脂肪族ポリエステルの環境分解制御
目次
第1章 序論 3
はじめに 4
生分解性高分子 4
ポリラクトン 6
ポリ(n-アルキレン n-アルキレンジカルボキシレート) (PAAD) 7
生分解性高分子の課題 9
生分解性制御方法 9
本研究の目的 11
第2章 ポリ(ブチレン n-アルキレンジカルボキシレート)の合成と生分解性評価 13
2-1 緒言 14
2-2 実験 16
試薬 16
核磁気共鳴スペクトル(NMR) 16
分子量測定 16
熱的物性測定 16
フィルム作成 17
広角X線回折(WAXD) 17
ポリ(ブチレン n-アルキレンジカルボキシレート)(PBAD)の合成 18 生物化学的酸素要求量(BOD)生分解度測定 18 PBAD分解微生物の計数および単離 19
16SrDNA配列解析 21
PBAD分解細菌のクリアゾーン形成能評価 22
2-3 結果 23
PBADの合成 23
PBADの熱的性質 24
BOD生分解度評価 28
クリアゾーン形成試験 29
2-4 考察 33
2-5 まとめ 35
第3章 ジスルフィド結合の還元的開裂反応を利用した生分解開始時期の制御 36
3-1 緒言 37
3-2 実験 41
試薬 41
核磁気共鳴スペクトル(NMR) 41
フーリエ変換赤外吸収スペクトル測定 41
分子量測定 41
熱的物性測定 42
PBDTsのプレスフィルム作成 42
広角X線回折 (WAXD) 42
酸化還元電位(ORP)測定 42
ポリデカンジチオール(PDDT)の合成 43 ジチオジグリコール酸(DTG)の合成 43
1,4-ブチレン ビズ(チオグリコレート)(BTG)の合成 43
ポリブチレンジチオジグリコレート(PBDTG)の合成 44 ジチオジプロピオン酸(DTP)の合成 44
1,4-ブチレン ビズ(チオグリコレート)(BTP)の合成 45
ポリブチレンジチオジプロピオネート(PBDTP)の合成 45
PBDTsの加水分解性および還元分解性の評価 46
生物化学的酸素要求量 (BOD) 生分解度測定 46
3-3 結果 48
PDDTの合成 48
PBDTGおよびPBDTPの合成 49
PDDT,PBDTGおよびPBDTPの熱的性質 54
PDDT,PBDTGおよびPBDTPの加水分解性および還元分解性の評価 58
第1章
序論
はじめに
プラスチック(高分子材料)は低コストで優れた物性と耐久性を有するため,人々の生 活に不可欠な材料となっている。一方で,自然環境中では分解しないという性質が環境問 題を引き起こしている。汎用プラスチックとして大量に利用されているポリエチレン(PE) やポリスチレン(PS)は自然環境中では分解に30年以上の歳月を必要とする1-3。そのため,
環境流出したプラスチックは河川 4,5を経由し,湖5-8や海 9-11へ流出し,岸,水面,水中お よび底に蓄積する12-16。実際,2010年に全世界で480万トンから1270万トンのプラスチッ クが海洋流出したことが報告されている17,18。主に,密度の低いプラスチックは浮き、密度 の高いものは沈むことが想定される19。しかし,プラスチックは密度に関係なく底に堆積す ることが確認されている14-16。その要因の一つして,プラスチックへのバイオフィルムの形 成による重量増加が報告されている20。
環境中に流出したプラスチックが引き起こす具体的問題として,漁網や梱包用ロープの 生物への物理的な絡まりがある21-23。このプラスチックの絡まりはアザラシなどの哺乳類で 顕著に見られ,これは哺乳類の個体数の減少の一因と考えられている23,24。特に,漁網によ る被害は深刻である25-27。環境中に流出しコントロールを失った漁網はゴーストネットと呼 ばれ海洋中を漂っている。ゴーストネットは,魚類や哺乳類といった海洋生物を捕獲(ゴ ーストフィッシング)し続ける。
プラスチックは,長期的には光・酸素などによって物理的に崩壊し,プラスチック小片
(マイクロプラスチッック)まで断片化する。これらのマイクロプラスチックに加えて,
衣服などの繊維から出たマイクロファイバーは環境中に拡散し続けている。これらは環境 中に蓄積するため,新たな環境問題となっている。海底から採取されたイガイ(Mytilus edulis),ゴカイ(Arenicola marina)28,および,海洋から捕獲されたエビ29の体内にはマイ クロプラスチックが含まれていた。魚のプラスチック摂取量調査によれば,魚個体の35 % がプラスチックを摂取しており,1個体あたりの消化管から平均2.1個のプラスチックが見 つかっている 30。また,海鳥やウミガメの体内からもプラスチックが見つかっている 31-33。
告された38。特に,ポリエステル型ポリウレタンは分解を受け,さらにポリエステルウレタ ンのモノマーユニットであるポリエチレンアジペート,ポリプロピレンアジペートおよび ポリブチレンアジペートも分解することがわかった。
Figure 1. Structures of polyester urethane, polyether urethane, and poly(alkylene adipate).
この研究は生物劣化する高分子構造に関する最初の報告であるが,これ以降高分子材料 が引き起こす問題の解決策として生分解性高分子が注目されるようになった。多くの環境 分解性の生分解性高分子は,次の2つのステップで生分解が進行することが知られている
(Figure 2)39。
1)環境中の微生物が産生した菌体外酵素による低分子量化
2)低分子量化された化合物が微生物により代謝されることで,無機物にまで分解
Figure 2. Environmental degradation mechanisms of PAADs.
生分解性高分子の代表例として,微生物の生体内で生産されているポリ(3-ヒドロキシブ タン酸)(P(3HB))がある(Figure 3)40。P(3HB)は微生物の生体内でエネルギー源として蓄 積されている高分子である。また,P(3HB)は,微生物により分解・代謝されるため,元来 優れた環境分解性を有している。土壌環境中では様々な種類の細菌や真菌が P(3HB)の生分 解に関与している 41,42。また,環境水中においても P(3HB)は速やかに分解され,土壌環境 中と同様に環境水中の細菌や真菌がその生分解に関与している43,44。すなわち,微生物が生
O (CH2)l C (CH2)4 C O
O
O
n
Poly(alkylene adipate) l = 2 ~ 4 O (CH2)l C (CH2)m
O O
O
Polyester polyurethane
C z
nO (CH2)l H
N (CH2)y C H O N C
O O
n z
O (CH2)x H
N (CH2)y C H O N C
O O
Polyester polyurethane O (CH2)l C (CH2)mC
O O
O
n
HO (CH2)l OH
C (CH2)mC O HO
O Poly(n-alkylene n-alkylene dicarboxylate) OH
(PAAD)
Hydrolysis products Hydrolysis
Hydrolase
Microorganism Metabolism
CO2 H2O
First step Second step
Biodegradable polymer
Water soluble products Degradation
Hydrolase
Microorganism Metabolism
CO2 H2O
First step Second step
産する P(3HB)は,その分解性において環境依存性が低い生分解性高分子であることがわか る。
Figure 3. Structure of poly(3-hydrokybutylate).
P(3HB)の分解性とは異なり,化学合成よって得られる生分解性高分子の生分解性は一般
的には安定的に発現しない。一方で,P(3HB)と比較して化学合成生分解性高分子は物性制 御のための一次構造制御が容易であるため,様々な構造の生分解性高分子が創出されてい る。とりわけ脂肪族ポリエステルは,物性と生分解性の両面で優れていることから,農業 分野など様々な分野で利用されている45。
現在,化学合成脂肪族ポリエステルとして,ラクトンやヒドロキシカルボン酸から合成 されるAB型のポリエステルであるポリラクトンと,AA型の直鎖脂肪族ジカルボン酸(DCA)
とBB型の直鎖脂肪族ジオールからなるポリAABB型のポリエステルであるポリ(n-アルキ レン n-アルキレンジカルボキシレート)(PAAD)が,生分解性高分子として研究が進めら れている(Figure 4)39。
Figure 4. Structure of polylactone and poly(n-alkylene n-alkylene dicarboxylate) (PAAD).
ポリラクトン
代表的な化学合成系ポリラクトンの一つとしてポリ乳酸(PLA)が挙げられる(Figure 5)。
PLA は,デンプンを微生物発酵することにより生産される乳酸を化学重合することで得ら
CH CH2
O O
CH3
C n
Poly(3-hydroxybutylate) [P(3HB)]
O (CH2)l C (CH2)m C O
O
O
n
Poly(n-alkylene n-alkylene dicarboxylate) (PAAD)
O R C O
n
Polylacton
Figure 5. Poly(lactic acid), lactic acid, and alanin, poly(ε-caprolactone).
ε-カプロラクタムの開環重合によって得られるポリカプロラクトン(PCL)(Figure 5)
は,クチン分解酵素であるクチナーゼにより分解されることが報告されている55。これは,
PCLが天然物であるクチンやスベリンと構造56,57の類似性を有しているためである。また,
PCL は環境中に存在する様々な種類の微生物によって分解され,その分解微生物は環境中 に広く存在している58-62。PCLは,エステラーゼやリパーゼによって加水分解され60,63,他
方,P(3HB)分解酵素によって分解されない63。また,PCLを含むメチレン鎖長n = 4-7の直
鎖構造を持つポリラクトン(Figure 6)は,n = 4, 5, 7は同じ種の微生物により生分解された が,n = 6のみ異なる微生物により生分解された64。この結果は,AB型ポリエステルでも主 鎖の脂肪族鎖長の違いが,生分解性やその分解機構に影響を及ぼすことを示唆している。
Figure 6. Structures of polylactones.
ポリ(n-アルキレン n-アルキレンジカルボキシレート) (PAAD)
ポリ(n-アルキレン n-アルキレンジカルボキシレート)(PAAD)は,基本骨格となるジオ ールとDCAの組み合わせで様々な類縁体が合成可能である。多くのPAADはCarothersに より1940年までにその合成に成功している65。当初は重合度を上げることが困難で,物性 上問題がありPDDAは注目されてこなかった。しかし,高分子量のPAADの合成方法が開 発され 66,さらに PAADの環境分解性が報告されたことから,PAADは生分解性高分子と して再び注目されている 58。いくつかの PAADは,現在生分解性高分子として市販されて おり(Figure 7),それらの生分解性については詳細に調べられている67。PBSuおよびポリ(ブ チレンサクシネート-co-ブチレンアジペート)(PBSA)は,昭和電工からビオノーレとして 販売されている66。また,PBSuは三菱化学からバイオPBSとして販売されている68。その 他に商用販売されていたPAADにポリ(エチレンサクシネート)(PESu)がある。そのため,
PAADの一次構造と生分解性との相関についてある程度の知見の蓄積がある。
O (CH2)5 C O
n
Poly(ε-caprolactone) (PCL) CH C
HO O CH3
OH H2N CH C O CH3
OH Lactic acid Alanine CH C
O n
O CH3
Poly(lactic acid) (PLA)
O (CH2)n C O
m
n = 4 : Poly(δ-valerolactone) n = 5 : Poly(ε-caprolactone) n = 6 : Poly(ζ-enantolactone) n = 7 : Poly(η-caprylolactone)
Figure 7. Structues of commercially available PAADs.
5種類のリパーゼによるPESu,PBSuおよびPBSAの分解性評価では,PBSAおよびPESu は全てのリパーゼにより分解された67。特に,すべての酵素によるPBSAの分解量は,PESu のそれよりも多かった。一方,PBSuはPseudomonas sp.由来リパーゼのみによる分解を受け た。土壌中から単離された細菌Acidovorax delafieldii BS-3株は,固体のPBSA分解能を有し ていることが報告され69,さらにこのBS-3株から精製された酵素は固体のPBSA分解能を 有していた70。
土壌環境中におけるPBSu分解微生物の存在分布調査71およびPBSu分解微生物の単離が 行われている71,72。Microbispora rosea subsp. aerata IFO 14046株73とAmycolatopsis sp HT-6 株は,固体の PBSu分解能を有していた 74,75。PBSu 分解真菌として単離された Aspergillus fumigatus NKCM1706株は固体のPBSuを分解した76。さらにFusarium solaniもPBSuを分 解することが報告されており77,またF. solaniから生成されたクチナーゼもPBSu分解能を 有していた78。
実環境中における PBSuおよびPLA製の農業用マルチフィルムの土壌分解性が調べられ た 79。PBSuからなるフィルムは40日後に崩壊したが,PLAは 4ヶ月間分解しなかった。
19地点におけるPAADsとポリラクトンの土壌埋設試験において12ヶ月で初期重量の40 % 以上が分解した地点の数は,PBSA(13地点)>PCL(10地点)>PBSu(5地点)>PLA(2 地点)であった80。PBSuとPCLの土壌埋設試験では,12ヶ月間でPCLが25 %,PBSuが
O (CH2)2 C (CH2)2 C O
O
O
n
Poly(ethylene succinate) (PESu) O (CH2)4 C (CH2)2 C
O O
O
n
Poly(butylene succinate) (PBSu)
O (CH2)4 C (CH2)2 O O
O
Poly(butylene succinate -co- butylene adipate) (PBSA)
O (CH2)4 C (CH2)4 C O
O
O
C n
近年,PAAD はその生分解性という特性に加え,バイオマス由来ポリエステル(バイオ ベースポリマー)としても注目を集めている82。ポリ(ブチレンサクシネート)(PBSu)のモ ノマーである 1,4-ブタンジオール(BD)とコハク酸(SuA)は,グルコースを原料として 発酵生産により製造されている 83,84。セバシン酸(SeA)はひまし油のアルカリ熱分解で得 られる85。さらに,長鎖脂肪族鎖を有するジオール,ジカルボン酸,およびヒドロキシカル ボン酸は植物油由来の不飽和脂肪酸から合成され,これらから構成される長鎖脂肪族ポリ エステルは新規生分解性高分子と注目されている86,87,88-90。しかしながら,合成に関する報 告例はあるものの,生分解に関する報告例はない。
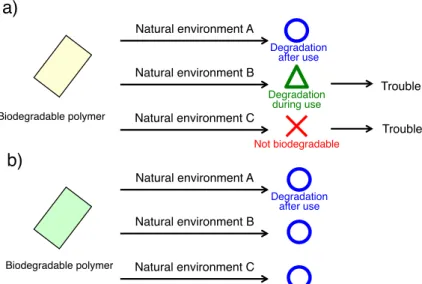
生分解性高分子の課題
これまで生産されてきた生分解性高分子は使用中にも徐々に分解が進行したり,使用後 に速やかに分解しないなどの欠点があった(Figure 8-a)。これは各々の環境に存在している 微生物の種類が異なっていることが原因であると考えられる。理想の生分解性高分子は,
暴露される環境を問わず使用時は分解せず,使用後に速やかに分解を開始するような仕組 みを持つことが望まれる(Figure 8-b)。
Figure 8. Environmental degradability of a) conventional biodegradable polymers and b) ideal biodegradable polymers.
生分解性制御方法
生分解性高分子は主に微生物の働きによって低分子量化される。しかし,環境中では高 分子分解酵素を生産する微生物は偏在しており,環境により生分解性高分子の分解性が大 きく異なる44,61,74。その一方で,低分子量化された水溶性モノマーは環境中の微生物により
Natural environment A Natural environment B
Natural environment C
Degradation during use
Not biodegradable Degradation
after use
Biodegradable polymer
Trouble Trouble
Natural environment A Natural environment B Natural environment C
Degradation after use
Biodegradable polymer
a)
b)
速やかに分解される91。そのため,生分解性高分子の生分解開始時期をコントロールするた めには,環境微生物種に依存しない低分子量化方法を提案する必要がある。
PLAは中温環境(30 °C)では分解せず,高温環境(50 °C),特にコンポスト中で分解す る92。これは,PLAのTgが55-60 °Cであり,ガラス転移温以下では分解が進行しにくいた めであると考えられる。そのために,PLA は使用中に分解されず,コンポスト投入で分解 開始する生分解性高分子として利用できる45。この場合,高温高湿のコンポスト環境が外部 刺激として働き生分解性を制御している。しかしながら,環境中に流出した高分子材料や 農業用マルチフィルムなど,オンサイト処理が望まれる生分解性高分子の生分解制御には 不向きである。
オンサイト処理を指向した生分解制御方法として,PBSA フィルムや PCL フィルムへの デンプン添加する方法が提案されている93,94。しかしデンプンの添加によりフィルム全体の 分解度は増大するが分解時期の制御には至っていない。酸化カルシウムを生分解性高分子 に添加する方法 95 は,酸化カルシウムが水酸化カルシウムとなり,フィルムから溶出しク ラックを形成し,さらに,塩基触媒的加水分解を併発し,劣化が促進する仕組みである。
劣化速度は増大するものの,使用中からの劣化が考えられるなど,分解時期の制御はでき ない。その他の生分解コントロール方法として,使用後の農業用マルチフィルムに生分解 性高分子分解能を持つ酵素を撒くことで,生分解を加速させる方法も提案されている96。こ の場合は,分解酵素の生産および散布に膨大なコストが必要となる。
以上は市販の生分解性高分子をそのまま利用して生分解性を制御する手法であるが,基 礎研究の分野では自然環境中の外部刺激に応答して化学構造が変化し,生分解を開始する 外部刺激応答性生分解性制御方法が検討されている。これまでに,PBSuオリゴマーをイミ ン結合で連結し,イミン結合の可逆的加水分解(Figure 9)により生分解開始時期を制御す る方法が報告されている97。このイミン結合を有するポリマーは,23 °C の水中で50時間 で数平均分子量が半減した。しかし,少量の水存在下で加水分解が進行するため,使用用 途が限定される。
本研究の目的
生分解性高分子の生分解開始時期を制御するためには,どのステップが律速であるのか を明らかにする必要がある。さらに生分解性高分子の一次構造により生分解開始時期を制 御できるか検討,その結果から新規生分解開始時期制御方法を考案する必要がある。これ までの生分解に関する研究においては特定の生分解性高分子を対象にしており,系統的な 生分解性評価が行われてこなかった。そこで本博士論文では,まず第 2 章において,生分 解性高分子の一群である脂肪族ポリエステルの脂肪族鎖長と環境分解性との関係を明らか にすることを目的として研究を進めた(Figure 10)。
Figure 10. Evaluation of biodegradeability of PAAD.
第 2 章で,生分解性高分子の低分子量化過程は環境中に存在している微生物に依存して いることを見出した。これを受けて第 3 章では,環境中に広く存在している外部刺激によ り生分解開始時期を制御する方法を検討した。まず高分子の鎖を非生物学的分解で断片化 し,その断片化物を微生物分解によって無機化することで,生分解性高分子の分解時期制 御実現を目的として研究を進めた。非生物学的な分解として,還元によってチオールへと 可逆的開裂をするジスルフィド結合に着目した。土中や底質における環境は還元的雰囲気 にあるため,ジスルフィド結合をリンカー部位として有する高分子は,環境中でチオール へと開裂を起こして低分子量化した後,微生物分解すると考えられる。モノマーユニット を結ぶリンカー部位としてジスルフィド結合を有する高分子を合成する。これを還元的環 境下で,非生物学的分解と微生物代謝を多段階的に分解することで高分子の分解時期制御 の実現を目指した(Figure 11)。
O (CH2)l C (CH2)m C O
O
O
n
HO (CH2)l OH
C (CH2)m C O
HO
O Poly(n-alkylene n-alkylene dicarboxylate) OH
(PAAD)
Hydrolysis products Hydrolysis
Hydrolase
Microorganism Metabolism
CO2 H2O
First step Second step
Biodegradable polymer
Water soluble products Degradation
Hydrolase
Microorganism Metabolism
CO2 H2O
First step Second step
Figure 11. Schematic mechanism for controlled degradation of polydisulfide.
HS SH S S n
S S n
Surface of River or Sea (Higher ORP Condition)
CO2 + H2O + SO3
Bottom of River or Sea (Lower ORP Condition) Release
after use Reductive
degradation Biodegradation
Trigger on
第2章
ポリ(ブチレン n- アルキレンジカルボキシレート)
の合成と生分解性評価
2-1 緒言
本章ではジオールとジカルボン酸からなる PAAD の脂肪族鎖の長さと生分解性との相関 関係の解明が,目的である。PAAD のモノマーとしては任意のジオールユニットとジカル ボン酸ユニットを用いることができるが,市販のPAADにおいてジオールユニットでは1,4- ブタンジオール(BD)が多く用いられている(BASF社が生産しているポリ(ブチレンアジ ペート-co-ブチレンテレフタレート)(PBAT),三菱化学株式会社などが生産しているポリ
(ブチレンサクシネート)(PBSu),昭和電工株式会社などが生産しているポリ(ブチレン サクシネート-co-アジペート)(PBSA),Figure 12)。そこで,本研究ではBDをジオールユ ニットとして用いる。そして,ジカルボン酸ユニットとしては長さの異なる直鎖脂肪族ジ カルボン酸を用いる。
Figure 12. Structures of PBAT, PBSA, and PBSu.
本博士論文では,カルボニル間のメチレン鎖数(n)の異なる9種類のジカルボン酸(DCA)
(コハク酸(n = 2; SuA),グルタル酸(n = 3; GlA),アジピン酸(n = 4; AdA),ピメリン酸
(n = 5; PiA),スベリン酸(n = 6; SbA),アゼライン酸(n = 7; AzA),セバシン酸(n = 8; SeA),
ウンデカン二酸(n = 9; UdA),およびドデカン二酸(n = 10; DdA))とBDとの縮合反応に より9種類のポリ(ブチレン n-アルキレンジカルボキシレート)(PBAD)を合成した(Figure
O (CH2)4 C (CH2)2 O O
O
Poly(butylene succinate -co- butylene adipate) (PBSA)
O (CH2)4 C (CH2)4 C O
O
O
C n
O (CH2)4 C (CH2)4 O O
O
Poly(butylene adipate -co- butylene terephthalate) (PBAT)
O (CH2)4 C O O
O
C C n
O (CH2)4 C (CH2)2 C O
O
O
n
Poly(butylene succinate) (PBSu)
Figure 13. Structure of poly(butylene n-alkylene dicarboxylate) (PBAD).
O (CH2)4 C (CH2)n C O
O
O
m
Poly(butylene n-alkylene dicarboxylate) (PBAD)
BD unit DCA unit
n 2 3 4 5 6 7 8 9 10
DCA SuA GlA AdA PiA SbA AzA SeA UdA DdA
PBAD PBSu PBGl PBAd PBPi PBSb PBAz PBSe PBUd PBDd
2-2 実験 試薬
AdA,AzA,DdA,GlA,SuA,SeA,メタノール,クロロホルム,クロロホルム-d1(0.05
v/v% TMS),Yeast Extract,Polypepton,塩化ナトリウム,水酸化ナトリウム,およびリン酸 二水素カリウムは和光純薬工業から購入した。BD,AdA,PiA,SbA,UdA,チタンテトラ シオプロポキシドは東京化成株式会社から購入した。2,5-ヒドロキシ安息香酸,クロロホル ム,ジクロロメタン,リン酸水素二カリウム,リン酸水素二ナトリウム二水和物,硫酸マ グネシウム七水和物,塩化カルシウム二水和物,塩化鉄六水和物は関東化学株式会社から 購入した。BDは蒸留精製したのちに使用した。
核磁気共鳴スペクトル (NMR)
NMRの測定には日本電子株式会社製ECA- 600を用いた。溶媒として0.05v/v%TMS含有 重クロロホルムを用いた。PBAD(10 mg)を重クロロホルム(0.6 mL)に溶解させ試料を 調整した。
分子量測定
合成した PBAD の分子量はゲルパーミエーションクロマトグラフィ(GPC)により算出 した。GPC測定にはLC-2000 GPC システム(日本分光株式会社)を使用し,流速1.0 mL·min−1,
温度40 °Cで測定した。検量線の作成には東ソー株式会社製のTSK標準ポリスチレン(数
平均分子量=1.0 106, 3.5 105, 9.5 104, 3.9 104, 8.9 103, 2.6 103, 2.7 102)を使用した。3種 類のカラム(TSKgelGMHXL,TSKguardcolumnHXL-H,TSKgelG2000HXL;東ソー株式会社)
を連結して使用し,RI検出器を用いて検出した。試料 (5.0 mg) をHFIP (0.1 mL)に溶解させ た後,クロロホルム (1.0 mL) を加えて測定溶液を調整した。調整した溶液を孔径0.45 µL のフィルターで濾過し,マニュアルインジェクション(100 µL)で測定した。
マトリックス支援レーザー脱離イオン化飛行時間型質量分析(MALDI-TOF-MS)測定に
式会社)を用いた。容器としてアルミ製パン(P/N SSC000E030およびP/N SSC000E032:セ イコーインスツル株式会社)を用い,窒素雰囲気下,10.0 °C·min−1で,室温から120 °Cま で昇温,3分間保持の後,10.0 °C min−1で,−70 °Cまで降温,3分間の保持の後,10.0 °C·min−1
で,120 °Cまで昇温して測定した。
動的粘弾性測定 (DMA) にはパーキンエルマー製DMA8000を用いた。試料10 mgをステ ンレス製板(マテリアルポケット)に挟み(Figure 14),120 °Cで試料を融解させた後,液 体窒素で急冷することで測定試料を調整した。測定は,−100~100 °Cの温度範囲で昇温速度 2.0 °C·min−1で行なった。
Figure 14. Sample preparation method of material pocket.
フィルム作成
PBADを20 20 0.1 mmのステンレス製金型の中央に置き,ポリテトレフルオロエチレン
が表面にコートされたポリイミドフィルム(カプトン®, 120HR616;東レ・デュポン株式会 社)で挟み,1 mm厚のステンレス板で挟んだ。130 °Cで15 MPa,2分間プレス成形の後,
徐冷することで各種PBADフィルムを得た。
広角 X 線回折(WAXD)
広角 X 線回折(WAXD)にはX 線回折装置(RINT2000V/PC;株式会社リガク)を用い た。測定試料としては,上記で作成した0.1 mm厚のフィルムおよびフィルムを凍結粉砕し た粉末を用いた。X 線源として CuKα 線を用い,電圧 40kV,電流 20 mA,スキャン速度
2.0 °·sec−1,2θ = 5-60°の条件で測定した。結晶化度は波形分離により,結晶由来のピークと
非晶由来のハローの面積を求め,下記の式に従って見積もった。
Ic:結晶性散乱強度,Ia:非晶性散乱強度 PBADs
Material Pocket
Bending
X
c= I
cI
c+ I
a×100
ポリ(ブチレン n-アルキレンジカルボキシレート) (PBAD)の合成 98
50 mLのナス型フラスコにBD (58.9 mmol),DCA (54.4 mmol) およびチタンテトライソプ ロポキシド (50 µmol)を加えた。窒素ガス雰囲気下200 °Cで1 時間撹拌した。その後,10 Pa
に減圧し240 °Cで4.5h撹拌した。生成した固体をジクロロメタン (15 mL) に溶解させ,メ
タノール (150 mL) に加えることで再沈殿を行なった。吸引濾過で沈殿物を回収し,室温で 減圧乾燥させることで白色固体を得た。
生物化学的酸素要求量 (BOD) 生分解度測定 44
BOD生分解度測定はBOD TESTER 200F (タイテック株式会社) を用いた。宮城県石巻市,
和歌山県和歌山市の土壌および静岡県浜松市佐鳴湖の底質を1 gずつ採取し,混合した。混 合した土壌(1 g)を蒸留水(1 L)に懸濁し濾過をした。濾液を1日静置後,上澄200 µlを 植種源としてフラン瓶に加えた。微生物が生育するのに必要な無機塩 (K2HPO4 ; 21.75 g·L−1, KH2PO4 ; 8.5 g·L−1, Na2HPO4・2H2O ; 33.4 g·L−1, NH4Cl ; 0.5 g·L−1, MgSO4・7H2O ; 22.5 g·L−1, CaCl2・2H2O ; 36.4 g·L−1, FeCl3・6H2O ; 0.25 g·L−1) を加え,全量が200 mLになるよう蒸留 水でメスアップした。コントロールとして,炭素源となる試料を加えていないフラン瓶を 一本用意し,20mgのアジ化ナトリウムを加えた。二酸化炭素吸収剤容器に4.0 % NaOH Aq.
(1.5 mL) を加え,フラン瓶上部に取り付けた。調整したフラン瓶をBOD TESTERに設置し,
スターラーを回し,30分後に消費酸素量の目盛をすべて同じ位置に調整した。25 °Cの暗室 下で撹拌を続け,定期的に消費酸素量を測定した。
<理論>
HO (CH2)4 OH C (CH2)n C O
HO
O
OH O (CH2)4 C (CH2)n C
O O
O
m
1,4-Butanediol (BD)
Dicarboxylic acid (DCA)
1) Ti(i-PrO)4 200°C, 1 h, in N2 2) 240°C, 4.5 h < 3.6 Pa
Poly(butylene n-alkylene dicarboxylate) (PBAD)
+
Table 1. Reaction formula and amount of sample for BODs
PBAD Reaction formula Formular weight
/ g·mol−1
Amount of Sample / mg
PBSu (C8H12O4)m + 9mO2 → 8mCO2 + 6mH2O 172 7.7 PBGl (C9H14O4)m + 10.5mO2 → 9mCO2 + 7mH2O 186 7.1 PBAd (C10H16O4)m + 12mO2 → 10mCO2 + 8mH2O 200 6.7 PBPi (C11H18O4)m + 15.5mO2 → 11mCO2 + 9mH2O 215 5.6 PBSb (C12H20O4)m + 15mO2 → 12mCO2 + 10mH2O 228 6.1 PBAz (C13H22O4)m + 17.5mO2 → 13mCO2 + 11mH2O 242 5.6 PBSe (C14H24O4)m + 18mO2 → 14mCO2 + 12mH2O 256 5.7 PBUd (C15H26O4)m + 19.5mO2 → 15mCO2 + 13mH2O 268 5.5 PBDd (C16H28O4)m + 21mO2 → 16mCO2 + 14mH2O 284 5.4
理論的酸素要求量 (ThOD)を9.0 mL に揃えるのに必要なサンプル量を,各酸化反応を用い て計算した。以下に代表してPBSuの計算式を示す。
X (mg) ÷ 172 (g·mol−1) × 9.0 (ThOD) × 22.4 (L·mol−1) = 9.0 (mL)
これより理論的酸素要求量 (ThOD) を9 mLに揃えるには7.7 mgのサンプルが必要となる。
生分解度の求め方
試料の生分解度は以下の式で求められる。
BOD生分解度 (%) = 測定した酸素消費量 (mL) / 理論的酸素要求量 (mL) × 100
PBAD 分解微生物の計数および単離 99
PBAD分解微生物の単離にはポリエステル乳化培地を用いた。PBAD(0.5 g)をジクロロ メタン (10 mL) に溶解した。ジクロロメタン溶液をプライサーフ (第一工業社) (0.01 v%) を含む蒸留水に加え,超音波処理(TOMY ultrasonic disruptor UD-200;トミー工業株式会社)
を 2 分間行なうことで,ポリエステル乳化液を作製した。これに微生物が生育するのに必 要な無機塩 (構成成分 (g·L-1): KH2PO4, 4.6: Na2HPO4・12H2O, 11.6: MgSO4・7H2O, 0.5:
FeCl2・6H2O, 0.1: NH4Cl, 1.0: Yeast extract, 0.1)を加えpH 7.0に調整した。その後,寒天粉末
2 % (w/v) を加え,110 °Cで20分間オートクレーブ滅菌後,ポリスチレン製薄型シャーレ
(90 15 mm;株式会社アテクト社製) に流し込んだ。
また単離したポリエステル分解微生物および大腸菌の培養は Luria broth (LB) 培地 (pH7.5, components (g·L−1): Yeast extract, 5.0: Polypepton, 10.0: NaCl, 5.0) で行った。培地は使 用前にオートクレーブ滅菌した。また固体LB培地には2 % (w/v) の寒天粉末を加えた。
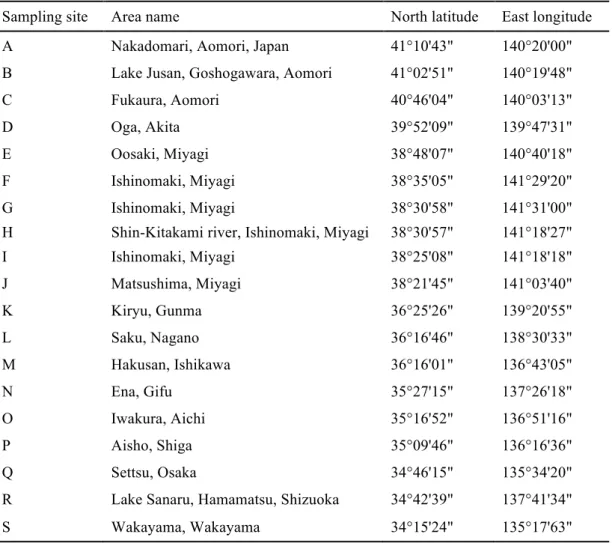
本研究では,全国19地点 (Table 2, Figure 15) から環境サンプル(土壌,砂,水および底 質)を採取した。環境サンプルを1.0 gずつ10 mLの滅菌超純水に懸濁し,10分間静置後,
100 µLを各種ポリエステル乳化培地上に塗布した。環境水サンプルは希釈せず100 µLを塗 布した。その後,30 °Cで培養し,クリアゾーンを形成したコロニーをポリエステル分解微 生物として計数し,各分解微生物の単離を行った。
Figure 15. Sampling sites of inocula in a) Japan and b) Miyagi prefecture of Japan.
Table 2. Latitude and longitude of sampling sites and condition of inoculum
Sampling site Area name North latitude East longitude A Nakadomari, Aomori, Japan 41°10'43" 140°20'00"
B Lake Jusan, Goshogawara, Aomori 41°02'51" 140°19'48"
C Fukaura, Aomori 40°46'04" 140°03'13"
D Oga, Akita 39°52'09" 139°47'31"
E Oosaki, Miyagi 38°48'07" 140°40'18"
F Ishinomaki, Miyagi 38°35'05" 141°29'20"
G Ishinomaki, Miyagi 38°30'58" 141°31'00"
H Shin-Kitakami river, Ishinomaki, Miyagi 38°30'57" 141°18'27"
I Ishinomaki, Miyagi 38°25'08" 141°18'18"
J Matsushima, Miyagi 38°21'45" 141°03'40"
K Kiryu, Gunma 36°25'26" 139°20'55"
L Saku, Nagano 36°16'46" 138°30'33"
M Hakusan, Ishikawa 36°16'01" 136°43'05"
N Ena, Gifu 35°27'15" 137°26'18"
O Iwakura, Aichi 35°16'52" 136°51'16"
P Aisho, Shiga 35°09'46" 136°16'36"
Q Settsu, Osaka 34°46'15" 135°34'20"
R Lake Sanaru, Hamamatsu, Shizuoka 34°42'39" 137°41'34"
S Wakayama, Wakayama 34°15'24" 135°17'63"
16S rDNA 配列解析 99
各ポリエステル分解細菌の16S rDNAをコロニーPCR法により取得した。使用したプライ
マー99,100は1525r (5'-AAAGGAGGTGATCCAGCC-3'),27f (5'- AGAGTTTGATCCTGGCTCA
G-3'),16Sr (5'- TACCTTGTTACGATTCA-3')および16Sf (5'- GTTTGATCATGGCTCAG -3')で あり,これらをそれぞれ20 pmolずつ用いた。爪楊枝でLB培地上のコロニーを採取し,滅 菌超純水に懸濁し,テンプレートとした。このテンプレートに10× Ex Taq Buffer (1.0 µL) , dNTP Mixture (1.0 µL) , プライマー (0.2 µL) ,TaKaRa Ex Taq (0.1 µL) を加え,滅菌超純 水で全量が10 µLとなるようメスアップした。サーマクルサイクルは,94 °C 5 分,(94 °C/20 sec,55 °C/30 sec,72 °C/90 sec) 25回,72 °C/10minで実行した。得られたPCR産物は
1.5 % (w/v) アガロースゲル中で電気泳動した。電気泳動後のアガロースゲルは10分間エチ
ジウムブロマイド溶液に浸漬し,蒸留水で10分間脱色した。その後,310nmのUV光をゲ ルに照射し,バンドの位置を確認した。
増幅したPCR産物 (3.0 µL) を,T-Vector pMD20 (Ampr, lacZ, TaKaRa社), 1.0 µL,T4 DNA Ligase, 1.0 µL, 10× Ligation Buffer, 1.0 µLおよび滅菌超純水4.0 µLを混合後,16 °Cで保温 することで,T-Vector pMD20に連結した。組み換えプラスミドは塩化カルシウム法により E. coli DH5α (supE44 ΔlacU169(φ80lacZΔM15) hsdR17 recA1 gyrA96 thi-1 relA1, TOYOBO社) に形質転換した。形質転換株の培養は,1 mM Ampicillin (60 µL),1 mM Isopropyl-β-D- thiogalactopyranoside (38 µL)および 0.1mM 5-Bromo-4-chloro-3-indolyl-β-D-galactopyranoside
(50 µL)が塗布された固体LB培地を用い,37 °Cの恒温槽で行った。プレート上に形成した
コロニーの中から白色のコロニーを選択することで,レプリカを作製した。プラスミドDNA のインサートチェックは制限酵素BamHⅠを用いて行った。プラスミドの消化はプラスミド (3 µL) , BamHI (1 µL),10 K Buffer (TaKaR社製) (1 µL) および滅菌超純水 (5 µL) を30 °C で 3 時間静置することにより行なった。インサートが確認されたコロニーからプラスミド を精製した。得られたプラスミドのシーケンシングは,オペロンバイオテクノロジー株式 会社にて依頼分析を行った。塩基配列および相同性解析はプログラム GENETYX あるいは プログラムblastn (http://www.ncbi.nlm.nih.gov/blast) を用いて行った。シーケンシングにより 得られた塩基配列データと,GeneBankより入手した塩基配列データを用いてclustalW法に よりアライメント解析をした。アライメント解析により得られたデータを用いて MEGA5 プログラムにより系統樹を発生させた。
PBAD 分解細菌のクリアゾーン形成能評価
「PBAD分解微生物の計数および単離」と同様に作成したPBAD乳化倍地上に,「PBAD 分解微生物の計数および単離」で単離したPBAD分解細菌19種を画線し,30 °Cで培養し 30日間培養した。形成したクリアゾーンの大きさが0.3 cm以下を+,0.3 cm以上0.5 cm未 満を++,0.5 cm以上を+++としてクリアゾーン評価能を評価した。
2-3 結果 PBAD の合成
BDと9種類のDCA(n = 2-10)の重縮合によりPBADを合成した。Figure 16にPBADの
1H NMRスペクトルを示す。
Figure 16. Partial 1H NMR spectra (600 MHz , CDCl3, 298 K) of PBAD.
得られたPBADの収率,分子量,熱的性質および結晶化度(Xc)をTable 3に示す。PBAd,
PBSbおよびPBUdの数平均分子量(Mn)は約1万程度であったが,その他のPBADはMn が4万以上の試料を得た。また再沈殿による精製後のサンプルであるため,いずれのPBAD の分散度(Mw/Mn)は2.0以下であった。
5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0
ppm PBSu
PBGl PBAd
PBPi
PBSb
PBAz
PBSe
PBUd
PBDd
Table 3. Yield and molecular weights of PBAD.
na PBAD Yieldb / % Mnc / 103 Mw/Mnc
2 PBSu 92 66.8 1.19
3 PBGl 82 63.7 1.20
4 PBAd 59 10.1 1.36
5 PBPi 68 40.3 1.73
6 PBSb 83 13.3 1.33
7 PBAz 82 57.7 1.23
8 PBSe 82 49.7 1.31
9 PBUd 81 9.4 1.68
10 PBDd 90 44.3 1.33
anumber of methylene unit in dicarboxylic acid unit. bmethanol insoluble part. cmeasured by GPC (eluent:chloroform) and calibrated with polystyrene standards.
PBSeのMALDI-TOF-MS測定結果をFigure 17に示す。片末端にBDが片末端にSeAを持 つPBSeの繰り返しが確認された。また,両末端にBDを有するPBSeの繰り返しが観測さ れた。また,カリウム,リチウムおよびカリウムが付加したPBSeのピークが観測された。
O (CH2)4 C (CH2)8 C O
O
O
m
PBSe
O (CH2)4 C (CH2)8 C O
O
O
mO CH2 CH2 CH2 CH2 OH
Na+
Li+ K+
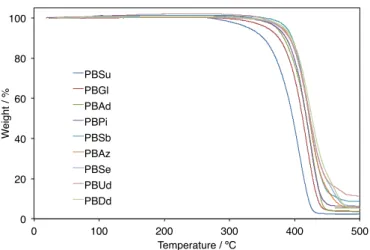
Figure 18. TGA charts of PBAD using a temperature ramp rate of 10 °C·min−1
Figure 19にDSC測定のチャートを,Table 4に結晶化温度(Tc)と融点(Tm)を示す。
Tcは降温測定時に,Tmは2回目の昇温測定時に観測された。PBSuのTmが最も高い114 °C であるが,その他のPBADのTmは70 °C以下であった。またDSC測定においてガラス転 移点(Tg)は観測されなかった。
Figure 19. DSC charts of PBAD in a) first cooling cycle at a rate of 10 °C·min−1 b) second heating cycle at a rate of 10 °C·min−1.
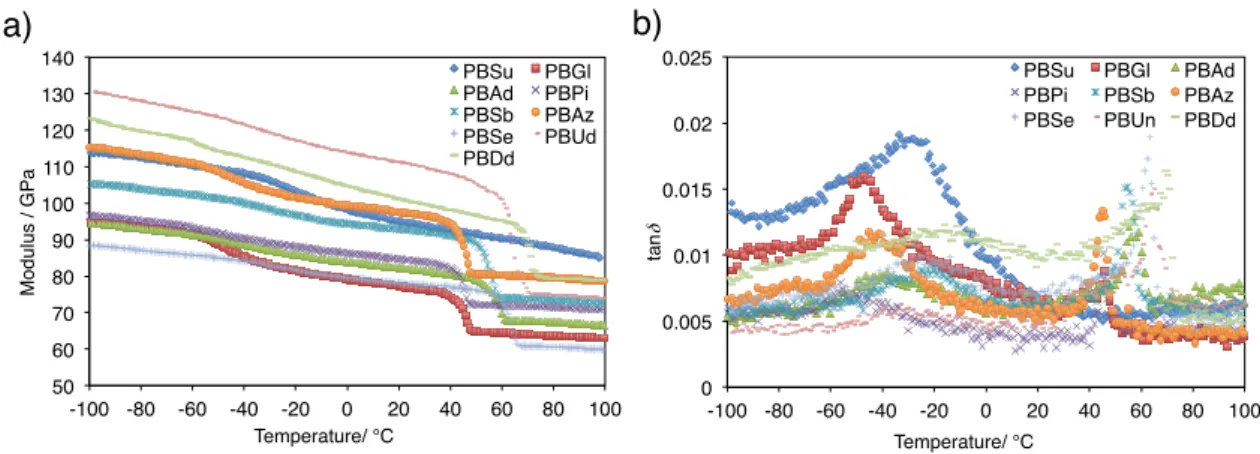
PBADのDMA測定のtanδの値よりTgを見積もった。Figure 20にPBDAのDMAチャー トを,Table 4にDMA測定で得られたPBDAのTgの値を示す。全てのPBADのTgは0 °C 以下であった。また,PBSuを除くポリエステルでは40~70 °Cに融解に由来するtanδのピ ークが観測された。DMA測定の温度範囲はDSC測定で観測されたPBSuの融点(114 °C) 以下であるため,PBSuの融解に由来するピークは確認されなかった。
0 20 40 60 80 100
0 100 200 300 400 500
Weight / %
Temperature / ºC PBSu
PBGl PBAd PBPi PBSb PBAz PBSe PBUd PBDd
Temperature/ ºC Temperature/ ºC
a) b)
Exo.
Exo.
-60 -40 -20 0 20 40 60 80 100 120 PBDd PBUd PBSe PBAz PBSb PBPi PBAd PBGl PBSu
-60 -40 -20 0 20 40 60 80 100 120 PBDd PBUd PBSe PBAz PBSb PBPi PBAd PBGl PBSu
Figure 20. DMA charts of PBAD. a) modulus, b) tanδ.
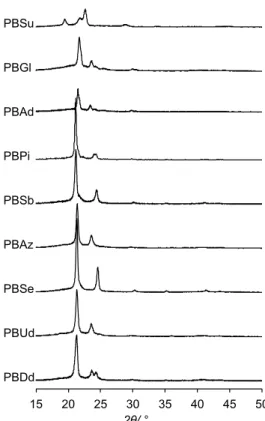
Figure 21にPBADのWAXDパターンを,Table 4にWAXDパターンを解析して得られた 結晶化度(Xc)を示す。PBSuでは2θ = 19.4,21.7,22.5,28.7,および33.6 °に回折ピーク が観測された。PBGlでは2θ = 21.7,23.6,25.3および29.9 °に回折ピークが観測された。
PBAdでは2θ = 21.4,21.7,23.4,24.1,25.2および29.7 °に回折ピークが観測された。PBPi では2θ = 21.1,24.1および29.9 °に回折ピークが観測された。PBASbでは2θ = 21.2,24.3,
29.9および34.5 °に回折ピークが観測された。PBAzでは2θ = 21.4,23.6,29.6および36.4 ° に回折ピークが観測された。PBSeでは2θ = 21.4,24.5,30.1,36.4および41.1 °に回折ピー クが観測された。PBUdでは2θ = 21.3,23.5,29.6,35.8および40.8 °に回折ピークが観測さ れた。PBDdでは2θ = 21.3,23.6,24.2,29.6,35.8および40.4 °に回折ピークが観測された。
0 0.005 0.01 0.015 0.02 0.025
-100 -80 -60 -40 -20 0 20 40 60 80 100 PBSu PBGl PBAd PBPi PBSb PBAz PBSe PBUn PBDd
50 60 70 80 90 100 110 120 130 140
-100 -80 -60 -40 -20 0 20 40 60 80 100 PBSu PBGl PBAd PBPi PBSb PBAz PBSe PBUd PBDd
a) b)
Temperature/ °C Temperature/ °C
Modulus / GPa tanδ
Figure 21. Partial WAXD patterns of PBAD. The region without peaks was omitted to clarify.
Table 4. Thermal properties, and crystallinity of PBAD.
PBAD Td5%a /°C Tgb /°C Tcc /°C ∆Hcc /J·g−1 Tmd /°C ∆Hmd /J·g−1 Xce /%
PBSu 330 −32 83 99 105, 114 99 41 ± 2
PBGl 357 −49 13 60 32, 40 63 24 ± 2
PBAd 372 −41 39 71 52, 56 78 27 ± 1
PBPi 374 −56 16 76 43 79 36 ± 0
PBSb 387 −23 36 103 59 110 59 ± 1
PBAz 381 −44 22 77 43 76 37 ± 1
PBSe 379 −28 49 116 65 125 29 ± 1
PBUd 379 −31 38 71 56 91 45 ± 3
PBDd 384 −16 56 120 71 125 61 ± 1
ameasured by thermal gravimetric analyzer. bmeasured by dynamic mechanical analyzer. cmeasured during the first cooling scan with a rate of 10 °C min−1. dmeasured during the second heating scan with a rate of 10 °C·min−1. emeasured using melt press film by X-ray diffractometer from 2θ = 5 ° to 60 °
15 20 25 30 35 40 45 50 2θ/ °
PBSu
PBGl
PBAd
PBPi
PBSb
PBAz
PBSe
PBUd
PBDd
BOD 生分解度評価
Figure 22-aにモノマーであるBDおよびDCAのBOD生分解度曲線を示す。DCAは培養
開始3日までに分解が始まり,30日目のBOD生分解度はそれぞれSuA:70 %,GlA:71 %,
AdA:80 %,PiA:66 %,SbA:75 %,AzA:88 %,SeA:56 %,UdA:68 %,DdA:59 % であった。BDは,培養開始5日目に分解が始まり30日目のBOD生分解度は76.3 %であっ た。曲線の立ち上がり部分の傾き(%·day−1)はSuA:14.9,GlA:15.9,AdA:15.5,PiA:
14.1,SbA:18.5,AzA:15.3,SeA:14.4,UdA:13.4,DdA:11.5,BD:16.4であった。
Figure 22-bに,PBADのBOD生分解度曲線を示す。PBSuおよびPBDd以外のPBAD (n =
3 ~ 9) は培養開始3日目に分解が始まった。PBSu(n = 2)は培養開始14日目に分解が始ま
った。PBPi のBOD生分解度が最も高く,28日目のBOD生分解度は 72.2 %であった。28 日目のBOD生分解度はPBAz,PBSb,PBAd,PBGl,PBSe,PBUd,PBSuの順に減少し,
それぞれ,69, 65, 63, 60, 57, 46, および27 %であった。PBDd(n = 10)の30日後のBOD生 分解度は,5 %であった。BOD曲線の立ち上がり部分の傾き(%·day−1)を比較すると,PBA の傾きが大きく 6.3 であった。そして,PBPi,PBGl,PBSb,PBAz,PBSe,PBUd,PBSu,
PBDdの順で傾きが減少し,それぞれ5.5, 4.9, 4.8, 2.9, 2.8, 2.2, 1.5, 0.1 %·day−1であった。
a) b)
20 30 40 50 60 70 80 90 100
BOD biodegradability / %
PBSu PBGl
PBAd PBPi
PBSb PBAz
PBSe PBUd
PBDd
20 30 40 50 60 70 80 90 100
BOD biodegradability / %
BD SuA
GlA AdA
PiA SbA
クリアゾーン形成試験
Table 5に,各環境サンプルの生菌数および生菌に対する分解微生物数を示す。各環境サ
ンプルの生菌数は1 × 103 ~ 1.7 × 106 cfu·g−1 であった。分解微生物の存在が確認された地点 数はPBAd (18 地点) > PBGl (17 地点) > PBUd (16 地点) > PBSb (15 地点) > PBAz (14 地点)
> PBPi (10 地点) > PBSe (7 地点) > PBDd (6 地点) > PBSu (5 地点)の順で低下した。DCAの メチレン数が 3~7 および 9 のポリエステルは,ほぼ全ての地点において分解微生物の存在 が確認された。
Figure 23に本研究で単離した31のPBAD分解細菌の16S rDNA配列を基に発生させた系
統樹を示す。TBTK040312A株,TBTK040711C株,TBTK040915A株およびTBTK040915B 株は Acinetobacter 属に,TBTK040306C 株,TBTK040506C 株および TBTK040703A 株は Peseudomonas属に近縁な種であった。TBTK040410B株はBurkholderia属に,TBTK041010A 株はCupriavidus属に近縁な種であった。TBTK040607C株はRhizobium属に,TBTK040402B 株,および TBTK040402D 株は Rhodobacter 属に近縁な種であった。TBTK040913A 株は Streptomyces属に,TBTK040209C株はArthrobacter属に,TBTK040502A株はMicrobacterium 属に近縁な種であった。また,TBTK040501B株はPaenibcillus属に,TBTK040606B株,お よびTBTK040404A株はExiguobacterium属に近縁な種であった。さらに31のPBAD分解細
菌のうち13株がBacillus属に近縁な種であった。
Table 6には同定した31種類のうち19株の細菌の,ポリエステル乳化培地上におけるク
リアゾーン形成能を示す。PBSu,PBGl,PBAd,PBPi,PBSb,PBAz,PBSe および PBUd のポリエステルに対しては,半数である 12 株以上の分解細菌がクリアゾーンを形成した。
しかし,PBDd に対しては,9株がクリアゾーン形成した。PBDdを分解する 9株のうち,
TBTK040705A株を除く8株はすべてのPBAD乳化培地上にクリアゾーンを形成した。
AD-degrading microorganisms to viable cell counts in 19 sites of Japan. Viable cell counts /×103 cfu
·
g−1PBAD-degrading microorganism counts / Viable cell counts/ % PBSuPBGl PBAdPBPiPBSbPBAz PBSePBUdPBDd 11 0.913129.13.62.51.29.01.2 160 NDND1.0NDNDNDND6.3ND 25 ND3.633NDND13ND2.0ND 130 1.64.8134.86.43.29.6615.0 1300 ND8.02012ND9.07.0129.7 200 ND5.7162.1182.8ND2.58.0 1400 ND126.5ND6.93.8ND4.1ND 100 ND2.09.7ND4.30.7NDNDND 580 1.08.0125.43.05.0ND0.9ND 1 0.1ND7.04.3NDNDND38ND 100 ND2.93.84.6194.88.76.029 500 ND8.3ND5.81011ND0.4ND 150 ND139.2ND18NDND662.0 1700 ND9.88.00.68.914ND12ND 460 ND1115ND7.2NDND5.5ND 110 ND8.711ND8.68.43.0NDND 250 ND106.8ND7.9ND0.16.0ND 47 6.41.06.44.38.5178.5NDND 360 ND2.015ND1610ND5.9ND ing PBAD-degrading an517181015147166Figure 23. Neighbor-joining phylogenetic tree of PBAD-degrading bacteria and related bacteria based on 16S rDNA sequence comparisons. The bar indicates 10 % estimated sequence divergence.
E.coli TBTK040312A
Acinetobacter calcoaceticus AY346313 TBTK040711C
Acinetobacter johnsonii AB099655 TBTK040915A
TBTK040915B TBTK040506C
Pseudomonas putida AY958233 TBTK040306C
Pseudomonas fluorescens EU439419 TBTK040703A
TBTK040410B
Burkholderia grimmiae strain R27 NR 18268 TBTK041010A
Cupriavidus necator N-1 CP002877 Ralstonia eutropha H16 AM260479 TBTK040607C
Rhizobium cellulosilyticum NR 043985 Rhizobium galegae Z79620
TBTK040402D
Rhodobacter sp SgZ-3 JX113682 TBTK040402B
Rhodobacter capsulatus SB 1003 CP001312 TBTK040913A
Streptomyces sp AC113
Streptomyces badius partial HF935087 TBTK040209C
Arthrobacter aurescens strain NI1 JQ435703 TBTK040502A
Microbacterium oxydans strain M66 1A JN644529 TBTK040501B
Paenibacillus sp. ZY-8 FJ232954 TBTK040606B
TBTK040404A Exiguobacterium undae NR 043477 TBTK040305D
TBTK040705A
Bacillus subtilis strain KT1002 AB115958 TBTK040405D
BTK040502C TBTK040711A Bacillus altidinis FM955870 TBTK040607A
TBTK040311B TBTK00707D TBTK040711B
Bacillus thuringiensis AY138288 TBTK040607B TBTK041005B TBTK040811B
TBTK040502D
0.1
γ-Proteobacteria
β-Proteobacteria
α-Proteobacteria
Actinobacteria
Firmicutes
Bacillus thuringnensis (AY138288)
0.1
TBTK040607A (LC057709)
TBTK040711B (LC057694)
TBTK040811B (LC057705) TBTK040707D (LC057693) TBTK040311B (LC034566)
TBTK040502C (LC057688) Bacillus altitudinis (FM955870)
TBTK040305D (LC057687) Bacillus subtilis KT1002 (AB115958)
TBTK040705A (LC057697) TBTK040405D (LC057701) TBTK040711A (LC057695)
TBTK040607B (LC057713) TBTK041005B (LC057710)
Exiguobacterium undae (NR043477) Paenibacillus sp. ZY-8 (FJ232954)
TBTK040404A (LC057714) TBTK040501B (LC057692)
TBTK040606B (LC057691)
Arthrobacter aurescens NI1 (JQ435703) TBTK040209C (LC057618)
Microbacterium oxydans M66_1A (JN644529) TBTK040502A (LC057706)
TBTK040913A (LC057704) Streptomyces sp. AC113 (FJ001763) Streptomyces badius partial (HF935087)
TBKT040402B (LC057702) TBTK040402D (LC057700)
Rhodobacter capsulatus SB 1003 (CP001312) Rhodobacter sp. SgZ-3 (JX113682) TBTK040607C (LC057690)
Rhizobium galegae (Z79620) Rhizobium cellulosilyticum (NR043985)
TBTK041010A (LC057703) Ralstonia eutropha H16 (AM260479) Cupriavidus necator N-1 (CP002877) TBTK040410B (LC057699)
Burkholderia grimmiae R27 (NR18268) TBTK040506C (LC057689)
Pseudomonas putida (AY958233) Pseudomonas fluorescens (EU439419)
TBTK040306C (LC057708) TBTK040703A (LC057698)
Acinetobacter johnsonii (AB099655) TBTK040312A (LC057686) TBTK040711C (LC057696)
Acinetobacter calcoaceticus (AY346313) Escherichia coli K-12 (CP010445)
TBTK040915A (LC057712) TBTK040915B (LC057711)
TBTK040502D (LC057707)