題 目
題 目
肝固有および骨髄由来 Kupffer 細胞の 異なる発生・分化と機能についての研究
五十嵐
い か ら し正
まさ巳
み(微生物・免疫学専攻)
防 衛 医 科 大 学 校
平成26年度
目 次
第1章 緒言 1頁
第2章 実験と方法 3頁
第3章 結果 9頁
第4章 考察 17頁
第5章 結論 22頁
謝辞 23頁 引用文献 24頁
略語一覧 30頁 図 32頁
1
第1章 緒 言
Kupffer細胞は肝のマクロファージ(M)であり、全身のMの約半数を占め
る。Kupffer細胞の主要な役割は、微生物、免疫複合体、エンドトキシン、門脈
由来抗原、腫瘍細胞の除去である(1-4)。血中に入り込んだ細菌の7-8割は肝に集 められ、Kupffer細胞や貪食型の肝B細胞によって除去される(2, 5-7)。そのため、
肝内のKupffer細胞による細菌除去は菌血症に対する生体防御にとって非常に重
要である(8)。Kupffer細胞はIL-1、IL-12、IL-18、TNF等の炎症性サイトカイン を産生し、病原体の貪食後には活性酸素(ROS)も産生し、さらにT細胞や NK/NKT細胞を含む他のリンパ球への抗原提示も行う(1, 2)。Kupffer細胞が産生 するIL-6やTNFのようなサイトカインは、肝細胞に急性期反応物質や補体を産 生させ、免疫グロブリンと共に菌をオプソニン化し、Kupffer細胞が病原体を貪 食しやすくする点でも非常に重要である(1, 5)。
近年、KinoshitaらはマウスF4/80+ Kupffer細胞がサイトカイン産生型CD11b+ 細胞と貪食/ROS産生型CD68+細胞の2種類に分けられることを報告している(2, 9)。この報告では、正常マウスのKupffer細胞は主としてCD68+細胞から成り、
他組織や全身循環中のM/単球であるCD11b+細胞とは異なる(2, 9)。またCD68+
細胞は、CD11b+細胞と異なり肝組織をcollagenase処理しないと分離できないこ
とから、肝類洞内皮や肝細胞に固着していると考えられている(2, 9)。Kleinらは
マウスKupffer細胞に骨髄由来のMと肝固着性のMと2グループがあると述
べている(10)が、Kinoshitaらの報告と比較すると前者がCD11b+細胞、後者が CD68+細胞であると思われる(9, 10)。HoltらもCD11blow F4/80high細胞とCD11bhigh
F4/80low細胞の2種類がマウス肝に存在すると報告しており、前者は放射線耐性
でClodronate(dichloromethylene diphosphonate)によって除去されるが、後者は 逆に放射線感受性でありClodronateでは除去されないと述べている(11)。したが
2
って、Kupffer細胞には機能的に異なるサブセットが存在するものと考えられる。
これらの報告ではフローサイトメトリー解析での測定条件設定が異なる可能性 があるが、CD11blow F4/80high細胞がCD68+細胞に、CD11bhigh F4/80low細胞が
CD11b+細胞にそれぞれ相当すると解釈できる(9)。
しかしながら、マウス肝F4/80+ Kupffer細胞にはCD11b陰性かつCD68陰性
(CD11b- CD68-)の細胞群が30%程度存在しており、これらの細胞群の位置づ けは必ずしも明確ではない。そこで本研究では、IgGがMに直接親和性がある ことにより同定された抗-FcR(CD16/CD32)抗体(12, 13)を新規に用いることで、
Kupffer細胞の包括的な分類を試みた。また、Kupffer細胞に特異的に発現し、細
菌の貪食に関与する補体受容体であるcomplement receptor of the Ig superfamily
(CRIg)(14)の発現についても調べた。
さらに、マウスだけでなくヒトの肝Kupffer細胞についてもCD11b、CD32
(FcRII)、CD68の発現による分類を行った。F4/80はマウスのKupffer細胞/M
に特異的な表面マーカーであり、ヒトには使えないため、本研究ではヒトMの 貪食やLPSシグナルの伝達に関与しているCD14 (15, 16)を用いて、CD14陽性の 肝単核球をヒトのKupffer細胞と定義した。
以上に基づき、本研究では、マウスおよびヒトKupffer細胞における、表面抗 原的な特徴、生体内における機能、および発生・分化についての包括的な分類 を新規に確立することを目的とした。本研究の成果は、Kupffer細胞が肝固有の 細胞群と骨髄由来の細胞群から成り、抗菌および抗腫瘍/炎症という生体防御に 必須な免疫学的役割を明確に分担していることを示すものである。また、致死 量放射線照射後も肝固有のCD32/CD68+ Resident Kupffer細胞が消失せず継続し て存在し、その機能を保持することから、Resident Kupffer細胞はその供給を骨 髄に依存せずYolk Sac(卵黄嚢)由来である(17, 18)ことを示唆するものである。
3
第2章 実験材料と方法
1. マウスおよびヒトからの肝単核球の分離
本研究の実施において、マウスを用いた実験に関しては防衛医科大学校動物 実験倫理委員会(承認番号12039)から、ヒトの肝組織を用いた実験に関しては 防衛医科大学校倫理委員会(承認番号807)からそれぞれ承認を得た。
7週齢の雄性C57BL/6マウス(B6マウス)を日本クレアから購入し使用した。
マウス摘出肝をミンチ状に細切した後、0.05% collagenase(和光純薬、大阪)を 含む Hanks’ Balanced Salt Solution(HBSS)15 mLを加え37℃で20分間振盪温 浴した。その検体をステンレスメッシュ(メッシュ径0.1mm)で濾過した後、
33% Percollによる比重遠心法で肝単核球分画を肝細胞から分離し採取した。赤
血球は浸透圧溶血によって除去した(9, 10, 19-21)。
ヒト肝組織については、外科肝切除9症例(年齢64-72歳、男性6例、女性3 例)からインフォームド・コンセントを得た上で肝組織検体を得た。9例中3例 は閉塞性黄疸があり、肝単核球分離が困難であったため除外した。残りの6例 は、肝内結石症1例、原発性肝癌3例、転移性肝癌2例であり、全て本研究の 解析対象に含めた。いずれの症例でも病変部から離れた背景肝組織から組織を 採取し検討に用いた。
2. フローサイトメトリー解析
CD16/CD32の染色を除き、抗体の非特異的結合を防ぐためにマウス肝単核球
をFc-blocker(2.4 G2; BD PharMingen, San Diego, CA, USA)で前処理した。その 単核球をPECy5標識抗-F4/80抗体(BM8; eBioscience, San Diego, CA, USA)、PE 標識抗-CD11b抗体(M1/70; eBioscience)、PE標識抗-CD16/CD32抗体(93;
eBioscience)、Biotin標識抗-CD68抗体(FA-11; AbD Serotec, Oxford, UK)とPE
4
標識Streptavidinで染色した。Kupffer細胞の特徴を調べるために、肝単核球を FITC標識抗-Gr-1抗体(RB6-8C5; Beckman Coulter, Miami, FL, USA)、FITC標 識抗-NK1.1抗体(PK136; eBioscience)、FITC標識抗-c-kit(CD117)抗体(2B8;
eBioscience)、FITC標識抗-CD45.1(Ly5.1)抗体(A20; eBioscience)で染めた。
ヒト単核球も同様にPECy7標識抗-CD14抗体(RMO52; Beckman Coulter)、PECy5 標識抗-CD11b抗体(Bear 1; Beckman Coulter)、PE標識抗-CD68抗体(Y1/82A;
eBioscience)、PE標識抗-CD32抗体(2E1; Immunotech, Marseille, France)で染色 した。
フローサイトメトリー測定および解析はCytomics FC500(Beckman Coulter) を使用した。
3. 細胞分取
肝単核球からCD11b陽性細胞を分取するため、PE標識抗-CD11b抗体および 磁気beads結合抗-PE抗体を用いて細胞を標識し、magnetic-activated cell sorting
(MACS; Miltenyi Biotec, Bergisch Gladbach, Germany)によって磁気的に採取し た。CD32陽性細胞とCD68陽性細胞についてもそれぞれ同様に分取した。採取 したこれらの細胞はPCR解析に用いた。
4. RNA抽出、逆転写、定量PCR
RNeasy Mini Kit(QIAGEN, Valencia, CA, USA)を用いて、脾単核球、肝単核 球、分取した肝単核球からtotal RNAを抽出した。SuperScript III First-Strand Synthesis Kit(Invitrogen Life Technologies, Carlsbad, CA, USA)を用いてRNA
500ngを逆転写し、cDNAを得た。マウス補体受容体CRIgとF4/80検出プライ
マーは以下の通り。
5
CRIg(Vsig4): sense 5’-GACTTGACCACTAATGGGACTGGAA-3’
antisense 5’-GTCCTGCAGCGGAACAAGATATAA-3’
F4/80(Emr1): sense 5’-CACCGGTATAGACAAGACTGACA-3’
antisense 5’-TCTCACCATCAGGAAGAGCA-3’
定量PCRはSYBR Green I Master Mixを用いて行いLightCycler480(Roche Diagnostics, Vilvoorde, Belgium)で測定した。測定結果は、同一検体のF4/80 mRNA 発現量との相対比で示した。
5. 免疫組織化学
肝をクリオスタットで6 μm厚に薄切した新鮮凍結切片をスライドガラスに載 せ伸展させ、アセトン固定後に洗浄した。1% Block Ace(DS Pharma Biomedical、
東京)で10分間浸漬した後に、100倍希釈の抗-CD68抗体(FA-11; AbD Serotec)、 抗-CD11b抗体(M1/70; eBioscience)、抗-CD16/CD32抗体(93; eBioscience)と 共に一晩4℃、湿潤箱中で抗原抗体反応させた。Histofine Simple Stain Mouse MAX-PO(ニチレイ、東京)で1時間反応後、Histogreen(Abcysonline, Paris, France)
で5分間発色させた。
6. FITC-E.coli、FITC-S.aureus貪食能の評価
生体におけるKupffer細胞の貪食能を評価するため、FITC標識E.coli、FITC 標識S.aureus(FITC-E.coli, FITC-S.aureus; BioParticles, Invitrogen Life
Technologies)、または直径0.75 μmのFITC- beads(Fluoresbrite YG 0.75 micron
6
microspheres; Polysciences, Warrington, PA, USA)をマウス尾静脈に静注し、5分 後に肝を摘出した。そして、その摘出肝から分離した単核球をフローサイトメ トリーで評価した。
7. ファゴライソゾーム形成の評価
pHrodoはpHが低下する酸性環境で初めて蛍光発色する色素で、ファゴソー
ムとライソゾームとの融合(ファゴライソゾームの形成)の検出に用いられる。
pHrodoで標識されたE.coli(BioParticles, Invitrogen Life Technologies)20 μLと肝 単核球(5 105細胞/200 μL)を混合し37℃、5% CO2で30分間培養した。その 後フローサイトメトリーで蛍光発色を検出することで、細胞内でのファゴライ ソゾームの形成を評価した。
8. Resident Kupffer細胞の除去
Clodronate(LKT Laboratories, St. Paul, MN, USA)をLiposomeに封入し25 mg/mLに調整したClodronate-liposome 100 μLを、実験開始36時間前に予めマウ スに腹腔内投与することでResident CD32/CD68+ Kupffer細胞を除去した(以下、
Clodronate前処置マウス)。
9. S.aureus感染に対する抵抗性の評価
Clodronate前処置マウス群に3 108 colony forming unit(CFU)のS.aureus(209P;
ATCC 6538P)を尾静注し、通常飼育下で観察し生存率を測定した。コントロー
ル群にはClodronate-liposomeの代わりに生理食塩水を腹腔内投与した生理食塩
水前処置マウスを用いた。
7
10. 残菌数の測定
3 108 CFUのS.aureus を静注後、経時的に肝脾の摘出と採血を行い、各臓器 をホモジェネートしPBSで懸濁液とした。各臓器の懸濁液と血液をPBSで10 倍段階希釈しBHI(brain-heart infusion)寒天培地に塗布して37℃で12時間培養 した。培養後のコロニー数を測定することで各臓器または血中の残菌数とした。
11. S.aureusに対するMCP-1産生能の評価
Clodronate前処置マウスに3 108 CFUのS.aureusを尾静注し経時的に眼窩底 から採血し血清を得た。また、分離した肝単核球(5 105細胞/200 μL)を1 x 107
CFUのS.aureusと37°C、5% CO2で共培養し、経時的に上清を収集した。マウ
ス血清または培養上清検体のMCP-1濃度をELISA kits for mouse MCP-1(BD Biosciences)を用いて測定した。
12. EL-4腫瘍に対する抵抗性の評価
C57BL/6マウス由来の腫瘍細胞であるEL-4細胞を転移実験に用いた。麻酔下
で脾を露出できる程度の小切開を左側腹に置き開腹した。直視下で1 106個の EL-4細胞を脾内に注入し、経門脈的腫瘍肝転移モデルを作成した(22)。閉腹後 は通常飼育を行い、生存率を測定した。また、EL-4細胞注入後20日目に肝を摘 出し、摘出肝表面の転移数を計測した。
13. EL-4細胞に対する細胞傷害活性、IL-12およびIFN-産生能の評価
EL-4細胞を10 μg/mLのcalcein-AM(Kohjin Bio、埼玉)で標識した後、分離 した肝単核球(5 105細胞/200 μL)と共に37°C、5% CO2で4時間培養した。
E/T比(effector-to-target ratio)は25:1、50:1、100:1とした。Terascan(Mineva Tech、
8
東京)で細胞傷害活性を測定した。
また、E/T比 100:1で4時間培養した上清を収集し、上清中のIL-12とIFN-
の濃度をELISA kits for mouse total IL-12(Pierce Biotechnology, Woburn, MA, USA)とELISA kits for mouse IFN-(BD Biosciences)を用いて測定した。
14. Ly5.1骨髄移植キメラマウスの作成
致死量(9 Gy)の放射線を全身照射したLy5.2 B6マウスにLy5.1 B6マウス(三 協ラボサービス、茨城)から分離した骨髄細胞1 107個を尾静注することで、
骨髄移植キメラマウスを作成した。作成したキメラマウスは全例が生存した。
15. 統計学的解析
各実験における結果は、平均値±標準誤差で算出した。SAS software program version 9.2(SAS Institute, Cary, NC, USA)を用いて統計学的解析を行った。生存 率の2群間比較はWilcoxon rank sum testを用いて検定した。その他はStudent’s t-testにて検定した。P < 0.05を以って統計学的に有意差ありとした。
9
第3章 結 果
1. マウス肝F4/80+ Kupffer細胞/マクロファージ(M)の分類
通常C57BL/6マウスの肝から分離した単核球の約4分の1がF4/80+ Kupffer 細胞/Mであった(図1A)。肝F4/80+細胞のほとんどはGr-1-であり、また好中 球を示唆するGr-1+ CD11b+のスポットがF4/80陰性ゲートに存在したため、肝
F4/80+細胞は肝内好中球とは異なる細胞群であることを確認した(図1A)。現
在、マウスのCD32(FcRII)を特異的に同定する抗体がないため、本研究では 抗-CD16/CD32抗体を代わりに用いた。F4/80+ CD16/CD32+細胞がNK1.1陰性で ある一方、F4/80- CD16/CD32+細胞がNK1.1陽性である(図1B)ため、前者を CD32+ Kupffer細胞、後者をCD16+ NK細胞と定義した。肝F4/80+ Kupffer細胞/M
は、CD11b+細胞とCD68+細胞に分けられるが、そのいずれにも属さないCD11b- CD68-細胞が32.0%程度存在した(図2A)。抗-CD16/CD32抗体を用いると、
CD11b+細胞とCD32+細胞に分類することができ、いずれにも属さないCD11b- CD32-細胞は19.9%であった(図2B)。CD32+細胞とCD68+細胞の関係をみると 19.4%が重複していた(図2C)。CD32とCD68を同色で共染色し、さらにCD11b を用いて展開すると、F4/80+細胞をこれらの2群に分けることができ、その陰性 細胞は9.8%であった(図2D)。したがって、この染色方法によってほとんどの
Kupffer細胞を抗原別に分類することができるものと考えられた。以上の結果を
まとめ、F4/80+ Kupffer細胞/Mにおける3つの型の割合を模式的に示した(図 2E)。
2. CD11b+細胞とCD32+細胞/CD68+細胞の形態ならびに細胞生物学的相違
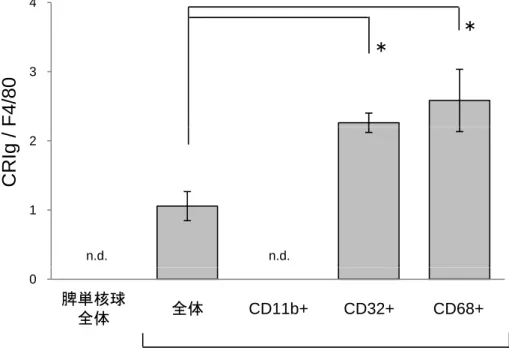
CRIgは、肝のMに特異的に発現し、かつ貪食能を反映している補体受容体
である(14)。そこで、マウスの単核球におけるCRIgの発現について定量PCRを
10
用いて調べたところ、脾単核球はCRIgを全く発現していなかったのに対し、肝 単核球でCRIgの発現を認めた(図3)。肝単核球をMACS法で分取したCD11b+ 細胞、CD32+細胞、CD68+細胞から抽出したRNAをもとに評価したところ、
CD11b+細胞ではCRIg発現を認めなかったのに対し、CD32+細胞とCD68+細胞で CRIgの発現を認めた(図3)。このことから、CD11b+細胞ではなくCD32+細胞 やCD68+細胞が特徴的な性質を有する細胞群である可能性が考えられた。実際 に、正常マウスの肝以外の臓器としての脾や末梢血のF4/80+単核球を調べたと ころ、CD11b+細胞がほとんどでCD32+細胞やCD68+細胞はごくわずかしか認め られなかった(図4)。したがって、CD32+細胞/CD68+細胞は肝固有に存在する 細胞群である可能性が考えられた。
肝のCD11b+細胞、CD32+細胞、CD68+細胞を免疫組織化学で染色しその形態 を観察したところ、CD32+細胞とCD68+細胞が星状または紡錘形であった(図
5A, C)のに対し、CD11b+細胞は円形~卵円形であり(図5B)、形態的に全く
異なる細胞群であることが分かった。また、CD11b+細胞の数はCD32+細胞や CD68+細胞よりも少なく、その大きさも小さかった(図5A-C)。肝組織内の分 布についても、CD32+細胞とCD68+細胞は共に類洞に沿い存在していた(図5A, C)のに対し、CD11b+細胞は肝全体に点在していた(図5B)。
フローサイトメトリー解析でも、細胞の大きさを反映するFS(forward scatter)
と細胞内成分や構造物の複雑さを反映するSS(side scatter)で展開した分布が、
CD32+細胞とCD68+細胞ではほとんど同じく広範に分布したのに対し(図6; 淡 青色と濃青色)、CD11b+細胞は限局的な分布を示していた(図6; 赤色)。FS 値についても、CD11b+細胞は425±5と低値であったのに対し、CD32+細胞では 466±7、CD68+細胞では481±13と高値を示した(P < 0.01;図6)。SS値の違い も考慮に入れると、CD11b+細胞とCD32+細胞/CD68+細胞は異なる細胞群に類別
11
できることが判明した。
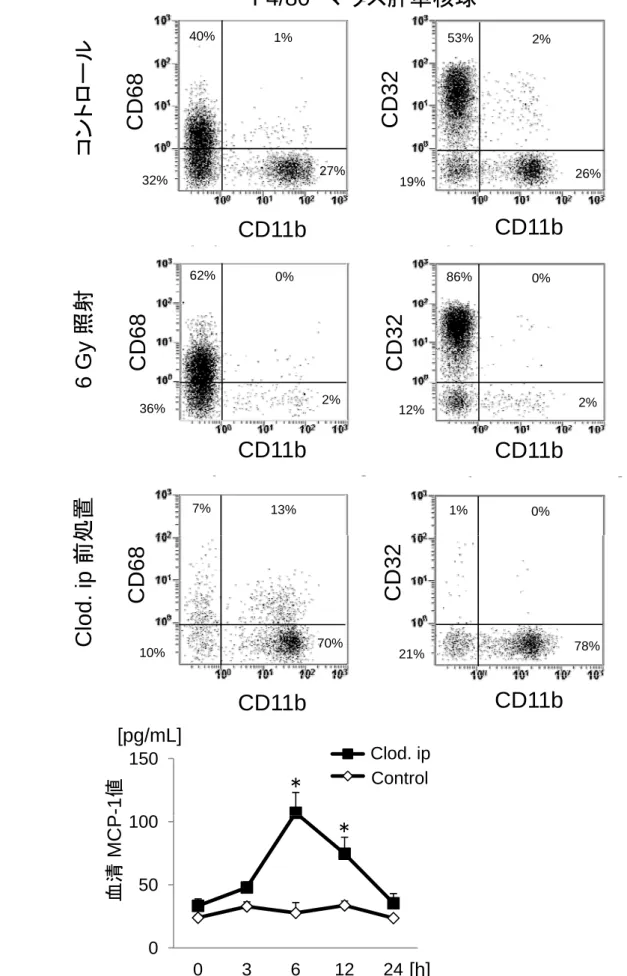
マウスに非致死量(6 Gy)の放射線を照射し、照射後6日目にF4/80+単核球を 観察したところ、肝内のCD11b+細胞はほとんど消失したが、CD32+細胞とCD68+ 細胞は残存した(図7B)。CD11b+細胞は、脾、骨髄、末梢血のいずれにおいて も同様に消失していた。また、このような放射線照射マウスに骨髄移植をする
と、肝CD11b+細胞の再構築が認められた(図示せず)。
一方、マウスをClodronateで前処置すると、放射線照射時とは逆に、CD32+ 細胞/CD68+細胞が除去され、CD11b+細胞は除去されずに増加した(図7C)。そ して、末梢血や脾のCD11b+細胞も除去されなかった(図示せず)。
以上から、F4/80+ Kupffer細胞/Mは、肝固有に存在し放射線抵抗性/Clodronate 感受性のResident CD32+細胞/CD68+細胞と、骨髄由来で放射線感受性/Clodronate
抵抗性のRecruited CD11b+細胞から成り、両者の発生・分化過程が大きく異なる
ことが示唆された。
3. マウスF4/80+ Kupffer細胞の貪食能とファゴライソゾーム形成能
マウスにFITC-E.coliまたはFITC-S.aureusを尾静注投与すると、Kupffer細胞 の貪食菌数はCD68+細胞>CD32+細胞>CD11b+細胞の順であった(図8A, B)。ま た、ファゴソームがライソゾームと融合して酸性環境になると蛍光発色する pHrodo-E.coliを肝単核球と共培養すると、CD11b+細胞ではpH低下に伴う蛍光 発色が全くないことから、有効な貪食とそれに引き続くファゴライソゾーム形 成、すなわち殺菌能が認められないことが判明した(図8C)。これに対し、CD32+ 細胞とCD68+細胞はそれぞれ4.9%、18.5%の細胞にファゴライソゾーム形成が認 められた(図8C)。また、CD32+細胞はCD68+細胞と比べて貪食・殺菌能が弱 かったため、CD68+細胞よりも機能的に未熟な細胞であることが示唆された。
12
4. マウスCD68+細胞の前駆細胞としてのCD32+細胞
マウスにFITC-E.coliまたはFITC-S.aureusを静注して5分後に肝単核球を調べ ると、F4/80+ CD32+細胞は数が減り、F4/80+ CD68+細胞は増加していた(図9A, B)。
同時に、CD32発現の程度を示すMFI(Mean fluorescence intensity)も菌投与後 に有意に減少した(図9A, B)。この結果から、CD32+ Kupffer細胞は菌を貪食 した後CD68を発現しCD32を失う可能性が考えられた。実際、CD32+細胞の多 くは造血幹細胞の分子マーカーであるc-kit(CD117)を多く発現していた(図 10A)。CD32+ CD68-細胞の約3分の2(67.7%)がc-kitを発現していたが、CD32+ CD68+細胞では18.3%、CD32- CD68+細胞は2.5%であった(図10B)。また、別 の幹細胞マーカーであるCD34の発現についても同様のパターンを示した(デー タ示さず)。したがって、肝F4/80+ CD32+細胞は、機能的にもF4/80+ CD68+細胞 よりも未熟な細胞群であったことからも、F4/80+ CD68+細胞の前駆細胞であると 考えられた。
5. Clodronate前処置マウスにおける抗菌能の低下と抗腫瘍能の亢進
マウスにClodronateを投与すると、肝CD32/CD68+ Kupffer細胞が除去され、
CD11b+細胞が残った(図7C)。このようなClodronate前処置マウスに3 108 CFU
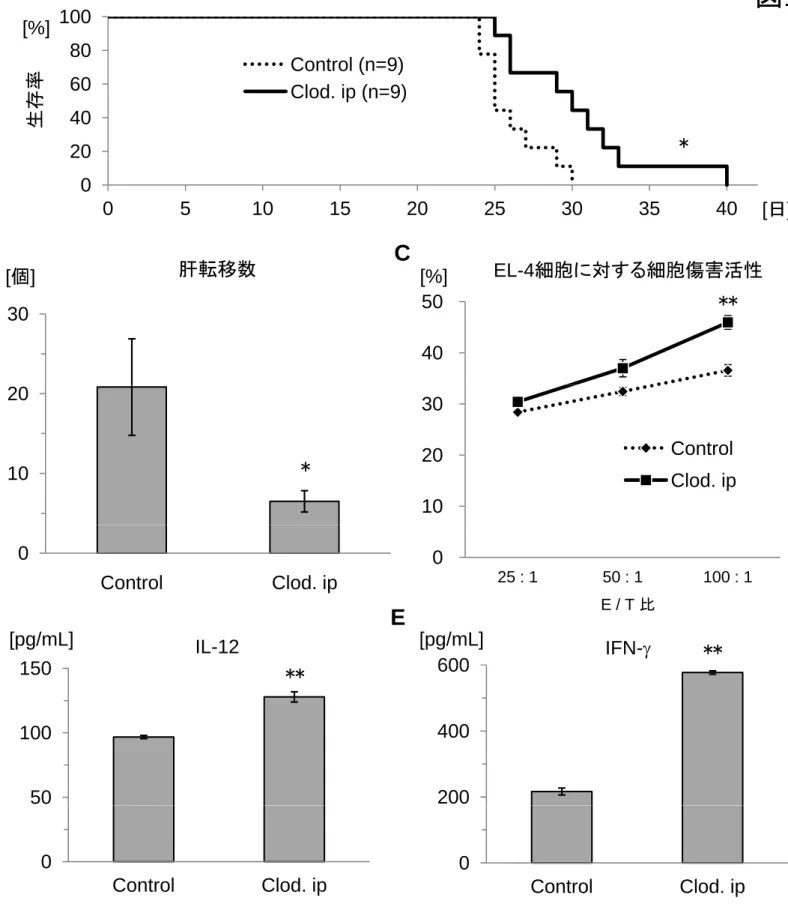
のS.aureusを静注感染させると、2日以内に全例が死亡した(図11A)。また、
Clodronate前処置によって肝内のCD32/CD68+細胞は除去されて肝での貪食活性
が落ちることで、感染20分後における肝内の菌数は有意に減少した(図11B)。
逆に、このようなマウスではその分血中からの菌排除が著しく遅延した(図11B)。
そして、感染11時間後では脾に多くの生菌が残存した(図11C)。この結果は、
CRIg欠損マウスにおいて血中を循環する病原体に対する貪食能が著しく低下す るという報告(14)と合致する。さらに、Clodronate前処置による肝CD32/CD68+
13
細胞の除去は、S.aureus感染後3時間目の血清MCP-1濃度上昇を有意に抑制し た(図11D)。Clodronate前処置マウスから分離した肝単核球をS.aureusと共培
養し、in vitroにおけるMCP-1を測定しても、48時間まで全く産生が認められな
かった(図11E)。以上のことから、Clodronate感受性であるCD32/CD68+細胞
が主にMCP-1を産生していることが示唆された。
しかしながら、Clodronate前処置マウスの脾にEL-4腫瘍細胞を注入し肝転移 を起こすと、驚くべきことに、コントロール群よりも長く生存した(図12A)。
EL-4細胞脾注後20日目における肝表面の転移数をみると、Clodronate前処置マ ウス群(6.5±1.3個)はコントロール群(20.8±6.1個)よりも低値を示した(図 12B、図13)。また、EL-4細胞に対する細胞傷害活性をみても、Clodronate前処 置マウスから分離した肝単核球の方がコントロールマウスの肝単核球よりも高 値を示した(図12C)。さらに、Clodronate前処置マウスの肝単核球をEL-4細 胞と共培養すると、コントロールマウスの肝単核球よりも高いIL-12および IFN-の産生能を示した(図12D, E)。Clodronate前処置マウスでは、CD11b+細 胞の一部が細胞表面にCD68を発現しており(図7C)、またフローサイトメト
リーのFS解析でCD11b+細胞の大きさが増大していた(図示せず)ことから、
CD32/CD68+細胞の除去がCD11b+細胞を増加させるだけでなく機能的にも活性
化させているものと考えられた。このような可能性の一方で、Clodronateによっ
てCD32/CD68+細胞にアポトーシスが誘導される際に一時的にMCP-1を産生し
(図7D)、肝に炎症状態をもたらした結果、CD11b+細胞が骨髄から誘導され増
加した可能性もある。そして、肝のCD11b+細胞がCD68+細胞の機能を代償して いたとも考えられる。以上から、Clodronate前処置マウスでは、CD11b+細胞の
IL-12産生能が亢進しており、このことが結果的に肝NK/NKT細胞の抗腫瘍活性
を増強しているものと推察された。
14
6. 骨髄移植キメラマウスを用いた肝CD11b+細胞、CD32/CD68+細胞の発生学的 由来の検索
B6(Ly5.2)マウスに致死量である9 Gyの放射線を照射した後、Ly5.1マウス から分離した骨髄細胞1 107個/匹を尾静脈から注入移植したところ、全例が生 存した。このような骨髄移植後4週目のマウスから肝単核球を分離して調べた ところ、F4/80+ CD11b+細胞はほぼ全てがLy5.1マウス由来細胞を示すCD45.1抗 原を発現していた(図14A左)。一方、F4/80+CD11b-細胞のほとんどがCD45.1 陰性であった(図14A左)。したがって、骨髄移植キメラマウスにおいて、肝 単核球中のCD11b+細胞は骨髄由来であることが分かった。逆に、CD11b-細胞は 骨髄造血幹細胞に由来しない肝固有の放射線抵抗性の細胞群であることが分か った。実際、このキメラマウスのF4/80+ CD32/CD68+細胞はCD45.1陰性であっ た(図14A右)。また、骨髄移植後に成熟分化した細胞が末梢に定着する8週 目においても4週目とほぼ同様の傾向を示した(図14B)。すなわち、F4/80+ CD32+ 細胞ならびにF4/80+ CD68+細胞は骨髄由来ではなく、発生学的にYolk Sac由来 である(17, 18)ことが示唆された。
7. 致死量9 Gy照射マウス肝Kupffer細胞の貪食・殺菌能
9 Gy全身照射後6日目のB6マウスにおいて、骨髄由来F4/80+ CD11b+細胞は、
肝、脾ともにほとんど消失した(図15A, B)。逆に、Resident MであるF4/80+ CD11b-細胞の比率は、肝では27%から42%へ(図15A)、脾では5%から22%
へ(図15B)と、共に増加した。
まず、9 Gy照射後6日目のマウスおよび9 Gy照射後骨髄移植を行ったキメラ マウスの肝Kupffer細胞における貪食能について評価した。非照射コントロール マウス、照射後6日目のマウス、骨髄移植キメラマウスに各々FITC-beadsを尾
15
静注し、5分後に肝を摘出して分離した単核球をフローサイトメトリーで評価す ると、CD68+細胞はCD32+細胞よりも多くFITC-beadsを取り込んでいた(図16A)。 また、9 Gy照射マウスおよびキメラマウスの貪食能は、非照射コントロールマ ウスとほぼ同程度であった(図16A)。
次に、pHrodo-E.coliを用いて非照射コントロールマウス、照射後6日目のマ ウス、骨髄移植キメラマウスから分離した肝単核球のファゴライソゾーム形成 について調べた。pHrodoの発色は、CD68+細胞の方がCD32+細胞よりも強い活 性を示した。9 Gy照射マウスおよびキメラマウスのファゴライソゾーム形成能 は非照射コントロールマウスと比べ有意な差は認められなかった(図16B)。キ メラマウスのCD11b+細胞は非照射コントロールマウスのCD11b+細胞と同様に ファゴライソゾーム形成を全く示さなかった(図16B)。以上のことから、
CD32/CD68+細胞は、肝Kupffer細胞の貪食殺菌の中心的役割を果たしており、
機能的にも強い放射線抵抗性を有していることが判明した。
8. ヒト肝CD14+ Kupffer細胞/マクロファージの分類と貪食能
ヒト肝から分離した単核球中のCD14+ Kupffer細胞についてCD11b、CD32、
CD68の発現を評価した。ヒト肝からのKupffer細胞の分離はマウスよりも困難 で、collagenase処理を行ってもCD14+ Kupffer細胞はヒト肝単核球の約10%を占 める程度であった。しかし、マウスのKupffer細胞と同様、ヒトCD14+ Kupffer 細胞もCD11b+細胞、CD32+細胞、CD68+細胞の3つの分画に分類することがで きた(図17A)。CD11b-細胞の約70%がCD32を、約60%がCD68をそれぞれ 発現していたため、CD11b-細胞中では大部分の細胞がCD32とCD68を共発現
(CD32+ CD68+)していることになる。ヒトでは、マウスと異なり、肝や末梢血 中にCD11b+ CD32+細胞が存在した(図17A, B)。しかし、ヒト肝でみられる
16
CD11b- CD32+細胞やCD11b- CD68+細胞(図17A)は、ヒト末梢血中にほとんど なかった(図17B)。そのため、ヒトにおいてもマウスと同様にCD11b+細胞で はないCD32/CD68+細胞が肝固有に存在するResident Kupffer細胞であると考え られた。
また、ヒト肝単核球をpHrodo-E.coliと共培養すると、CD11b+ Kupffer細胞/M
はほとんどファゴライソゾーム形成を示さなかった(3.5%のみ)のに対し、
CD32/CD68+ Kupffer細胞は顕著なファゴライソゾーム形成を示した(51.4%)(図 17C)ことから、マウスと同様の機能分担が存在することが示唆された。
17
第4章 考 察
肝マクロファージ(M)であるKupffer細胞は、その特別な解剖学的、生理 学的な微小環境により、貪食殺菌、TNF等のサイトカイン産生、抗腫瘍作用等 いくつかの独特な機能を持つと考えられてきた(23, 24)。しかし、Kupffer細胞を 発生・分化および機能的な観点から明確に分類できる有用な方法は、これまで 存在しなかった。
本研究では、マウスF4/80+ Kupffer細胞およびヒトCD14+ Kupffer細胞を、
CD11b、CD32、CD68という細胞表面抗原の発現によって3つのグループに分類
できることを示した。本分類は、ほとんど全てのKupffer細胞を漏れなく類別す ることができる。この方法で分類したCD32/CD68+細胞は肝固有のResident Kupffer細胞であり、骨髄由来のCD11b+ Recruited Kupffer細胞/Mとは異なる系
統のMであることが明らかとなった。
免疫組織化学による観察では、CD11b+細胞が肝内に点在したのとは異なり、
CD32/CD68+細胞は肝類洞に沿って分布したことから、これらは肝類洞内皮細胞
や肝細胞に固着している細胞群であると考えられた。これは、肝組織を
collagenaseで処理しなくてもCD11b+細胞は分離できるが、CD68+細胞の分離に はcollagenase処理が必要であること(9)と合致する。また、CD32/CD68+細胞は
CD11b+細胞よりも大型で星形や紡錘形を呈したことから、形態学的にも大きく
異なる。
肝に特異的に発現し、かつ貪食能を反映するCRIg (14)が、CD32/CD68+細胞に は存在するがCD11b+細胞や脾単核球では認められなかったことは、CD32/CD68+ 細胞が肝固有のResident Kupffer細胞であることを示唆する。さらに、B6マウス に6 Gyの放射線を照射すると、肝のCD32/CD68+細胞は抵抗性を示し生残した のに対し、CD11b+細胞は放射線感受性で照射後にほとんどが消失した。放射線
18
照射後の骨髄移植によって肝内に再構築できた細胞群がCD11b+細胞であったこ とから、CD11b+細胞は骨髄由来のRecruited Kupffer細胞/Mであることが証明さ れた。
実験的に肝Kupffer細胞除去効果を持つとされるClodronate ip投与モデルマウ スの肝単核球を観察したところ、Resident CD32/CD68+細胞は除去されるが Recruited CD11b+細胞は除去されないことが分かった。これまで、Clodronateや GdCl3(塩化ガドリニウム)投与で全てのF4/80+ Kupffer細胞を除去できると考 えられてきたが、それは単に、免疫組織化学によって切片上でF4/80+細胞が激 減することから推定されたことである。しかし、本研究のフローサイトメトリ ーならびに免疫組織化学によって、Clodronate投与はResident CD32/CD68+細胞 を選択的に除去し、CD11b+細胞は末梢血単球や脾Mと同様に除去されないこと が明らかとなった(9, 11, 25, 26)。
MにおけるCRIgの発現は病原体の貪食に必要であることが知られている
(14)。FITC-E.coliや FITC-S.aureusを静注感染させたマウスで肝単核球による貪 食を観察すると、CRIgを発現するCD32/CD68+細胞を中心として活性が認めら れた。その中で特に、CD68+細胞が強い貪食能と機能的なファゴライソゾーム形 成を示したことは、本細胞がROS産生能を有するというこれまでの報告(9)と合 致している。CD68はスカベンジャー受容体の1つであり、膜融合や抗原処理に 関与する(27-29)。したがって、こうした機能を反映するCD68+細胞がResident
Kupffer細胞として肝における病原体排除を司っているものと考えられる。
FITC-E.coliやFITC-S.aureusを静注すると、その直後にCD68+ CD32-細胞が増え、
CD68- CD32+細胞が減るという現象が観察された。そこで本研究では、「CD32+ 細胞はCD68+細胞の前駆体であり、貪食・活性化しCD68を発現するようになる」
という仮説を立てた。未熟前駆細胞が発現する造血幹細胞の分子マーカーであ
19
るc-kit(CD117)およびCD34を調べたところ、大部分のCD32+細胞はc-kitや CD34を発現していたが、CD68を発現するに伴ってc-kitとCD34の発現率は減 少した。したがって、CD32+細胞はより未分化な分画であり、肝固有のResident
Kupffer細胞であるCD68+細胞の前駆体であると考えられた。放射線照射実験か
らも分かるように、CD32/CD68+細胞は骨髄からの細胞供給を受けることなく肝 内で高い分化・再生能を有することが示された。このことは、c-kit+ CD32+ Kupffer 細胞が単球やCD11b+ Mとは異なりYolk Sacで独自に発生する細胞系譜である という考え(17, 18)を支持する。
他方、pHrodo-E.coliを用いた実験から、CRIgを持たないCD11b+細胞は、
CD32/CD68+細胞とは対照的に、機能的なファゴライソゾームを形成できないこ
とが分かった。したがって、肝のCD11b+細胞には効果的な殺菌能は備わってお らず、E.coli感染後の肝CD11b+細胞がROSを産生しないという過去の報告(9) にも合致する。
Clodronate処理によってResident Kupffer細胞(CD32/CD68+細胞)を除去する
と、S.aureus静注感染後マウスの生存率は顕著に低下した。しかし、驚くべきこ
とに、EL-4腫瘍細胞の経門脈性肝転移を抑制し、生存率も改善させた。すなわ
ち、肝CD32/CD68+細胞は抗腫瘍免疫には直接的に関与しておらず、おそらく
CD11b+細胞がIL-12を産生することでNK/NKT細胞を活性化し抗腫瘍免疫に重
要な役割を演じていると考えられる。「肝における抗腫瘍作用に直接関与する のはasialo GM1陽性細胞(NK細胞)であって、GdCl3で除去しえるKupffer細 胞ではない」と述べているShiratoriらの報告(30)は、これを支持する。また、Shono ら(31)も、「高脂肪食で飼育したマウスではCD11b+細胞が増え、それが活性化
してIL-12産生が増加することによって肝NK/NKT細胞の抗腫瘍活性が亢進す
る」ことを示している。
20
貪食能旺盛なCD32/CD68+細胞は、S.aureusを投与するとそれらを貪食し活性 化することで、C-C chemokine receptor 2(CCR2)のリガンドであるMCP-1を産 生する。CCR2欠損マウスを用いた実験では、炎症刺激を加えた際に腹腔や肝へ のマクロファージ集積が劇的に減少したとの報告があること(32, 33)からも、
CD11b+細胞の集積にはCD32/CD68+細胞が産生するMCP-1が必要不可欠である と考えられる。
Clodronate前処置によるCD32/CD68+細胞の除去は、感染マウスにおける肝内
の細菌数を減少させ、逆に循環血中や脾の細菌数を増加させた。したがって、
肝CD32/CD68+細胞は、血中の細菌を積極的に肝内に取り込み、殺菌することで、
全身の細菌排除に貢献していると考えられた。すなわち、CD32/CD68+ Kupffer 細胞は、敗血症/菌血症を防ぐ重要な担い手となっていることが推測される。こ のことは、リコンビナントCRPがKupffer細胞のFcRII(CD32)に結合してオ プソニン効果を発揮し致死的細菌感染モデルマウスを救命しえたという過去の 報告(34)とも一致する。
Kupffer細胞の起源を確かめるため、Ly5.2 B6マウスに致死量の放射線(9 Gy) を照射後にLy5.1 B6マウスの骨髄細胞を移植しキメラマウスを作成した。そし て、肝に定着したKupffer細胞/Mの表面抗原を検索したところ、骨髄移植後4 週、8週において、肝F4/80+ CD11b+細胞はいずれもCD45.1陽性(ドナー型)で あり、骨髄由来であることが示された。これに対し、肝F4/80+ CD32/CD68+細胞
はCD45.1陰性であったため、骨髄造血幹細胞に由来しない肝固有のResident細
胞群がレシピエントマウスに残存し続けているものと考えられた。
また、9 Gyの放射線照射によって、肝だけでなく脾のF4/80+ CD11b+細胞も消 失し、その後の骨髄移植によって再構築された。他方、肝脾共に、F4/80+ CD11b- 細胞は放射線抵抗性であり、骨髄移植を行わなくとも照射後6日目にはその細
21
胞の比率が増加したことから、組織内での自己再生能が示唆された。さらに、9 Gy照射後6日目のマウスや骨髄移植キメラマウスにおける肝Kupffer細胞の貪 食・殺菌能を評価したところ、FITC-beadsの貪食能、pHrodo-E.coliによるファ ゴライソゾーム形成能共に、非照射正常コントロールの機能と全く変わらなか った。したがって、Resident Kupffer細胞は、放射線抵抗性を示して生残するだ けでなく、貪食・殺菌という機能面においても障害されないことが明らかにな
った。Yamadaらも、ストロンチウム-89による骨髄細胞除去後にみられる
Resident Kupffer細胞が増殖能を保持していることを報告(35)しており、本研究の
結果と合致する。以上のように、本研究において、Resident CD32/CD68+ Kupffer 細胞の機能や発生・分化について解明できたことは意義深いと考えられる。
マウスと同様ヒト肝CD14+ Kupffer細胞も、CD11b+細胞、CD32+細胞、CD68+ 細胞の3つの細胞群に分けることができた。CD11b+ CD32+細胞がヒトに認めら れる点はマウスと異なるものの、ヒトにおいても肝で認められ末梢血にはない
CD11b陰性分画の CD32/CD68+細胞がマウス同様に肝固有の細胞群と考えられ
た。pHrodo-E.coliを用いた貪食実験から、効果的な殺菌能を有する細胞は、
CD11b+ Kupffer細胞/MでなくCD32/CD68+ Kupffer細胞であることが分かった。
このように、ヒトにおいてもマウスで確認できたKupffer細胞/Mの基本的特徴 を保持していると考えられた。
22
第5章 結 論
1. マウスF4/80+およびヒトCD14+肝Kupffer細胞/Mは、CD32/CD68+細胞と
CD11b+細胞に包括的に分類することができた。
2. CD32/CD68+細胞は、放射線抵抗性/Clodronate感受性で、肝固有の細胞群で あり、CRIgを発現していた。一方、CD11b+細胞は、CRIgを持たず、放射線
感受性/Clodronate抵抗性の骨髄由来細胞であった。
3. CD32+細胞は、表面抗原的特徴(c-kit+、CD34+)からCD68+細胞より未熟で あり、機能的特徴から貪食活性の強いCD68+細胞の前駆体と考えられた。
4. Clodronate前処置マウスを用いた検討から、CD32/CD68+細胞は抗菌能、
CD11b+細胞は抗腫瘍能という、生体防御に必須な2大機能を明確に分担して
いることが分かった。
5. 致死量の9 Gy放射線照射後も、CD32/CD68+細胞は肝内に残存し、その機能 を維持しえた。
23
謝 辞
稿を終えるにあたり、本研究における直接の御指導ならびに御校閲を賜りま した防衛医科大学校免疫微生物学講座教授 関 修司先生に深甚なる感謝の意 を表します。また、多岐にわたり御指導と御配慮をいただいた防衛医科大学校 分子生体制御学講座教授 四ノ宮成祥先生、ならびに臨床検査医学講座講師 緒方 衝先生に感謝の意を表します。さらに全般にわたり御助力いただきました 防衛医科大学校免疫微生物学講座准教授 木下 学先生、同助教 中島 弘幸先生、
同助教 中島 正裕先生に感謝の意を表します。
24
引 用 文 献
1. Seki S, Habu Y, Kawamura T, Takeda K, Dobashi H, Ohkawa T, et al. The liver as a crucial organ in the first line of host defense: the roles of Kupffer cells, natural killer (NK) cells and NK1.1 Ag+ T cells in T helper 1 immune responses.
Immunol Rev. 2000;174(1):35-46.
2. Seki S, Nakashima H, Kinoshita M. The liver as a pivotal innate immune organ.
Immuno-Gastroenterology. 2012;1(2):76-89.
3. Thomson AW, Knolle PA. Antigen-presenting cell function in the tolerogenic liver environment. Nat Rev Immunol. 2010;10(11):753-66.
4. Toth CA, Thomas P. Liver endocytosis and Kupffer cells. Hepatology.
1992;16(1):255-66.
5. Benacerraf B, Sebestyen MM, Schlossman S. A quantitative study of the kinetics of blood clearance of P32-labelled Escherichia coli and Staphylococci by the reticuloendothelial system. J Exp Med. 1959;110(1):27-48.
6. Cancro MP. Editorial: phagocytic B cells: deja vu all over again? J Leukoc Biol.
2012;91(4):519-21.
7. Nakashima M, Kinoshita M, Nakashima H, Habu Y, Miyazaki H, Shono S, et al.
Pivotal advance: characterization of mouse liver phagocytic B cells in innate immunity. J Leukoc Biol. 2012;91(4):537-46.
25
8. Ashare A, Monick MM, Powers LS, Yarovinsky T, Hunninghake GW. Severe bacteremia results in a loss of hepatic bacterial clearance. Am J Respir Crit Care Med. 2006;173(6):644-52.
9. Kinoshita M, Uchida T, Sato A, Nakashima M, Nakashima H, Shono S, et al.
Characterization of two F4/80-positive Kupffer cell subsets by their function and phenotype in mice. J Hepatol. 2010;53(5):903-10.
10. Klein I, Cornejo JC, Polakos NK, John B, Wuensch SA, Topham DJ, et al.
Kupffer cell heterogeneity: functional properties of bone marrow derived and sessile hepatic macrophages. Blood. 2007;110(12):4077-85.
11. Holt MP, Cheng L, Ju C. Identification and characterization of infiltrating macrophages in acetaminophen-induced liver injury. J Leukoc Biol.
2008;84(6):1410-21.
12. Berken A, Benacerraf B. Properties of antibodies cytophilic for macrophages. J Exp Med. 1966;123(1):119-44.
13. Ravetch JV, Bolland S. IgG Fc receptors. Annu Rev Immunol. 2001;19:275-90.
14. Helmy KY, Katschke KJ, Jr., Gorgani NN, Kljavin NM, Elliott JM, Diehl L, et al. CRIg: a macrophage complement receptor required for phagocytosis of circulating pathogens. Cell. 2006;124(5):915-27.
26
15. Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593-623.
16. Devitt A, Moffatt OD, Raykundalia C, Capra JD, Simmons DL, Gregory CD.
Human CD14 mediates recognition and phagocytosis of apoptotic cells. Nature.
1998;392(6675):505-9.
17. Schulz C, Gomez Perdiguero E, Chorro L, Szabo-Rogers H, Cagnard N, Kierdorf K, et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science. 2012;336(6077):86-90.
18. Takahashi K. Development and differentiation of macrophages and related cells:
Historical review and current concepts. J Clin Exp Hematop. 2001;41(1):1-33.
19. Gangopadhyay A, Bajenova O, Kelly TM, Thomas P. Carcinoembryonic antigen induces cytokine expression in Kuppfer cells: implications for hepatic metastasis from colorectal cancer. Cancer Res. 1996;56(20):4805-10.
20. Nakashima H, Kinoshita M, Nakashima M, Habu Y, Shono S, Uchida T, et al.
Superoxide produced by Kupffer cells is an essential effector in concanavalin A-induced hepatitis in mice. Hepatology. 2008;48(6):1979-88.
21. Toth CA, Thomas P, Broitman SA, Zamcheck N. Receptor-mediated endocytosis of carcinoembryonic antigen by rat liver Kupffer cells. Cancer Res.
1985;45(1):392-7.
27
22. Nakagawa R, Nagafune I, Tazunoki Y, Ehara H, Tomura H, Iijima R, et al.
Mechanisms of the antimetastatic effect in the liver and of the hepatocyte injury induced by alpha-galactosylceramide in mice. J Immunol.
2001;166(11):6578-84.
23. Gordon S. Alternative activation of macrophages. Nat Rev Immunol.
2003;3(1):23-35.
24. Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2005;5(12):953-64.
25. Kinoshita M, Uchida T, Nakashima H, Ono S, Seki S, Hiraide H. Opposite effects of enhanced tumor necrosis factor-alpha production from Kupffer cells by gadolinium chloride on liver injury/mortality in endotoxemia of normal and partially hepatectomized mice. Shock. 2005;23(1):65-72.
26. Sato A, Nakashima H, Nakashima M, Ikarashi M, Nishiyama K, Kinoshita M, et al. Involvement of the TNF and FasL produced by CD11b Kupffer cells/macrophages in CCl4-induced acute hepatic injury. PLoS One.
2014;9(3):e92515.
27. Kurushima H, Ramprasad M, Kondratenko N, Foster DM, Quehenberger O, Steinberg D. Surface expression and rapid internalization of macrosialin (mouse CD68) on elicited mouse peritoneal macrophages. J Leukoc Biol.
2000;67(1):104-8.
28
28. Rabinowitz SS, Gordon S. Macrosialin, a macrophage-restricted membrane sialoprotein differentially glycosylated in response to inflammatory stimuli. J Exp Med. 1991;174(4):827-36.
29. Smith MJ, Koch GL. Differential expression of murine macrophage surface glycoprotein antigens in intracellular membranes. J Cell Sci. 1987;87(1):113-9.
30. Shiratori Y, Nakata R, Okano K, Komatsu Y, Shiina S, Kawase T, et al.
Inhibition of hepatic metastasis of colon carcinoma by asialo GM1--positive cells in the liver. Hepatology. 1992;16(2):469-78.
31. Shono S, Habu Y, Nakashima M, Sato A, Nakashima H, Miyazaki H, et al. The immunologic outcome of enhanced function of mouse liver lymphocytes and Kupffer cells by high-fat and high-cholesterol diet. Shock. 2011;36(5):484-93.
32. Dambach DM, Watson LM, Gray KR, Durham SK, Laskin DL. Role of CCR2 in macrophage migration into the liver during acetaminophen-induced hepatotoxicity in the mouse. Hepatology. 2002;35(5):1093-103.
33. Kuziel WA, Morgan SJ, Dawson TC, Griffin S, Smithies O, Ley K, et al. Severe reduction in leukocyte adhesion and monocyte extravasation in mice deficient in CC chemokine receptor 2. Proc Natl Acad Sci U S A. 1997;94(22):12053-8.
34. Inatsu A, Kinoshita M, Nakashima H, Shimizu J, Saitoh D, Tamai S, et al. Novel mechanism of C-reactive protein for enhancing mouse liver innate immunity.
Hepatology. 2009;49(6):2044-54.
29
35. Yamada M, Naito M, Takahashi K. Kupffer cell proliferation and glucan-induced granuloma formation in mice depleted of blood monocytes by strontium-89. J Leukoc Biol. 1990;47(3):195-205.
30
略 語 一 覧 BHI : brain-heart infusion
CCR2 : C-C chemokine receptor 2 CD : cluster of differentiation
cDNA : complementary deoxyribonucleic acid CFU : colony forming unit
CRIg : complement receptor of the Ig superfamily CRP : C-reactive protein
Cy : cyanine
E.coli : Escherichia coli
ELISA : enzyme-linked immunosorbent assay E/T : effector-to-target
FcR : Fc gamma receptor
FITC : fluorescein isothiocyanate FS : forward scatter
HBSS : Hanks’ Balanced Salt Solution IFN- : interferon-gamma
IgG : immunoglobulin G IL : interleukin
LPS : lipopolysaccharide
MACS : magnetic-activated cell sorting MCP-1 : monocyte chemotactic protein-1 M : macrophages
MFI : Mean fluorescence intensity
31
NK : natural killer NKT : natural killer T
PBS : phosphate-buffered saline PCR : polymerase chain reaction PE : phycoerythrin
RNA : ribonucleic acid
ROS : reactive oxygen species S.aureus : Staphylococcus aureus SS : side scatter
TNF : tumor necrosis factor
A
24% 2%
15% 4%
肝単核球 図 1
3% 1%
CD1 1b
F4/80
71% 80%
Gr-1 Gr-1
B B
D16 / CD32
F4/80
CD
CD16 / CD32+ F4/80-gated
CD16 / CD32+ F4/80+gated
70.4% 3.5%
NK1.1 NK1.1
図1. マウス肝F4/80+細胞の同定 コントロールB6マウスの肝から分離した単核球をフローサイトメト リーで測定・解析した。肝単核球全体の4分の1程度がF4/80+であった(A)。また肝内Gr-1+好中球は F4/80陰性ゲートにスポットとして存在した(A)ため、肝F4/80+細胞と鑑別できた。CD16/CD32+F4/80- 細胞はNK1.1陽性であり、CD16/CD32+F4/80+細胞はNK1.1陰性であった(B)。
32
A
F4/80
+肝単核球
8
38.8±2.0%
3 2
50.9±2.0%
図 2
0.9±0.1%
B
1.4±0.2%CD6
32.0±2.0%
28 3±2 0%
CD 3
19.9±2.0%
27 8±1 3%
CD11b
28.3±2.0%
CD11b
8
59.4±1.4%31.1±3.2% 19.4±0.8%
27.8±1.3%
D C
2.5±0.2%
CD32 + CD6
CD32
CD11b
9.8±0.3%
28.3±1.1%
CD68
30.3±2.1%
19.2±2.1%
30%
CD32+
CD68+
20%
20%
E
CD11b+
20%
30%
CD11b+
図2. マウス肝F4/80+Kupffer細胞/マクロファージ(M)の分類 (A-D) フローサイトメトリーでF4/80+ にゲートを掛けた細胞におけるCD11b CD32 CD68の発現 (D)の縦軸にある「CD32+CD68」は 共 にゲートを掛けた細胞におけるCD11b、CD32、CD68の発現。(D)の縦軸にある「CD32+CD68」は、共 染色したことを示す。マウス5匹において同様の結果が得られた。図中の数値は5匹の結果を、平均値
±標準誤差で示す。(E) A-Dの結果をもとに、CD11b+細胞、CD32+細胞、CD68+細胞の関係を示した 模式図。
33
図 3
3 4
*
*
F4/80
1 2
n.d.
n.d.
CRIg /
0
Spleen MNCs
Liver MNCs
CD11b P
CD32 P
CD68
全体 P
脾単核球
全体 CD11b+ CD32+ CD68+
肝単核球
図3. RT-PCRによる脾単核球、肝単核球、分取した単核球各群のCRIg mRNA発現量 結果は同一 検体のF4/80 mRNA発現量との相対比で示す。独立した4回の実験にて同様の結果が得られ、平均 値±標準誤差で示す。*P < 0.05 vs.他群。n.d.はnot detectedの略。
34
A
図 4
F4/80
+マウス脾単核球
1% 1%
0%
0%
A
65%
33%
66%
34%
CD68 CD32
F4/80
+マウス末梢血単核球
33%
B
CD11b CD11b
F4/80 マウス末梢血単核球
0% 0%
6% 5%
68 3 2
80%
20%
75%
14%
CD CD 3
CD11b CD11b
図4. 脾および末梢血中のF4/80+単核球のCD11b発現、CD32発現、CD68発現
(A)脾F4/80+単核球のCD11b、CD32、CD68発現。(B)末梢血F4/80+単核球のCD11b、CD32、CD68 発現。マウス3匹から同様の結果が得られた。
35
図 5
A B
C
CD68 CD32
CD11b
図5. 免疫組織化学によるマウス肝組織のCD11b+細胞、CD32+細胞、CD68+細胞の形態と分布 正常B6マウス肝の新鮮凍結切片を抗-CD68抗体(A)、抗-CD11b抗体(B)、抗-CD16/CD32抗体(C)で 染色した。マウス3匹から同様の結果が得られた。400倍光学顕微鏡にて観察し白線は50 mを示す。
右下窓は800倍の強拡大像を示す。
36
マウス肝単核球 図 6
SS
FS
SS
FS
425±5 ** 466±7 481±13
赤色点 :F4/80+CD11b+細胞 淡青色点:F4/80+CD32+細胞 濃青色点:F4/80+CD68+細胞
図6. マウス肝F4/80+細胞中のCD11b+細胞とCD32+細胞/CD68+細胞のFS/SS分布の違い マウス 肝単核球のフローサイトメトリーによるFS (forward scatter)とSS (side scatter)の分布。赤色は CD11b+細胞を、淡青色はCD32+細胞を、濃青色はCD68+細胞を示す。CD32+細胞とCD68+細胞は FS/SS分布が類似しているが、CD11b+細胞は明らかに異なる分布を示す。独立した4回の実験で同 様の結果が得られた。FS値は平均値±標準誤差で示す。**P< 0.01 vs.他群。
37
図 7
40% 1%
C D68
53% 2%
C D32
F4/80
+マウス肝単核球 A
ト ロール
32% 27%
CD11b
C
19% 26%
CD11b
C
B
コン ト
62% 0%
2%
CD68
86% 0%
CD32
B
6 Gy 照射
7% 13% 1% 0%
36% 2%
CD11b
12% 2%
CD11b C
処 置
10% 70%
CD68
21% 78%
CD32
Clod. ip 前 処
D
100
150 Clodronate
PBS
[pg/mL]
*
-1値
Clod. ip Control
CD11b CD11b
0 50 100
血清MCP
*
図7. マウス肝F4/80+細胞中のCD11b+細胞とCD32+細胞/CD68+細胞の放射線照射およびClodronate 前処置に対する感受性 (A)正常コントロール。(B)放射線によるCD11b+細胞の除去効果。マウスに6 Gy 照射し、照射後6日目のCD11b+細胞、CD32+細胞、CD68+細胞を調べた。マウス4匹から同様の結果が 得られた。(C) Clodronate投与によるCD32+細胞/CD68+細胞の除去効果。マウスにClodronate-
0 3 6 12 24 [h]
liposomeを腹腔内投与(Clod. ip)し36時間後の分画を調べた。マウス4匹から同様の結果が得られた。
(D) Clodronate投与後の血清MCP-1値の推移。Clodronate-liposome投与後経時的に採血し、血清中の MCP-1値を測定した。独立した4回の実験結果を平均値±標準誤差で示す。*P < 0.05 vs.コントロール 群。図中のClod.はClodronate-liposomeの略。
38
F4/80+ CD11b+ gated F4/80+CD32+gated F4/80+CD68+gated
図 8
A
g
41.1%
g
80.3%
/80 C 3 ga ed
73.4%
FITC-E.coli in vivo
B
4.4% 15.7% 77.6%
FITC-S.aureus in vivo
C
0.0% 4.9% 18.5%
pHrodo-E.coli in vitro
図8. FITC-E.coli、FITC-S.aureusを用いた貪食能およびpHrodo-E.coliを用いたファゴライソゾー ム形成の測定 B6マウスの尾静脈にFITC-E.coli (A)またはFITC-S.aureus (B)を静注し5分後に肝を 摘出した。摘出肝から分離した単核球中のFITC発色をフローサイトメトリーで評価した。マウス3-4匹 から同様の結果が得られた。また、正常B6マウスの肝単核球をpHrodo-E.coliと共培養し、ファゴライ ソゾーム中の酸性環境で発色する蛍光をフローサイトメトリーで測定した(C)。独立した3回の実験で同 様の結果が得られた。
39