J. Brew. Soc. Japan. Vol.101, No.12, p.949-956 (2006)
研 究 報 文酵母 の酢酸代 謝酵 素遺伝 子 の破 壊及 び高発 現 が アル コール発 酵 中の酢 酸生成 に及 ぼす影 響
後 藤(山 本)奈 美1・DANG HongA nh2
(1(独)酒 類 総 合 研 究 所 ・2FoodI ndustries Research Institute, Vietnam)
平 成18年5月24日 受 理
Effects of the gene disruption and the overexpression of acetate metabolic enzymes of yeast on acetate production during alcohol fermentation
Nami GOTO-YAMAMOTO1, DANG Hong Anh2
(1 National Research Institute of Brewing, 3-7-1 Kagamiyama, Higashi-Hiroshima 739-0046, Japan);
2Food Industries Research Institute , 301 Nguyen Trai Road, Thanh Xuan, Hanoi, Vietnam)
To study the mechanism of acetate production by yeast during alcohol fermentation, the effects of gene disruption of ALD2, ALD3, ALD6, and ACH1 as well as the overexpression of ACS1 and ACS2 on acetate production were examined using laboratory strains. Disruption of ACH1 and double disrup- tion of ALD2 and ALD3 did not affect the acetate production. Although the overexpression of ACS2 reduced the acetate production, it also inhibited alcohol fermentation. As was reported, the disruption of ALD6 reduced the acetate production. However, it caused an overproduction ofpyruvate, which was probably caused by a shortage of the reducing force.
Key words: 酵 母,酢 酸,ア ル デ ヒ ド デ ヒ ド ロ ゲ ナ ー ゼ ー
緒 言
酢 酸 は,清 酒,ワ イ ン な ど の 酒 類 に とっ て 望 ま し く な い 成 分 の 一 つ で あ り,発 酵 時 や 貯 蔵 中 に 酢 酸 菌 や 乳 酸 菌 に汚 染 さ れ る とそ の 濃 度 が 著 し く上 昇 す る 。 しか し,こ れ ら の 雑 菌 汚 染 が な い 場 合 で も,酢 酸 は 酵 母 に よ っ て 生 産 され,菌 株 や 発 酵 条 件 に よ っ て は 問 題 と な る程 度 に ま で 高 濃 度 に な る こ とが 知 ら れ て い る1>。
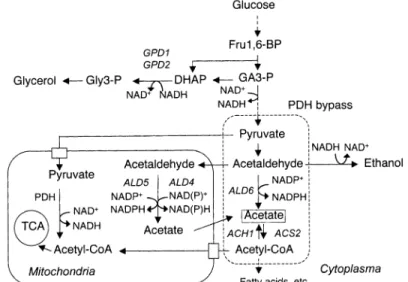
酢 酸 は,酵 母 細 胞 質 で ピ ル ビ ン 酸 か ら ア セ チ ル CoAを 生 合 成 す る 経 路(ピ ル ビ ン酸 デ ヒ ドロ ゲ ナ ー ゼ(PDH)バ イ パ ス)の 中 間 体 と し て 生 成 さ れ る と 考 え ら れ て き た(Fig.1)。PDHバ イ パ ス で は,ピ ル ビ ン酸 か ら生 成 した ア セ トア ル デ ヒ ドが ア ル デ ヒ ドデ ヒ ドロ ゲ ナ ー ゼ(ALD)に よ っ て 酸 化 さ れ て 酢 酸 と な り,さ ら に ア セ チ ルCoA合 成 酵 素(ACS)に よ っ
て ア セ チ ルCoAが 生 成 さ れ る 。PDHバ イ パ ス は 脂 肪 酸 生 合 成 等 の 原 料 と な る 細 胞 質 の ア セ チ ルCoAを 生 成 す る 重 要 な 経 路 で あ る が,何 ら か の 理 由 で 酢 酸 が 余 剰 に な っ た 場 合 は 酢 酸 が 菌 体 外,す な わ ち 酒 類 中 へ 排 出 さ れ る と考 え られ る。 実 験 室 酵 母 で はALD活 性 が 低 く,ACS活 性 が 高 い 株 の 酢 酸 生 成 が 低 い こ とが 報 告 さ れ て い る2)。最 近,Mizuno他3)は,酢 酸 生 成 が 低 く,ア ル コー ル 生 成 が 高 い ビー ル 酵 母 の 変 異 株 を 分 離 し,こ の 株 は ア ル コ ー ル デ ヒ ド ロ ゲ ナ ー ゼ 活 性 が 高 く,ALD活 性 が 低 い こ と を報 告 し た 。
ALDは,PDHバ イ パ ス に 関 与 す る 細 胞 質 型 と エ タ ノ ー ル の 資 化 に 関 与 す る ミ トコ ン ド リ ア 型 が5つ の 遺 伝 子,aLD2〜aLD6に コ ー ド さ れ て い る4)。 な お, ALD1は ク ロ ー ニ ン グ ・ア ー テ ィ フ ァ ク ト に 対 し て 命 名 さ れ た た め,欠 番 と な っ て い る。 こ の う ち,
949
後 藤(山 本)・DANG Hong Anh: 酵 母 の酢 酸 代 謝酵 素遺 伝 子 の 破壊 及 び高 発 現 が アル コール 発酵 中の酢 酸 生成 に及 ぼす 影響
Fig. 1 Schematic representation of the pyruvate dehydrogenase (PDH) bypass and related metabolism.
Fru 1, 6-BP, fructose 1, 6-bisphosphate; GA 3-P, glyceraldehyde 3-phosphate; DHAP, dihydrox- yacetone phosphat ; Gly 3-P, glycerol 3-phosphate.
AILD2, ALD3, 及 びALD6 が 細 胞 質 型 のALDを コ ー ド し て い る 。ALD6は 恒 常 的 に 発 現 し て お り, そ の 破 壊 株 は 生 育 が 遅 く な る と報 告 さ れ て い る5)が, Rernize他6)は シ ャ ン パ ン酵 母 か ら分 離 し た ハ プ ロ イ
ド株 を 用 い た 実 験 で,AL1)6破 壊 に よ り酢 酸 生 成 が 減 少 し,生 育 速 度 に は有 意 な 影 響 が 認 め られ な か っ た と報 告 し た 。 ま た,ALL)6の 高 発 現 で,酢 酸 生 成 が 約2倍 に な る こ と を 示 し た 。 し か し,a/d6△ 株 の 酢 酸 の 生 成 は ゼ ロ に は な らず,他 のALDも 関 与 し て い る だ ろ う と推 察 し て い る。
ALD3の 発 現 は 生 育 段 階 依 存 型 で,ス トレ ス で 誘 導 さ れ る こ と が 報 告 さ れ て い る 。 ま た,ALD2の 発 現 はALD3よ り も低 く,diauxic shiftと 高 浸 透 圧 条 件 で 誘 導 さ れ る4)。 ま た,Akamatsu他7)は,清 酒 膠 で は 膠 初 期 にALD2及 びALD3のmRNAが 蓄 積 す る こ と を報 告 した 。 高 糖 濃度 条 件 で は酢 酸 生 成 が 高 く な る こ と が 報 告 さ れ て お り1β),同 様 に,清 酒 醸 造 で は糖 化 が 進 み す ぎ て 糖 濃 度 が 高 くな っ た 場 合 に,ワ イ ン醸 造 で は 貴 腐 ワ イ ン の よ う に 果 汁 の 糖 濃 度 が 高 い 場 合 に 酢 酸 濃 度 が 高 くな る こ とが 経 験 的 に 知 ら れ て い る 。 従 っ て,高 浸 透 圧 条 件 でAILD2及 びALD3が 高 発 現 す る こ とが 酒 類 醸 造 中 の 酢 酸 生 成 に 関 与 す る 可 能 性 が
あ る と考 え,両 遺 伝 子 の 破 壊 株 の 酢 酸 生 成 を検 討 し た 。 酢 酸 か ら ア セ チ ルCoAを 生 合 成 す る 酵 素 が ア セ チ
ルCoAシ ン セ タ ー ゼ(ACS)で,好 気 条 件(非 発 酵 条 件)で 発 現 す るACS1と 発 酵 条 件 で 発 現 す る ACS2の2つ の 遺 伝 子 に コ ー ド さ れ て い る。Remize 他6)はACS2を 高 発 現 さ せ て も 酢 酸 生 成 が 減 少 し な い と報 告 し た 。 ま た,発 酵 条 件 で は な い が,deJong‑
Gubbels他9)はACS1及 びAC52を 高 発 現 さ せ て も, 呼 吸 条 件 で 増 殖 させ た 酵 母 に グ ル コ ー ス ・パ ル ス を与 え た 場 合 の 高 濃 度 の 酢 酸 生 成 を 抑 制 す る こ と は で き な か っ た と 報 告 し て い る 。 一 方,Akamatsu他7)は ACS2を 高 発 現 さ せ た 清 酒 酵 母 を 用 い る と液 体 静 置 培 養 お よ び 清 酒 小 仕 込 み で 酢 酸 生 成 量 が 減 少 す る と報 告 して お り,使 用 菌 株 に よ る差 異 が あ る もの と推 定 さ れ る 。
ACSと は 逆 に,ア セ チ ルCoAを 酢 酸 とCoAに 分 解 す る 酵 素 が ア セ チ ルCoAハ イ ドロ ラ ー ゼ でACHZ
に コ ー ド さ れ て い る。ACH1が 酢 酸 生 成 に 関 与 す る か ど うか は,こ れ ま で 報 告 さ れ て い な い。
本 研 究 で は,発 酵 条 件 に お け る 酵 母 の 酢 酸 生 成 機 作 を検 討 す るた め,酢 酸 を 生 成 す る 細 胞 質 型ALD及 び ACHZの 遺 伝 子 破 壊,並 び に 酢 酸 を 消 費 す るACS1 及 びACS2の 高 発 現 が,酵 母 の 酢 酸 生 成 に 及 ぼ す 影 響 を検 討 し た 。
950 醸 協 (2006)
後 藤(山 本)・DANGHongAnh:酵 母 の酢 酸 代謝 酵 素遺 伝 子 の破 壊 及 び高 発 現 が アル コール 発酵 中の 酢酸 生 成 に及 ぼす影 響
実 験 方 法 1. 使 用 菌 株
ALD2とALD3は 酵 母 のVIII番 染 色 体 に 並 ん で コ ー ド さ れ て い る た め,両 遺 伝 子 の 二 重 破 壊 株 は Navarro‑Avino他4)の 方 法 に ほ ぼ 準 じ て 作 成 し た 。 ま ずALD2の 上 流 配 列 とAL1)3の 下 流 配 列 を増 幅 し, pB luescriptllに 並 べ て ク ロ ー ニ ン グ し た 。 二 つ の 挿 入 配 列 の 間 に あ るHindIIIサ イ トに,pAG61プ ラ ス ミ ド 由 来 のCaURA3遺 伝 子 をSP6プ ラ イ マ ー 及 び T7プ ラ イ マ ー 配 列 にHindIIIサ イ トを つ け た プ ラ イ
マ ー で 増 幅 し(SP6プ ラ イ マ ー サ イ ト とCaURA3の 間 にHindIIIサ イ トが あ る た め,増 幅 断 片 は 両 端 に HlndIIIサ イ トを 持 つ),HindIII消 化 し て 挿 入 し た 。
ク ロ ー ニ ン グ し た(ALD2上 流 配 列‑CaURA3‑
ALD3下 流 配 列)のDNA断 片 をPCRで 増 幅 し,こ れ を 用 い て 半 数 体 酵 母,YPH499(MATa, ura3‑
52)を 形 質 転 換 し,ウ ラ シ ル 要 求 性 が 相 補 さ れ た 菌 株 を選 択 し た 。ALD2/ALL)3の 破 壊 は,ALD2上 流 配 列 の フ ォ ワ ー ドプ ラ イ マ ー とALL)3下 流 配 列 の リバ ー ス プ ラ イ マ ー を用 い たPCRで 確 認 した(datanot shown)。
ACH1破 壊 株 は,CaURA3遺 伝 子 を マ ー カ ー に 用 い た 相 同 組 み 換 え で 作 成 し た 。 ま ず,ACH1の 翻 訳 開 始 点 か ら550塩 基 と1135塩 基 の 間 を,clalサ イ ト 及 び5Pelサ イ トを つ け た プ ラ イ マ ー を 用 い て 増 幅 し た 。 次 に,増 幅 産 物 を両 制 限 酵 素 で 消 化 し,同 じ く両 制 限 酵 素 で 消 化 し た 後,フ ォ ス フ ァ タ ー ゼ 処 理 を し た pAG61プ ラ ス ミ ド に ラ イ ゲ ー シ3ン し た 。 得 ら れ た プ ラ ス ミ ド をACπ1フ ラ グ メ ン トの 中 程 に1カ 所 切 断 サ イ トが あ るNarIで 消 化 し て 線 状 に し,YPH 499 株 を形 質 転 換 し て ウ ラ シ ル 要 求 性 が 相 補 され た 菌 株 を 選 択 し た 。 形 質 転 換 株 は,ゲ ノ ムDNAをHind III 消 化 し,DIGラ ベ ル し たACH1フ ラ グ メ ン ト(開 始
コ ド ン か ら368塩 基 と1958塩 基 の 間)を プ ロ ー ブ に サ ザ ン解 析 を行 っ てACH1遺 伝 子 が 破 壊 さ れ て い る
こ と を確 認 した (data not shown)。
また, Saccharomyces Genome Deletion Project (http: //web. uni‑frankfurt. de/fb 15/ mikro/ euros‑
carf/ projects. htm1) で 作 成 さ れ た 半 数 体 及 び2倍 体 のACH1, ALD2, ALD3, 及 びALD6の 各 破 壊 株, 並 び に 親 株BY4741 (A4ATa, ATCC 201388),
BY4743 (MATa/al, ATCC201390)をATCCか ら購 入 し て使 用 し た 。
ACS遺 伝 子 の 過 剰 発 現 株 はAkamatsu他7)の 方 法 に従 い,オ ー レ オ バ シ ジ ン耐 性 遺 伝 子 とADH1プ ロ モ ー タ ー を 持 つ 多 コ ピ ー 型 プ ラ ス ミ ド,pAUR123(タ カ ラ バ イ オ)にACS1,ま た はACS2の コ ー ド領 域 を ク ロ ー ニ ン グ し,YPH499ま た はYPH499achヱ ム 株 を 形 質 転 換 し て 作 成 し た 。 コ ン ト ロ ー ル 株 に は pAUR 123を 形 質 転 換 し,培 地 に は オ ー レオ バ シ ジ ン Aを0.5g/1添 加 し た 。
2.発 酵 試 験
Wickerhamの 合 成 培 地(Yeast Nitrogen Base without Amino Acids, Difco)10)に 必 要 な ア ミ ノ 酸, 核 酸 を10mg/1補 い,断 らな い 限 りグ ル コ ー ス 濃 度 は 11%と し た 。 酵 母 は グ ル コ ー ス 濃 度5%の Wicker‑
hamの 合 成 培 地 で30℃,24時 間 振 と う培 養 培 養 後, 続 い て24時 間 静 置 培 養 し,遠 心 分 離 で 集 菌 ・洗 浄 後, 滅 菌 水 に 懸 濁 し,107cells/1に な る よ う に培 地 に 添 加 した 。 発 酵 試 験 は,シ リ コ 栓 を した 三 角 フ ラ ス コ を 用 い,25℃ で 静 置 し て 行 っ た 。
3。 分 析 方 法
酢 酸,そ の 他 の 有 機 酸 濃 度 の 定 量 は 島 津 製 作 所 製 モ デ ルLC‑10HPLCシ ス テ ム 及 びShim‑packSCR‑
102Hカ ラ ム2本 を用 い た 。 エ タ ノ ー ル 濃 度 及 び ピ ル ビ ン酸 濃 度 の 測 定 に はFキ ッ ト(ロ ッ シ ュ)を 用 い た 。
結 果
1.AL1)2,AL1)3,AL、06及 び.ACH1破 壊 が 酵 母 の 酢 酸 生 成 に 及 ぼ す 影 響
ald2△,ald3△,ald6△,ach1△ の 各 株,並 び に ald2△ald3△ 株 の 発 酵 速 度 は,対 照 株 と比 較 し て 大 き な 差 異 は 認 め ら れ な か っ た(Fig.2)。 酢 酸 の 生 成 は,Remize他6)の 報 告 と 同 様,ald6△ 株 で は 減 少 し た が,ald2△,ald3△,ach1△ の 各 株(半 数 体 及 び2 倍 体),並 び にald2△ald3△ 株 で は そ れ ぞ れ の 対 照 株
と ほ ぼ 同 様 で あ っ た(Fig.3)。 し か し,ald6△ 株 で は 発 酵 液 の ピル ビ ン 酸 濃 度 が 対 照 株 お よ び 他 の 供 試 株 よ り高 く な っ た(Table1)。 そ の 他 の 有 機 酸 濃 度 に は 有 意 な 差 異 が 認 め られ な か っ た(datanotshown>。
2.ald2△ald3△ 株 とald6△ 株 の 酢 酸 生 成 に 及 ぼ す培 地 浸 透 圧 の 影 響
ALD2及 びALD3が 高 浸 透 圧 条 件 で 高 発 現 す る 現 951
後 藤(山 本)・DANGHongAnh:酵 母 の酢 酸代 謝 酵 素遺 伝 子 の破 壊 及 び高 発現 が アル コー ル発 酵 中 の酢 酸 生成 に及 ぼす影 響
Fig. 2 Changes in ethanol concentrations during a static liquid culture of YPH 499, BY 4741, BY 4743, and
their ALDs and ACH1 disruptants. Wickerham's minimal media containing 11% glucose was
fermented at 25•Ž by ald2 A (closed triangle), ald3 A (open triangle), ald6 A (closed square), ald2 A
ald3 A (open square), ach1A (closed circle), and a control strain (open circle).
Fig. 3 Changes in acetate concentrations during a static liquid culture of YPH 499, BY 4741, BY 4743, and their ALDs and ACH1 disruptants. Fermentation condition and symbols are the same as for Fig. 2.
象 は,高 浸 透 圧 条 件 で の 酢 酸 高 生 産 と何 ら か の 関 連 が あ る の で は な い か と 推 定 さ れ た が,ald2△ald3△ 株 の11%グ ル コ ー ス 培 地 に お け る 酢 酸 生 成 は 対 照 株 と 同 様 で あ っ た 。 そ こで,ald2△ald3△ 株 とald6△ 株 を 用 い て 異 な る 浸 透 圧 条 件 で 発 酵 試 験 を行 っ た 。 そ の 結 果(Fig.4),各 株 と も2%グ ル コ ー ス 培 地 と比 較 し て,2%グ ル コ ー ス+0.5Mソ ル ビ トー ル 培 地 及 び11
%グ ル コ ー ス 培 地(両 培 地 は初 発 の 浸 透 圧 が 同 じ)で
は発 酵 終 了 後 の 酢 酸 濃 度 が 高 くな っ た 。2%グ ル コ ー ス 培 地 と,2%グ ル コ ー ス+0.5Mソ ル ビ トー ル 培 地 の 酢 酸 濃 度 の 差 は,主 に浸 透 圧 に よ っ て 生 じ た も の と 考 え られ る。 し か し,ど の 培 地 で もald2△ald3△ 株 の 酢 酸 生 成 は,対 照 株 と差 異 が 認 め られ な か っ た 。 一 方,ald6△ 株 で は,ど の 条 件 で も親 株 よ り酢 酸 生 成 が 低 下 し て い る こ とが 確 認 さ れ た 。
952 醸 協 (2006)
後藤(山 本)・DANGHongAnh:酵 母 の酢 酸代 謝 酵 素遺 伝 子 の破 壊 及 び高 発現 が ア ル コー ル発 酵 中 の酢 酸 生成 に及 ぼ す影 響
Table 1 Acetate and pyruvate concentrations in fermented media
Fig. 4 Acetate concentrations after static liquid
cultures of YPH 499, BY 4741, and their
ALD2 ALD3 and ALD6 disruptants. Wick-
erham's minimal medium containing 2%
glucose (closed bar), 2% glucose and 0.5 M
sorbitol (hatched bar), and 11% glucose
(open bar) was fermented at 25•Ž.
3.ACSI, ACS2高 発 現 及 びACH1破 壊 が 酢 酸 生 成 に 及 ぼ す 影 響
ACSI, ACS2とACH1と は 逆 反 応 を 触 媒 す る 酵 素 を コ ー ド し て い る 。 発 酵 条 件 で 発 現 す るaCS2を
高 発 現 させ た 場 合 に,酢 酸 生 成 が 減 少 した と い う報 告 と変 化 しな か っ た と い う報 告 が あ る が,ACH活 性 が 強 い 場 合 に はACS2高 発 現 の 酢 酸 生 成 に 対 す る 効 果 が 打 ち 消 さ れ る の で は な い か,と 考 え,achl△ 株 及 び 野 生 型 株 で のACS1及 びACS2高 発 現 が 酢 酸 生 成 に 及 ぼ す 影 響 を検 討 し た 。
そ の 結 果(Fig.5),ACS2の 高 発 現 で 酢 酸 生 成 は 減 少 した が,ア ル コ ー ル 発 酵 も同 様 に 抑 制 さ れ,特 に ach1△ 株 で 顕 著 で あ っ た 。 一 方,ACS1の 高 発 現 で も若 干 ア ル コ ー ル 発 酵 及 び 酢 酸 生 成 の 抑 制 が 認 め ら れ た が,4CS2ほ ど顕 著 で は な か っ た 。
清 酒 酵 母 で はACS2の 高 発 現 に よ る 発 酵 阻 害 は認 め られ な か っ た η。 シ ャ ンパ ン酵 母 由 来 の 半 数 体 株 で は,AGS2の 高 発 現 に よ っ て 増 殖 が 抑 制 さ れ,初 期 の 発 酵 速 度 も対 照 よ り遅 くな っ た が,後 半 の 発 酵 速 度 は対 照 よ り も速 く,最 終 的 に 発 酵 が 早 く終 了 した と報 告 さ れ て い る6)。今 回 の 結 果 は,清 酒 酵 母,シ ャ ン パ ン 酵 母 の い ず れ と も 異 な っ て お り,こ れ は遺 伝 的 な バ ッ ク グ ラ ウ ン ドの 違 い に 起 因 す る と考 え られ る 。 少 な く と も今 回 用 い た 実 験 室 酵 母YPH499で は,ACS2 高 発 現 及 びACH1破 壊 に よ る 過 剰 な ア セ チ ルCoAの 生 成 に よ っ て,代 謝 系 に 障 害 を 生 じ て い る の で は な い
か と推 察 さ れ る 。
考 察
1.酵 母 の 酢 酸 生 成 とALD遺 伝 子
本 実 験 の 結 果 か ら,ALD2及 びALD3が 高 浸 透 圧 条 件 で 高 発 現 す る 現 象 は,酢 酸 の 高 生 産 との 関 連 は な い と考 え られ た 。
酢 酸 の 合 成 に 関 し て は,細 胞 質 のPDHバ イ パ ス に さ ら に ミ トコ ン ド リア のALDが 関 与 す る バ イ パ ス が 存 在 す る こ とが 報 告 さ れ た11)。す な わ ち,細 胞 質 の ア セ トア ル デ ヒ ドが ミ トコ ン ド リ ア 内 のALDで 酢 酸 に 酸 化 さ れ,こ れ が 再 度 細 胞 質 に戻 っ て ア セ チ ルCoA の 基 質 に な る,と い う 経 路 で あ る(Fig.1)。Saint‑
Prix他12)は,各ALD遺 伝 子 破 壊 株 の 酢 酸 生 成 能 を 調 べ,ald6△ 株 だ け で な く,ミ トコ ン ド リ ア のALD
を コ ー ドす るALD5の 破 壊 株 も酢 酸 生 成 が 低 下 す る 953
後藤(山 本)・DANGHongAnh:酵 母 の酢 酸代 謝 酵 素遺 伝 子 の破 壊 及 び高 発 現 が アル コ ール 発酵 中 の酢 酸 生成 に及 ぼす影 響
Fig. 5 Changes in ethanol and acetate concentrations during a static liquid culture of YPH 499 (solid line) and its ACH1 disruptant (broken line) with or without an overexpression of ACS1 and ACS2. Open circle, control; closed triangle, overexpression of ACS]; closed circle, overexpression of ACS2.
こ と を 明 ら か に し た 。 ま た,a♂46△ 株 で は ミ ト コ ン ド リ ア のALDを コ ー ド す るALD4が 高 発 現 し て お り,a146△a144△ 株 で は 増 殖 が 抑 制 さ れ,a146△
aJ44△a145△ 株 で は さ ら に 著 し い 増 殖 抑 制 が 認 め ら れ た 。 こ の グ ル ー プ はa1謡2△ald3△ 株 の 検 討 を行 っ て い な い が,単 独 の 破 壊 株 で は酢 酸 生 成 に 変 化 が 認 め ら れ な い こ と,並 び にa146△a144△a1諾2△a143△ 株 の酢 酸 生 成 がa/46△aZ44△ 株 と変 わ ら な い こ と か ら, ALD2,ALD3両 遺 伝 子 の 酢 酸 生 成 へ の 関 与 は ほ と ん ど な い と推 定 し て お り,本 実 験 は,こ の 結 論 を 裏 付 け る も の と な っ た 。 従 っ て,酢 酸 は 主 にALD6が 関 与 す る 細 胞 質 のPDHバ イ パ ス,並 び にALD4, AしL)5に よ る ミ トコ ン ド リ ア の 経 路 の 両 者 に よ っ て
生 成 す る と考 え ら れ る 。 実 際 の 清 酒 膠 か ら採 取 し た 酵 母 か ら も,/1LD6と と も にALD4のmRNAが 検 出 さ れ た(ALO5は 未 検 討)7)。 し か し,興 味 深 い こ と に全 て のALDを 破 壊 した 株 で も,エ ル ゴ ス テ ロ ー ル, オ レ イ ン酸 及 びTween80を 添 加 した 培 地 で 増 殖 ・発 酵 が 可 能 で,少 量 で は あ る が 酢 酸 が 生 成 した こ とか ら, さ ら に 異 な る 酢 酸 の 生 成 系 が あ る だ ろ う,と 報 告 さ れ て い る12)。
さ ら に,ALD2及 びALD3に つ い て は,パ ン トテ ン酸 生 合 成 系 の3一 ア ミノ プ ロ パ ン ア ル デ ヒ ドか ら β‑
ア ラ ニ ン(ひ ア ミノ プ ロ パ ン 酸)を 生 成 す る 反 応 を触 媒 す る こ とが 報 告 さ れ だ3>。 本 実 験 で 用 い たWicker‑
hamの 合 成 培 地 に は パ ン トテ ン酸 が 添 加 さ れ て い る こ と か ら,a162△a143△ 株 の パ ン トテ ン酸 要 求 性 が 現 れ な か っ た もの と考 え られ る。
2.酵 母 の 酢 酸 生 成 と酸 化 還 元 バ ラ ン ス
高 浸 透 圧 条 件 で は,酢 酸 生 成 が 上 昇 す る と と も に, 菌 体 内 の 浸 透 圧 調 製 の た め グ リセ ロ ー ル が 高 生 産 され
る こ とが 知 られ て い る。 グ リ セ ロ ー ル 合 成 系 の グ リセ ロ ー ル ーひ リ ン酸 デ ヒ ド ロ ゲ ナ ー ゼ はG翔 刀,GPO2 の2遺 伝 子 に コ ー ド さ れ て お り,こ の う ち,GP加 を 高 発 現 さ せ た と こ ろ,グ リセ ロ ー ル と と もに 酢 酸 や コ ハ ク 酸 な どが 高 生 産 さ れ た と報 告 され て い る14)。ま た, 別 の グ ル ー プ も,GLIP2を 高 発 現 さ せ た 場 合 に 同 様 の 結 果 と な っ た が,君LO6を 破 壊 す る こ とで 酢 酸 の 高 生 産 を抑 制 で き た と報 告 し だS)。 グ リセ ロ ー ル 生 産 に は浸 透 圧 調 節 の 他,過 剰 なNADHを 消 費 し て 酸 化 還 元 バ ラ ン ス を と る意 義 が あ る こ とが 知 ら れ て お り絡), NAD(P)Hを 生 産 す る 酢 酸 生 成 と は酸 化 還 元 の 観 点 か ら は反 対 の 作 用 が あ る 。 従 っ て,高 浸 透 圧 で グ リセ ロ ー ル が 高 生 産 さ れ た 場 合 は,消 費 さ れ たNADHを 補 う た め に酢 酸 が 過 剰 に 生 産 さ れ た と推 定 され る。
な お,ミ トコ ン ド リア 内 のNADHは ミ ト コ ン ド リ ア 内 膜 を透 過 で き な い た め,直 接 細 胞 質 に は輸 送 され な い が,最 近,酵 母 に も動 物 細 胞 で 知 ら れ て い る リ ン ゴ酸‑ア ス パ ラ ギ ン酸 シ ャ トル(リ ン ゴ酸,ア ス パ ラ ギ ン酸 が ミ ト コ ン ド リ ア 内 膜 を透 過 す る こ とで,間 接
954 醸 協 (2006)
後藤(山 本)・DANG Hong Anh:酵 母 の酢 酸代 謝 酵素 遺 伝 子 の破 壊及 び高発 現 が ア ル コー ル発 酵 中 の酢 酸 生成 に及 ぼす 影 響
的 にNADHを 輸 送 す る 系)が 酵 母 に も存 在 す る 可 能 性 が 報 告 さ れ た17)。従 っ て,こ の 系 を介 し て ミ ト コ ン ド リア 内 のNADHが 細 胞 質 へ 輸 送 さ れ る と推 定 さ れ る 。
ま た,細 胞 質 型 のALD6を 欠 損 さ せ る こ と で,高 浸 透 圧 条 件 で の 酢 酸 生 成 が 減 少 し た(Fig。4)こ と か ら,ALL)6も グ リセ ロ ー ル 高 生 産 時 の 酢 酸 生 成 に 寄 与 し て い る と 考 え ら れ る 。Ald6PはNADPを 補 酵 素 とす る が,酵 母 に はtransdehydrogenase活 性 が な く,NADPHとNADHを 直 接 変 換 す る こ と は で き な い 。 ま た,NADHか らNADPHへ の 変 換 は,グ リ セ ロ ー ル の 生 産 と分 解 を行 う こ とで 可 能 で あ る と考 え られ て い る が18),そ の 逆 の 系 は現 在 の と こ ろ報 告 さ れ て い な い 。 お そ ら く,NADH,NADPHの 両 者 を 補 酵 素 に す る 酵 素 な どが 両 者 の バ ラ ン ス を と る た め に 関 与 し て い る の で は な い か と推 定 さ れ る が,こ の 点 は さ ら に検 討 が 必 要 で あ る 。
さ ら に,GOD1高 発 現 株 で は,ア セ トア ル デ ヒ ドや ピ ル ビ ン 酸 が 発 酵 途 中 に 高 蓄 積 さ れ,こ れ はNADH の 不 足 に よ っ て エ タ ノ ー ル 発 酵 が 阻 害 さ れ た た め と推 定 さ れ て い る19)。今 回 の 実 験 で も,ald6△ 株 で は 発 酵 液 の ピル ビ ン 酸 濃 度 が 上 昇 し て お り,こ れ も還 元 力 の 不 足 が 原 因 と 考 え ら れ る 。Remize他6)は,aJ46△
株 で ア セ トア ル デ ヒ ド濃 度 が 一 時 的 に上 昇 し,発 酵 終 了 後 に は ア セ トア ル デ ヒ ドか ら生 成 し た と考 え ら れ る 2,ひ ブ タ ン ジ オ ー ル 濃 度 が 高 くな っ た と報 告 した 。
以 上 の こ と か ら,酵 母 の 酢 酸 生 成 に は,代 謝 系 の 酵 素 活 性 だ け で な く,NAD+/NADH等 の 酸 化 還 元 バ ラ ン ス が 強 く影 響 し て い る こ とが 推 察 さ れ た 。
要 約
実 験 室 酵 母 を 用 い た 実 験 で,/1LD2,/1LD3,及 び ACH1の 遺 伝 子 破 壊 は,発 酵 条 件 で の 酢 酸 生 成 に 影 響 し な い こ と が 示 さ れ た 。 ま た,ノ1CS2の 高 発 現 は 酢 酸 生 成 を 抑 制 した が,ア ル コ ー ル 発 酵 も 阻 害 し た 。 既 報 の とお り,ALD6の 破 壊 は 酢 酸 生 成 を減 少 さ せ た が,ピ ル ビ ン酸 を 高 生 産 して お り,こ れ は 還 元 力 の 不 足 が 原 因 と考 え ら れ た 。 以 上 の こ と か ら,酢 酸 の 生 成 に は 代 謝 系 の 酵 素 活 性 の 他,酸 化 還 元 バ ラ ン ス の 維 持 が 強 く関 わ っ て い る こ とが 推 察 さ れ た 。
文 献
1) Y. SHIMAZU and M. WATANABE: J. Ferment.
Technol., 59, 27-32 (1981)
2) C. VERDUYN, E. POSTMA, W. A. SCHEFFERS, and J. P. VAN DIJKEN: J. Gen. Microbiol., 136, 395-403 (1990)
3) A. MIZUNO, H. TABEI, and M. IWAHUTI: J.
Biosci. Bioeng., 101, 31-37 (2006)
4) J. P. NAVARRO -AvISTo, R. PRASAD, V. J.
MIRALLES, R. M. BENITO, and R. SERRANO:
Yeast, 15, 829-842 (1999)
5) P. G. MEADEN, F. M. DICKINSON, A. MIFSUD, W. TESSIER, J. WESTWATER, H. BUSSEY, and M. MIDGLEY: Yeast, 13, 1319-1327 (1997) 6) F. REMIZE, E. ANDRIEU, and S. DEQUIN: Appl.
Environ. Microbiol., 66, 3151-3159 (2000) 7) S. AKAMATSU, H. KAMIYA, N. YAMASHITA, T.
MOTOYOSHI, N. GOTO -YAMAMOTO, T. ISHIK- AWA, N. OKAZAKI, and A. NISHIMURA: J.
Biosci. Bioeng., 90, 555-560 (2000)
8) 上 田 隆 蔵, 西 村 公 臣: 改 訂 清 酒 酵 母 の研 究, p.571‑584. 清 酒 酵 母 研 究 会 (1980)
9) P. DE JONG-GUBBELS, M. A. VAN DEN BERG, M.
A. H. LUTTIK, H. Y. STEENSMA, J. P. VAN DIJKEN, and J. T. PRONK: FEMS Microbiol.
Lett., 165, 15-20 (1998)
10) M. J. ZIMBRO, and D. A. POWER (eds.): Difco
& BBL Manual, p. 638-642, Becton, Dickin- son and Company, Sparks (2003)
11) S. BOUBEKEUR, 0. BUNOUST, N. CAMOUGRAND, M. CATROVIEJO, M. RIGOULET, and B. GUERIN:
J. Biol. Chem., 274, 21044-21048 (1999) 12) F. SAINT-PRIX, L. BONQUIST, and S. DEQUIN:
Microbiology, 150, 2209-2220 (2004)
13) W. H. WHITE, P. L. SKATRUD, Z, XUE, and J.
H. TOYN: Genetics, 163, 67-77 (2003)
14) F. REMIZE, J. L. ROUSTAN, J. M. SABLAYR- OLLES, P. BARRE, and S. DEQUIN: Appl. Envi-
ron. Microbiol., 65, 143-149 (1999)
15) J. M. EGLINTON, A. J. HEINRICH, A. P. POLL- NITZ, P. LANGRIDGE, P. A. HENSCHKE, and M.
DE BARROS LOPES: Yeast, 19, 295-301 (2002) 16) C. GANCEDO and R. SERRANO: The Yeasts, 2
nd ed. vol. 3, ed. by A. H. ROSE and J. S.
HARRISON, p. 205-259, Academic Press, Lon- 955
後 藤(山 本)・DANGHongAnh:酵 母 の酢 酸代 謝 酵 素遺 伝 子 の破 壊 及 び高 発現 が ア ル コー ル発 酵 中 の酢 酸生 成 に及 ぼ す影 響
don (1989)
17) S. CAVERO, A. Vozza, A. DEL ARCO, L. PAL- MIERI, A. VILLA, E. BLANCO, M. J. RUNSWICK, J. E. WALKER, S. CERDAN, F. PALMIERI, and J.
SATROSTEGUI: Mol. Microbiol., 50, 1257-1269 (2003)
18) J. NORBECK, and A. BLOMBERG: J. Biol.
Chem., 272, 5544-5554 (1997)
19) 5. MICHNICK, J-L. ROUSTAN, F. REMIZE, P.
BARRE, and S. DEQUIN: Yeast, 13, 783-793 (1997)
956 醸 協 (2006)