に も述 べ た よ うにGR/Aマ
ウス で は 繁殖 中 に発 生
した 乳 腺 腫 瘍 は 妊 娠(な い しは ホ ル モ ン)依 存 性 を示 す
が,未 経 産 個 体 や 繁 殖 終 了 後 に発 生 した もの は悪 性 度 も
高 く,ホ ル モ ン依 存 性 を 示 さな い 。 これ ら乳 腺 腫 瘍 な い
し乳 が ん の 発 生 は 触 診 に よ り調 べ た 。 繁 殖 中 は,妊 娠 中
期 か ら分 娩 後,腫 瘍 が 消 失 す る ま で 隔 日に 調べ,繁 殖 終
了 後 は,週1回,死
ぬ ま で チ ェ ッ ク した 。 未経 産 マ ウス
で は,5カ
月 令 以 降,週1回,同
様 に して調 べ た 。発 生
した 腫 瘍 の 一 部 で 組 織 学 的 検 索 を 行 な った 。
繁 殖 中 の 乳 腺 腫 瘍 の 発 生 状 況 をFig.5に
示す 。発 が
Fig.3.Estrous
cycle
ん 率(Fig.5A)は,世
代 が進 む につ れ て徐 々に 高 くな
り,F78で
は 第1産
目58%,2産
目で はす でに100%
近 くに 達 した 。1匹 あ た りの腫 瘍 の数(Fig.5B)は
平
均 す る と1産 目で1個,2産
目で は2個 で あ った 。 腫 瘍
発 生 時 の 平 均 妊 娠 回 数(Fig.5c)は
ど の世 代 で も2以
下 で あ り,1回
目の 妊 娠 時 に発 生す る ものが 大部 分 で あ
った 。 平 均 発 生 月 令(Fig.5-D)は,ほ
ぼ3カ
月で あ
った 。2産 終 了 ま で に 発 生 した 腫 瘍 のす べ て は妊 娠 依存
性(分 娩 後 退 行 した)で あ った が,3産
終 了 後 は 約4%
の 腫 瘍 が 妊 娠 依 存 性 を 示 さな くな っ た(Table1)。
繁 殖 終 了 後 一 度 腫 瘍 が 退 行 して 消 失 した マ ウス をそ の
ま ま放 置 して お くと,新 ら し く,ま た は一 度 退 行 した 同
じ場 所 か ら再 び 腫 瘍 が 発 生 す る。 この 繁 殖 終 了後 の発 が
ん 率 と発 が ん 月令 をFig.6に
示 す 。 繁 殖 終 了後 の発 が
ん 率(Fig.6A)は
ほぼ90%以
上 で あ った 。 発 が ん 月 令
(Fig6B)は11カ
月前 後 で,世 代 に よ る変 化 は み られ
なか った 。 繁 殖 中 に 発 生 した 腫 瘍 の うち の 約30%が,繁
殖 終 了 し て一 旦 退 行 後,数 か 月で また 再 発 した 。 そ れ は
繁 殖 終 了 後 に 発 生 した 全 腫 瘍 の 約37%に
相 当
した(Tab-le2)。
未 経 産 雌 の発 が ん 率 をFig.7に
示 した 。 未 経 産 雌 で
は18か 月で ほぼ95%が
発 が ん した 。 平 均 発 が ん 月 令 は
13.8か 月で あ った 。GRお
よびGRと
他 系 統 とのFl
Fig. 4. Representative
wholemount
preparations
of mammary
glands
and appearance
of mammary
hyperplastic
nodules (HN).
340
Fig.
5.
Pregnancy-dependent
mammary
tumorigenesis
in breeding
females
Table
1.
Percentage
of
pregnancy-dependent
mammary
tumors
which
changed
into
autonomous
tumors
during
reproducti-ve periods.
* Total
no.
of
pregnancy-dependent
rpaj
nary
tumors
examined
に お け る発 が ん成 績 に 関 し ては,オ
ラ ン ダ[27,28]お
よび ア メ リカ[19,20]の
研 究室 で の結 果 が 報 告 され て
い る。
繁 殖 中,繁 殖 終 了後,お
よび未 経 産 の腫 瘍 を組 織 学 的
に分 類す る[18]とTable3の
通 りで あ り,妊 娠 中に
で き た腫 瘍 は大 部 分 がplaqueとtumor
typepで
あ
った の に 対 し,繁 殖 終 了 後,お
よび未 経 産 雌 に発 生 し た
もの は,ほ
とん どがcarcinomaで
あ った 。 未 経産 雌,
forced breeding雌,下
垂 体 移植 雌,お
よび卵 巣 を除
去 してestrone投
与 を した マ ウ スで 発 生 し た 腫 瘍 の
morphologyに
関 してVan
Nie and Duxが
報 告 し
てい る[28]。
3.乳
腺 腫 瘍 の ホ ル モ ン依 存 性
GRマ
ウ ス の腫 瘍 は 妊 娠 後 半 期 に増 殖 し,分 娩 と 同時
に 急 速 に 退 行 し,次 の 妊 娠 時 に 再発 す る こ とか ら ホル モ
ン依 存 性 で あ る と考 え られ た 。1965年,Van
Nie and
Thung[27]は
分 娩 後 消 失 した 腫 瘍 が,progesterone
(P)+estrone(E)の
投 与 に よ り,ふ た た び 増 殖す るが,
P単 独 投 与 で は,腫 瘍 は 再 発 しな い こ とを報 告 した 。 ま
たVan
Nie and Dux[28]は,こ
の 妊 娠 依 存性 の腫
瘍 を 移 植 す る際,P+E+下
垂 体移 植 したhostで
は増
殖 す るが,正 常 雌,去 勢 雄,あ る いは 下 垂 体 移 植,P+
Eの み で は,増 殖 しな い こ とを認 め た 。Janikら[9]
Fig. 6. Mammary tumorigenesis in retired fem-ales Means•}SEM
胞 産 生 の 速 度 の 増 加 に よ る と 報 告 し た 。 我 々 は こ のGR
の 妊 娠 依 存 性 腫 瘍 へ の3H-thymidineの
と り こ み か ら,
DNA合
成 能 を 検 討 し た[31]。
そ の 結 果 をFig.8に
示 す 。 分 娩 前 日 に は 正 常 乳 腺 も 腫 瘍 も3H-thymidine
の と り こ み は 高 か っ た が,分
娩 の 翌 日 に は 両 者 と も と り
こ み は 急に
低 下 し た(Fig.8-A)。
減 少 の 程 度 は 正 常
乳 腺 よ り も腫 瘍 の 方 が 著 し か っ た 。 分 娩 前 日 か ら 種 々 の
ホ ル モ ン処 理 を し た 場 合 の,分
娩 翌 日 に お け る と り こ み
をFig.8-Bに
示 す 。 腫 瘍 へ の と り こ み はestradiol
benzoate(EB),ヒ
ト胎 盤 性 ラ ク トゲ ン(HPL),下
垂
体 移 植(3AP)な
ど で は 全 く 対 照 群 と違 い が な か っ た が,
P,P+EBで
は,と
り こ み は 高 く,P+HPLで
は,P
単 独 よ り さ らに 上 昇 し,こ
の 腫 瘍 の 増 殖 は,Pに
大 き く
依 存 し て い る こ と が 推 測 さ れ た 。 一 方,autonomousな
Fig. 7. Mammary tumor incidence in virgins *Mammary tumor age (means•}SEM) and no. of estimates
Table 2. The incidence
of autonomous
mammary
tumors
which once disappeared
at the end of reproductive
periods
in GR/A female mice
Table 3. Types of mammary
tumors
which appeared
during
pregnancy,
after
342
Abbreviations
: C=control ; EB=estradiol benzoate ; P=progesterone ; HPL-human placental lactogen ; 3 AP=isograftsof 3 pituitaries
aMeans•}SEM
** P <0 .01 vs Control * P <0.05 vs Control •õ P <0.01 vs P-group
Fig. 8. DNA synthesis
of normal mammary
glands
and mammary
tumors
[31]
SHNの
腫 瘍 で は ホル モ ン処 理 に 関 係 な く,常 に と り こ
み は 高 か った(Fig.8-c)。
妊 娠15日 に卵 巣 を 除 去 し,
Pあ るい はP+EBを
毎 日投 与 してDNA合
成 に 対す る
ホル モ ンの 影 響 を 妊 娠19日 に 見 た場 合[32]に
は,P単
独 で は 正 常 雌 に 比 べ て3H-thymidineの
と り こみ は低
く,P+EB投
与 で は 正 常 の レベ ル に 達 した(Fig.9)。
しか し 乳 腺 で はPの
量 を増 加 して もと りこみ は 変 らず
に 低 い ま ま に と ど ま った のに,腫
瘍 で はPの
量 を 増 す
こ と に よっ て と りこみ も増 した 。 これ らの 結 果 か ら,正
常 乳 腺 よ りも腫 瘍 の方 がPに 対 す る感 受 性 の 高 い こ とが
Fig. 9. The in vivo incorporation of 3H-thymi-dine into DNA of normal and neoplastic mammary glands on day 19 of pregna-ncy in each group. [32]
Ovex=ovariectomy ; P=progesterone ; EB=estradiol benzoate.
aMeans•}SEM
bNumber of estimates
Mammary glands : I/II, III : II, III/IV, V : IV/V : P <O. O1.
Mammary tumors : I/II, III ; II/III, iv, v; III/IV,V : P<O.01.
Fig. 10. Specific prolactin binding activities of normal mammary glands and mammary tumors and plasma prolactin levels on day 19 of pregnancy and 1 day after parturition (means•}SEM).
*Number of estimates
a/c, b/c, d/e, d/f, g/h, g/i: P <0.01.
Table 4. Transplantation
of pregnancy-dependent
mammary
tumors
of GR/A mice into female nude mice
*Total
no. of mice
receiving
tumors
344
Fig. 11. Growth curve of mammary tumors in each group. 2P and 3P are the 2nd and the 3rd parturitions, respectively. [18]
*Mammary tumor size was expressed in terms of the major two diameters.
•¬ of parturition
示 唆 さ れ た 。 ま たSluyser
and
Van
NieE22]は,
卵 巣 除 去 し たGRマ
ゥ ス にP+Eを
投 与 す る と腫 瘍 を
発 生 し,そ
れ は 妊 娠 中 に 発 生 し た 腫 瘍 と 同 様,そ
の90%
以 上 が ホ ル モ ン 依 存 性 で あ る こ と を 見 出 し た 。 こ の 腫 瘍
を 移 植 す る と,最
初 は ホ ル モ ン 依 存 性 の 腫 瘍 が 増 殖 す る
が,移
植 を く り返 す う ち に,つ
い に は ホ ル モ ン依 存 性 が
な くな る こ と が わ か っ た 。 そ し て,こ
の ホ ル モ ン依 存 性
と 非 依 存 性 腫 瘍 のE-receptorを
測 定 し た と こ ろ,ホ
ル
モ ン 依 存 性 腫 瘍 で は 高 く,非
依 存 性 腫 瘍 で は 低 い 結 果 を
得 た 。TereniusもE-receptorに
つ い て 同 様 の 結 果
を 報 告 し て い る[26]。
最 近,Sluyserら[23]は,ホ
ル モ ン 依 存 性 腫 瘍 で はE-recePtorお
よ びP-receptor
と も 高 く,非
依 存 性 腫 瘍 で は 前 者 は 低 く,後
者 は ほ と ん
ど 欠 如 し て い る こ と を 明 ら か に し た 。
Fi9.10は
分 娩 前 後 のProlactin
recePtorを
測 定 し
た 結 果 で あ る(Yanai,R.eta1.未
発 表)。 妊 娠19日
に
Fig. 12. Percentage of various types of mam-mary tumors in each group [18].•¬ Adenocarcinoma type A ;•¬
Adeno-carcinoma type B ;•¬ Mixed type of A and B ; •¬ Adenocarcinoma type
C ;•¬ Adenoacantoma ;•¬ Pale cell carcinoma ; •¬Mammary tumor type p and plaques. Number at the right of each column represents the number of tumors examined.
は 正 常 乳 腺 も腫 瘍 もprolactin
binding
activityが
低 か っ た 。 一 方,分
娩 翌 日 の 結 果 は,正
常 乳 腺 で は 乳 子
を 哺 育 さ せ た 場 合 と,分
娩 後 泌 乳 を さ せ な い 場 合 で は そ
のactivityが
非 常 に 異 な る の に 対 し て,腫
瘍 で は 泌 乳
の 有 無 に よ る ホ ル モ ン 条 件 で 全 く 影 響 を 受 け な い こ と が
わ か っ た 。Aidells
and
Daniel[1-3]は,GRの
ホ
ル モ ン依 存 性 乳 腺 腫 瘍 を 同 系 統 マ ウ ス のfatpadに
移
植 し,PreneoPlasticあ
る い はneoPlasticな
性 質 を 詳
し く調 べ て い る 。 一 方,Briandら[7,8,22]は
こ の
腫 瘍 の 酵 素 系 を 測 定 し て,正
常 乳 腺,あ
る い
はautono-mousな
腫 瘍 と の 比 較 を 行 な っ て い る 。
GRの
妊 娠 依 存 性 腫 瘍(組
織 学 的 に は 主 にplaqueあ
る い はmammary
tumor
tyPePよ
り な る)を,雌
のnudeマ
ウ ス に 移 植 し た 結 果 をTable4に
示 し た 。
移 植 後20週
ま で に 腫 瘍 を 発 生 し た 個 体 は,P-pellet移
植 群 で は10匹
中3匹,対
照 群 で は9匹
中4匹
で,全
く,
る よ うに,GRのhormone
responsiveな
腫 瘍 は1
つ の 腫 瘍 の 中 にhormone
dependentな
良 性 の 細 胞 か
ら,autonomousな
悪 性 の 細 胞 ま で い ろ い ろ なtyPe
の 細 胞 を 含 む も の で あ り,そ
の 中 の,よ
り活 性 の 高 い,
言 い か え れ ば 悪 性 度 の 高 い 細 胞 が,増
殖 し て く る も の と
考 え ら れ る 。
4.乳
腺 腫 瘍 の 発 生,発
育 に お け るgenotypeと
mammary
tumor
virusと
の 関 係 に つ い て
は じ め に 述 べ た よ う にGRの
腫 瘍 はMTVに
よ る も
の で あ り,そ
のMTVは
雌 の 乳 汁 以 外 に 卵 お よ び 雄 の
精 子 か ら もそ の 子 に 受 け つ が れ る 。 し か し,こ
のMTV
を 他 の 系 統 の マ ウ ス に 導 入 し た 時,そ
の 新 し いhostか
ら は 次 の 雄 に は 導 入 さ れ な い の で,乳
汁 経 由 以 外 の 方 法
で こ れ を 伝 え る た め に は,GRのgenomeが
必 要 で あ
る と 考 え ら れ た[4,33]。Bentvelzen[5]は,こ
の
MTVがGRのgenomeに
伴 っ て 一 つ のgenetlc
factorと
し て そ の 子 に 受 け つ が れ る と 考 え た 。 ま
たNa-quesを
発 生 す る か,そ
れ と も ホ ル モ ン 依 存 性 の な い 腫
瘍 を 作 る か はMTVを
受 け るhostの
日令に
よ る と報
告 し て い る 。GRのMTVに
つ い て の 詳 細
は,Bent-velzenら
に よ る 広 範 な 実 験 お よ び 綜 説 が あ る の で,そ
れ ら を 参 照 さ れ た い[4,5,6,15,29]。
こ こ で は,
我,々 の 研 究 室 で 作 ら れ たSHN系
マ ウ ス[16]と
の 関 係
に つ い て 述 べ る 。
SHN系
はSwiss
albinoよ
り選 抜,育
種 さ れ た 乳 が
ん 高 発 系 マ ウ ス で あ り,Bittner
typeのMTVを
も
つ 。 そ の 乳 が ん の 発 育 は 完 全 にautonomousで
あ る
[17,31]。
こ のSHNとGRと
のF1,お
よ び 両 系 統
を そ れ ぞ れ 相 互 にfoster
nurseさ
せ た 場 合 の 発 が ん 成
績 をTable5に
示 す 。 繁 殖 中 の 乳 腺 腫 瘍 の 発 生 率,お
よ び 発 生 し た 腫 瘍 の 妊 娠 依 存 性 と も,両
系 統 のF1は
完
全 にGRと
同 じ結 果 を 示 し た 。 こ れ はVan
Nie
and
Thung[27],Nandi
and
Helmich[20]ら
のC57BL
やBALB/c(こ
れ ら の 系 統 は い か な る タ イ プ のMTV
も 持 っ て い な い)と
のF1を
用 い た 実 験 結 果 と 一 致 す
Table 5. Genotype,
type of MTV and mammary
tumorigenesis
in each group. [18]
a) Each group of mice was subjected to force-breeding beginning about 60 days of age until the 3rd to the 5th pregnancy. Females of Groups I-IV were mated with males of the same groups, respectively, and females in Groups V and VI were mated with SHN males.
b) Groups II and VI were significantly below the other groups in all parameters (P <0.05 or 0.01), while there was little difference between Groups II and VI and between Groups I, III,
IV and V, respectively. c) From males through sperms.
d) Number of mice with mammary tumors/total number of mice examined. e) Number of pregnancy-dependent tumors/total number of tumors. f) Mean•}S.E.M.
346
る 。 ま た,GRの
子 をSHNにfoster
nurseさ
せ た
場 合 はGRに
哺 育 さ せ た 場 合 と ほ ぼ 同 じ 結 果 で あ っ た が,
SHNの
子 をGRにfoster
nurseさ
せ た 場 合 は
,M
TVのtypeはGRで
あ る と考 え られ る の に,SHNと
同 じ よ うな 発 が ん 性 を 示 し,発
生 し た 腫 瘍 のtypeは,
妊 娠 依 存 性 の も の は 少 な か っ た 。 こ の こ と か ら,GRの
MTVは,GRのgenomeを
伴 わ な い と 伝 え ら れ に
く い こ と が わ か る 。 各 群 の 典 型 的 な 腫 瘍 の 発 育 状 態 を
Fig.11に
示 し た 。 ま た 各 群 の 腫 瘍 の 組 織 学 的 分 類[18]
をFig.12に
示 す 。GR
typeの
発 が ん 成 績 を 示 す 両
系 統 のF1と,GR/fSHNはGRと
同 じtypeの
腫 瘍
で あ り,SHN
typeの
発 が ん 成 績 を 示 すSHN/fGR
はSHNと
類 似 の 組 織 学 的typeを
示 した 。
最 近,Van
Nie and
Hilgers[30]は,17α-ethynyl
-19-nortestosteroneを
,卵
巣 除 去 し たGRマ
ウ ス に
投 与 す る と3週
間 で 全 て の マ ゥ ス にearly
stages
of
mammary
tumorを
発 生 す る が,GRにfoster
nurse
し たBALB/cやMAS系
マ ウ ス で は 腫 瘍 は 発 生 し な
い こ と を 見 出 し,こ
の 腫 瘍 の 発 生 にgeneが
必 要 条 件 で
あ る こ と を 報 告 し て い る 。
お わ
り に
ヒ トの 乳 が ん の モ デ ル と しては,主 に 発 が ん 剤 に よ る
ラ ッ トの 乳 が んや,マ
ウス の 自然 発 生 乳 が ん が 用 い られ
て い る 。 と くに マ ウス の 自然 発 生 乳 が ん は 系 統 も多 く,
それ ぞ れ 特 徴 が あ る。 モ デル と して どれ が 最 も よい か は,
各 々一 長 一 短 が あ り,一 概 に は 言 え な い が,各 人 の実 験
の 目的 に 応 じた 系 統 を 選 ぶ こ とが 大切 で あ る 。そ の ため
に は 各 系統 の特 徴 を よ く知 る こ とが 必要 で あ る と考 え,
こ こで はGRマ
ウス に 関 す るい ろ い ろ な成 績 を 挙 げ た 。
GR系
マ ウス は乳 が ん 発 生 に おけ る,virusお
よび ホ
ル モ ンの 両 方 の立 場 か ら,非 常 に 有 用 な マ ウ ス と考 え る。
今後,乳
が ん の研 究 に 寄 与 す る と ころ 大 で あ ろ う。
乳 腺腫瘍 の組織 学的検索は,関 西医科大学病理学教室森井外
吉教授,お よび螺良愛郎助手に よった。記 して謝意を表 する。
またGRマ
ウスの飼育,管 理な どに御 協力頂 いた谷 口寿,宮
本盛吉両技官 に感謝す る。われわれの研究室 で行なわれ た研究
の一部 は,文 部省科学研究費補助金(が ん特別研究1)の
助成
による。
文
献
[1]
Aidells, B. D. and Daniel,
C. W. (1974)
. Hormone-dependent mammary tumors in strain GR/A mice. I. Alteration between
ductal and tumorous phases of growth du-ring serial transplantation. J. Natl. Cancer
Inst., 52, 1855-1863.
[2] Aidells, B. D. and Daniel, C. W. (1976) . Hormone-dependent mammary tumors in
strain GR/A mice. II. Preneoplastic and neoplastic properties. J. Natl. Cancer Inst.,
57, 916-526.
[3] Aidells, B. D. and Daniel, C. W. (1976). Hormone-dependent mammary tumors in strain GR/A mice. III . Effectiveness of
supplementary hormone treatments in
in-ducing tumorous phase growth. J. Natl. Cancer Inst., 57, 527-537.
[4] Bentvelzen, P. (1968). Genetical control of the vertical virus transmission of the Mð¹-hlbock mammary tumor virus in the GR
mouse strain. Ph. D. Thesis, Univ. Leiden, Hollandia Publ. Co., Amsterdam.
[5] Bentvelzen, P.(1972). Hereditary infections with mammary tumor viruses in mice. In RNA Viruses and Host Genome in
Oncoge-nesis. Emmelot P. and Bentvelzen, P., edi-tors, North Holland Publ. Co., Amsterdam,
pp.309-337.
[6] Bentvelzen, P. and Daams, J. H. (1969) . Hereditary infections with mammary
tu-mor viruses in mice. J. Natl. Cancer Inst., 43, 1025-1035.
[7] Briand, P. and Daehnfeldt, J. L. (1973). Enzyme patterns of glucose catabolism in hormone-dependent and -independent
ma-mmary tumours of GR mice. Europ. J. Ca-ncer, 9, 763-770.
[8] Daehnf eldt, J. L., Schð¹lein, M. and Briand, P. (1975). Biochemical changes in GR
mo-use mammary tissue during hormonal
tu-mour induction. Europ. J. Cancer, 11, 509 -515.
[9] Janik, P., Briand, P. and Hartmann, N. R. (1975). The effect of estrone-progesterone
lopment in BR6 mice; ovarian influences
and 5-hydroxytryptamine. Brit. J. Cancer, 24, 561-567.
[11] Little, C. C. (1941). Biology of the Labora-tory Mouse. Snell, G. D., editor, Blakiston,
Philadelphia.
[12] Mð¹hlbock, O. (1955). The influence of ho-rmones on spontaneous experimental
tu-mours. In: Simposio sugli antimitotici San Remo, p.41-53.
13] Mð¹hlbock, O. (1956). Biological studies on the mammary tumor agent in different st-rains of mice. Acta Unio Int. Contra. Ca-ncr. XII, 665-681.
[14] Mð¹hlbock, O. (1965). Note on a new inbred mouse strain GR/A. Europ. J. Cancer, 1, 123-124.
[15] Mð¹hlbock, O. and Bentvelzen, P. (1968). The transmission of mammary tumor
viru-ses. Perspect. V irol., 6, 75-87.
[16] Nagasawa, H., Yanai, R., Taniguchi, H., Tokuzen, R. and Nakahara, W. (1976). Two -way selection of a stock of Swiss albino
mice for mammary tumorigenesis:
Esta-blishment of two new strains (SHN and SLN). J. Natl. Cancer Inst., 57, 425-430.
[17] Nagasawa, H., Tokuzen, R. and Nakahara, W. (1976). Growth of mammary tumors in a high and a low mammary tumor strains
of mice established from the same basal
stock of Swiss albino. GANN, 67, 913-915.
[18] Nagasawa, H., Morii, S., Tsubura, A. and Yanai, R. (1977), Relative importance of
genotype and type of mammary tumor vi-rus on mammary tumorigenesis in mice. Europ. J. Cancer., in press.
[19] Nandi, S. and Helmich, C. (1974). Trans-mission of mammary tumor virus by the
GR mouse strain. I. Role of the virus in the production of lesions. J. Natl. Cancer
Inst., 52, 1285-1290.
J. Natl. Cancer Inst., 52, 1567-1570.
[21] Schð¹lein, M., Daehnf eldt, J. L. and Briand, P. (1974). Lactose synthetase induction by
hormones in normal and tumorous GR mou-se mammary tissue. Int. J. Cancer, 14,
372-378.
[22] Sluyser, M. and Van Nie, R. (1974). Est-rogen receptor content and
hormone-res-ponsive growth of mouse mammary tumors. Cancer Res., 34, 3253-3257.
[23] Sluyser, M., Evers, S. G. and De Goeij, C. C. J. (1976). Sex hormone receptors in
ma-mmary tumours of GR mice. Nature, 263,
386-389.
[24] Squartini, F. (1962). Responsiveness and
progression of mammary tumors in high-cancer strain mice. J. Natl. Cancer Inst., 28, 911-926.
[25] Strong, L. C. (1942). The origin of some inbred mice. Cancer Res., 2, 531-539.
[26] Terenius, L. (1972). Parallelism between oestrogen binding capacity and hormone
responsiveness of mammary tumours in GR
/A mice. Europ. J. Cancer, 8, 55-58.
[27] Van Nie, R. and Thung, P. J. (1965). Res-ponsiveness of mouse mammary tumours
to pregnancy. Europ. J. Cancer, 1, 41-50.
[28] Van Nie, R, and Dux, A. (1971). Biological and morphological characteristics of
ma-mmary tumors in GR mice. J. Natl. Cancer
Inst., 46, 885-897.
[29] Van Nie, R., Hilgers, J. and Lenselink, M.
(1972). Genetical analysis of mammary tumor development and mammary tumor virus expression in the GR mouse strain.
In Colloque Inserum Recherches Fondame-ntales sur les Tumeurs Mammaires.
Mou-riquand, J., editor, Lyon, Imprimerie Bose Frð¤res, pp.21-29.
[30] Van Nie, R. and Hilgers, J. (1976). Genetic analysis of mammary tumor induction and

![Fig. 6. Mammary tumorigenesis in retired fem- fem-ales Means•}SEM 胞 産 生 の 速 度 の 増 加 に よ る と 報 告 し た 。 我 々 は こ のGRの 妊 娠 依 存 性 腫 瘍 へ の3H‑thymidineの と り こ み か ら,DNA合成 能 を 検 討 し た[31]。そ の 結 果 をFig.8に示 す 。 分 娩 前 日 に は 正 常 乳 腺 も 腫 瘍 も3H‑thymidineの と り こ み](https://thumb-ap.123doks.com/thumbv2/123deta/6854584.1171640/7.756.64.698.105.835/FigMammary胞産生速度増加よる報告し々こ.webp)
![Fig. 8. DNA synthesis of normal mammary glands and mammary tumors [31]](https://thumb-ap.123doks.com/thumbv2/123deta/6854584.1171640/8.756.397.687.105.430/fig-dna-synthesis-normal-mammary-glands-mammary-tumors.webp)


![Table 5. Genotype, type of MTV and mammary tumorigenesis in each group. [18]](https://thumb-ap.123doks.com/thumbv2/123deta/6854584.1171640/11.756.70.700.588.785/table-genotype-type-mtv-mammary-tumorigenesis-group.webp)
