九州大学学術情報リポジトリ
Kyushu University Institutional Repository
過剰歯幹細胞の再生医療学的応用に関する研究
牧野, 友祐
九州大学大学院歯学府
https://doi.org/10.15017/21979
出版情報:Kyushu University, 2011, 博士(歯学), 課程博士 バージョン:
権利関係:
§4. 研究Ⅱ
「過剰歯幹細胞移植による免疫療法の解析」
§4-1. 目的
現在、難治性疾患である全身性エリテマトーデス、SLEの治療に対し、骨髄間葉系 幹細胞の応用が期待されている。研究Ⅱの目的は過剰歯幹細胞が有する免疫調節能に 注目し、この機能を応用したin vivo細胞療法への可能性を検討することである。こ の目的を達成するために、ヒトSLEモデルマウスMRL/lprマウスに過剰歯幹細胞を 経静脈的に全身投与し、特徴的なヒトSLE様症状に対する治療学的効果を検討した。
また、細胞移植した過剰歯幹細胞によるレシピエントでの免疫抑制効果を検討するた めに、レシピエント骨髄間葉系幹細胞の組織構築能に注目し解析を行った。さらにT 細胞養子移植法やレシピエント骨髄間葉系幹細胞移入法を併用し、過剰歯幹細胞の全 身性細胞移植療法の治療メカニズムの解明を試みた。
4-2. 材料と方法
4-2-1. MRL/lprマウスへのSupSC細胞移植
16週齢雌性MRL/lprマウスに、体重10g当り0.1×106のSupSCまたはhBMMSC を尾静脈より全身投与した。同週齢のMRL/lprマウスにPBSを注入したものを対照 群とした。マウスを死亡するまで日々観察した。また、20週齢時にマウスを屠殺し、
末梢血、尿、腎臓、脾臓、腋下リンパ節を採取した。
4-2-2. 末梢血中、尿中、組織サンプル中の自己抗体、免疫グロブリン、生体マーカー、
サイトカインの測定
マウス後眼窩静脈叢から末梢血を採取し、遠心分離法で血清を得た。組織サンプル からM-PER Mammalian Protein Extraction Reagent (Thermo Scientific, South
Logan, UT) を用いて総タンパクを抽出した。尿は遠心後、上清を使用した。抗dsDNA IgG抗体、抗dsDNA IgM抗体、抗核抗体 (anti nuclear antigen, ANA)、補体第3 成分 (complementary 3, C3)、IL-10、IL-17、IFNγの測定には、Mouse anti-dsDNA IgG-specific ELISA Kit、Mouse anti-dsDNA IgM-specific ELISA Kit、Mouse Anti-Nuclear Antigens (ANA/ENA) Ig's (total (A+G+M))、Mouse C3 ELISA Kit、 Mouse Albumin ELISA Kit (alpha diagnostic, San Antonio, TX)、Mouse IL-10 Quantikine ELISA KitおよびMouse IL-17 Quantikine ELISA Kit、Mouse IFNγ Quantikine ELISA Kit (R&D Systems)を使用した。クレアチニンレベルはcreatinin parameter assay kit (R&D Systems)を、尿タンパク濃度測定にはBio-Rad Protein
Assay (Bio-Rad)を使用した。免疫グロブリンの測定は、予め各抗マウスイムノグロ
ブリン抗体 (各10 µg/ml, Invitrogen)でコートしたwellを作製し、Mouse Mono-AB ID/SP kit (Invitrogen)を使用して行った。
4-2-3. マウス腎の組織学的解析
腎組織を4%PFA含有PBS (pH7.4)にて4℃、一晩浸漬固定した。通法通りパラフ ィンに包埋した。パラフィン切片を作製し、H&E染色、Gomorriトリクローム染色、
Schiff過ヨウ素酸 (Periodic Acid Schiff; PAS) 染色を行った。一部の組織から凍結切 片を作製しマウスの IgG 抗体を用いた免疫蛍光染色に使用した。細胞核の染色には DAPI核染色を行った。
4-2-4. マウス末梢血中のTh17細胞、Th1細胞、Tregの解析
末梢血中の Th17 およびTh1 細胞を測定するために 、マウスPBMNC をPerCP 標識抗マウス CD4 抗体、FITC 標識抗マウス CD8a 抗体と反応させた後、Foxp3 staining buffer kit (eBioscience) を使用してR-PE標識抗マウスIL-17抗体、APC 標識抗マウスIFNγ抗体で染色した。また、マウスPBMNCをPerCP標識抗マウス CD4抗体、FITC 標識抗マウスCD8a抗体、APC標識抗マウスCD25抗体で染色し た後、Foxp3 staining buffer kit (eBioscience) を用いてR-PE標識抗マウスFoxp3 抗体で染色した試料を Treg の解析に使用した。フローサイトメトリーの解析には FACSCalibur (BD Bioscience) を用いた。
4-2-5. in vivoトレーシング解析
移植細胞をマウス体内でトレーシングするために、carboxyfluorescein diacetate succinimidyl ester (CFSE)を 用 い た 。 細 胞 (10 106/ml)を 2 mmol/ml CFSE (Dojindo)と37℃、10分間反応させた。CFSE標識細胞 (1×106) をMRL/lprマウス 尾静脈から注入し、注入後24時間または1週間後、マウスを屠殺し、腎臓、脾臓、
骨を採取した。摘出組織を 4%PFA 含有 PBS 溶液で固定し、凍結切片を作製後に DAPI染色を行った。
4-2-6. マウスPanT細胞の単離と培養
MRL/lprマウス脾臓をすり潰し、0.82% NH4Cl含有PBS溶液を室温、15分間作用 させて、赤血球を溶解除去し、脾細胞を得た。PanT cell isolation kit (Miltenyi Biotec) を使用し、脾臓細胞からマウスのPanT細胞を磁気分離した。PanT細胞 (1×106/well)
を、抗CD3抗体 (5 µg/ml, eBiosicence, San Diego, CA) でコーティングした24ウ ェルプレートにて、抗CD28抗体 (1 µg/ml, eBiosicence)とともに完全培地中で3日 間培養した。浮遊細胞および培養上清を回収し解析に使用した。
4-2-7. 骨のマイクロ−コンピューター断層解析
マイクロCT、Skyscan 1076 scanner (Skyscan, Kontich, Belgium)を使用して、マ ウ ス 大 腿 骨 遠 位 端 の マ イ ク ロ CT 撮 影 を 行 い 、InstaRecon/NRecon software (Skyscan)上でデータの 3 次元画像構築を行った。3-dimensional image analysis system CT Analyzer software (Skyscan)にて、骨密度 (bone mineral density, BMD) およびbone volume/trabecular volume [BV/TV]、trabecular thickness [Tb.Th]、 trabecular number [Tb.N]、trabecular separation [Tb.Sp、trabecular space [Tb.Spac] 等の各種骨パラメーターを計測した。骨密度 (bone mineral density,
BMD) の標準として、2種類のハイドロキシアパタイトファントム(ハイドロキシア
パタイト密度0.25および0.75 g/cm3, Skyscan) を用いた。
4-2-8. 骨の組織学的解析
脛骨を4%PFAで固定し、10%EDTAで脱灰した後、パラフィンに包埋した。パラ フィン切片を作製し H&E 染色を行った。また、酒石酸耐性酸ホスファターゼ (Tartrate-resistant acid phosphate, TRAP) 染色を行い、骨幹端におけるTRAP陽 性細胞の定量を行なった (Miura et al., 2006; Yamaza et al., 2009)。具体的には、各 実験群から5つの画像を任意に選択し、NIH イメージソフトウェア Image-Jを使用 して TRAP陽性細胞を計測し、全骨組織領域に対する TRAP陽性細胞の数として示
した。
4-2-9. 血中マーカーによる破骨細胞機能の解析
マウス後眼窩静脈叢から末梢血を採取し、遠心分離法で血清を得た。Mouse TRANCE/RANKL/TNFSF11 Quantikine ELISA Kit (R&D Systems) お よ び RatLAps ELISA kit (Nordic Bioscience Diagnostics A/S)を用いて、soluble receptor activator for nuclear factor κB ligand (sRANKL)およびI型コラーゲンC末端テロ ぺプチド(C-terminal telopeptides of type I collagen; CTX)の血中濃度を測定した。
4-2-10. in vitro破骨細胞の解析
マウス大腿骨および脛骨から骨髄細胞を採取した。骨髄液を 0.82% NH4Cl 溶液で 15 分間、室温で溶血させ、骨髄有核細胞を得た。骨髄有核細胞 (1 106/well) は 10 ng/ml macrophage colony stimulating factor (R&D Systems)および 25 ng/ml sRANKL (Peprotech) を加えた10%FBS含有αMEMで5~6日間培養した。その後 Acid Phospatase, Leukocyte (TRAP) kit (Sigma)を用いてTRAP染色し、TRAP陽 性多核細胞 (3核以上)を破骨細胞様細胞として計測した。また破骨細胞特異的遺伝子 解析のために培養細胞からtoral RNAも抽出した。
4-2-11. 骨芽細胞および破骨細胞特異的遺伝子の発現解析
SV total RNA isolation kit (Promega)を用いてサンプルからtotal RNAを抽出した。
DNaseⅠ処理後、SuperscriptⅢ (Invitrogen)を使用しcDNAを合成した。各遺伝子 特異的プライマーペアとPlatinum PCR supermix (Invitrogen)を混合し、PCRを行
った。本実験で使用したヒト特異的 PCR プライマーペアの配列は章末の表 4 に記載し た。
4-2-12. マウスBMMSCの単離と培養
マウスBMMSCの単離と培養は過去の報告 (Miura et al., 2006; Yamaza et al., 2009; 2010) を参考にした。マウス大腿骨および脛骨から全有核細胞 (all nuclear cells; ANC) を採取した。ANCを100 mm培養ディッシュに10~20×106で播種し、
37℃、5% CO2条件下にて3時間培養した。培養ディッシュをPBSにて2回洗浄し、
非付着性の細胞を除去した。その後、付着性細胞を14~16日間培養し、付着性コロ ニーを形成させた。コロニー形成細胞を一度継代し、増殖培養させた細胞をマウス BMMSCとして実験に使用した。マウス増殖培地として、20% FBS (Equitech Bio) お よ び 2 mM L-glutamine (Nakarai Tesque) 、55 µM 2-mercaptoethanol (Invitrogen)、100 U/ml penicillinおよび100 µg/ml streptomycin (Nakarai Tesque) を添加したα-MEM (Invitrogen)を使用した。フローサイトメトリー法で単離培養細 胞でのMSCマーカーの発現を解析した。単離培養細胞は、CD14, CD34, CD45に対 しては陰性を、CD73, CD105, CD146, Sca-1, SSEA-4に陽性反応を示し (データ未 提)、単離細胞がMSCであることが確認された (Yamaza et al., 2011)。
4-2-13. CFU-Fの解析
1フラスコ当り1.5×106個のANCをT-25フラスコ (Nunc) に播種した。3時間後 フラスコをPBSで洗浄し、上記培地にて16日間培養を行った。PBSで洗浄後、2%
PFAおよび1%トルイジンブルー含有溶液で培養細胞を染色した。光学顕微鏡下で50
個以上の細胞を含む集団をコロニーと見なし、1フラスコ当りの全コロニー数を計測 した。
4-2-14. マウスBMMSC増殖能の解析
BMMSCを1ウェル当り10×103個の濃度で8-well chamber slides (Nunc)に播種 し、2~3日間培養した。BrdU溶液 (1:100) (Invitrogen)を培地に添加し、20時間後 BrdU staining kit (Invitrogen)を用いて染色を行った。その後、ヘマトキシリン染色 を行った。BrdU陽性細胞と全細胞の数を計測し、全細胞数あたりのBrdU陽性細胞
数の割合 (BrdU陽性細胞数/全細胞数の百分率) を算出した。
4-2-15. colony forming unit-osteoblasts (CFU-OB) の解析
マウスCFU-F形成後、マウス増殖培地に2 mM β-glycerophosphate (Sigma)、100 µM L-ascorbic acid 2-phosphate (Wako Pure Chemicals)、10 nM デキサメタゾン
(Sigma)を添加した硬組織誘導培地を用いて CFU-F コロニーを7日間培養した。そ
の後アルカリフォスファターゼ染色を施した(Yamaza et al., 2010)。光学顕微鏡下 で 50 個以上の細胞を含む集団をコロニーと見なし、ALP 陽性コロニーを CFU-OB とした。ALP陽性コロニーと全コロニーの数を計測し、CFU-OB割合 (CFU-OB数
/全コロニー数の百分率) を算出した。
4-2-16. in vitro骨形成能の解析
マウスBMMSC を硬組織誘導条件下にて培養を行った。培養1週後、ALP染色と
ALP活性試験を行った (Yamaza et al., 2010)。誘導2週後、培養細胞からtotal RNA
を単離し、遺伝子発現解析を行った。誘導 4 週後、培養群を 1%アリザンレッド
(Sigma)にて染色した。石灰化領域を NIH イメージソフト Image-J 上で計測し、ア
リザンレッド陽性エリアの割合 (アリザンレッド陽性面積/全面積の百分率) を算出 した (Miura et., 2006; Yamaza et al., 2010)。
4-2-17. in vivo骨形成能および骨髄ニッチ形成能の解析
過去の論文 (Miura et al., 2006; Yamaza et al., 2008; 2009) を参考に、マウス BMMSC のin vivo骨形成能および骨髄ニッチ形成能を検索した。4.0×106のマウス BMMSC を、40 mgのHA/TCPキャリアー (Zimmer)として混和し、8~10週齢の 免疫不全マウスBalb/c nu/nuマウスの背部皮下に移植した。移植8週後、移植体を 回収し、4%PFA で固定した。10%EDTA (pH7.4)溶液で脱灰し、パラフィン切片作 製後、H&E染色を行った。各実験群から7画像を任意に選択し、形成された骨組織 とニッチの面積をNIHイメージソフトウェアImage-Jで計測し、全組織エリアに対 する骨またはニッチエリアの割合(骨組織面積/全面積またはニッチ面積/全面積の 百分率)として算出した (Miura et., 2006; Yamaza et al., 2010)。
4-2-18. マウス造血性コロニー形成細胞 (hematopoietic colony forming cell; CFC) の解析
Miura et al. (2006) の報告を基に CFC 解析を実施した。メチルセルロース培地 (MethoCult GF M3444, Stem Cell Technologies)を使用し、正常マウス由来全骨髄細 胞とレシピエントMRL/lprマウス由来BMMSCとの共培養実験を行った。1×104細 胞を2% FBS含有Iscove’s modified Dulbecco’s medium (Stem Cell Technologies)
100µlで懸濁し、メチルセルロース培地1 mlで混和後、35 mm dishに細胞を播種、
培養した。培養12~14日後、顕微鏡下でコロニー数を計測した。
4-2-19. マウスT細胞の細胞増殖能の解析
過去の報告 (Zhang et al., 2009; Yamaza et al., 2011) を参考に、マウス脾臓細胞 と マ ウ ス BMMSC と の 共 培 養 実 験 を 行 っ た 。BMMSC を 0, 2×103, 20×103, 200×103/wellで播種し、一晩培養した。200×103 cells/wellのマウス脾臓細胞を播き、
PHA (5 μg/ml)を添加した完全培地で培養した。72時間後、浮遊細胞を回収し、Cell Counting Kit-8 (Dojindo)を用いて生存細胞を測定した。
4-2-20. in vitroマウスTh17細胞の分化能の解析
Mouse CD4+CD25+ regulatory T cell isolation kit (Miltenyi Biotec)を使用して、
マウス脾臓細胞からCD4+CD25-T細胞を磁気分離した。抗マウスCD3抗体 (5 µg/ml, eBiosicence, San Diego, CA)にてコーティングしたプレートにて、抗マウスCD28抗 体 (1 µg/ml, eBiosicence)を用い、CD4+CD25-T細胞 (1×106/well) を3日間刺激した。
活性化T細胞 (1×106/well) とMSC (20×103/well) とを完全培地で共培養した。Th17 細胞を分化誘導するためにリコンビナントマウスTGFβ1 (2 µg/ml, R&D Systems) とIL-6 (50 µg/ml, R&D Systems)を添加した。3.5日間共培養し、浮遊細胞と培養上 清を回収した。浮遊細胞 (1×106) をPerCP標識抗マウスCD4抗体、FITC標識抗マ ウス CD8a 抗体で染色した後、Foxp3 staining buffer kit (eBioscience) を使用し R-PE標識抗マウスIL-17抗体とAPC標識抗マウスIFNγ抗体と反応させた。染色細 胞をFACSCalibur (BD Bioscience)で解析した。また、培養上清中のIL-17をMouse
IL-17 Quantikine ELISA Kit (R&D Systems)で測定した。
4-2-21. レシピエントマウスBMMSCのMRL/lprマウス投与実験
20週齢レシピエントMRL/lprマウスから単離培養したBMMSCをマウス体重10g 当り 0.1 106個の割合で 16 週齢 MRL/lpr マウスに経静脈的に投与した。同一週齢
MRL/lpr マウスを対照群として使用した。マウスが死亡するまで飼育した。また
BMMSC投与4週後 (20週齢MRL/lprマウス)、マウスより末梢血血清と尿を採取し た (図4-1)。
図4-1. レシピエントMRL/lprマウスBMMSCのMRL/lprマウスへの投与の概略図。
MSC 移植群および非移植群のレシピエント MRL/lpr マウスから骨髄を採取し、
CFU-F 法を用いて BMMSC を単離した。レシピエント由来 BMMSC を 16 週齢の
MRL/lprマウスに経静脈的に投与した。
4-2-22. レシピエントマウスT細胞の養子細胞移植実験
レシピエントMRL/lprマウスの脾臓より磁気分離したマウスPanT細胞 (マウス当 り10×106) を尻静脈から免疫不全マウスNOD SCIDマウス (8週齢) に移植した。
PanT細胞養子移植後2日目にレシピエントMRL/lprマウス由来のBMMSCをマウ
ス体重 10g 当り 0.1 106個の割合で養子移植マウスに経静脈投与した。いずれのマ
ウスも死亡するまで飼育した (図4-2)。
Formation of!
CFU-fs !
ex vivo expansion!
of mBMMSCs!
Bone Marrow of!
Femurs & Tibiae!
Isolation of ! Bone Marrow Cells!
16 days! 7-10 days!
hMSC!
Transplantation of!
mBMMSC ! mBMMSC!
Transplantation of!
human MSC !
16-week-old!
MRL/lpr mice!
16-week-old!
MRL/lpr mice!
図 4-2. MRL/lpr マウス PanT 細胞の SCID マウスへの養子移植ならびに養子移植 SCID マウスへのレシピエント BMMSC の全身的投与の概略図。hMSC 移植または
非移植の MRL/lpr マウスの脾臓から磁気分離システムを利用して純化した PanT 細
胞 (PanT)を経尾静脈でSCIDマウスに移植した。hMSC移植または非移植のレシピ エントMRL/lprマウスから単離したレシピエントBMMSCをPanT移植SCIDマウ スに経静脈投与した。
4-2-23. 統計学的処理
2グループ間の有意差検定にはStudent’s t-testを使用した。0.05以下のP値を有意 差ありと見なした。
Isolation of ! Spleen Cells!
Formation of!
CFU-fs !
ex vivo expansion!
of BMMSCs!
Bone Marrow of!
Femurs & Tibiae!
Isolation of ! Bone Marrow Cells!
16 days! 7-10 days!
hMSC!
Sorting of ! PanT Cells!
Transplantation of!
human MSC !
Analysis!
Transplantation of!
mBMMSC ! BMMSC!
PanT!
MRL/lpr mice!
Spleen!
Adaption of PanT cells !
4-3. 結果
4-3-1. MRL/lprマウスにおけるSupSC細胞移植の治療学的効果
ヒト由来のBMMSC、SHED、臍帯血間葉系幹細胞などのヒト間葉系幹細胞をSLE
モデルMRL/lprマウスへ全身的投与した場合、種々のSLE様症状を改善する治療効
果が報告されている (Zhou et al., 2008; Sun et al., 2009; Yamaza et al., 2010)。そ こで、SLEに対するSupSCの治療効果を検討する為に、16週齢MRL/lprマウスに
SupSC を経静脈的に細胞移植した。hBMMSC も同様に投与した。また、同一週齢
MRL/lprマウスにPBSを経静脈投与したマウスを対照群として使用した。
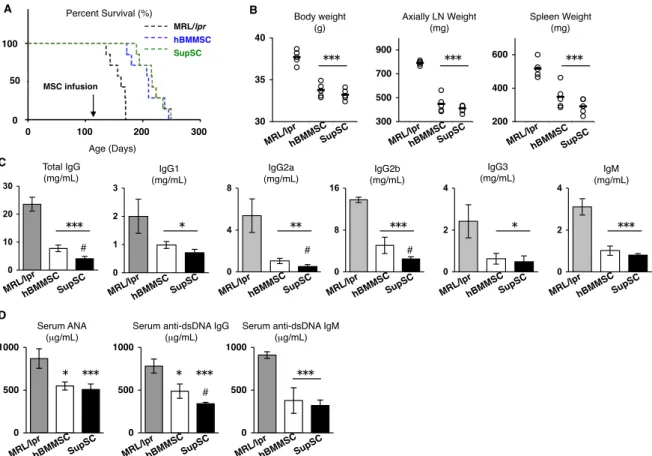
Kaplan-Meyer法による生存率の解析では、SupSC 移植群 MRL/lprマウスでは対 照 群 MRL/lpr マ ウ ス と 比 較 し て 寿 命 の 延 長 が 認 め ら れ た(図 4-3 A)。 Mantel-Haenszel試験により各実験群の平均生存日数は、対照群が162日、hBMMSC 移植群が 210日、SupSC 移植群が216 日であった。対照群と比較してSupSC移植 群および hBMMSC 移植群の平均寿命には有意差が認められた (p<0.005)。SupSC 移植群の平均寿命とhBMMSC移植群の平均寿命では統計学的有意差はなかった。
20週齢対照群MRL/lprマウスでは体重の増加やリンパ節肥大、脾腫が観察された。
SupSC 移植マウスではこれらの増大現象に対し抑制効果が認められた (図 4-3 B)。
hBMMSC移植群もSupSC移植マウスと同等の抑制効果が得られた。この結果から、
SupSC移植がMRL/lprマウスの重度のリンパ増殖症を改善する可能性が示唆された。
SLE 患者において、血中免疫グロブリン値や自己抗体価は SLE の発症および進行 とその治療効果を評価する指標と知られている。対照群末梢血では、免疫グロブリン (IgG1、IgG2a、IgG2b、IgG3、IgM) (図4-3 C)および自己抗体 (抗核抗体[anti-neuclear antibody; ANA]、抗二本鎖 DNA[anti-double strand DNA; dsDNA]抗体である抗
ds-DNA IgG抗体や抗ds-DNA IgM抗体) (図4-3 D)の濃度が顕著に上昇していた。
従って、20 週齢 MRL/lpr マウスでは、重度の SLE 様症状を発症していることを示
唆している (Sun et al.,2009; Yamaza et al., 2010)。
SupSCを移植した場合、対照群 MRL/lprマウスと比較して、免疫グロブリンや自
己抗体 (ANA、抗ds-DNA IgG抗体や抗ds-DNA IgM抗体) の血清濃度が顕著に低 下していた (図 4-3 D)。hBMMSC移植群でもSupSC移植と同等の結果を得たが、
SupSC群との間に顕著な差は認められなかった (図4-3 C)。
図4-3. SupSC移植によるMRL/lprマウスの生存と血中免疫グロブリンおよび自己抗
体レベルへの効果。(A): Kaplan-Meier生存率曲線。SupSC移植はMRL/lprマウス の生存期間を延長させた。SupSC 移植群 (緑線, SupSC, n=7) は対照群 (黒線, MRL/lpr, n=7)と比べ、MRL/lprマウスの生存が延長していた。hBMMSC移植群 (青 線, hBMMSC, n=7) はSupSC移植群と同等の生存率を示した。(B): 20週齢SupSC 移植群において体重、腋下リンパ節 (LN) および脾臓の重量が減少していた。
P<0.005 (vs. MRL/lpr)。n=5。バーは平均値を示す。(C): SupSC移植群では血清
A!
C!
B!
0 500 1000
1 0
500 1000
0 1
500 1000
1
Serum ANA!
(µg/mL)!
***!
*!
Serum anti-dsDNA IgM!
(µg/mL)!
***! Serum anti-dsDNA IgG!
(µg/mL)!
***!
*!
#!
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC! 0
1 2 3
1 0
4 8
1 0
8 16
1 0
2 4
1 0
2 4
1
IgG1!
(mg/mL)!
IgG2a!
(mg/mL)!
IgG2b!
(mg/mL)!
IgG3!
(mg/mL)! IgM!
(mg/mL)!
MRL/lpr
! hBMMSC
! SupSC
!
MRL/lpr
! hBMMSC
! SupSC
! MRL/lpr
! hBMMSC
! SupSC
!
MRL/lpr
! hBMMSC
! SupSC
!
MRL/lpr
! hBMMSC
! SupSC
!
200! 300!
Age (Days)!
Percent Survival (%)!
MRL/lpr!
hBMMSC!
SupSC!
MSC infusion!
100!
50!
0!
0! 100!
D!
*! **!
#!
***!
#!
*! ***!
0 10 20 30
1
MRL/lpr
! hBMMSC
! SupSC
!
***!
#!
Total IgG!
(mg/mL)!
30 35 40
0 0.5 1 1.5 2 2.5 3 3.5 300
500 700 900
0 0.5 1 1.5 2 2.5 3 3.5 200
400 600
0 0.5 1 1.5 2 2.5 3 3.5
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC
! Body weight!
(g)!
Axially LN Weight!
(mg)!
Spleen Weight!
(mg)!
***!
***!
***!
免疫グロブリン値が減少していた。P<0.05, P<0.01, P<0.005 (vs. MRL/lpr);
#P<0.05 (vs. hBMMSC)。n=5。棒グラフは平均値±標準偏差を示す。(D): SupSC移 植群では血清自己抗体 (抗核抗体 (ANA) と抗 dsDNA IgG抗体および IgM抗体)が 減少していた。P<0.05, P<0.005 (vs. MRL/lpr); #P<0.05 (vs. hBMMSC)。n=5。
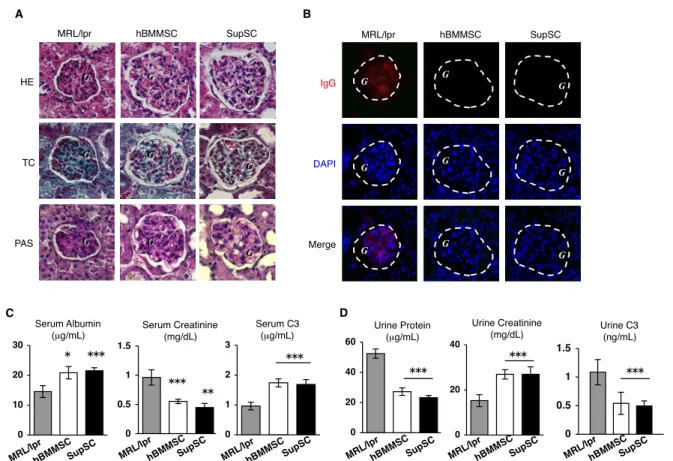
ヒト SLE 患者と同様、腎炎は MRL/lpr マウスの主な障害の一つである (Sun et al.,2009; Yamaza et al., 2010)。組織学的および免疫蛍光染色による解析から、20週
齢対照群 MRL/lpr マウスの腎組織ではメサンギウム基質の過剰成長、基底膜障害、
糸球体へのIgG沈着などの重度の糸球体腎炎を示す症状が観察された (図4-4 A, B)。 血清および尿を用いた腎機能評価では、対照群では血清アルブミンレベルが低下し、
尿タンパクが上昇していた。クレアチニンは血清中で顕著に上昇しており、尿中では 減少し、逆にC3レベルは血清中で顕著に下降しており、尿中では上昇していた。つ まり、20週齢対照群MRL/lprマウスは重篤な腎機能不全を呈していた (図4-4 C, D)。
SupSC 移植群マウスで腎の組織学的および機能的評価を行うと、萎縮した糸球体や
IgG沈着などの糸球体腎炎様所見が顕著に消失し、腎濾過機能も著しく改善されてい た (図4-4)。同様の改善傾向がhBMMSC移植群でも認められた (図4-4)。
図4-4. SupSC移植によるMRL/lprマウスの腎障害の改善効果。(A): H&E染色 (HE)、 Gomorriトリクローム染色(TC)、過ヨウ素酸染色 (Periodic acid-Schiff; PAS)法によ る組織解析。SupSC 移植は MRL/lpr マウスで認められる 腎炎や基底膜障害、糸球 体(G)におけるメサンギウム細胞の過成長などの典型的腎障を改善した。(B): 免疫蛍 光染色法の結果。SupSC移植はMRL/lprマウスの糸球体におけるIgG沈着(赤)を 減少させていた。DAPIによる対比染色 (青)。(C, D): SupSC移植MRL/lprマウスに おいて、著明な血清中アルブミン値の上昇 (C) と尿タンパク値の低下が観察された。
対照群と比較し、SupSC 移植群では血清中クレアチニン値と補体3 (C3)値の低下 (C)と尿中クレアチニン値と C3値の上昇をもたらした(D)。P<0.05, P<0.01,
P<0.005 (vs. MRL/lpr)。n=5。 棒グラフは平均値±標準偏差を示す。
4-3-2. SupSC細胞治療によるMRL/lprマウスにおける全身性免疫抑制効果
SLE の発症に Th17 細胞や Th1 細胞、Treg の関与が報告されている(Sun et al.,2009; Shah et al.,2010; Hou et al.,2011)。Th17細胞は自己免疫疾患や炎症状態 を悪化させる (Sato et al.,2006; Garrett-Sinha et al.,2008)。またSLE患者の血清で
A! B!
HE!
TC!
PAS!
MRL/lpr! hBMMSC! SupSC!
G G
G
G G
G
G G
G
IgG!
DAPI!
Merge!
MRL/lpr! hBMMSC! SupSC!
G G G
G G G
G G G
0 20 40 60
1
***!
MRL/lpr
! hBMMSC
! SupSC! Urine Protein!
(µg/mL)!
0 0.5 1 1.5
1
***!
MRL/lpr! hBMMSC
! SupSC! Urine C3!
(ng/mL)!
0 20 40
1
***!
MRL/lpr
! hBMMSC
! SupSC! Urine Creatinine!
(mg/dL)!
0 0.5 1 1.5
0 1
10 20 30
1
Serum Creatinine!
(mg/dL)!
***!
**! Serum Albumin!
(µg/mL)!
*! ***!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC! C!
0 1 2 3
1
***!
MRL/lpr! hBMMSC
! SupSC! Serum C3!
(µg/mL)!
D!
はIL-17レベルが異常に上昇していた (Kurasawa et al.,2000; Wong et al.,2008)。 また、SLE 患者の末梢血単核細胞では Th1 細胞に関連するサイトカイン IFNγの mRNAが上昇していた (Akahoshi et al., 1999)。従って、SLEにおいてTh17細胞 とTh1細胞がSLE発症の重大な病因因子であると示唆している。フローサイトメト リー解析より、対照群 MRL/lpr マウスと比較して SupSC 移植群マウスでは末梢血 CD4+IL-17+IFNγ−Th17細胞が著しく減少していた (図4-5 A)。ELISA解析により、
SupSC 投与群における血清 IL-17 レベルの減少も明らかとなった (図4-5 C)。末梢 血 CD4+IL-17−IFNγ+Th1細胞ならびにIFNγのレベルは、対照群と比べSupSC移 植群では著明に減少していた (図4-5 A, C)。hBMMSC移植群においても同様のTh17 細胞および Th1 細胞の分化、サイトカイン産生において抑制効果が認められたが、
その抑制効果はSupSC移植の方が有意に高かった (図4-5 A, C)。
Tregは、ヘルパーT細胞の細胞増殖とサイトカイン分泌を抑えることで自己免疫の 発症を抑制し、免疫寛容の維持へも関わり深いT細胞である (Akahoshi et al., 1999)。 過去の研究 (Sun et al.,2009; Yamaza et al., 2010) と同じく、SupSC移植群での末 梢血 CD4+CD25+Foxp3+Treg 細胞割合および血清 IL-10 の濃度は、対照群または hBMMSC移植群との間に有意な差が認められなかった (図4-5 A, C)。
Tregと Th17細胞および Tregと Th1細胞のバランスは、SLE を含む自己免疫疾 患の発症や進行に重要な因子として考えられている (Sun et al.,2009; Shah et al., 2010; Yamaza et al., 2010)。SupSC移植群におけるTreg/Th17またはTreg/Th1の 比率は対照群と比べ有意に上昇していた (図4-5 B)。hBMMSC移植群においても同 様の傾向が認められたが、SupSC移植群においてはTreg/Th17細胞比が著しく上昇 していた (図4-5 B)。以上の結果を踏まえると、SupSC移植はTh17やTh1等に起
因する過剰に活性化した免疫システムを抑制制御することによって Treg優位の免疫 環境を再構築し、MRL/lprマウスのSLE様症状に対して治療効果をもたすことが示 唆している。
図4-5. SupSC移植による末梢血Th17細胞およびTh1細胞、Tregの変化。(A): 対 照群と比較して、SupSC 移植群では、末梢血 CD4+IL17+IFNγ-Th17 細胞および CD4+IL17-IFNγ+Th1 細 胞 が 著 明 に 減 少 し て い た 。 一 方 、 末 梢 血 CD4+CD25+Foxp3+Tregは実験群間で有意な差は認められなかった。(B): SupSC移 植群は、非移植群と比較して、Treg/Th17またはTreg/Th1の細胞比が有意に高かっ た。P<0.005 (vs. MRL/lpr), #P<0.05 (vs. hBMMSC)。(C): 血清IL-17およびIFNγ
値がSupSC移植群において著明に低下した。末梢IL-10値は 実験群間で有意な差を
認めなかった。P<0.005 (vs. MRL/lpr); #P<0.05, ###P<0.005 (vs. hBMMSC)。n=5。
4-3-3. SupSC細胞治療によるMRL/lprマウス脾臓および腎臓における免疫抑制効果
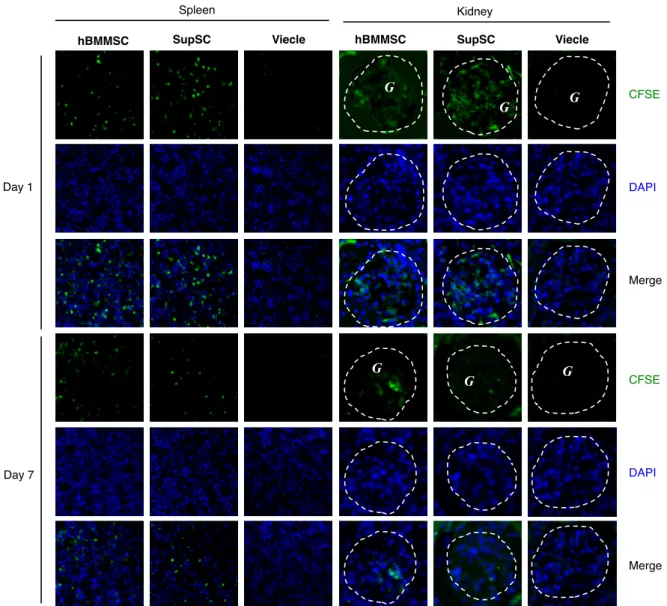
SupSC が障害組織へ遊走する能力を検索するために、CFSE でラベリングした
SupSC をMRL/lprマウスへ静脈内投与した。注入1日後、CFSE 陽性SupSCは腎 の糸球体ならびに脾臓組織において認められた (図 4-6)。注入7日後、SupSC は両 組織で確認できたが、その数は低下していた。hBMMSC も SupSC と同様の動態を
0 10 20
1
0 40 80
1
0 2 4 6
1
***!
#!
IL-17+IFN"- (Th17)!
(% in CD4+ T cells)!
Serum IFN"!
(pg/ml)!
***!
###!
MRL/lpr! hBMMSC
! SupSC!
0 10 20 30
1
***!
###!
Serum IL-17!
(pg/ml)!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC! A!
0 5 10
1
IL-17-IFN"+ (Th1)! (% in CD4+ T cells)!
MRL/lpr! hBMMSC
! SupSC!
***!
###!
0 10 20 30
1
CD25+Foxp3+!
(% in CD4+ T cells)!
Serum IL-10!
(pg/ml)!
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC!
0 25 50
1
***! #!
Treg/Th17!
MRL/lpr! hBMMSC
! SupSC! B!
0 5 10
1
***! **! Treg/Th1!
MRL/lpr
! hBMMSC
! SupSC!
C!
示した (図4-6)。一方、非標識SupSC移植群の脾臓および腎臓ではCFSE陽性細胞 は観察されなかった (図4-6)。
図4-6. MRL/lprマウスの脾臓と腎臓における移植SupSCの局在。CFSE標識SupSC
または hBMMSC を経静脈的にMRL/lpr マウスに移植し、CFSE陽性細胞の局在を
調べた。移植後 1日ではCFSE 陽性SupSC がMRL/lprマウスの脾臓と腎臓の糸球 体 (点線で囲まれた部分、G) に観察できた。移植後 7日では移植 1日に比べ少量の
CFSE 陽性SupSC が確認された。hBMMSC 移植群においても同様の標識細胞の局
在が認められた。非標識SupSC移植群では陽性細胞は認められなかった。
Spleen!
SupSC!
hBMMSC! Viecle!
Day 1!
Day 7!
Kidney!
SupSC!
hBMMSC! Viecle!
G
G G
G G G
CFSE!
DAPI!
Merge!
CFSE!
DAPI!
Merge!
SupSC細胞移植による MRL/lprマウス脾臓ならびに腎臓への治療効果を検討する ために、各組織を摘出し、IL-17とIFNγの濃度をELISA法で解析した。対照群の脾 臓および腎臓において亢進したIL-17とIFNγ濃度は、SupSC移植群において顕著に 抑制されていた (図4-7 A, B)。SupSC移植群ではhBMMSC移植群に比べこれらサ イトカインの顕しく抑制されていた (図4-7 A, B)。次に、SupSCのT細胞に対する 抑制効果を調べる為に、各実験群 MRL/lpr マウスの脾臓から PanT 細胞を単離し、
抗CD3抗体および抗CD28抗体刺激下で3日間培養を行い、培養上清でのIL-17お よびIFNγの分泌を解析した。対照群由来のPanT細胞 (T-MRL/lpr) の培養上清では 高いレベルで IL-17 と IFNγが検出された。一方、SupSC 移植群由来 PanT 細胞 (T-SupSC) と hBMMSC 移植群由来 PanT 細胞 (T-hBMMSC) の培養上清では、
T-MRL/lpr のレベルと比較して IL-17 と IFNγはともに有意に減少していた (図 4-7 C)。T細胞の組織為害作用を調べる為に、各実験群のPanT細胞を免疫不全マウスに 養子移植し、その生存試験を行った。Kaplan-Meyer 生存率曲線の結果から、
Fronziano et al. (1994) の報告と同様、T-MRL/lpr養子細胞移植が免疫不全マウスに 致死的状態を招いた (図 4-7 D)。T-SupSC移植群ならびにT-hBMMSC移植群では 生存期間の延長が認められた (図4-7 D)。Mantel-Haenszel試験から、PanT細胞注 入後の生存日数の平均値が T-MRL/lpr 養子移植群では10 日に対し、T-SupSC養子 移植群では37日、T-BMMSC移植群では36日を示した。T-SupSC養子移植群およ
び T-hBMMSC 養子移植群の生存期間は T-MRL/lpr養子移植群と比べ有意に延長し
ていた (P<0.005)。T-SupSC養子移植群とT-hBMMSC養子移植群との間では生存 期間に有意差は認められなかった。
図4-7. SupSC移植による免疫抑制効果。(A, B): 対照群と比較し、SupSC移植群の 腎臓(A)と脾臓(B)における IL-17 と IFNγのレベルは著明に低下していた。P<0.05,
P<0.005 (vs. MRL/lpr); #P<0.05 (vs. hBMMSC)。n=5。(C): 対照群(T-MRL/lpr)
およびhBMMSC移植群 (T-SupSC)、SupSC移植群 (T-SupSC) より単離したPanT 細胞を抗 CD3抗体と抗 CD28抗体で 3日間刺激培養した。 T-MRL/lpr群で上昇し ていた IL-17 および IFNγが T-SupSC 群では著しく低下した。P<0.05, P<0.01,
P<0.005 (vs. T-MRL/lpr); #P<0.05 (vs. T-hBMMSC)。 n=5。 (D): PanT 細胞 (10 107) を 8~10 週齢の SCID マウスに経静脈投与した。T-MRL/lpr 養子移植群 (T-MRL/lpr, 赤線, n=7) と比較してT-SupSC養子移植群 (T-SupSC, 緑線, n=7) お よび T-hBMMSC 養子移植群 (T-hBMMSC, 青線, n=7) では生存期間が延長してい た。P<0.005 (vs. T-MRL/lpr)。黒線: 非養子移植群 (Control, n=7)。
4-3-4. SupSC移植のMRL/lprマウスでの骨粗鬆症および骨芽細胞ニッチへの効果
MRL/lpr マウスではその骨格系において顕著なBMDの低下や骨梁減少を伴う骨粗
鬆症様症状を呈していた (図4-8 A)。本研究でのマイクロCT解析から、MRL/lprマ
SupSC BMD ( 4-8 A)
0 15 30
1
0 10 20 30
1 0
40 80
1
0 15 30
0 1
4 8
1 0
50 100 150
0 1
20 40
1
IL-17 (pg/ml)! IFN! (pg/ml)!
Spleen!
IL-17 (pg/ml)! IFN! (pg/ml)!
Kidney!
Spleen-PanT!
IL-17 (ng/ml)! IFN! (ng/ml)!
***"
#"
***"
#"
***"
#"
***"
#"
**"
#"
*
**"
#"
*
MRL/lpr
! hBMMSC
! SupSC! MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC!
MRL/lpr
! hBMMSC
! SupSC!
T-MR L/lpr!
T-hBMMSC
! T-Su
pSC! T-MR
L/lpr! T-hBMMSC
! T-Su
pSC!
B! A!
C! D!
Days! 0!
50!
100!
20! 40!
0!
Percent Survival (%)!
uninfused control!
T-MRL/lpr!
T-hBMMSC!
T-SupSC!
らに、BV/TV、Tb.Th、Tb.N、Tb.Spなどの各種骨パラメーター解析結果から、SupSC 移植によるMRL/lprマウスの骨梁構造の回復を示した (図4-8 B)。hBMMSC移植群 ではSupSC移植群と同等のBMDと骨梁構造の改善が認められた(図4-8 A, B)。マイ クロ CT および組織学的解析から、対照群と比較して SupSC 移植群ならびに hBMMSC 移植群では骨梁の形成が促進されていた (図 4-8 C)。半定量的 RT−PCR 解析により、対照群MRL/lprマウス脛骨組織に比べ、SupSC移植群やhBMMSC移 植群脛骨での Runx2、Alp、Osteocalcin などの骨芽細胞特異的遺伝子発現が上昇し
ていた (図 4-8 D)。さらに骨芽細胞ニッチを計測したところ、対照群と比較して
SupSC 移植群ならびに hBMMSC 移植群では骨組織における骨芽細胞ニッチが増加
していた (図 4-8 E)。以上の結果は、SLE モデルマウスの骨格系において、SupSC 細胞移植が骨形成を促し、骨芽細胞ニッチの再構築を促進していることが示唆された。
図4-8. SupSC移植によるMRL/lprマウスでの骨粗鬆症および骨芽細胞ニッチ形成へ の影響。(A): SupSC移植により骨塩濃度 (BMD)の改善効果が認められた。(B): 骨パ ラメーター解析。P<0.05, P<0.01, P<0.005 (vs. MRL/lpr)。n=5。棒グラフは平 均値±標準偏差を表す。(C): マイクロCT像と組織学的解析によるMRL/lprマウスに おける骨構造。SupSC 移植により著明な海綿骨の増加が認められた(黄線と青線で 囲まれた領域)。(D): 半定量的RT-PCR解析の結果、非移植群に比べSupSC移植群 ではRunx2, Alp, Ocn等の骨芽細胞特異的遺伝子の発現が上昇していた。P<0.005 (vs. MRL/lpr)。n=5。(E): MRL/lprマウスの脛骨近位骨幹端部において、骨髄領域当 たりの骨芽細胞の数を計測した。SupSC 移植群において骨芽細胞の数が著明に増加 していた。P<0.005 (vs. MRL/lpr)。n=5。棒グラフは平均値±標準偏差を表す。
0 0.5 1
0 1
1 2
0 1
1 2
1
Runx2 Ratio!
(To G3PDH)!
Alp Ratio!
(To G3PDH)!
Ocn Ratio!
(To G3PDH)!
0 0.2 0.4
0 1
2 4
0 1
0.2 0.4
0 1
20 40 60
0 1
0.5 1
1
BMD (g/cm3)! BV/TV (%)! Tb.Th (mm)! Tb.N (1/mm)! Tb.Sp (mm)!
*! **!
**! **! *! ***! ***! **!
*!
MRL/lpr! hBMMSC! SupSC!
A! B!
C! MRL/lpr!
hBMMSC
! SupSC!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr! hBMMSC
! SupSC!
D!
MRL/lpr! hBMMSC
! SupSC!
Runx2!
Apl!
G3pdh!
Ocn!
MR L/lpr!
hBMMSC
! SupSC
! MRL/lpr!
hBMMSC
! SupSC
! MR
L/lpr! hBMMSC
! SupSC
!
***! ***!
***!
E!
0 20 40
1
MRL/lpr! hBMMSC! SupSC! OB (/Niche Area)!
***!
4-3-5. SupSC移植によるMRL/lprマウス骨組織における破骨細胞分化と活性の抑制
SupSC 移植により骨梁の増加が認められたことから、MRL/lpr マウス骨組織にお
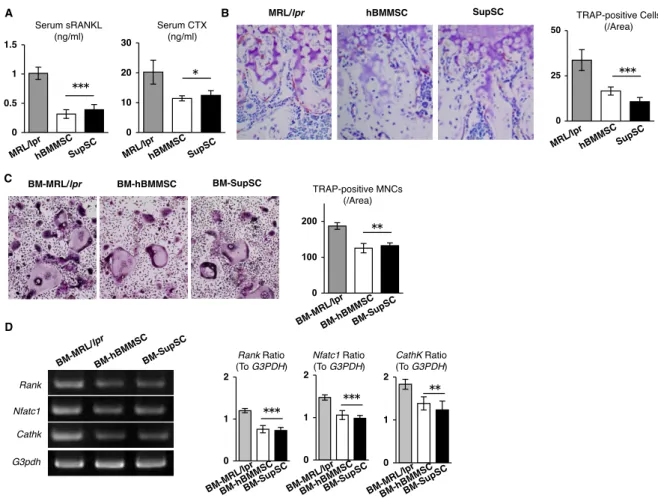
け る 破 骨 細 胞 の 活 性 を 解 析 し た 。 破 骨 細 胞 分 化 決 定 的 因 子 で あ る soluble runt-related NF-κB ligand (sRANKL) や骨吸収マーカーであるI型コラーゲンC末 端テロペプチドの血清中濃度をELISA法で検索した。対照群MRL/lprマウスで著し く高い濃度を示していたこれらのマーカーは、SupSC 移植群において顕著な低下を 示した (図4-10 A)。TRAP染色による組織学的解析から、対照群と比較してSupSC 移植群の脛骨骨端部においてTRAP陽性細胞の著明な減少が観察された (図4-10 B)。
hBMMSC移植群においても同等の破骨細胞活性の抑制効果が認められた (図4-10 A,
B)。
破骨細胞の分化能を検索する為、各実験群のレシピエント骨髄細胞をマクロファー ジコロニー刺激因子と sRANKLの存在化の下、破骨細胞様細胞を分化誘導した。対 照群と比較して、SupSC移植群由来骨髄培養系では著明なTRAP陽性多核細胞の減 少が観察された (図4-9 C)。半定量的RT-PCR法により破骨細胞特異的遺伝子を解析 すると、対照群と比べ SupSC 移植群の破骨細胞培養系において Rank、Nfatc1、
Cathepsin Kの発現が低下していた (図4-9 D)。hBMMSC移植群の破骨細胞培養系 においても同等の破骨細胞分化抑制が認められた (図 4-9 C, D)。以上のことから、
SupSC移植によりMRL/lprレシピエントマウスにおける破骨細胞の分化と活性の下
方制御がSupSC移植による骨量増加にも寄与している可能性が示唆された。
図4-9. SupSC移植によるMRL/lprマウスにおける破骨細胞の分化と活性の抑制効果。
(A): MRL/lpr マウスの血清中における可溶性 RANKL (sRANKL)と C-terminal telopeptides of type I collagen (CTX) の上昇がSupSC移植群では抑制されていた。
P<0.05, P<0.005 (vs. MRL/lpr)。n=5。棒グラフは平均値±標準偏差を表す。(B): 対 照群と比較して、SupSC移植群では脛骨近位骨幹端におけるTRAP陽性細胞が著し く減少していた。P<0.05, P<0.005 (vs. MRL/lpr)。n=5。(C, D): in vitro RANK 誘導性破骨細胞分化培養系において、非移植群骨髄細胞(BM-MRL/lpr)に比べSupSC 移植マウス由来骨髄細胞 (BM-SupSC)では、TRAP 陽性多核細胞 (TRP-positive MNCs) の数が減少していた。(D): 破骨細胞特異的遺伝子Rank, Nfatc1, cathepsin K (CathK)の 発 現 は 非 移 植 群 に 比 べ SupSC 移 植 群 で は 顕 著 に 低 下 し て い た 。 BM-hBMMSC: hBMMSC移植 MRL/lprマウス由来骨髄細胞。P<0.01, P<0.005 (vs. BM-MRL/lpr)。n=5。
0 10 20 30
0 1
0.5 1 1.5
1
*!
***! Serum sRANKL!
(ng/ml)!
Serum CTX!
(ng/ml)!
MRL/lpr! hBMMSC
! SupSC!
MRL/lpr! hBMMSC
! SupSC! A!
0 1 2
1 0
1 2
1 0
1 2
1
0 100 200
1
0 25 50
1
BM-MRL/lpr! BM-hBMMSC! BM-SupSC!
TRAP-positive MNCs!
(/Area)!
BM-MR L/lpr! BM-hBMMSC
! BM-Su
pSC!
**!
B! MRL/lpr! hBMMSC! SupSC!
***! TRAP-positive Cells!
(/Area)!
MRL/lpr! hBMMSC
! SupSC!
BM-MR L/lpr!
BM-hBMMSC
! BM-Su
pSC!
Rank!
Nfatc1!
Cathk!
G3pdh!
BM-MR L/lpr! BM-hBMMSC
! BM-Su
pSC!
***! **!
Rank Ratio!
(To G3PDH)!
Nfatc1 Ratio!
(To G3PDH)!
CathK Ratio!
(To G3PDH)!
D! C!
***!
BM-MR L/lpr! BM-hBMMSC
! BM-Su
pSC! BM-MR
L/lpr! BM-hBMMSC
! BM-Su
pSC!
4-3-6. SupSC移植によるレシピエントBMMSCの形成能への影響
BMMSC は骨芽細胞の前駆細胞にあたるため、レシピエント MRL/lpr マウス
BMMSCへのSupSC移植の効果を検索した。対照群、hBMMSC移植群、SupSC移 植群の各レシピエント MRL/lpr マウスより BMMSC を単離した。各レシピエント BMMSC を便宜上、MSC-MRL/lpr、MSC-hBMMSC、MSC-SupSC と名付けた。
CFU-F 解析の結果、MSC-MRL/lprと比較して MSC-SupSCではコロニー数が減少 していた (図4-10 A)。またポピュレーションダブリング解析とBrdU取り込み解析 の結果から、MSC-SupSCではMSC-MRL/lprよりも高い細胞増殖活性を備えていた (図4-10 B, C)。 MSC-SupSCと同等の結果がMSC-hBMMSCで得られた (図4-10 A-C)。
次にレシピエントBMMSCのin vitro骨形成能を検索した。CFU-F形成培養系を 用いてcolony forming unit-osteoblasts (CFU-OB)を解析した。コロニーを骨誘導培 地で培養し、1週間後ALP染色した。ALP陽性コロニーをCFU-OBとみなし、ALP 陽性コロニー数を計測した。ALP 陽性コロニー数は各群で違いが認められなかった
が、CFU-OB/CFU-F比率を比較すると、MSC-SupSC群において顕しく高い比率を
示した (図 4-10 D)。各群のコロニー形成細胞を播種し増殖させた後、骨誘導培地で
培養した。骨誘導1週後のALP活性 (図4-10 E)ならびに骨誘導4週後のアリザリン レッド陽性面積 (図4-10 F)はMSC-MRL/lpr群と比較してMSC-SupSC群で促進し
ていた。MSC-MRL/lpr 群と比較して MSC-SupSC 群では骨芽細胞特異的遺伝子
Runx2、Alp、osteocalcinが高く発現していた (図4-10 G)。MSC-SupSCと同等の 骨形成促進作用がMSC-BMMSC群で認められた (図4-10 D-G)。