⽣分解性ポリエステルの環境分解性と材料寿命制御
群⾺⼤学⼤学院 理⼯学府 理⼯学専攻 物質・⽣命理⼯学領域 環境調和型材料科学研究室
鈴⽊ 美和
⽬次
第1章 序論
汎⽤性プラスチックが引き起こす海洋汚染
⽣分解性⾼分⼦とは
⽣分解性⾼分⼦の⽣物分解
⽣分解性⾼分⼦の酵素による分解
⽣分解性⾼分⼦の課題
芽胞形成細菌およびその応⽤例 本研究の⽬的
第2章 ポリエチレンスクシネートおよびポリ(3-ヒドロキシブタン酸) の環境分解性の差異
実験 試薬 環境試料
培地および培養条件 細菌株
⽣菌数およびポリマー分解菌数の測定 ポリマー分解細菌の単離
遺伝系統学的解析 基質分解試験
海⽔を微⽣物接種源としたポリマーの⽣物化学的酸素要求量(BOD)
⽣分解度測定 DNAアクセッション番号
結果
環境試料中の⽣菌数に対するP(3HB)およびPESu分解微⽣物の存在割合
海⽔中のP(3HB)およびPESuのBOD⽣分解度
単離されたP(3HB)およびPESu分解細菌の遺伝系統学的解析 考察
第3章 海洋性ポリエチレンスクシネートおよびポリ(e-カプロラクトン) 分解細菌の特徴付け
実験 試薬
PCL分解細菌の単離
5 6 8 13 15 18 20 22
23 24 24 24 24 25 25 25 25 26 26
26 27 27 29 30 32
35
36 36 37
キノン解析
グアニン-シトシン(GC)含率 脂肪酸メチルエステル(FAME)解析 リン脂質解析
⽣理的・⽣化学的解析 ポリエステル分解活性
⽣育およびポリエステル分解活性に塩濃度が与える影響
⽣育およびポリエステル分解活性に温度が与える影響 フィルム分解試験
表⾯形態観察
⽣育およびポリエステル分解活性に炭素源が与える影響 ザイモグラム
塩基配列アクセッション番号 結果
ポリエステル分解細菌,TKCM64株の特徴付け 遺伝系統学的解析
PCLおよびPESuのフィルム分解試験
上清のPCLおよびPESu分解活性と増殖に炭素源が与える影響 ザイモグラムによるPCL分解活性
考察
第4章 ⽣物製剤を利⽤した時限⽣分解性⾼分⼦の創製 実験
試薬
PESu分解芽胞形成細菌のスクリーニングおよび同定 PESu分解芽胞形成単離株の特徴付け
PESuフィルム分解試験
脂肪酸メチルエステル(FAME)解析
⽣理的・⽣化学的解析 ポリエステル分解活性
⽣育およびポリエステル分解活性に塩濃度が与える影響
⽣育およびポリエステル分解活性に温度が与える影響 表⾯形態観察
37 38 38 38 38 39 39 39 39 40 40 40 41 42 42 45 45 47 48 49
51 52 52 52 52 52 53 53 53 53 53 53
芽胞含有PESuフィルムの作成
培地中での芽胞含有PESuフィルム分解試験 実海⽔中での芽胞含有PESuフィルム分解試験
結果
PESu分解芽胞形成細菌の単離と特徴付け
YKCMOA1株の芽胞の特徴付け
培地中での芽胞含有PESuフィルムの分解試験 実海⽔中での芽胞含有PESuフィルムの分解試験
考察
第5章 総括
参考⽂献
54 54 54
55 56 58 58 59
61
63
68
第 1 章
序論
汎⽤性プラスチックが引き起こす海洋汚染
⽯油化学⼯業の勃興に伴いプラスチック⽣産は,1930年から1940年頃より,急速に拡⼤
し,その市場規模は今なお拡⼤を続けている[1,2]。2015年には,全世界で3億2200万トン のプラスチックが⽣産された [3]。プラスチックはその優れた物性および安定性から,様々 な製品の材料として⽤いられているが,他⽅,⼀旦環境に流出した際には,分解されないた めに,そこに留まり続ける[4–6] (Fig. 1)。プラスチックが原因の海洋汚染は,その⽣産が開 始されて以降,ここ40年以上もの間,報告され続けている[2,7–10]。特に,現代の海洋ゴミ の多くはプラスチック廃棄物で占められており,その割合は,60-80%と推定されている [11,12]。
Fig. 1. Plastic pollution in Kuju beach (Shimoda city, Shizuoka).
⾃然環境中に流出したプラスチックは,海洋⽣態系に様々な負の影響を及ぼすことが知 られている。例えば,海洋環境中でのプラスチック廃棄物の⽣物への誤飲や絡まりなどが挙 げられる [13,14]。Choyらは,プラスチックの⿂による誤飲量は,その種類によって差があ ったものの,平均すると19%の⿂にプラスチックが含まれていることを報告している [15]。
Bondらが,海⿃であるウミガラスおよびハシブトウミガラスのプラスチック摂取量を1985 年から2012年にかけて調べたところ,いずれの年代でも,⼀部の海⿃は体内にプラスチッ クを含んでおり,その割合は約7 %であることがわかった[16]。これらのプラスチックを誤 飲した⼩さい⿂や海⿃は,その後⾷べ物の取り込み量の制限および消化管内損傷を起こし,
死亡に⾄ることがある[10]。また,プラスチック表⾯は疎⽔性が⾼いため,環境中に希薄に 存在するヒトに対して発ガン性が認められる物質(ベンゾピレンを含む多環式芳⾹族化合 物(PAHs))を濃縮し吸着することがわかっている。このような汚染されたプラスチックを⽣
物が誤⾷することにより,⽣態系内において⽣物濃縮が⽣じる。最近になって,PAHs は,
⾷物連鎖を通じてヒトの体内を汚染している可能性が危惧されている[10,17–19]。ところで,
漂流する漁具は,海洋⽣物に絡まり、その⽣態系に深刻な影響を与えている[20] (Fig. 2,3)。
これらの漂流する漁具は,ゴーストネットと呼ばれる。⼀⽅これらと⽣物の絡み合いは,ゴ ーストフィッシングと呼ばれている。ゴーストフィッシングが⽣じる期間および被害をも たらす⽣物個体数は,Matsuokaらによれば,30から328⽇間であり,84から455個体の海 洋⽣物を死亡させると推測されている [21]。ゴーストネットに⽣物が絡むと,それは沈降 し,海底の⽣物にも影響を与える。さらに,海底において絡まった⽣物が完全に分解される と,再び浮上し,再度ゴーストフィッシングを続けると考えられている[22]。また,ゴース トフィッシングは,ゴーストネットの回収費⽤や,⽣物資源の損失を考慮すると,経済活動 に対しても少なからず影響を及ぼしている [23,24]。
Fig. 2. Derelict fishing gear in coral reef [25].
⽣分解性⾼分⼦とは
⽣分解性⾼分⼦は,⾃然環境中で微⽣物の作⽤により分解される環境分解性⾼分⼦と,ヒ トの体内の作⽤(代謝)により分解,吸収される⽣体内分解性⾼分⼦がある[27]。本博⼠論
⽂では,前者の⾃然環境中で⽣分解する⾼分⼦材料を⽣分解性⾼分⼦として扱う。
Nathalieらによれば,⽣分解性⾼分⼦の環境分解は以下の3つの過程を経て⽣じる(Fig. 4)
[28]。
Step 1:微⽣物および⾮⽣物因⼦(機械的破断,光分解,熱分解,酸化分解,加⽔分解,化 学分解)による,細かい断⽚化(⽣物劣化)。
Step 2:微⽣物が⽣産する触媒作⽤物質(酵素やフリーラジカル)によるポリマー主鎖の分 解と,それに伴うオリゴマー,ダイマーおよびモノマーの⽣成(脱重合)。
Step 3:微⽣物細胞表⾯のレセプターによる分⼦の認識および分⼦のペリプラズムへの移
⾏と,細胞質内での代謝を経由したエネルギー⽣産(異化),貯蔵物質の合成,⼀次および
⼆次代謝産物の⽣成(同化)。分⼦の完全酸化によって⽣じた,⼆酸化炭素,窒素,メタン,
⽔および塩の排出(無機化)
Fig. 4. Biodegradation mechanism of polymer [28].
Table 1 に,国際標準化機構(ISO)および経済協⼒開発機構(OECD)が制定した⾼分⼦材料
の⽣分解性評価⽅法およびその規格概要を⽰す。これらの規格は,主としてstep 3を定量的 に評価することにより成⽴している。
Table 1. Biodegradability test methods for plastics materials.
Standards Outline Media Evaluation criteria
ISO 14851 Determination of the ultimate aerobic biodegradability of plastic materials in an aqueous medium
Aqueous
Oxygen demand
ISO 14852 Carbon dioxide evolved
ISO 17556
Determination of the ultimate aerobic biodegradability of
plastic materials in soil Soil Carbon dioxide evolved
ISO 14855-1
Determination of the ultimate aerobic biodegradability of plastic materials under controlled composting conditions Part 1: General method
Compost Carbon dioxide evolved
ISO 14855-2
Determination of the ultimate aerobic biodegradability of plastic materials under controlled composting conditions Part 2:
Gravimetric measurement of carbon dioxide evolved in a laboratory scale test ISO 17556
Determination of ultimate aerobic biodegradability of
plastic materials in soil Soil Oxygen demand or carbon dioxide evolved
ISO 14853
Determination of ultimate aerobic biodegradation of plastic materials in an aqueous system
Aqueous
Anaerobic Biogas production
ISO 15985
Determination of the ultimate anaerobic biodegradation and disintegration under high-solids anaerobic- digestion conditions
Anaerobic Biogas production
ISO 13975
Determination of the ultimate anaerobic biodegradation of plastic materials in controlled slurry digestion systems
Anaerobic
Slurry digestion system Biogas production
ISO/DIS 18830
Test method for determining aerobic biodegradation of plastic materials sunk at the sea water/sandy sediment interface
Seawater
Sandy sediment interface Oxygen demand
ISO/DIS 19679
Determination of aerobic biodegradation of non- floating plastic materials in a seawater/ sediment interface
Seawater
Sandy sediment interface Carbon dioxide evolved OECD 301A
The screening of chemicals for ready biodegradability in
an aerobic aqueous medium Aerobic aqueous medium
Dissolved organic carbon
OECD 301B Carbon dioxide evolved
OECD 301C Oxygen consumption
OECD 301D Dissolved oxygen
OECD 301E Dissolved organic carbon
OECD 301 Oxygen consumption
⽣物化学的酸素要求量(BOD)は,以下の式(1)で表される。
BOD=被験物質を⽤いた試験酸素消費量−ブランク試験による酸素消費量
容器中の被験物質 (1)
ここでのThOD(理論的酸素要求量)とは,被験物質が完全酸化されるときに必要な理論的 酸素量を⽰す。
⼆酸化炭素(CO2)発⽣量は,以下の式(3)で表される。
CO2発⽣量 = 被験物質存在下でのCO2発⽣量-ブランク試験でのCO2発⽣量 (3)
⼆酸化炭素発⽣量を⽤いた⽣分解度は,以下の式(4)および(5)で表される。
ここでの ThCO2(理論的⼆酸化炭素発⽣量)は,被験物質が完全に酸化された時の理論的 CO2発⽣量,3.67は炭素を⼆酸化炭素に変換する係数,TOCは総有機炭素量を⽰している。
溶存有機炭素量(DOC)を⽤いた⽣分解度は,以下の式(6)で表される。
DOC(t), DOCbl(t), DOC(0)および DOCbl(0)は,それぞれ時間tおよび0における被験物質と 微⽣物を添加した分解媒体のDOC,時間tおよび0における微⽣物を添加した分解媒体の みのDOCを⽰している。
⽣物ガス発⽣量を⽤いた⽣分解度は,以下の式(7)で表される。
⽣分解度(%) = BOD
ThOD×100 (2)
生分解度(%) = CO2発生量
ThCO2×被験物質添加量×100 (4)
生分解度(%) = CO2発生量
3.67×試験で添加したTOC×100 (5)
生分解度(%) = (1- DOC(t)-DOCbl(t)
DOC)0*-DOCbl(0)+ ×100
(6)
CB, CBbおよびCはそれぞれ被験物質存在下でのガス化下炭素発⽣量,ブランク試験におけ るガス化した炭素発⽣量および被験物質に含まれている炭素量を⽰している。
⽇本国内においては⽇本バイオプラスチック協会(JBPA)が,Table 1に⽰すOECD 301C, ISO 14851, ISO 14852, ISO 17556, ISO-14855-1, ISO 14855-2に準拠し,対象とする⾼分⼦の⽣
分解度が,所定期間内に60 %以上であるものを,⽣分解性⾼分⼦と定めている [29]。
Fig. 5に,各⽣分解性⾼分⼦の名称および構造を⽰す。
Fig. 5. Chemical structures of biodegradable polyesters. (a): poly(3-hydroxy butyrate) [P(3HB)], (b): poly(ethylene succinate) (PESu), (c): poly(butylene succinate) (PBSu), (d): poly(butylene succinate-co-adipate) (PBSA), (e):
poly(butylene adipate-co-terephthalate) (PBAT), (f): poly(lactic acid) (PLA), (g):
poly(e-caprolactone) (PCL)
P(3HB)に代表される微⽣物産⽣脂肪族ポリエステル(polyhydroxyalkanoate: PHA)は,環境 中で過剰に炭素源が存在し,栄養源が抑制されているとき,多くの細菌の体内でエネルギー 源として貯蔵される。その蓄積率は,最⼤で,細菌の乾燥重量あたり90%にもなる。他⽅,
栄養源の供給が開始され,外部からの⼗分な炭素源が得られないとき,PHA は体内の分解 生分解度(%) =CB-CBb
C ×100 (7)
b=1.320 nm, c(繊維軸)=0.596 nmで,直⽅晶系,2分⼦がアンチパラレル鎖となり,充填さ れている。分⼦は,左巻きの 21ヘリックス構造をとる。その⾻格構造は左巻きゴーシュ,
左巻きゴーシュ,トランス,トランスである[33]。P(3HB)のガラス転移点は4 ˚C付近で,融
点は180 ˚C付近である。P(3HB)の⾮晶および結晶領域の密度はそれぞれ,1.26 g/cm3および
1.18 g/cm3である。P(3HB)の機械的特性(ヤング率および引張り強度)は,アイソタクチック
ポリプロピレン(PP)のものと類似している。しかし,P(3HB)の破断伸度は5 %であり,PPの もの(400 %)よりはるかに低く,P(3HB)は硬くて脆い材料として知られている[32]。
PESu およびPBSu などの,直鎖脂肪族カルボン⼆酸(DCA)と直鎖脂肪族ジオールからな る化学合成ポリエステル,ポリ(n-アルキレン n-カルボキシジオナート)(PAAD)の多くは,
⽣分解性を⽰す[34]。PESu および PBSu の融点は,低密度ポリエチレン(LDPE)のそれ(110
˚C)に近似しており,ガラス転移温度は,PE(-45 ˚C)とポリプロピレン(PP) (-5 ˚C)の間に位置 する。その密度は,ポリエチレンテレフタレート(PET)に近く1.25 g/m3であり,引っ張り強 度はPEとPPの値の間に位置する。PESuは,エチレングリコールとコハク酸との重縮合体 であり半結晶性の⾼分⼦である。PESuは,かつて昭和⾼分⼦株式会社(現 昭和電⼯株式 会社)から製品名Bionolle ♯6000として製造されていた[35]。また,⽇本触媒からはLunare SEという製品名で販売されていた。PESuの結晶は,a=0.760 nm, b=1.075 nm, c(繊維軸)
=0.833 nmで,直⽅晶系,4分⼦で1ユニットを取っており,その⾻格構造は,トランス,
右回りゴーシュ,トランス,左回りゴーシュである [36]。⼀⽅で,PESuを室温で延伸する と,全てトランス構造であるβ晶が観察された [37]。PESuの平衡融点,ガラス転移点,結 晶化温度, および熱分解温度は,それぞれ115.6, -11.5, 55, および413˚Cである。[36,37]。ヤ ング率は,約287 MPa[40],引張強度は,25.1 MPa[41], 弾性率は,402 MPa, 破断応⼒は,
33.2 MPa[42]である。破断伸度は,200~503%の間に位置する[35,41,42]。また,PAADの共重
合体も,⽣分解性⾼分⼦として知られている。例えば,ポリ(ブチレンスクシネート-co-アジ ペート)(PBSA)は,様々な環境下で⾼い⽣分解性を⽰す[43] 。
PBATは,その構造に芳⾹族を含むポリエステルで,現在Ecoflexという商品名で,BASF 社によって⽣産および販売されている。PBATの融点はPEと同等であり,機械的性質はPE より優れている[44]。⼀⽅で,PBATは,含まれるテレフタル酸の組成により,その⽣分解 速度が異なる[45]。
PLA は乳酸の重縮合あるいはラクチドの開環重合により得られる。このラクチドは,バ イオマスを原料とし微⽣物の発酵により得られることから,PLA はバイオマスプラスチッ クとしても知られている[46]。
PCL は,e-カプロラクトンを開環重合させることで,合成される。PCL の結晶は直⽅晶 系であり,構造の各軸は, a=0.7496 nm, b=0.4974 nm, c(繊維軸)=1.7297 nmである[47]。
ガラス転移点,融点および平衡融点はそれぞれ,-65 ~ -60,56 ~ 65および86.05 ˚Cである。
ヤング率は,210-440 MPa, 引張強度は,20.7-42 MPa, 破断伸度は, 300 ~ 1000 %である[48–
パッケージング[51],ドラックデリバリーシステム[52]など,様々なアプリケーションへの 応⽤が期待されている[53,54]。Table 2に各⽣分解性⾼分⼦の性質を⽰す。
Table 2. The properties of biodegradable polymers and commodity plastics.
Polymer Melting
temperature (°C) Glass-transition
temperature (°C) Tensile strength (MPa) Elongation at break (%) References
P(3HB) 180 4 43 5 [32]
PESu 104 -11.5 - -10 21-25 200-503 [35,38,41]
PBSu 114 -32 33 560 [35]
PBAT 110-115 - 36 820 [44]
PLA 120-170 55-56 53 10-100 [46]
PCL 56-65 -65 - -60 20.7 - 42 300 - 1000 [48]
HDPEa 120-140 -133 - -100 14.5-38 2.0-130 [55]
LDPEb 105-116 -125 40-78 90-800 [55]
PPc 160-176 -10 - -23 26-41.1 15-700 [55]
a: high density polyethylene, b: low density polyethylene, c: polypropylene
⽣分解性⾼分⼦の⽣物分解
Table 3に,⽣分解性⾼分⼦に対する環境とそれらの分解性を⽰す。⽣分解性⾼分⼦は,
その⼀次構造の違いにより環境分解性が異なる。⼀⽅で,P(3HB)のみは,環境の違いに依ら ず,優れた⽣分解性を⽰す[56–60]。
⽣分解性⾼分⼦の環境分解性は,その環境中の⽣分解性⾼分⼦分解微⽣物の存在割合と 相関がある[34,66]。Table 4 に,今までに⽣分解性⾼分⼦分解微⽣物として単離された株の 属種名,単離源およびそれらが分解する⾼分⼦を⽰す。
Table 3. Environmental degradabilities of biodegradable polymers.
Polymer Environmental degradability References
Excellent Depending on site Poor
PHA Soil
Freshwater Brackish water Seawater Aerobic sludge Anaerobic sludge Compost
- -
[56–60]
PESu Soil
Freshwater Compost Activated sludge
- Seawater
[35,61]
PBSu Compost Soil Seawater
Activated sludge
Freshwater [35,61]
PBAT Compost Soil Freshwater
Seawater [62–64]
PLA Compost Soil Seawater [62,64]
PCL Soil
Freshwater Seawater Compost
- -
[61,65]
Table 4. Biodegradable polymer-degrading microbes.
Strain Degrading polymers Source of isolates References
Bacteria Actinobacteria
Arthrobacter ilicis PHA Compost [67]
Clavibacter michiganense subsp PHA Freshwater [68]
Streptomyces
thermocarboxydovorans PHA Soil [69]
Proteobacteria a-proteobacteria
Ochrobactrum anthropi PHA Sludge [68]
b-proteobacteria
Acidovorax delafieldii PHA, PBSu Soil, Compost, Freshwater, Sludge [68]
Alcaligenes faecalis PHA Seawater [68]
Burkholderia vietnamiensis PBSu Soil [70]
Burkholderia cepaci PHA Sludge [68]
Comamonas testosteroni PHA Soil, Compost, Seawater [59,60,71]
Delftia acidovorans PHA Freshwater [72]
Duganella zoogloeoides PHA, PCL, PBSu Soil [70]
Leptothrix sp. PBSA, PESu, PCL Soil [73]
Ralstonia pickettii PHA, PCL Activate sludge [70]
Roseateles depolymerans PCL, PBSu Soil [70]
Variovorax paradoxus PHA, PCL Soil, Compost, Freshwater, Sludge [70]
Zoogloea ramigeru PHA Seawater [68]
Paucimonas lemoignei PHA Soil, Compost [74]
g-proteobacteria
Acinetobacter calcoaceticus PHA, PCL Soil [70]
Acinetobacter junii PCL, PBSu Soil [70]
Acinetobacter johnsonii PHA Seawater [68]
Marinobacter sp. NK-1 PHA Seawater [75]
Pseudoalteromonas haloplanktis PHA Compost, Freshwater, Seawater [76]
Pseudomonas alcaligenes PHA Compost [68]
Pseudomonas putida PHA Soil [68]
Pseudomonas stutzeri PHA Soil [68]
Stenotrophomonas maltophilia PHA Soil [68]
Serratia marcescens PCL [70]
Vibrio ordalii PHA Seawater [68]
Firmicutes
Clostridium acetobutylicum PHA, PCL Sewage sludge enrichment culture [57]
Amycolatopsis sp. HT-6 PHA, PCL, PBSu, Soil [77]
Amycolatopsis sp. HT-32 PCL,PLA Soil [78]
Microbispora rosea PCL,PBSu Cultured collection [79]
Excellospora japonica PBSu Cultured collection [79]
Excellospora viridilutea PBSu Cultured collection [79]
Bacillus sp. TT96 PHA, PCL, PBSu, PESu Soil [80]
Bacillus pumilus KT1012 PCL, PESu Freshwater [81]
Bacillus pumilus 1-A PBSA, PBSu, PCL Soil [82]
Bacillus megaterium PHA Soil, Compost, Freshwater [68]
Paenibacillus polymyxa PHA Soil [68]
Staphylococcus aureus PHA Freshwater [68]
Bacteroidetes
Flavobacterium johnsoniae PHA Soil, Compost, Freshwater, Seawater [68]
Fusobacteria
llyobacter delafieldii PHA Estuarine sediment [68]
Fungi
Acremonium sp PHA Soil [68]
Aspergillus sp. XH0501 PBSu Soil [83]
Aspergillus clavatus NKCM1003 PHA, PESu Terrestrial environment [84]
Aspergillus fungatus PHA Soil, Compost [68]
Aspergillus penicilloides PHA Soil [68]
Aspergillus versicplor PBSA, PBSu Compost [85]
Candida guiliermondii PHA Deep sea [86]
Paecilomyces marquandii PHA Soil [68]
Penicillium ochrochloron PHA Soil [68]
Verticillium leptobactrum PHA Soil [68]
⽣分解性⾼分⼦の酵素による分解
Table 4 に⽰した⽣分解性⾼分⼦分解微⽣物は,その菌体外に加⽔分解酵素を⽣産する
[87,88]。これらの酵素は,⽣分解性⾼分⼦を低分⼦量化させる[72,88]。
⽣分解性⾼分⼦のうち,微⽣物産⽣脂肪族ポリエステルであるP(3HB)およびその共重合 体は,P(3HB)分解酵素により加⽔分解される。これまでに,様々なP(3HB)分解酵素が精製 され,特徴付けされた。その結果,P(3HB)分解酵素は,以下の特徴を有することが明らかに なった[89]。
1. 幅広いpH,温度,イオン強度で安定。
2. 分⼦量は100kDa以下で,四次構造を形成しない。
3. 疎⽔性担体に強⼒に結合する。
4. ⼀般的には,⾄適pHが7.5から9.8である。
5. ⼀次構造は,活性残基を含む触媒ドメイン,P(3HB)表⾯への吸着ドメインおよびこれ らを連結するリンカードメインから構成されている。触媒ドメインは,活性残基とし て,Ser, HisおよびAspを有している。この活性残基であるSerはリパーゼボックス内 に存在する(Fig. 6)。
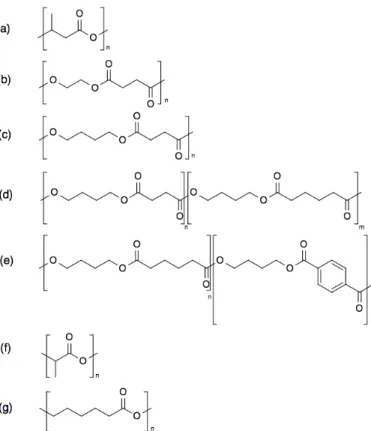
Penicillium funiculosum IFO6345のP(3HB)分解酵素および3HBの三量体との複合体の構造 解析が⾏われ,P(3HB)分解酵素は,基質結合部位として連続したモノマーユニットに対す る3つ以上の結合部位を持っていることが明らかになった。基質結合部位は,主に疎⽔的相 互作⽤により基質の側鎖と結合している。その⼀⽅で,親⽔的な相互作⽤によるエステル結 合の認識も⾏われている。これにより、主鎖および側鎖の炭素鎖⻑特異的および光学異性体 特異的な認識が⽣じる[92]。また,3種類の細菌由来のP(3HB)分解酵素の基質特異性を調べ たところ,P(3HB)分解酵素は本来の基質であるP(3HB)の他に,PHAの⼀種である正式名称 (P(3HP))お よ び 正 式 名 称(P(4HB))と , 化 学 脂 肪 族 ポ リ エ ス テ ル で あ る PESu お よ び poly(ethylene adipate)(PEA)を加⽔分解することが明らかになった[93]。P(3HB)分解酵素の結 晶構造解析および基質特異性試験の結果から,P(3HB)分解酵素は,基質となるポリエステ ルの側鎖がメチル基以下であり,2つのカルボニル基に挟まれる主鎖の原⼦数が、3あるい は4である場合に,その活性が⾒られると推測されている[92,93]。PESuではエチレングリ コール-コハク酸からなるユニットの 2量体が,3HBの 4量体の構造に類似しているため,
PESuはP(3HB)分解酵素により分解されると考えられる(Fig. 7)[94]。
Fig. 7. 3HB tetramer and Ethylene glycol - succinic acid (ESu) unit dimer [93].
⼀⽅,化学合成脂肪族ポリエステルは,⼈⼯的に製造されたものであり,天然に存在しな い。つまり,化学合成脂肪族ポリエステルを本来の基質とする酵素は存在しない。これに対 して,1977 年に,常盤らは,脂肪を分解する酵素であるリパーゼが種々の化学合成脂肪族 ポリエステルを加⽔分解することを⾒出した[95]。これ以降,化学合成脂肪族ポリエステル を加⽔分解できる酵素が,数多く発⾒され特徴付けられている(Table 5)。
Table 5. Substrate specificitiesand specific activitiesof purified lipases and esterase.
Enzyme Origin Degradable polymers References
Bacteria Actinobacteria
Cutinase Thermobifida fusca PBAT, PCL [96]
Esterase Thermobifida halotolerans PLA [97]
Proteobacteria b-proteobacteria
Esterase Roseateles depolymerans PESu, PCL, PBSu, PBSA, PBAT [98]
g-proteobacteria
Lipase Pseudomonas alcaligenes PCL, PESu [99]
Lipase Pseudomonas aeruginosa PCL [99]
Lipase Burkholderia cepacia PCL, PESu [99]
Esterase Pseudomonas fluorescens GK13 PCL [99]
Esterase Pseudomonas pseudoalcaligenes PBAT [100]
Lipase Pseudomonas sp. PBAT [101]
Firmicutes
Lipase Bacillus subtilis PCL, PESu
Cutinase Bacillus pumulus PBAT, PBSA, PBSu, PCL, PESu [102]
Lipase Pelosinus fermentans PBAT [103]
Fungi
Cutinase Cryptococcus magnus PBSA, PBSu, PCL, PLA [104]
Cutinase Fusarium solani PCL [88]
Cutinase Paraphoma related strain PBSu, PBSA, PBAT, PCL, PDLLA [105]
Cutinase Pseudozyma antarctica PBSu, PBSA, PCL, PLA [106]
多くの化学合成脂肪族ポリエステルは,リパーゼ,エステラーゼおよびクチナーゼにより 加⽔分解される(Table 5)。リパーゼ(トリアシルグリセロール アシルハイドロラーゼ, EC 3.1.1.3)とは,油と⽔の界⾯で,トリアシルグリセロールをグリセロールと脂肪酸に加⽔分解 させる加⽔分解酵素のクラスを指す[107]。クチナーゼ(EC 3.1.1.74)は,植物のクチクラ層に ある,ヒドロキシ脂肪酸およびエポキシ脂肪酸で構成されるポリエステルであるクチンを 加⽔分解する酵素である[108]。Liuらは,PCL分解活性を有する Aspergillus oryzae および
Fusarium solani 由来のクチナーゼの構造とその機能解析を⾏なった。A. oryzae のクチナー
ゼには,活性部位の周辺に,ヘキサン酸エステル部位が容易に近づくことができる⼤きな溝 が存在した。⼀⽅,F. solani のクチナーゼでは,2つのアミノ酸(81 位のロイシンおよび 184番のバリン)が障壁となり,ヘキサン酸エステルが活性部位に近づくことを妨げていた。
この基質に対する酵素の溝へのアクセスのしやすさが,ポリエステルの加⽔分解速度に⽐
例していた[109]。Paalらは,Pseudomonas pseudoalcaligenes由来のPBAT分解活性を有する 酵素(PpEst)をクローニングし,モデリングを⾏なった。その結果,PpEstの酵素活性部位に は,PBAT のモデル基質であるbis(4-(benzoyloxybutyl)terephthalateが⼊るポケットの存在が 確認できた[99]。これらのことから,化学合成脂肪族ポリエステルとトリアシルグリセロー ルやクチンとの間には構造類似性があり,各種のリパーゼやクチナーゼは,化学合成脂肪族 ポリエステルも基質として認識し,加⽔分解できると考えられる。また,各種酵素の分解す る⽣分解性⾼分⼦の種類は,それぞれ異なっている。
また,リパーゼは,アモルファスPHA を分解できたが[110],結晶性のPHA をまったく 分解できなかった[98,111]。
⽣分解性⾼分⼦の課題
現在利⽤可能な多くの⽣分解性⾼分⼦では,その⽣分解速度および分解開始時期の制御 が困難であり,このことが,⽣分解性⾼分⼦の普及を妨げている。理想的な⽣分解性⾼分⼦
は,使⽤期間中,機械的強度を保ち,不要時あるいは廃棄後に速やかに⽣分解されることが 望まれる。本博⼠論⽂では,分解開始時期および分解速度が⼗分に制御された⽣分解性⾼分
⼦を,時限⽣分解性⾼分⼦(timing biodegradable plastics)と定義づける(Fig. 8)。
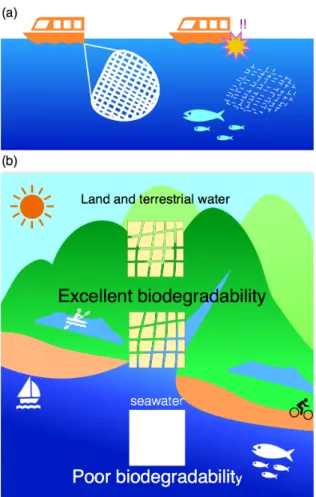
また,Table 3に⽰すように,化学合成⽣分解性⾼分⼦の⽣分解性は環境に依存しており,
このことも⽣分解性⾼分⼦の⽤途を狭める要因の⼀つとなっている。しかし⼀⽅で,このよ うな⽣分解性⾼分⼦の流出環境は多岐にわたるため,⽣分解性⾼分⼦の普及を想定するな ら,どのような環境においても分解される必要があるだろう。本博⼠論⽂では,このような 環境に応じて⽣分解速度が極端に異なる⽣分解性⾼分⼦を,潜在的⽣分解性⾼分⼦
(potentially biodegradable plastics)として定義付ける(Fig. 8)。
Fig. 8. Timing biodegradable and potentially biodegradable plastics. (a): timing biodegradable plastics. Mechanical properties of timing biodegradable plastics are kept during use, whereas they are biodegraded rapidly after use. (b): potentially biodegradable plastics in seawater. The potential-biodegradable plastics are never biodegraded in seawater in spite of the fact that they are
⽣分解性⾼分⼦の分解速度および分解開始時期を制御するために,これまで,様々な⽅法 が研究されてきた。
中温環境下(30 ˚C)でのPLAの加⽔分解速度は,極めて低い[112]。PLAの加⽔分解速度は,
環境温度をガラス転移温度以上にしたり, 環境pH を酸性あるいは塩基性にしたり,また は⾼温で加圧させることにより,上昇することが知られている[113,114] 。PBAT では,添 加剤の種類および UV 照射時間によって⽣分解速度を制御できる [115,116]。P(3HB)では,
ドデカノール,ラウリン酸,トリブチリンおよびトリラウリンの添加量により,その酵素加
⽔分解速度制御できる[117]。ポリ(ブチレンスクシネート-co-アジペート)(PBSA)の表⾯にマ ンノシルエリスリトール脂質(MEL)をコーティングさせることで,その⽣分解速度を低下さ せることができた。このMELはエタノールによってPBSAから除去でき,再度⽣分解速度 を上昇させることが可能であった[118]。また,⽣分解性⾼分⼦に,デンプンや酸化カルシウ ムを添加させることで,その⽣分解速度は上昇する[119–121]。
⼀⽅,⽣分解開始時期の制御⽅法として,Fukuda らは,ポリマー主鎖中に,容易に加⽔
分解されるイミン結合を導⼊することで,⽔を制御因⼦とした分解開始時期制御⽅法を開 発した[122]。
しかし,上記の研究例で⽰された分解速度および分解開始時期の制御⽅法は,実環境中での 分解速度制御には必ずしも適していない。特にこれらの⽅法では使⽤中あるいは保管中に 材料の物性が低下してしまうという⽋点から,⽣分解性⾼分⼦の時限分解性を確⽴できた とは⾔い難い。また,⾺場らは,ポリマー主鎖中に,還元条件下で開裂されるジスルフィド 結合を導⼊することで,海底や湖底といった還元環境を制御因⼦とした新しい時限⽣分解 性⾼分⼦創製の⼿法を提案している[123,124]。
芽胞形成細菌およびその応⽤例
芽胞(endospore)とは,ある種の細菌が,栄養制限下で形成する細胞形態を指す[125]。
芽胞を形成する好気性細菌として,Sporisarcina, Paenisporosarcina, Brevibacillus, Paenibacillus,
Geobacillus,および Bacillus 属が知られている[126]。芽胞を形成する嫌気性細菌として,
Acetonema, Pelosinus, Peptoclostridium, Gottschalkia, Ruminiclostridium, Lachnoclostridiumおよ
びClostridium属が知られている[127]。これらの芽胞形成細菌のすべては,Firmicutes⾨に属
している。芽胞は,代謝休眠状態にあり,環境抵抗性(極端な温度,乾燥,湿気,熱,UV,
酸化および電離放射線)を有している。芽胞の寿命は,数百から数千年と推定されている
[125,128]。もっとも研究が進んでいる芽胞形成細菌は,Bacillus sublitisであり,その芽胞形
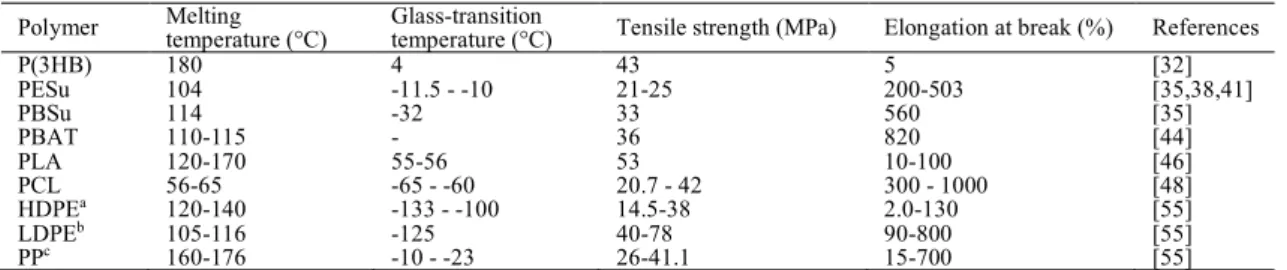
成プロセスは,以下に⽰す通りである。栄養制限下で成⻑が停⽌し,⾮対称の細胞分裂が開 始する。⼤きい細胞は⺟細胞,⼩さい細胞は前芽胞と呼ばれる。DNA分離が終了したのち,
⺟細胞は,前芽胞を吸い込む段階へと移る。芽胞の代謝休眠状態とDNAの圧縮が開始され る間,⺟細胞は,胞⼦⽪層および胞⼦の内膜と外膜の形成を媒介する。このプロセスが完了 すると,⺟細胞は溶解し,芽胞を放出する[125]。芽胞の構造は,外側から,外壁,外殻,外 膜,⽪層,発芽細胞壁,内膜および中核の順で構成されている。このように,芽胞は多層構 からなり,このことが⾼い環境抵抗性の原因の⼀つとなっている[129]。芽胞が発芽する際 は,発芽誘起条件が整う必要がある。⾃然環境中での芽胞発芽誘起物質は,特定の栄養素で ある。例えば,B. subtilisの芽胞においては,L-アラニン, L-バリン, L-アスパラギンが発芽 を誘引する[130]。これら誘起物質が芽胞内膜上のレセプターと結合し,発芽が始まる。その 後,芽胞のコアより H+, K+, Na+と共に芽胞特異的な Ca2+と pyridine-2,6-dicarboxylic acid (dipicolinic acid)のキレート(CaDPA)が放出される。その後,トランスグリコシラーゼによっ て,芽胞⽪層が分解され,その内部に⽔が取り込まれることにより,芽胞のコアが拡⼤する。
⽔はコア中での代謝を開始させる。コア内で代謝が始まると芽胞は増殖細胞として増殖す る[131,132](Fig. 9)。⼀⽅で、これらの芽胞形成細菌のうち B. cereus, B. licheniformis, B.
thuringiensisなどは,ヒトに対する病原菌としても知られている。これに対して乳業を始め
とする⾷品産業では,芽胞形成細菌および芽胞の混⼊や,バイオフィルム形成を阻⽌するた めに,様々な⼿法がとられている[126]。他⽅で,芽胞形成細菌の環境耐性を利⽤して,薬 品,⾷品製造および環境浄化への応⽤例がある。例えば,Bacillus clausiiの芽胞は,乳児下 痢症の薬品製剤として利⽤されている。我々の⾝近な⾷品である納⾖は,B. subtilis var. natto を⽤い,⼤⾖を発酵させることで製造されている。また,B. subtilis や B. megaterium, B.
licheniformis, Paenibacillus polymyxaの混合物は,⽔環境の浄化剤として台湾,USAおよびベ ルギーの企業から販売されている[133]。
Fig. 9. Life cycle of spore [131,132].
本研究の⽬的
⽣分解性⾼分⼦のさらなる普及のためには,分解開始時期および分解速度が⼗分に制御 された時限⽣分解性⾼分⼦の創製が必要不可⽋である。本博⼠論⽂では,時限⽣分解性⾼分
⼦を実現させるために,化学合成⽣分解性⾼分⼦であるポリエチレンスクシネート(PESu)に 注⽬した。PESuは,⼟壌,淡⽔中で優れた⽣分解性を⽰すが,⼀⽅で海⽔中での⽣分解速 度が極めて低い。このことは,PESuが海⽔中で潜在的⽣分解性⾼分⼦候補となりうること を⽰唆している。すなわち海⽔環境中でのPESuに対して,何らかの⽣分解性トリガーを与 えることができれば,時限⽣分解性⾼分⼦を創製できる可能性がある。⼀⽅で,PESu が,
海⽔中で⽣分解されない原因は明らかになっていない。
PESuはin vitroでP(3HB)分解微⽣物により⽣産されるP(3HB)分解酵素によって加⽔分解
される[93]。これはP(3HB)分解酵素がPESuの⼆量体をP(3HB)の4量体のアナログとして 認識することができるためである(Fig 7)。PESu とP(3HB)の双⽅が P(3HB)分解酵素によっ て分解されるにもかかわらず,P(3HB)が様々な条件下で優れた⽣分解性を⽰す⼀⽅で,PESu は海⽔中で⽣分解されない。このことは,PESuの環境分解性がP(3HB)分解酵素による酵素 分解性で説明できない。そこで,本博⼠論⽂第2章では,P(3HB)とPESuの環境分解性の差 異を理解し,PESuが海⽔中で⽣分解されない原因を明らかにするために,⼟壌,淡⽔およ び海⽔中のP(3HB)およびPESu分解菌の分布の相関およびP(3HB)およびPESuの海⽔環境 分解性を調べる。
PESuは,PCLをはじめとする他の化学合成⾼分⼦と同様,クチナーゼあるいはリパーゼ により加⽔分解されることが知られている[97,100]。Table 4に⽰すように,ポリエステル分 解細菌種の中には,PCLとPESuの双⽅を分解するものもいる。他⽅,PCLは様々な条件下 で,優れた⽣分解性を⽰すのとは対照的に,PESuは海⽔中で⽣分解されない[61,134]。そこ で,第3章では,PCLおよびPESuの分解活性を有する海洋性細菌に着⽬し,その詳細な特 徴づけを⾏うことで,海⽔中でPESu⽣分解速度が極めて遅い理由を議論する。
第4章では,第2章および第3章で得られた結果に基づいて,PESuを利⽤した時限⽣分 解性⾼分⼦の創製を検討する。本章では,PESuの⽣分解性トリガーとして芽胞形成細菌に 着⽬した。⼀般的に芽胞は,PESuの融点以上(104 ˚C)での耐熱性があり,そのため,PESuの 添加剤として使⽤できると期待される。そこでPESuを海⽔中で時限分解させるために,こ の芽胞を⽤いた時限⽣分解性⾼分⼦材料の構築を試みる。
第 2 章
ポリエチレンスクシネートおよびポリ(3-ヒドロキシ
ブタン酸)の環境分解性の差異
実験⽅法
試薬
ポリエチレンスクシネート (PESu) (Mn = 4.8×104, Mw/Mn = 1.9)は,株式会社⽇本触媒よ り提供された。ポリ(3-ヒドロキシブタン酸) (P(3HB)) ( Mn = 6.8×104, Mw/Mn = 2.6)は,三菱 ガス化学株式会社より提供された。⾼分⼦をクロロホルムに溶解させ,メタノールに加える ことにより再沈殿した。⾼分⼦フィルムは,ガラスシャーレをキャスト表⾯として⽤いたソ ルベントキャスト法で作成された。他の試薬は東京化成⼯業株式会社,和光純薬⼯業株式会 社,関東化学株式会社あるいはSigma-Aldrich Japanから購⼊された。これらの試薬は未精製 のまま⽤いられた。
環境試料
47 個の⼟壌試料は森(5 地点: location no. A, S, T, U, AO (Table 7)),校庭(1 地点: AX),
⼤学構内(7 地点: B, AV, BE, BF, BG, BH, BI),市街地(8 地点: E, F, G, H, I, J, K, V),河 原(2 地点: N,Y),底質(8 地点: M, Q, AE, AI, AN, BB, BC, BD),公園(5 地点: K, O, P, AP, AQ),畑(9 地点: Z, AF, AM, AR, AS, AT, AU, AY, AZ)および道路沿い(2 地点: AC, BA)から採取された。11 個の淡⽔試料は, 雪(1 地点: C),川(5 地点: L, R, X, AA, AD),
湖(2 地点: D, AB),池(2 地点: AG, AH)および噴⽔(1 地点: W)から採取された。10 個の海⽔試料は沿岸⽔域(10 地点: AJ, AK, AK’, AK’’ AL, AL’, BJ, BK, BL, BM)から採取 された。採取試料のその他の情報を,Table 7に記載した。
培地および培養条件
PESuを,ジクロロメタン(30 mL)に溶解した。この溶液を1Lのミネラル培地溶液に添加し,
TOMY ultrasonic disruptor UD-200を⽤い,乳化させた。ミネラル培地の試薬組成は以下の通 りである。4.6 g/LのKH2PO4, 11.6 g/LのNa2HPO4·12H2O, 0.5 g/LのMgSO4·7H2O,0.1 g/Lの FeCl3·6H2O,1.0 g/LのNH4Cl,0.1 g/LのYeast extractおよび 0.1 g/LのPlysurfを混合し,pH 7.0 に調整した。ジクロロメタンは,40 ˚Cで撹拌することで除去された。⼀⽅,P(3HB)粉末は 直接ミネラル培地に混合させた。Luria-Bertani (LB) 培地の試薬組成は以下の通りである。
10 g/Lのtryptone peptone,5.0 g/Lのyeast extractおよび5.0 g/LのNaClを混合し,pH 7.5に調整し た。培地を固化させるために,寒天(15 g/L)を培地に添加した。海⽔試料を試験に⽤いる 時は,培地のNaCl濃度を3%に調整した。培地は使⽤前に,121 ˚Cで15分間オートクレーブ された[81]。
細菌株
本研究で標準株として使⽤された細菌をTable 6に⽰す。
Table 6. Bacterial strains used in this study.
Strain Culture collection nos. Relevant characteristics References/Source
Escherichia coli DH5α
F−, Φ80d lacZΔM15, Δ(lacZYA- argF)U169, deoR, recA1, endA1, hsdR17(rK- mK+), phoA, supE44, λ−, thi-1, gyrA96, relA1
TaKaRa
Acidovorax delafieldii DSM 64 Reference strain [135]
Alcanivorax sp. NBRC 102022 Reference strain [136]
Bacillus megaterium NBRC 15308 Reference strain [137]
Microbacterium flavum NBRC 103923 Reference strain [138]
Microbispora rosea NBRC 14044 Reference strain [139]
Pseudomonas aeruginosa PAO1 DSM 1707 Reference strain [140]
Shewanella putrefaciens NBRC 3908 Reference strain [141]
Comamonas testrosteroni YM1004 JCM 10170 P(3HB)-degrading bacteria [142]
Delftia acidovorans YM1609 JCM 10172 P(3HB)-degrading bacteria [72,143]
Ralstonia pickettii T1 JCM 10169 P(3HB)-degrading bacteria [144]
Shewanella sp. JKCMAJ-6,1A - P(3HB)-degrading bacteria [89]
⽣菌数およびポリマー分解菌数の測定
環境試料中の⽣菌数とポリマー分解菌数を,それぞれプレート計数法とクリアゾーン法 で計数した[134]。1 gの環境試料を10 mLの滅菌⽣理⾷塩⽔に分散させ,10分間静置した。こ の上清から10-1から10-5の濃度の希釈溶液を作製した。各濃度の希釈液(50 µL)を固体のLB培 地およびポリマー含有培地に塗布した。⽔試料は,希釈せずに原液のまま培地に塗布され た。これらの固体培地をプラスチックテープで密封し,30 ˚Cで1 週間培養した。LB培地上 に形成されたコロニーおよびポリマー含有培地上に形成されたクリアゾーンを計測し,そ の数をそれぞれ環境試料1 gあたりのコロニー出現数 (c.f.u.) およびクリアゾーン形成数 (cz.f.u.)で表した。
ポリマー分解細菌の単離
PESuおよびP(3HB)分解細菌は環境試料から,それぞれのポリマーを含む固体培地を⽤い たクリアゾーン法によって単離された。P(3HB)含有培地上でP(3HB)分解活性によりスクリ ーニングされた細菌をP(3HB)分解細菌と,PESu含有培地上でPESu分解活性によりスクリー ニングされた細菌をPESu分解細菌と定義した。ポリマー含有培地上にクリアゾーンを形成 したコロニーをLB培地上に画線し,培養することで純化した。単離株の純度は,ポリマー 含有培地とLB培地の双⽅を⽤い確認された。
遺伝系統学的解析
16S rRNA遺伝⼦はポリメラーゼ連鎖反応(PCR)法で増幅された。Forwardプライマーは 16Sf : 5'-GTTTGATCATGGCTCAG-3' (⼤腸菌の16S rDNA塩基配列の36-53に該当)および27f:
5' -AGAGTTTGATCCTGGCTCAG-3' (8-27)で あ っ た 。Reverseプ ラ イ マ ー は16Sr: 5' - TACCTTGTTACGACTTCA-3' (1517-1533)お よ び1525r: 5'-AAAGGAGGTGATCCAGCC-3' (1543-1525)であった。反応は, 1.25 U ExTaq DNA polymerase (TaKaRa),100nM 各primer (16Sf,
のアニーリング,72 ˚Cで5分間の伸⻑を25サイクル⾏い,72 ˚Cで10分間保温した。PCR⽣成 物は1.5 % (wt./vol.)のアガロースゲルを⽤いた電気泳動により解析され,その後,pMD 20 (TaKaRa)に連結された。16S rDNA配列はダイデオキシ法で決定され,NCBIサイト上のプロ グラムblast (http://www.ncbi.nlm.nih.gov/BLAST/)を⽤いGenBankデータと⽐較された。16S rDNA塩基配列を,DDBJサイト上のプログラムClustalW (http://clustalw.ddbj.nig.ac.jp) を⽤い アライメントした。系統樹はプログラムMEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods [145]を⽤
い,近隣結合法[146]により構築された。
基質分解試験
単離株のPESuおよびP(3HB)加⽔分解能⼒はクリアゾーン法で調べられた。固体のポリマ ー含有培地に細菌を植菌し,30 ˚Cで1週間培養した。加⽔分解活性はコロニーの中⼼からク リアゾーンの縁までの距離で評価された。
海⽔を微⽣物接種源としたポリマーの⽣物化学的酸素要求量 (BOD) ⽣分解度測定
3 種類の海⽔(⼤洗(location no. BJ),新潟(BK),熱海(AL))を,BOD⽣分解試験 の微⽣物接種源として⽤いた。全ての試験は,好気条件下BOD reactor (Taitec)内で⾏われた。
フィルム試料(1 x 1 cm,約5 mg)および197.4 mLの海⽔を300 mLのBOD反応容器に⼊れた。
さらに,2.6 mLのミネラル溶液を反応容器に加えた。ミネラル溶液の試薬組成は以下の通り
である。0.085 g/LのKH2PO4, 0.2175 g/LのK2HPO4, 0.446 g/LのNa2HPO42H2O, 0.017 g/LのNH4Cl, 0.225 g/LのMgSO4.7H2O, 0.275 g/LのCaCl2および0.0025 g/LのFeC13.6H2Oを蒸留⽔に溶解した。
⽣分解試験は,25 ˚Cで28⽇間⾏われ,BODデータは⼀⽇ごとに計測された。ポリマー試料 のBOD⽣分解度は,試験溶液のBOD値 (BODt)からブランクコントロール値(BODb)を引 き,その値(BODt-BODb)を試験試料の理論的酸素要求量で割り,算出された。
DNAアクセッション番号
単離された株の16S rRNA遺伝⼦の塩基配列はDDBJ nucleotide sequence datebaseに登録さ れた。Fig. 12に,各株のアクセッション番号を⽰す。
結果
環境試料中の⽣菌数に対するP(3HB)およびPESu分解微⽣物の存在割合
68 個の環境試料中の,P(3HB)およびPESu分解微⽣物を計数した(Table 7)。Fig. 10は,各 環境試料中の⽣菌数とP(3HB)およびPESu分解微⽣物数との相関を⽰している。
Table 7. Samples aspect
No. North latitude East longitude Sampling site Sample aspect Viable microorganism counts
(c.f.u./g)
P(3HB)- degrading microorganism counts (cz.f.u./g)
PESu-degrading microorganism counts (cz.f.u./g)
A 36°28'43.83" 139°24'50.14" Gunma, Kiryu Forest soil 5.7×106 1.2×106 2.8×105
B 36°25'23.99" 139°20'50.52" Gunma, Kiryu University soil 1.6×106 3.0×105 1.4×105
C 36°25'23.99" 139°20'50.52" Gunma, Kiryu University snow 9.6×103 1.6×103 6.0×102
D 36°28'43.83" 139°24'50.14" Gunma, Kiryu Lake water 2.2×104 4.4×103 1.4×102
E 35°41'53.01" 139°46'26.11" Tokyo, Akihabara
City area soil 3.5×106 8.0×105 2.2×105
F 35°38'26.54" 139°17'36.11" Tokyo, Hachiouji City area soil 4.6×106 1.1×106 2.4×105 G 35°37'44.78" 139°44'14.21" Tokyo,
Shinagawa
City area soil 2.0×107 9.3×105 1.2×105
H 35°41'30.70" 139°46'17.37" Tokyo, Kanda City area soil 2.0×107 1.0×106 1.1×105
I 35°40'58.44" 139°45'53.59" Tokyo, Tokyo City area soil 3.1×107 9.3×105 6.6×104
J 35°39'22.35" 139°45'29.41" Tokyo, Hamamatsutyou
City area soil 3.5×107 9.4×105 1.5×105
K 35°42'58.03" 139°46'25.27" Tokyo, Ueno Park sand 1.3×107 1.0×106 8.6×104
L 36°20'4.91" 139°26'22.03" Tochigi, Ashikaga
River water 3.3×104 8.0×102 5.2×101
M 36°20'4.91" 139°26'22.03" Tochigi, Ashikaga
Sludge 1.3×105 3.3×104 3.3×103
N 36°20'4.91" 139°26'22.03" Tochigi, Ashikaga
River side soil 4.8×106 1.4×106 1.4×105
O 36°20'19.88" 139°26'41.33" Tochigi, Ashikaga
Park sand 9.3×106 1.7×106 9.3×104
P 36°20'19.88" 139°26'41.33" Tochigi, Ashikaga
Park soil 5.3×106 1.6×106 4.0×104
Q 36°26'58.03" 139°22'6.39" Gunma, Kiryu Sludge 7.1×106 2.1×106 2.3×105
R 36°26'58.03" 139°22'6.39" Gunma, Kiryu River water 5.8×104 1.0×103 1.3×102
S 36°28'17.87" 139°24'2.52" Gunma, Kiryu Forest soil 3.3×106 1.1×106 1.6×105
T 36°28'27.85" 139°24'8.50" Gunma, Kiryu Forest soil 3.6×106 6.0×105 1.1×105
U 36°28'19.13" 139°24'1.02" Gunma, Kiryu Forest soil 9.3×106 6.0×105 1.1×105
V 35°10'13.56" 136°52'49.50" Aichi, Nagoya City area soil 2.1×107 9.3×105 6.0×105
W 35°10'13.56" 136°52'49.50" Aichi, Nagoya Fountain water 3.1×105 6.0×103 9.3×101
X 36°29'23.58" 139°24'46.86" Gunma, Kiryu River water 5.4×103 8.1×102 8.0×102
Y 36°29'23.58" 139°24'46.86" Gunma, Kiryu River side sand 4.5×106 9.3×105 1.5×105
Z 36°31'55.95" 139°0'40.01" Gunma, Shibukawa
Field soil 1.2×107 2.1×106 3.1×105
AA 36°31'55.95" 139°0'40.01" Gunma, Shibukawa
River water 2.3×102 5.7×101 3.4×10
AB 36°23'50.26" 139°17'13.07" Gunma, Midori Lake water 1.1×103 1.3×102 4.0×10
AC 36°26'18.11" 139°10'35.62" Gunma, Maebashi
Roadside soil 5.9×106 6.7×105 1.6×105
AD 36°23'17.15" 139°9'11.18" Gunma, Maebashi
River water 1.3×103 3.3×102 5.3×101
AE 36°23'17.15" 139°9'11.18" Gunma, Maebashi
Sludge 2.9×106 6.7×105 2.9×105
AF 36°23'17.82" 139°9'9.96" Gunma, Maebashi
Field soil 2.2×106 5.9×105 1.1×105
AG 36°23'40.64" 139°18'19.90" Gunma, Midori Pond water 3.3×103 6.7×102 2.3×104
AH 36°23'37.83" 139°18'20.47" Gunma, Midori Pond water 3.2×105 1.9×103 1.6×102
AI 36°23'37.94", 139°18'18.44" Gunma, Midori Sludge 1.5×106 2.7×105 2.5×105
AJ 34°52'9.84" 138°19'30.41" Shizuoka, Yaizu Coastal seawater 1.6×104 5.4×102 0 AK 35°6'44.58" 138°35'8.43" Shizuoka,
Kanbara
Coastal seawater 5.9×103 1.4×101 0
AK’ 35°6'44.58" 138°35'8.43" Shizuoka,
Kanbara Coastal seawater 1.3×104 2.0×102 0
AK’
’
35°6'44.58" 138°35'8.43" Shizuoka, Kanbara
Coastal seawater 1.7×104 4.3×101 0
AL 35°5'53.03" 139°4'38.06" Shizuoka, Atami Coastal seawater 1.7×102 4.8×101 0
AL’ 35°5'53.03" 139°4'38.06" Shizuoka, Atami Coastal seawater 1.3×102 5.7×101 0
AM 36°18'50.19” 139°26'14.94” Tochigi, Field soil 1.4×106 1.2×104 4.6×103

![Fig. 7. 3HB tetramer and Ethylene glycol - succinic acid (ESu) unit dimer [93].](https://thumb-ap.123doks.com/thumbv2/123deta/6305904.1096289/16.892.256.637.569.820/fig-tetramer-ethylene-glycol-succinic-acid-esu-dimer.webp)
![Fig. 9. Life cycle of spore [131,132].](https://thumb-ap.123doks.com/thumbv2/123deta/6305904.1096289/21.892.262.637.154.533/fig-life-cycle-of-spore.webp)