Carcinological Society of Japan
シンポジウム報告
Cancer 25: 143–148 (2016)
エビヤドリムシ科等脚類の研究:
だから寄生虫の研究はやめられない!
A short revision of some biological notes on Japanese bopyrid isopods, with homage for the
“
cute as crustacean parasites” in my life
齋藤暢宏
1Nobuhiro Saito
はじめに
鮮魚店の店頭などで,鰓域が膨れ上がったホッコ クアカエビPandalus eous Makarov, 1935やクロザコ エビArgis lar (Owen, 1839)などを見かけたことはな いだろうか.これはエビの鰓に寄生したエビヤドリ
ムシ類による“虫コブ”である(図1).ホッコク
アカエビの鰓腔にはアマエビエラヤドリ(新称) Bopyroides hippolytes (Krøyer, 1838),クロザコエビ にはエビジャコヤドリムシArgeia pugettensis Dana, 1852が寄生することが知られる(齋藤ら,2001). エビヤドリムシ科Bopyridaeは等脚目甲殻類の1分 類群であり,十脚目甲殻類を宿主とする寄生生物で ある(和名に“エビ”とあるが,寄生対象は十脚類 全般である).宿主の鰓腔や体腔,腹部腹面・背面 等に寄生し,時に寄生部位を著しく膨瘤させる(椎 野,1952; 齋藤ら,2001).宿主への寄生主体は成 熟雌であり,この体制は寄生適応のため属種によっ ては大きく変形する.各体節は癒合化し,付属肢も 退化・消失し,体型が左右非対称や蠕虫状となるた め,等脚類の基本体型(下村・布村,2010)から著 しく逸脱した形態となる(図2).雄は矮雄で,雌 の腹部腹面等に懸着する.寄生を受けた宿主は栄養 分の一部が寄生種の成長に費やされるとともに,雌 では不妊化し,雄では寄生去勢されることが知られ て い る(Hiraiwa & Sato, 1939; O’Brien & Van Wyk, 1985). 筆者は学部生時代に,本来の研究テーマ(Saito & Kubota, 1995)とは別にこの寄生虫に魅せられて しまい,以来本業の傍らエビヤドリムシ類の研究に 従事している.まだまだ勉強不足で,本テーマを十 分に理解しているとは甚だ言い難いが,本稿では, この魅力あふれるエビヤドリムシ類の生物学を読者 諸兄に紹介してみたい. エビヤドリムシ類の研究意義 私的見解ではあるが,以下に示す5項目をエビヤ ドリムシ類の研究意義と考えている.なお,これら は寄生虫の生物学的,病理学的研究においても,基 礎的な重要項目に当てはまるものと思われる. 1)エビヤドリムシ類相の解明 エビヤドリムシ類は,世界では約600種が知られ
(Boyko & Schotte, 2015),日本からは98種が報告され て い る(齋 藤 ら,2000; Shimomura et al., 2006, 2012; Saito et al., 2010; 三浦ら,2014; Saito & Shimomura, 2015; Saito, 2015).本邦産ヤドリムシ類に関する論 文は約80報が刊行されている(このうちの32報は 椎野季雄博士による)(齋藤,2002).本邦における 等脚類全般の分類学的研究は,欧米に比べ発展途上 1 株式会社水土舎 〒214–0038 川崎市多摩区生田8–11–11
Suido-sha Co. Ltd., Ikuta 8–11–11, Tama-ku, Kawasaki, Kanagawa 214–0038, Japan
的な状況にあるが(布村,1984),エビヤドリムシ 類については椎野等の業績により,世界中で最も解 明された海域となっている(伊谷,2004).しかし ながら,これまでの調査地は千葉県以南の本州太平 洋岸,瀬戸内海,九州西岸に集中しており,北海 道, 東 北 地 方, 日 本 海 等 か ら の 報 告 は 少 な い (図3).今後調査が進むにつれ,さらに多くの知見 が得られるものと期待される(齋藤,2002).実際, 筆者自身も3種のエビヤドリムシ類(下記)を新種
記載している(Saito et al., 2010; Saito & Shimomura, 2015; Saito, 2015);
a) ショキタテナガノエラヤドリProbopyrus irio-motensis Saito, Shokita & Naruse, 2010,ショキ タテナガエビMacrobrachium shokitai Fujino &
Baba, 1973を宿主とする,純淡水産エビヤド リムシ類の本邦初記録.本種の研究の経緯に
ついては,本誌20号に詳しい(齋藤,2011).
b) サラサノハラヤドリAnisarthrus okunoi Saito & Shimomura, 2015, サ ラ サ エ ビRhynchocinetes uritai Kubo, 1942を宿主とする,サラサエビ科 エビ類寄生性エビヤドリムシ類の世界初記録. 雌の体型は著しく左右非対称であるが,胸部 両側に全7胸脚を備える特徴がみられた(Saito & Shimomura, 2015). c) アカホシカクレノコシヤドリIzuohshimaphryx-us hoshinoi Saito, 2015,アカホシカクレエビ Ancylomenes speciosus (Okuno, 2004)を 宿 主 と する,コエビ類腹部背面寄生性エビヤドリム 図1. エビヤドリムシ類に寄生されたコエビ類2種.A,エビジャコヤドリムシArgeia pugettensis Dana, 1852
に寄生されたクロザコエビArgis lar (Owen, 1839);B,サラサノハラヤドリAnisarthrus okunoi Saito & Shimomura, 2015に寄生されたサラサエビRhynchocinetes uritai Kubo, 1942.矢印はエビヤドリムシ類の寄 生部位を示す.(Aは齋藤ら(2001),BはSaito & Shimomura (2015)より出版元の許可を得て転載).
エビヤドリムシ科の研究 シ類の第3種.コエビノコシヤドリ属(新称) Filophryxus Bruce, 1972に類似するが属レベル での相違がみられたほか,宿主の腹部背面へ の 寄 生 状 況 に も 他 種 と の 違 い が み ら れ た (Saito, 2015). 2)宿主と寄生種の関係 国内のエビヤドリムシ類の宿主は106種の十脚甲 殻類が知られ,エビヤドリムシ類とほぼ同数が報告 さ れ て い た(齋 藤,2002; Shimomura et al., 2006, 2012; Saito et al., 2010; 三 浦,2014; 三 浦 ら,2014; Saito & Shimomura, 2015; Saito, 2015).これは宿主
とエビヤドリムシ類の関係が概ね1:1ということ を示唆している.エビノハラヤドリHemiarthrus ab-dominalis (Krøyer, 1840)は複数の十脚類を宿主とし て利用するが(齋藤・本尾,2010),エビヤドリム シ科としては宿主特異性を高める方向で進化してい ると考えられている(Markham, 1986).しかし,こ れには注意が必要である.エビヤドリムシ類の研究 はいまだ不十分であり,知見の不足がこの関係に反 映された可能性もある.このことを,伊谷博士は “見 か け の 宿 主 特 異 性” と 表 現 し て い る(伊 谷, 2004). 3)寄生率の時空間的把握 エビヤドリムシ類の寄生状況の地域差や季節差に ついて興味が持たれる.筆者は,日本海区水産研究 所が実施した「東海黄海等生物資源調査」によって 漁獲された5種のコエビ類に寄生するエビヤドリム シ類を観察することができた(齋藤ら,2001).調 査曳網ごとにエビへの寄生率を調べたが,なんと いっても,十分な標本数のデータを利用できたこと は魅力であった.ホッコクアカエビ(4100個体) の鰓腔寄生性アマエビエラヤドリは0~4.2%,ハサ
ミモエビEualus biunguis (Rathbun, 1902)(1100個体) とアシナガモエビLebbeus longipes (Kobjakova, 1936)
(554個体)の腹部に寄生するエビノハラヤドリは
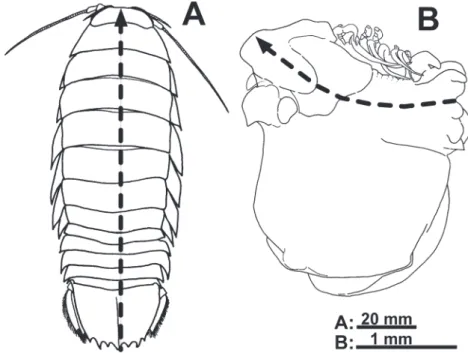
それぞれ5.2~64.7%および10.5~15.0%,クロザコ エビ(117個体)とトゲザコエビArgis toyamaensis (Yokoya, 1933)(29個体)の鰓腔に寄生するエビジャ 図2. 自由性等脚類(A, オオグソクムシBathynomus doederleini Ortmann, 1895)とエビヤドリムシ類(B, アカ
ホシカクレノコシヤドリIzuohshimaphryxus hoshinoi Saito, 2015)の体制の違い.破線矢印は体正中線(体 軸)を示す.自由性種は体型が左右相称で,流線型の美しいフォルムであるのに対し,エビヤドリムシ
類では体軸がゆがみ,育房が側方に大きく膨瘤し,左右不相称のユニークな形態である(A, 原図;B,
コヤドリムシはそれぞれ0.9%と27.6%であった. ま た, 東 京 湾 湾 奥 部 の 潟 湖 干 潟“新 浜 湖” で 2シーズン分の周年調査で採集されたニホンスナモ グリNihonotrypaea japonica (Ortmann, 1891)に寄生す るスナモグリヤドリムシIone cornuta Bate, 1864の寄 生状況の季節変化を調べることができた(齋藤・木 下,2004).本種は最近の分類学的再検討によりエ ビ ヤ ド リ ム シ 科 か ら 独 立 し て い る(Boyko et al., 2013)が,エビヤドリムシ類の寄生状況の参考にな るデータとして意義があると思われる.宿主は計 328個体が得られ,寄生がみられたのは22個体で あった(寄生率6.7%).各月の採集個体は0~31個 体 で, 被 寄 生 個 体 は0~3個体(寄生率0~25%), 明瞭な季節性はみられなかった.寄生は主に片側の 鰓腔にみられたが,両側の鰓に寄生するケースや, 片側の鰓に2個体が寄生するケースもみられた.ニ ホンスナモグリは第1胸脚の左右片方が巨大化して “大鉗脚”を形成するが,スナモグリヤドリムシの 寄生と大鉗脚の左右性に関係はみられなかった. 今後の展望 研究意義の最後の2項目は今後取り組んでいきた いテーマである. 4)宿主の個体への影響と個体群への影響 エビヤドリムシ類の寄生は宿主の雌雄による違い はないといわれている.スジエビモドキPalaemon serrifer (Stimpson, 1860)に寄生するエビヤドリムシ Bopyrus squillarum Latreille, 1802では,宿主の雌雄
エビヤドリムシ科の研究 間で寄生率に違いがみられなかったと報告されてい る(伊藤・渡邊,1992). しかし筆者の調査では宿主の性に対する選択性が みられた.東京湾湾奥の調査において,スナモグリ ヤドリムシは,主にニホンスナモグリの雌に寄生 し,雄への寄生は1例のみであった(齋藤・木下, 2004).寄生を受けた雌の宿主はいずれも未抱卵で, 寄生により不妊化したようであった.なお,本海域 の宿主の性比(雌雄に対する雄の割合)は19%で あり,寄生の雌雄差が宿主の性比に左右されている 可能性も考えられた. 一方,西表島でのショキタテナガノエラヤドリの 例では,宿主はいずれも雄であった(Saito et al., 2010).伊豆大島で採集された,アカホシカクレノ コシヤドリが寄生した2個体のアカホシカクレエビ も雄であった(Saito, 2015).なお,このうちの1個 体は第2腹肢内肢に雄性突起が見られず,寄生去勢 を受けた可能性が考えられた. エビヤドリムシ類の寄生は,個体レベルでは宿主 の 生 殖 機 能 を 阻 害 す る(Hiraiwa & Sato, 1939; O’Brien & Van Wyk, 1985).しかし,寄生虫の生存 には宿主の存在は不可欠であり,宿主個体群を滅ぼ すような害を与えるとは考えにくい.おそらくは, 宿主個体群へのエビヤドリムシ類の寄生率が,宿主 と寄生虫の共存バランスを保つキーとなっているも のと思われる.このことについて,さらに多くの事 例を集めて,今後検討していきたいと考えている. 5)宿主と寄生種の進化の関係 今後調査が進むにつれ,“真”の宿主特異性が明 らかになるものと思われる.宿主特異性が強く,限 られた宿主にのみ寄生するもの,逆にこれが弱くて 複数の十脚類を宿主に選択するもの,また,宿主の 多様性とエビヤドリムシ類の多様性に対応が見られ るケースもあるかもしれない.宿主の進化とエビヤ ドリムシ類の進化の関係は非常に興味の持たれる テーマであるが,Boyko & Williams (2009)に示され るように,その関係は単純なものではない.多角的 なアプローチによってその複雑な関係が解明されて いくことと期待する. 謝 辞 本稿は,日本甲殻類学会第53回大会自由集会「フ クロエビ類の生物学」(2015年10月9日開催)によ る講演内容をまとめたものである.本集会を企画, 運営された,日本甲殻類学会会長朝倉 彰博士(京 都大学瀬戸臨海実験所),ならびに集会のコンビ ナーを務められ,筆者に発表の機会を与えていただ いた,田中克彦博士(東海大学海洋学部),草稿を 御校閲いただきました岩下 誠博士(日本水産資源 保護協会)と伊谷 行博士(高知大学教育学部) に,記して謝意を表します. 文 献
Boyko, C. B., Moss, J., Williams, J. D., & Shields, J. D., 2013. A molecular phylogeny of Bopyroidea and Cryp-toniscoidea (Crustacea: Isopoda). Systematics and Bio-diversity, 11: 495–506.
Boyko, C. B., & Schotte, M., 2015. Bopyridae Rafinesque, 1815. In: C. B. Boyko, N. L. Bruce, K. L. Merrin, Y. Ota, G. C. B. Poore, S. Taiti, M. Schotte, G. D. F. Wil-son (eds.), World Marine, Freshwater and Terrestrial Isopod Crustaceans database. Accessed through: World Register of Marine Species at http://marinespecies.org/ aphia.php?p=taxdetails&id=1195 on 2015-10-23. Boyko, C. B., & Williams, J. D., 2009. Crustacean parasites
as phylogenetic indicators in decapod evolution. In: J. W. Martin,, K. A. Crandall, & D. L. Felder (eds.), De-capod Crustacean Phylogenetics. CRC Press, pp. 197– 220.
O’Brien, J., & Van Wyk, P., 1985. Effects of crustacean para-sitic castrators (epicaridean isopods and rhizocephalan barnacles) on growth of crustacean hosts. In: A. M. Wenner (ed.), Factors in Adult Growth, Crustacean Is-sues, 3, A. A. Balkema, Rotterdam, pp. 191–218. Hiraiwa, Y., & Sato, M., 1939. On the effect of parasitic
Isopoda on a prawn, Penaeopsis akayebi Rathbun, with a consideration of the effect of parasitization on the higher Crustacea in general. Journal of Science of the Hiroshima University, series B, div. 1 (Zoology), 7: 105–124. 伊谷 行,2004.エビの上で二人で暮らす:エビヤド リムシの生活史と種の多様性.フィールドの寄生 虫学:水族寄生虫学の最前線(長澤和也編).東 海大学出版会,神奈川,pp. 13–26. 伊藤 円・渡邊精一,1992.イソスジエビとスジエビ モドキにおけるBopyrus squillarumの寄生による影 響について.日本ベントス学会誌,43:29–39. Markham, J. C., 1986. Evolution and zoogeography of the
Isopoda Bopyridae, parasites of Crustacea Decapoda. In: R.H. Gore, & K.L. Heck (eds.), Crustacean Bioge-ography (=Crustacean Issues, 4). A. A. Balkema, Rot-terdam, pp. 143–164. 三浦知之,2014.鹿児島湾のニッポンウミシダ上に棲 息するウミシダヤドリエビに超寄生するエビヤド リムシ類の記録.Nature of Kagoshima, 40: 101–106. 三浦知之・宇都宮美樹・北嶋雄太・富岡 宏,2014. 海産甲殻類に寄生する等脚目エビヤドリムシ上科 に関する宮崎県での初めての記録(予報).宮崎 大学農学部研究報告,60: 29–39. 布村 昇,1984.海産等脚類研究の現状.日本ベント ス研究会誌,26: 38–42. 齋藤暢宏,2002.日本産ヤドリムシ(甲殻綱・等脚目) の宿主リスト.タクサ,13: 18–31. 齋藤暢宏,2011.寄生性甲殻類の1新種,ショキタテ ナガノエラヤドリについて:アマチュア研究者新 種記載顛末記.Cancer, 20: 39–41.
Saito, N., 2015. A new hemiarthrine bopyrid, Izuohshi-maphryxus hoshinoi (Crustacea: Isopoda) from Izu-Ohshima Island, Sagami Sea, Pacific coast of central Japan. Crustacean Research, 44: 67–73.
齋藤暢宏・伊谷 行・布村 昇,2000.日本産等脚目 甲殻類目録(予報).富山市科学文化センター研 報,23: 11–107. 齋藤暢宏・木下今日子,2004.東京湾奥部の潟湖干潟 に生息するニホンスナモグリから得られたエビヤ ドリムシ科等脚類,Ione cornutaの寄生状況.日本 ベントス学会誌,59: 1–7.
Saito, N., & Kubota, T., 1995. Species composition of cope-pods related to surface water masses in squid fishing grounds off Argentina. Proceedings of the NIPR Sym-posium on Polar Biology, 8: 139–153.
齋藤暢宏・本尾 洋,2010.クボイバラモエビLebbeus kuboi Hayashi, 1992から得られたエビヤドリムシ類 について.Cancer, 19: 15–17. 齋藤暢宏・養松郁子・橋詰和慶,2001.日本海産エビ 類にみられたエビヤドリムシについて.日本海区 水産試験研究連絡ニュース,394: 6–9.
Saito, N. & Shimomura, M., 2015. A new abdominally para-sitizing bopyrid, Anisarthrus okunoi sp. nov. (Crustacea: Isopoda), infesting the hinge-beak shrimp Rhynchocine-tes uritai Kubo, 1942 (Crustacea: Decapoda: Rhyncho-cinetidae). Species Diversity, 20: 37–43.
Saito, N., Shokita, S., & Naruse T., 2010. A new species of freshwater bopyrid, Probopyrus iriomotensis (cea: Isopoda), parasitizing Macrobrachium spp. Crusta-cea: Decapoda), from Iriomote Island, Ryukyu Islands, southwestern Japan. Species Diversity, 15: 169–183.
椎野季雄,1952.エビヤドリムシ科の系統発生につい
て.三重県立大学研究年報,第2部自然科学,1:
33–56.
下村通誉・布村 昇,2010.日本産等脚目甲殻類の分
類(1).海洋と生物,186: 78–82.
Shimomura, M., Ohtsuka, S., & Hashimoto, J., 2012. New distribution and host record of a penaeid ectoparasitic isopod, Minicopenaeon intermedium intermedium Bourdon, 1981 (Crustacea: Bopyridae). Bulletin of the Kitakyushu Museum of Natural History and Human History, Series A (Natural History), 10: 1–8.
Shimomura, M., Ohtsuka, S., & Sakakihara, T., 2006. Two bopyrid isopods infesting caridean shrimp Hippolyte sp. in the Seto Inland Sea, western Japan (Crustacea: Pera-carida). Bulletin of the Kitakyushu Museum of Natural History and Human History, Series A (Natural History), 4: 1–7.