1
渓流魚における遺伝的多様性低下の影響の把握
水産総合研究センター 増養殖研究所 栃木県水産試験場 三重大学 要旨 奇形個体が出現するイワナ集団および近隣の奇形個体が出現しない集団を対象に、マイクロサ テライト DNA 分析を実施し、奇形個体が出現する集団の遺伝的多様性を検討した。その結果、 奇形が出現する集団の遺伝的多様性は同一水系内の奇形個体が出現しない集団と比べ著しく低 いレベルであることが明らかになった。移植による遺伝的回復を図ったイワナ在来個体群につい て、適応度に関連する形質の変化を検証するため、発眼率の調査を行った。その結果、平均発眼 率は、2009 年の移植区間、移植区間外および 2010 年の移植区間よりもわずかに低い値を示した が、昨年度と同様に発眼率の有意な変化は認められなかった。遺伝的多様性の低下がイワナ個体 群に何を引き起こしているのかということを明らかにするため、堰堤で隔離された個体群の遺伝 的多様性と左右対称性のゆらぎ(FA:Fluctuating asymmetry)、及び遺伝的多様性と当歳魚のプロ ポーションの関係性について調査した。その結果、平均 Asymmetric score 及び FA 頻度と平均へ テロ接合度との間にいずれも有意な相関は認められず、プロポーションの平均因子座標と平均へ テロ接合度の間に有意な相関は認められなかった。文献調査の結果、次のことが考えられた。す なわち、本州の渓流魚在来個体群においては、キリクチや古座川流域のアマゴを除いて、現時点 で近交弱勢や遺伝的劣化によって個体群の存続可能性が危ぶまれている例は多くないかもしれ ない。しかしながら、隔離された小個体群という現状を考えると、近い将来にそのような状態に 陥ることは避けえないと考えられる。さらに、近交弱勢や遺伝的劣化が顕在化していない個体群 においても、遺伝的多様性の低下に伴って、適応可能性は不可避的に低下しつづける可能性が高 い。 緒言 サケ科魚類自然集団における奇形個体の出現は日本各地で報告がなされており(Morita & Yamamoto 2000; Sato 2006; Mori et al. 2009; 本報告書など)、遺伝多様性低下との関連性が指摘され ている(Morita et al. 2009)。しかしながら、奇形個体が出現する集団を対象に、実際に遺伝子分 析をおこなった研究はとても少ない。ここでは、奇形個体が出現するイワナ集団および近隣の奇 形個体が出現しない集団を対象に、マイクロサテライト DNA 分析を実施し、奇形個体が出現す る集団の遺伝的多様性を検討した。2
伝的回復(Genetic restoration)が様々な動植物種で行われ、一部で集団の遺伝的多様性や適応度 の増大に明らかな効果を上げている(Vrijenhoek et al. 1989;Westemeier et al. 1998)。移植による遺 伝的回復を図ったイワナ在来個体群について、適応度に関連する形質の変化を検証するため、発 眼率の調査を行った。また、遺伝的多様性の低下がイワナ個体群に何を引き起こしているのか、 ということを明らかにするため、堰堤で隔離された個体群の遺伝的多様性と左右対称性のゆらぎ (FA:Fluctuating asymmetry)、および遺伝的多様性と当歳魚のプロポーションの関係性について 調査した。 遺伝的多様性の低下は天然魚の存続可能性に短期的・長期的な負の影響を及ぼすと考えられる。 そのような負の影響が起こるメカニズムについて取りまとめることを目的とする。また、天然魚 の生息状況を把握するための文献調査を行うことも目的とした。 方法 奇形集団の遺伝子分析 福島県阿武隈川水系片貝川 A 集団、片貝川 B 集団、秋田県米代川水系大蓋沢川 C 集団、D 集 団から、遺伝子分析に供するサンプルを収集した。これらのうち奇形個体が出現する集団は、片 貝川 A 集団、米代川 C 集団であり、片貝川 A 集団ではすべての個体が胸鰭に奇形があり、米代 川 C 集団では 2011 年 11 月に担当者らが現地調査をおこなったところ、採集したイワナ 100 個体 中 2 個体が上顎欠損の奇形個体であった(図 1)。その他の集団からは今のところ奇形個体は確認 されていない。
使用したマイクロサテライト DNA 遺伝子座は、阿武隈川では 8 遺伝子座[Sfo-12 (Angers et al. 1995)、u-85 (Estoup et al. 1998)、Sle-1、Sle-5、Sle-6 (Yamaguchi et al. 2008)、Sco-200、Sco-211、Sco-220 (Dehaan and Ardren 2005)]、米代川では 16 遺伝子座 [(Sfo-8、Sfo-12、Sfo-103、Sfo-218、Sfo-290 (Angers et al. 1995)、u-85 (Estoup et al. 1998)、Ots-2 (Banks et al. 1999)、Fgt-3 (Sakamoto et al. 1994)、 Sle-1、Sle-4、Sle-5、Sle-6 (Yamaguchi et al. 2008)、Sco-200、Sco-211、Sco-216、Sco-220(Dehaan and Ardren 2005)]を用いた。
3 移植による遺伝的回復後の発眼率の変化 調査場所と支流間移植の実施 水系内で最も遺伝的多様性が低下した萱の手沢に対して、近隣 の支流である足沢からの支流間移植を 2009 年 9 月に実施した(栃木県水産試験場、2010)。萱の 手沢内の移植先区間を、堰堤間の距離が長く、最も多い生息個体数が見込まれる区間 1(流程長 276.6m、平均勾配 8.1%)と区間 2(流程長 223.7m、平均勾配 7.5%)とした(図 2)。移植に先立 ってエレクトリックショッカーによる採捕を行い、ほぼ雄だけが生息する区間(区間 1)とほぼ 雌だけが生息する区間(区間 2)を設定した(詳細は栃木県水産試験場、2011)。足沢で採捕した 成熟雄 16 個体と成熟雌 10 個体を、それぞれ区間 2 と区間 1 に移植した。 発眼率の確認 移植による適応度の変化を調べるための指標として、産着卵の発眼率を調査した。移植を実施 した 2009 年には、移植区間である区間 1 および 2 と移植区間の上下の 2 区間(区間 L と U)に おいて、ホースポンプを利用して産卵床から卵を回収し(岸ら 2009)、発眼卵数および死卵数を 計数した。平均発眼率は、観察された発眼卵数と死卵数の合計から産出した。移植翌年の 2010 年と翌々年の 2011 年には、 両移植区間の発眼率を同様 の方法で調べた。発眼率の 調査は、積算水温で産卵か ら 300℃・日を超えたと見込 まれた 2009 年 12 月 24 日、 2010 年 12 月 21 日および 2011 年 12 月 22 日に行った。 図 2 萱の手沢の移植先区間と移植の概要 図1 大芦川水系支流群の在来特定結果と遺伝的 多様性 青で示された 3 支流が在来個体群生息支流。A お よび He はそれぞれマイクロサテライト DNA8 遺伝 子座の分析から求められたアリル数と平均へテロ 接合度。
4
遺伝的多様性の低下と FA およびプロポーションの関係性
図 4 FA とプロポーションの調査対象河川
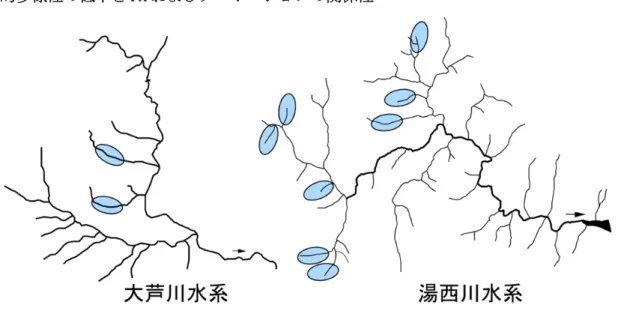
調査は利根川水系の 10 河川(大芦川支流 2、湯西川支流 8)において行った(図 4)。FA の計 数は当歳魚の胸鰭と腹鰭の鰭条数について行い(図 5)、FA の指標値として Asymmetric score (Iguchi et al. 2005)を次式により計算した。 Asymmetric score = 100 {|(PTL-R)| / (PTL+R) + |(PVL-R)| / (PVL+R)} また、集団中に含まれる FA が観察された個体の割合(FA 頻度)を求め、もう一つの指標値とし た。 プロポーションの測定は当歳魚を対象に行った。体側方向からデジタルカメラにより撮影し、 鰭の起点など 9 点のランドマークを設定した(図 6)。ランドマーク間の 17 の距離を Adobe Photoshop(Adobe)上で計測し、計量形質として扱った。全ての計量計質は、尾叉長で除すこと により標準化した。個体間で生じるプロポーションの違いが、体のどの部位に現れるかを検出す る目的で主成分分析を行った。 遺伝的多様性は上記解析を行った当歳魚と同所的に生息していた 1 歳以上の個体(1 河川につ き原則として 50 個体を解析した)を対象として解析した。 腹鰭鰭条数 PV 胸鰭鰭条数 PT L R L R 図 5 鰭条数の計数 鰭条数は顕微鏡下で 計数した
5 図 6 プロポーションの測定(上:ランドマーク、下:測定距離) A:吻端、B:背鰭起点、C:アブラ鰭起点、D:尾鰭起点 1、E:尾鰭叉点、 F:尾鰭起点 2、G:尻鰭起点、H:腹鰭起点、I:鰓蓋下端 文献調査 保全遺伝学に関する最新のレビュー論文を収集し、遺伝的多様性の低下が天然魚の存続可能性 に与える短期的・長期的影響について取りまとめた。また、分子生物学的手法によって、天然魚 集団の判別を行っている文献を収集して、(1)流域における天然魚の生息場所、(2)生息場所 のサイズ、(3)生息個体数、および(4)遺伝的多様性の観点から、生息状況を取りまとめた。 また、VORTEX (個体群存続可能性分析モデル; Lacy 1993) を用いて、平均的な個体数の在来個体 群が今後 100 年間に経験する可能性がある遺伝的多様性の減少を予測した。 結果及び考察 奇形集団の遺伝子分析 マイクロサテライト DNA を指標とした秋田県米代川水系の奇形集団および隣接する集団の遺伝 的多様性を表1に、福島県阿武隈川水系の多様性を表 2 に示した。米代川水系のイワナ集団、阿 武隈川の集団ともに、奇形が出現する集団の遺伝的多様性は同一水系内の奇形個体が出現しない 集団と比べ著しく低いレベルであることが明らかとされた(図 2)。同様の低い遺伝的多様性は、 熊野川水系の奇形(背びれ湾曲、あるいは尾鰭基部の欠損)イワナ集団にも報告されており(Sato et al. 2010)、イワナに見られるさまざまなタイプの奇形出現に遺伝的多様性の低下が関係する 可能性が考えられた。ただし、遺伝的多様性の低下が必ず奇形個体の出現につながるのかどうか は現時点で不明であり、今後多くの集団について奇形出現と遺伝的多様性との関係を検討してい く必要がある。 A I B H G C D F E A I B H G C D F E
6 遺 伝 子 座 S c o -2 2 0 S fo -1 2 S le -1 u-8 5 S co -200 S c o -21 1 S le -5 S le-6 F gt -3 O ts -2 S c o -216 S fo -103 S fo -2 9 0 S fo -2 18 S fo -8 S le-4 平 均 自 然 滝 下 流 集 団 H e 0. 57 0. 6 7 0. 3 8 0. 1 8 0. 40 0. 73 0 .66 0 .50 0. 73 0 .49 0. 8 0 0. 5 9 0. 4 8 0. 49 0. 72 0 .62 0. 5 6 H o 0. 55 0. 6 8 0. 4 8 0. 1 3 0. 29 0. 87 0 .68 0 .48 0. 71 0 .55 0. 8 4 0. 5 2 0. 4 2 0. 45 0. 70 0 .4 2 0 .5 5 P 0 .1 8 0 .0 6 0 .4 6 0 .2 3 0 .1 0 0 .1 0 0 .0 2 1 .0 0 0 .5 2 0 .7 1 0 .1 4 0 .5 4 0 .4 8 0 .4 7 0 .1 8 0 .00 N u m be r o f al le le s 343234 32 4 2 7 3 3 4 4 4 n 3 1 3 1 3 1 3 13 13 13 13 1 3 1 3 1 3 1 3 1 3 1 3 1 3 1 3 1 自 然 滝 上 流 集 団 H e 00000 0 .4 9 00 0 0 0 0 0 0 0 0 0 .0 3 (奇 形 集 団 ) H o 00000 0 .4 7 00 0 0 0 0 0 0 0 0 0 .0 3 P ----- 0 .3 7 - - - - - - - - - - N u m be r o f al le le s 000003 00 0 0 0 0 0 0 0 0 n 4 7 4 7 4 7 4 74 74 74 74 7 4 7 4 7 4 7 4 7 4 7 4 7 4 7 4 7 S fo -12 u-8 5 S co -220 S le -1 S co -211 S c o -20 0 S le -5 S le-6 平 均 自 然 滝 下 流 集 団 H e 0. 84 0. 6 4 0. 9 2 0. 8 5 0. 89 0. 79 0 .86 0 .57 0 .80 H o 0. 81 0. 3 5 0. 8 3 0. 8 5 0. 73 0. 42 0 .79 0 .54 0 .67 P 0 .0 9 0 .0 00 .0 10 .0 30 .0 00 .0 00 .0 40 .0 1 N u mb e r o f alle le s 9 7 1 9 1 3 2 0 1 0 9 5 n 4 8 4 8 4 8 4 84 84 84 84 8 自 然 滝 上 流 集 団 H e 0. 29 0. 1 5 0. 2 7 0. 3 8 0. 29 0. 63 0 .29 0 .10 0 .30 (奇 形 集 団 ) H o 0. 30 0. 2 5 0. 3 1 0. 3 9 0. 33 0. 61 0 .33 0 .18 0 .33 P 0 .0 6 0 .0 00 .0 00 .0 00 .0 10 .0 00 .0 10 .0 0 N u m be r o f al le le s 433445 43 n 4 8 4 8 4 8 4 84 84 84 84 8 表1 秋田県 米代川 水系にお け る奇形 イ ワナ 集団 と 近 接集団 の遺伝 的多様 性。 H e : ヘテ ロ 接合度 期待値 、H o : ヘ テ ロ 接 合 度 観察値 、 N u m be r o f all e le s: 対立 遺伝子 数、 P : ハ ー デ ィ ー ワ イ ンベルグ 平 衡検定 のP 値。 表2 福島県 阿武隈 川水系 に お け る 奇 形 イ ワ ナ 集 団と 近接集 団の遺 伝的多 様性。 H e : ヘ テ ロ 接合 度期待 値、 H o : ヘ テ ロ 接 合 度観察 値 、 N u m be r o f al le le s: 対立 遺伝子 数、 P : ハ ー デ ィ ー ワ イ ンベルグ 平 衡検定 のP 値。
8 発眼率 産卵床数と発眼率 2009 年には、移植区間で 6 カ所(区間 1:1 カ所、区間 2:6 カ所)、移植 区間外で 5 カ所(区間 U:2 カ所、区間 L:3 カ所)の産卵床が確認された。区間 1 で確認さ れた 1 カ所の産卵床では、わずかな死卵(19 粒) のみが観察されたことから、発眼率の算出から 除外した。区間 2 で確認された 6 カ所の産卵床 の平均発眼率は 69.4%だった(図 3)。いっぽう、 移植区間外で確認された合計 5 カ所の産卵床の 平均発眼率は 70.8%だった。移植翌年の 2010 年 には移植区間で合計 9 カ所(区間 1:5 カ所、 区間 2:4 カ所)の産卵床が確認された。その 平均発眼率は 73.6%だった。2011 年には移植区 間で合計 5 カ所(区間 1:3 カ所、区間 2:2 カ 所)の産卵床が確認され、その平均発眼率は 64.8%であった。2009 年の移植区間、移植区間 外および 2010 年と 2011 年の移植区間の発眼率に有意な差異は認められなかった(一元配置分散 分析、F = 1.09、P > 0.05)。 2011 年の平均発眼率は、2009 年の移植区間、移植区間外および 2010 年の移植区間よりもわず かに低い値を示した(64.8%)が、これまで同様に発眼率の有意な変化は認められなかった。こ れらの値は、鬼怒川支流蛇王沢の自然産卵床の 60.1%よりも高く、同沢の人工産卵場の 78.0%や 蒲田川に設置された人工産卵河川における 90.0%よりも低かった(中村 1999、中村ら 2009)。現 状では、過去に報告されている発眼率の範囲内にあったといえる。移植によって支流間の交配個 体が生まれ、それらが卵質や精子の質に関して現状よりも高い適応度を有する仮定とすると、そ の効果が現れるのは交配個体が繁殖に参加する 2012 年秋からになると考えられる。移植区間に おける発眼率やその他の形質の変化について、今後もモニタリングを継続する予定である。 FA と遺伝的多様性の関係性 データの得られた 9 河川(各河川 28~41 個体)の平均 Asymmetric score および FA 頻度につい て、平均へテロ接合度との関係性を Spearman の順位相関係数により検討したところ、いずれも 有意な相関は認められなかった(図 7、Asymmetric score、rs = -0.483、P > 0.05;FA 頻度、rs = -0.244、 P > 0.05)。 当歳魚プロポーションと遺伝的多様性の関係性 個体毎に得られた 17 の計測データについて主成分分析を行ったところ、第 1(PC1)から第 3 主 図 3 2009 年の移植区間外と移植区間および移植翌 年以降の移植区間における発眼率 バーは産卵床間の標準偏差、ボック スは範囲、黒四角は全産卵床から
9 成分(PC3)までで全分散のおよそ 63%が説明された(表 1)。PC1 は、CD、CG、DF、EF で因 子負荷量が高く、「尾柄部の発達の程度」を示す軸と考えられた。PC2 は、DG で因子負荷量が高 く、やはり「尾柄部の発達の程度」を示す軸と考えられた。PC3 は、AB で因子負荷量が高く、 「体躯前半部の発達の程度」を示す軸と考えられた。各集団の主成分得点を平均し、PC1 と PC2 を軸として図示したところ、明瞭な関係性は認められなかった(図 8)。特に遺伝的多様性が低下 していた 3 集団は、それぞれ離れた場所にプロットされており、遺伝的多様性の低下がある方向 へのプロポーションの変化(例えば、尾柄部の発達が悪い、など)をもたらすことはないと考え られる。また、各主成分(PC1~PC3)における集団毎の平均因子座標と平均へテロ接合度の関 係を検討したところ、いずれの主成分においても有意な相関は認められなかった(図 9、それぞ れ、r = -0.35、-0.20、-0.31、いずれも P > 0.05)。 図 7 Asymmetric score および FA 頻度と遺伝的多様性との関係性 各プロットは集団の平均値を示す。 表 1 5 つの主成分における 17 計量形質の因子負荷量と固有値
10 網掛けは第 1-第 3 主成分において高い因子負荷量を示した形質を示す。 図 8 PC1 および PC2 座標における各集団の平均主成分得点のプロット 色つきのプロットは特に遺伝的多様性が低い 3 集団 PC1 PC2 PC3 PC4 PC5 A-B 0.145 0.228 -0.724 -0.301 0.417 A-I -0.684 0.167 0.199 -0.289 0.381 B-C -0.39 -0.473 0.096 0.333 -0.437 B-G -0.597 -0.523 -0.112 -0.114 -0.411 B-H -0.665 -0.401 -0.231 -0.419 -0.143 B-I 0.608 -0.061 -0.675 0.069 -0.022 C-D 0.712 -0.339 0.372 -0.243 0.008 C-F 0.506 -0.322 0.349 -0.491 -0.057 C-G -0.703 -0.453 -0.248 -0.271 0.089 C-H -0.585 -0.511 -0.212 0.388 0.262 D-E -0.659 0.515 0.257 -0.041 -0.145 D-F -0.825 -0.124 -0.066 -0.342 0.06 D-G 0.308 -0.709 0.191 -0.073 0.334 E-F -0.761 0.319 0.24 0.014 -0.077 F-G 0.697 -0.346 0.384 0.043 0.186 G-H -0.344 -0.332 -0.089 0.55 0.284 H-I 0.671 -0.049 -0.491 -0.107 -0.444 Eigen.values 6.251 2.543 2.024 1.443 1.234 Proportion 36.773 14.96 11.905 8.488 7.259 Cumulative.prop. 36.773 51.733 63.638 72.126 79.384
11 図 9 各主成分(PC1~PC3)における集団毎の平均因子座標と平均へテロ接合度の関係 以上の結果から、調査対象河川のイワナ当歳魚(全て陸封型個体群)について、FA およびプ ロポーションと遺伝的多様性に、何らかの関係性を見いだすことはできなかった。北海道に生息 する降海型のイワナでは、ダムによる隔離に伴い遺伝的多様性が低下した集団で FA が増大する 有意な関係性が見いだされている(Morita et al. 2009)。両者の違いの原因は不明であるが、陸封 型と降海型の個体群では集団の創始に大きな違いがあり、ベースとなる遺伝的多様性も大きく異 なることから、単純な対比はするべきでないかもしれない。本州の陸封型イワナにおいては、遺 伝子分析によらずに遺伝的多様性の低下を検出する目的で、あるいは遺伝的多様性低下の影響を 検出する目的で FA やプロポーションの変化を指標として利用することはできないと考えられる。 遺伝的多様性の低下が天然魚の存続可能性に与える短期的・長期的影響及び在来個体群の生息現 状 遺伝的多様性の低下は、大きく 3 つのプロセスで天然魚の存続可能性に負の影響を及ぼすと考 えられる(図1)。すなわち、集団サイズが小さくなると、(1)近交弱勢:近親交配の頻度が高 まることで、本来は劣性な有害遺伝子がホモ化して発現し、適応度の低い個体の割合が集団内で 増加する;(2)遺伝的劣化:遺伝的浮動によって、複数の弱有害な遺伝子が集団内に広まり、 個体の平均的な適応度が低下する;(3)適応可能性の低下:遺伝的変異が失われることで、将 来の環境変動に対する適応可能性が低下する、である。1、2は天然魚の存続可能性を短期的に 低下させる可能性が高く、3は長期的な存続可能性を低下させると考えらえる。 分子遺伝学的手法を用いて在来個体群の判別を行っている論文は、イワナ(4 報)とアマゴ(1 報)であった。このうち、イワナの1 報は北海道の河川での研究例であり、本事業で想定してい る本州の渓流漁場とは生息条件が大きく異なるため、解析からは除外した。 渓流魚の魚種に関わらず、在来個体群は各流域の中で、最上流部に自然の滝や堰堤で隔離され た状態で生息していた(Kikko et al. 2008; Kubota et al. 2008; Sato et al. 2010)。在来個体群(キ
リクチを除くイワナ個体群)の平均的な生息場所サイズは、流程で1.2 km 程度であった(図2
12 来個体群の遺伝的多様性は非在来個体群と比較して非常に低く、ミトコンドリアDNA では多く の集団で単型的であり(図3)、多型マーカーであるマイクロサテライト DNA では、平均的に XX であった(図4)。特に、紀伊半島のキリクチ(イワナ)と古座川流域のアマゴ個体群では遺 伝的多様性が著しく低くかった(図3、4)。これらの在来個体群を除いたイワナ在来個体群の マイクロサテライトDNA の遺伝的多様性は、平均で 0.3 であり、非在来個体群の 0.5 程度と大 きな違いがなかった。しかしながら、キリクチと古座川のアマゴ以外の在来個体群においても、 その生息現状を考慮すると、遺伝的多様性は今後、急速に失われると予想される。実際に、平均 的な生息個体数(200 個体)と個体群パラメータをもとに、VORTEX を用いて遺伝的多様性の 長期的な変化を予測した結果、今後100 年の間に多くの在来個体群が遺伝的多様性を大きく失う と予想された(図5)。 保全遺伝学において指摘されているように、遺伝的多様性を保持することの意義は、すでに顕 在化している近交弱勢や遺伝的劣化を改善するだけでなく、集団の適応可能性を保持することに もある。本州の渓流魚在来個体群の生息現状を概観すると、紀伊半島のキリクチとアマゴ(古座 川流域)では、遺伝的多様性が極めて低い状態にあり、近交弱勢や遺伝的劣化の影響が危惧され た。実際に、いくつかのキリクチ個体群では、適応度指標の低い奇形個体が高頻度で確認されて いる(Sato 2006)。一方、これらの個体群を除くその他の在来個体群では、ミトコンドリア DNA が単型的であるものの、マイクロサテライトDNA の平均ヘテロ接合度は 0.3 程度であった。近 交弱勢や遺伝的劣化が顕在化する遺伝的多様性の閾値は分かっていないため、これらの在来個体 群が遺伝的問題により、短期的にも存続可能性を低下させているのかは現時点で判断できない。 しかしながら、多くの在来個体群が隔離された小さな個体群として存続していることから、これ らの個体群においても、比較的短期間のうちに、遺伝的多様性が大きく失われると予測されてい る(100 年間で、平均ヘテロ接合度が 0.3 程度低下)。 まとめると、本州の在来個体群においては、キリクチや古座川流域のアマゴを除いて、現時点 で近交弱勢や遺伝的劣化によって個体群の存続可能性が危ぶまれている例は多くないかもしれ ない。しかしながら、隔離された小個体群という現状を考えると、近い将来にそのような状態に 陥ることは避けえないと考えられる。さらに、近交弱勢や遺伝的劣化が顕在化していない個体群 においても、遺伝的多様性の低下に伴って、適応可能性は不可避的に低下しつづける可能性が高 い。これらのことを考えると、在来個体群の資源増大技術を開発する上で、人為的な隔離・分断 化とそれに伴う小集団化が進行する前の遺伝的集団構造を復元することは重要だろう。
13
図1.遺伝的多様性の低下に伴う存続可能性の低下に関する概念図
図2.在来個体群の生息場所サイズ(A)と推定生息個体数(B)
14
図3.在来個体群(天然)と非在来個体群(野生)のハプロタイプ多様度
図4.在来個体群(天然)と非在来個体群(野生)の平均ヘテロ接合度
15
図5.在来個体群(イワナ)の100年後の平均ヘテロ接合度
平均的な個体群パラメータをもつ個体群が、安定個体群として推移した場合の予測結果
引用文献
Angers B, Bernatchez L, Angers A, Desgroseillers L (1995) Specific microsatellite loci for brook charr reveal strong population subdivision on a microgeographic scale. J. Fish Biol. 47: 177-185.
Banks MA, Blouin MS, Baldwin BA, Rashbrook VK, Fitzgerald HA, Blankenship SM, Hedgecock D (1999) Isolation and inheritance of novel microsatellites in Chinook salmon (Oncorhynchus tshawytscha). J. Hered. 90: 281-288.
Dehaan PW, Ardren WR (2005) Characterization of 20 highly variable tetranucleotide microsatellite loci for bull trout (Salvelinus confluentus) and cross-amplification in other Salvelinus species. Mol. Ecol. Notes 5: 582-585.
Estoup A, Rousset F, Michalakis Y, Cornuet JM, Adriamanga M, Guyomard R (1998) Comparative analysis of microsatellite and allozyme markers: a case study investigating microgeographic differentiation in brown trout (Salmo trutta). Mol. Ecol. 7: 339-353.
Mori S. Akishinonomiya F. Sohkawa K. Hasegawa M. Matsumoto S. (2009) Red-spotted masu salmon (Oncorhynchus masou ishikawae) with unique jaw morphology collected from central Japan. Ichthyol. Res. 56: 208-209.
Morita K. Yamamoto S. (2000) Occurrence of a deformed white-spotted charr, Salvelinus leucomaenis (Pallas), population on the edge of its distribution. Fisheries Management and Ecology 7: 551-553.
16
Morita K. Morita SH. Yamamoto S. (2009) Effects of habitat fragmentation by damming on salmonid fishes: lessons from white-spotted charr in Japan. Ecol. Res. 24: 711-722.
Sakamoto T. Okamoto N. Ikeda Y. 1994. Dinucleotide repeat polymorphism of rainbow trout. J. Anim. Sci. 72: 2766-2767.
Sato T. (2006) Occurrence of deformed fish and their fitness-0related traits in Kirikuchi charr, Salvelinus leucomaenis japonicus, the southernmost population of the genuns Salvelinus. Zool. Sci. 23: 593-599. Sato T. (2010) Hybridization, isolation, and low genetic diversity of Kirikuchi char, the southernmost
populations of the genus Salvelinus. Trans. Amer. Fish. Soc. 139: 1758-1774.
Yamaguchi K. Nakajima M. Taniguchi N. 2008. Development of microsatellite markers in Japanese char Salvelinus leucomaenis and their applicability to closely related species. Fish. Genet. Breed. Sci. 38: 123-130.
岸大弼・苅谷哲治・徳原哲也. 2009. サケ科魚類の産卵床からのホースポンプによる発眼卵採集方 法. 水産技術 1: 25–28
Morita K, Morita HS, Yamamoto S. 2009. Effects of habitat fragmentation by damming on salmonid fishes: lessons from white-spotted charr in Japan. Ecol Res 24: 711–722.
中村智幸. 1999. 人工産卵場におけるイワナの産卵と産着卵のふ化. 日水誌 65: 434–440
中村智幸・徳田幸憲・高橋剛一郎. 2009. 人工産卵河川におけるイワナの産卵と当歳魚の動態. 応 用生態工学 12: 1–12
栃木県水産試験場. 2010. 平成 21 年度渓流資源増大技術開発事業報告書. 水産庁.
栃木県水産試験場. 2011. 平成 22 年度渓流資源増大技術開発事業報告書. 水産庁.
Vrijenhoek RC. 1994. Genetic diversity and fitness in small populations, pp. 37–53 in Conservation Genetics, edited by V. Loeschcke and S. Jain. Birkhauser, Basel.
Westemeier RL, Brawn JD, Simpson SA et al. 1998. Tracking the long-term decline and recovery of an isolated population. Science 282: 1695–1698.
Yamamoto S, Maekawa K, Tamate T et al. 2006. Genetic evaluation of translocation in artificially isolated populations of white-spotted charr (Salvelinus leucomaenis). Fish Res 78: 352–358.
Allendorf F.W., Hohenlohe P.A. & Luikart G. (2010). Genomics and the future of conservation genetics. Nature Reviews Genetics, 11, 697-709.
Fraser D.J., Weir L.K., Bernatchez L., Hansen M.M. & Taylor E.B. (2011). Extent and scale of local adaptation in salmonid fishes: review and meta-analysis. Heredity, 106, 404-420.
17
Hedrick P. (2005). 'Genetic restoration:'a more comprehensive perspective than'genetic rescue'. Trends in Ecology & Evolution, 20, 109.
Hedrick P.W. (1995). Gene flow and genetic restoration: the Florida panther as a case study. Conservation Biology, 9, 996-1007.
Hedrick P.W. & Fredrickson R. (2010). Genetic rescue guidelines with examples from Mexican wolves and Florida panthers. Conservation Genetics, 11, 615-626.
Ingvarsson P.K. (2001). Restoration of genetic variation lost-the genetic rescue hypothesis. Trends in Ecology & Evolution, 16, 62-63.
Kawamura K., Kubota M., Furukawa M. & Harada Y. (2007). The genetic structure of endangered indigenous populations of the amago salmon, Oncorhynchus masou ishikawae, in Japan. Conservation Genetics, 8, 1163-1176.
Kikko T., Kai Y., Kuwahara M. & Nakayama K. (2008). Genetic diversity and population structure of white-spotted charr, Salvelinus leucomaenis, in the Lake Biwa water system inferred from AFLP analysis. Ichthyological Research, 55, 141-147.
Kikko T., Kuwahara M., Iguchi K., Kurumi S., Yamamoto S., Kai Y. & Nakayama K. (2008).
Mitochondrial DNA population structure of the white-spotted charr (Salvelinus leucomaenis) in the Lake Biwa water system. Zoological Science, 25, 146-153.
Kubota H., Doi T., Yamamoto S. & Watanabe S. (2007). Genetic identification of native populations of fluvial white‐spotted charr Salvelinus leucomaenis in the upper Tone River drainage. Fisheries Science, 73, 270-284.
Lacy R.C. (1993). VORTEX: a computer simulation model for population viability analysis. Wildlife Research, 20, 45-65.
Narum S.R., Campbell N.R., Kozfkay C.C. & Meyer K.A. (2010). Adaptation of redband trout in desert and montane environments. Molecular Ecology, 19, 4622-4637.
Sato T. (2006). Occurrence of deformed fish and their fitness-related traits in Kirikuchi charr, Salvelinus leucomaenis japonicus, the southernmost population of the genus Salvelinus. Zoological Science, 23, 593-599.
Sato T., Demise T., Kubota H., Nagoshi M. & Watanabe K. (2010). Hybridization, Isolation, and Low Genetic Diversity of Kirikuchi Char, the Southernmost Populations of the Genus Salvelinus. Transactions of the American Fisheries Society, 139, 1758-1774.
Sato T. & Harada Y. (2008). Loss of genetic variation and effective population size of Kirikuchi charr: implications for the management of small, isolated salmonid populations. Animal Conservation, 11, 153-159.
18 rescue. Trends in Ecology & Evolution, 19, 489-496.
Taylor E.B., Tamkee P., Keeley E.R. & Parkinson E.A. (2011). Conservation prioritization in widespread species: the use of genetic and morphological data to assess population distinctiveness in rainbow trout (Oncorhynchus mykiss) from British Columbia, Canada. Evolutionary Applications, 4, 100-115. Weeks A.R., Sgro C.M., Young A.G., Frankham R., Mitchell N.J., Miller K.A., Byrne M., Coates D.J.,
Eldridge M.D.B., Sunnucks P., Breed M.F., James E.A. & Hoffmann A.A. (2011). Assessing the benefits and risks of translocations in changing environments: a genetic perspective. Evolutionary Applications, 4, 709-725.