病原微生物の病態発現機序にかかわる諸因子
に関する分子性状解析
山梨大学大学院
医学工学総合教育部
博士課程学位論文
2015 年 3 月
菅野 武史
目次
総合緒論 1 第Ⅰ部 集団発生した食品媒介レンサ球菌感染症原因菌の産生する病原因子の解析 緒論 2 第1 章 集団発生した食品媒介レンサ球菌感染症原因菌の産生する病原因子の解析 第1 節 材料および方法 3 第2 節 結果 6 第3 節 考察 15 要旨 19 引用文献 22 第Ⅱ部 病原性酵母における抗真菌剤標的候補の探索・同定 緒論 25 第1 章 TS-株の分離と性状解析 第1 節 緒言 28 第2 節 材料および方法 材料 28 方法 29 1) 生菌数測定と発育制限温度の設定 2) TS-株の分離とその性状解析 3) TS-株のスクリーニング 第3 節 結果と考察 29第2 章 C.glabrata Genomic DNA Library の構築
第1 節 緒言 32 第2 節 材料および方法
材料 32 方法 33
1) C.glabrata Genomic DNA の調製
2) C.glabrata Genomic DNA Library の調製
3) C.glabrata Genomic DNA Library の構築
4) C.glabrata Genomic DNA Library の性能確認
第3 節 結果と考察 37
第3 章 Genomic Library から TS-株(W12) 相補能保有 DNA 断片の分離
と性状解析
第1 節 緒言 40 第2 節 材料および方法
材料 41 方法 41 1) TS-株に対するOsmotic Stabilizer (Sorbitol) の影響を検討
2) TS-株のC.glabrata Genomic DNA Library による TS+株への復帰
3) TS+株からPlasmid DNA 抽出・増幅 4) Yeast Transformation による確認 5) TS-株および復帰株の各温度におけるコロニー形成能 第3 節 結果と考察 42 第4 章 TS-株 (W12)の必須遺伝子領域の解析と ORF 領域の探索 第1 節 緒言 45 第2 節 材料および方法 材料 45 方法 46 1) 相補遺伝子断片の制限酵素地図 (Restriction Map) の作製 およびTS-相補領域の同定 2) S.cerevisiaeゲノム配列からのORF 領域の推定
3) TS-株のEMS 処理による突然変異箇所 (point mutation) の探索
第5 章 C. albicansにおけるC.glabrata(W12)と同様のROM2 point mutation の導入とROM2 null mutant 構築の試み
第1 節 緒言 52
第2 節 材料および方法 材料 52
方法 52
1) C.albicansにおけるROM2 point mutant の構築 2) C.albicansにおけるpoint mutant の TS 性と Sorbitol 回復の検証 第3 節 結果と考察 53
第6 章 C. glabrata およびC. albicansにおけるCell lysis の検討 第1 節 緒言 55
第2 節 材料および方法 材料 55
方法 55
1) C. glabrata およびC.albicansにおけるCell lysis の分析 第3 節 結果と考察 56
第7 章 Tet System と Real Time RT-PCR による遺伝子解析 第1 節 緒言 58
第2 節 材料および方法 材料 58
方法 59
1) TET system の原理 2) C.glabrata およびC.albicansにおけるROM2のTet 株 (CgROM2 Tet, CaROM2 Tet)の構築 3) TET System による必須性の検討 4) RT-PCR を用いた CgROM2 Tet, CaROM2 Tet 株の Dox の有無 におけるROM2 mRNA 発現量の検討 第3 節 結果と考察 65
総合考察 67
要旨 70
総合結論 76
和文要旨 77
英文要旨 79
関係論文 82
1
総合緒論
われわれをとりまく環境中には人体(宿主)それ自体を含めて多種多様な微生物が存在 している。そのなかで病原微生物といわれているものがしばしば宿主に侵入して増殖する 場合があり、感染とよばれている。特定の微生物種が感染によって特定の宿主に病気を惹 起させる性質、すなわち病原性の発現に際しては、病原微生物側の要因として様々な病原 因子が関与している。 病原微生物にはウイルス、細菌、真菌など様々なものが存在するが、本研究では、その 一つとして、食中毒の原因菌となったA 群レンサ球菌 (Streptococcus pyogenes) に焦点を あて、これが産生する主要な病原因子について検討した。 A 群レンサ球菌には菌体成分に 由来する病原因子も存在するが、菌体外に産生される主要な病原因子である溶血毒素、 cysteine protease streptococcal pyrogenic exotoxin B (SpeB) および NAD glycohydrolase (NADase) の 3 種に着目し、食中毒の病態との関連性について知見を得るため、これら諸 因子の分子性状の解析を行なった。次に、皮膚真菌症や内在性真菌症の原因となる病原性真菌として最近とくに重要視され ているカンジダ属酵母であるCandida glabrata およびC. albicans に焦点をあてて研究を 行なった。医療上重要とされているカンジダ属酵母のなかからこの2菌種を研究対象に取 り上げた理由としては、C. glabrata のみゲノムが一倍体であるため遺伝解析が比較的容易 である点、また、C. albicans がもっとも病原性が強い点、さらに、他の病原性真菌に先駆 けてこれら2菌種でゲノムの全塩基配列が決定された点があげられる。こうした背景から、 この研究では本菌により惹起される感染症(カンジダ症)に対する対策として、新規抗真 菌剤の開発という視点から、菌の発育と病態発現に不可欠とされる必須遺伝子(およびそ の遺伝子産物)を探索・同定することで、これが菌にとって致死的作用を及ぼす薬剤の標 的候補となり得るかについて分子生物学的手法を用いて検討・解析を行なった。 このような病原微生物の病態発現機序にかかわる諸要因についてそれらの分子性状を解 析・理解することは感染症の予防・治療において重要な点であり、これらの知見は感染症 対策の一助になると期待される。
2
第Ⅰ部
集団発生した食品媒介レンサ球菌感染症原因菌の産生する
病原因子の解析
緒論
A 群レンサ球菌感染症はStreptococcus pyogenesを原因菌とした呼吸器感染症である。 本菌は健常人の咽頭や皮膚などにも存在する「常在菌」でもあるが、ヒトに様々な疾患を 引き起こすことが知られており、咽頭痛、発熱等の風邪様症状を惹起するほか、時として 急性糸球体腎炎やリウマチ熱などの続発症を誘発することもある。また、A 群レンサ球菌が 原因となる感染症には呼吸器感染症だけでなく軟部組織壊死、血液凝固異常や多臓器不全 等の重篤な症状を伴う劇症型レンサ球菌感染症(STSS)も報告(Shiseki et al., 1999)されて おり、再興感染症として注目されている。 近年、食品を介して起こる感染症は大きな社会問題となっており、これまでにイスラエ ル、ヨーロッパ、アメリカおよび中国などで食品媒介A 群レンサ球菌感染症の集団発生の 報告(Pournaras et al.,2000; Asteberg et al.,2006; Kuluski et al.,2006; Yang et al.,2007)が 散見される。わが国においても 1990 年代以降 7 例の食品媒介レンサ球菌感染症の報告 (Tanaka et al., 2006)がある。しかしながら、これらの報告は疫学調査が中心であり病態と 病原因子の関係性についての報告は見られない。2005 年 7 月には、神奈川県下(大学のオープンキャンパスの運営役員が弁当を喫食した 489 名中 218 名が発症)で A 群レンサ球菌(S. pyogenes T-25 型)による食品媒介感染症、い わゆる集団食中毒事件が発生した(Suzuki and Nikkawa, 2006)。著者はこの事例に関して、 T-25 型患者由来株の産生する主要な代謝 物質である、溶血毒素、cysteine protease streptococcal pyrogenic exotoxin B (SpeB)および NAD glycohydrolase (NADase)の活性に ついて検討を行ない、これまでに報告されている T-28 型および T-B3264 型(鈴木、坂口, 2009)と比較し、さらに 3 菌型の病原因子の SDS-PAGE によるタンパク質分析を行なった。 また、T-25 型においては溶血毒素である streptolysin O (SLO)、SpeB および NADase の 発現時期について検討した。これまでSLO と SpeB の活性量は反比例すると報告(小林、浜 田, 1981)されているが、これらの関係についてタンパク質分析の成績からも確認した。こ のように食中毒の病態を知る上で病原因子間の発現時期を知ることは不可欠と考えられる。
3
第
1 章
【集団発生した食品媒介レンサ球菌感染症原因菌
の産生する病原因子の解析】
材料および方法
1. 菌株および培養上清の調製
A 群レンサ球菌感染症患者由来株(T-25 型、T-28 型および T-B3264 型)を 5%馬血液寒天 培地に塗抹して、37℃で 24 時間分離培養した。その後、菌型間の病原因子の性状を比較す る目的で単一コロニーを1% Yeast Extract 含有 Todd-Hewitt broth(Difco)に植菌し、37℃ で6 時間前培養を行ない、前培養液を同培地に植菌し、6 時間および 18 時間本培養を行な った。また、T-25 型においては発現時期を特定する目的で 0.3% Yeast Extract 含有 Brain Heart Infusion broth(栄研化学)2mL に植菌し、37℃で 6 時間前培養を行ない、前培養液 0.3mL を同培地 120mL に植菌し、20 時間まで 2 時間ごとに 10mL ずつ回収して培養を終 了した。なお、SpeB 分析に用いる培養上清は 30 時間まで継続した。培養終了後に、培養 液を3,000rpm、15 分間遠心分離し、上清をメンブランフィルター(pore size 0.2μm)で濾過 滅菌した。なお、供試菌株は神奈川県衛生研究所(T-25 型)および東京都健康安全研究センタ ー(T-28 型および T-B3264 型)より分与を受けた。2. 溶血力価および溶血効率測定
培養上清をKusama et al. (1958)の方法で 2.5% rabbit red blood cells(RBC)を用いて試 験管法(log0.1 希釈系列)により溶血力価を求めた。0.05M 2-mercaptoethanol(2-ME)含有 pH6.5, phosphate buffered saline(PBS)を加えた還元条件下にて 37℃で 1 時間反応させた 後、2,500rpm、15 分間遠心分離し、上清に 2%塩酸アセトンを加え、510nm で吸光度を測 定した。完全非溶血を対照とし、完全溶血の力価を100%として各々の試験管の溶血%を算 出し、50%終点法により毒素の力価を得た。また、各々の溶血曲線から溶血効率(hemolytic efficiency;HE)を求めた。なお、溶血効率は完全溶血液の吸光度を 100%として、培養上清 各希釈液の溶血%を求め、これをプロビット換算して、培養上清の対数用量値に対してプロ ットし、得られた直線の傾きから求めた。力価はHD50/mL で表し、1HD50/mL は 2.5% RBC 0.5mL を 50%溶血させる最少量で表した。なお、還元剤依存性は 2.5% RBC を用いて 0.05M 2-ME 存在および非存在下にて、Kusama らの試験管法(log0.1 希釈)により力価測定を行な った。
4
3. 各種阻害物質における溶血阻害態度
2.5% RBC を用いた試験管法にて、還元状態における各種阻害物質による溶血阻害態度を 観察した。培養上清を用いてlog0.1 希釈系列を作り、1 系列を対照とし、残り 3 系列には 各々1 管ずつ阻害物質であるコレステロール(SIGMA)10μL(0.15μg)、γ-グロブリン(化血 研)10μL(0.5μg) 、 リ ン 脂 質 (WAKO)250μL(0.25mg) を 加 え 、 さ ら に bovine serum albumin(BSA) 含有 PBS で計 2mL とし、力価測定を行ない、溶血阻害態度を溶血率の変 化より求めた。さらに、エピガロカテキンガレート(EGCg)による T-25 型の溶血毒素阻害能 について検討した。1mg の EGCg (WAKO)を 1mL の PBS に加えたものを用いて、EGCg を1000μL(1mg)、500μL(0.5mg)加え計 2mL とし、力価測定を行い、溶血阻害態度を溶血 活性の変化として観察した。
4. SpeB の定量
Andreas and Lars (1995)の方法で培養上清 250μL に対して pH7.6, 0.1M phosphate buffer(PB)を用いて液量 750μL に希釈して、さらに酵素の活性化のために 0.3M 2-ME を含 むpH7.6, 0.1M PB を 250μL 加え計 1mL とし、37℃恒温槽で 30 分間反応させた。次に活 性化させた酵素に1%カゼイン(Difco)含有 pH7.6, 0.1M PB を 1mL 加え 37℃恒温槽で 20 分間基質と反応させた。次に 5%トリクロロ酢酸(WAKO)を 3mL 加え反応を停止させ、 3,000rpm、30 分間遠心分離を行なった後、上清の吸光度を 280nm で測定した。対照とし ては植菌していない培養液を用いた。SpeB 活性は単位既知のStreptomyces griseusより 精製した市販のプロテアーゼ(SIGMA)を基準として unit 数を算出した。
5. NADase の定量
Tatsuno et al.(2007)の方法に順じて測定した。まず、マイクロプレートに植菌していな いBrain Heart Infusion 培養液 9μL と培養上清 1μL を加えた。さらに pH7.4, 0.1% BSA 含有 0.1M PB を 80μL 加え計 90μL とした。そこに β-NAD(2.70mg/mL;ORIENTAL YEAST)を 10μL 加え、37℃で 60 分間反応させた。その後、5M NaOH を 35μL 加え、室 温で暗所に60 分間置き、340nm で吸光度を測定した。β-NAD 既知量の吸光度から検量線 を作成し、培養上清中のNADase による β-NAD の減少量(μg)を算出した。unit 数は反応時 間1 時間、培養上清 1μL あたりの β-NAD の減少量(μg)で表した。
6. SDS-PAGE 分析
5 (最終濃度 10%)、-20℃で 1 時間冷却した。その後、アセトンを 700μL 加え、-20℃で 1 時間冷却した後、10,000g、15 分間遠心分離した。上清を取り除き再びアセトンを 700μL 加え、同様に遠心分離した。その後、上清を取り除き、マイクロチューブ内に残存するア セトンを超小型遠心式濃縮機(VC-15SP; TAITEC)で乾燥させ、試料緩衝液(2% SDS, 5% ス クロース, 5% 2-ME, 0.002%ブロモフェノールブルー含有 pH6.8, 0.06M Tris-HCl 溶液) を20μL 加え、SDS-PAGE 試料とした。 SDS-PAGEはLaemmli(1970)の方法に従って行なった。SDS-PAGE試料を沸騰水浴中で 3分間加熱処理し、10% ポリアクリルアミドゲル(e-PAGEL; ATTO)に20μL添加した。通電 条件は定電流20mAでゲルの下端5mmの位置まで約90分間の電気泳動を行なった。分子量 マーカーとしてSDS-PAGE Molecular Weight Standards Broad Range (BIO RAD)を使用 した。市販のクマシーブリリアントブルー(CBB)染色液(EzStain Aqua; ATTO)による染色 を30分間行なった後、蒸留水で一晩脱色を行なった。
7. ウェスタンブロット(WB)分析
WB 装置(ATTO HorizBlot)を用いて SDS-PAGE 後のゲルを polyvinylidene fluoride 膜 (PVDF; BIO RAD)に 60 分間、定電流 1mA/cm2で転写を行なった。次に、PVDF 膜を 0.05%
ポリオキシエチレン(20)ソルビタンモノラウレート(Tween 20 相当品)含有 pH 7.5, 20mM Tris-buffered saline(TTBS)により 5%に調製したスキムミルク溶液でブロッキングした。 その後、TTBS で 5 分間、3 回の洗浄を行なった。次に一次抗体反応を 60 分間、37℃で行 なった。一次抗体は各々rabbit anti-SLO antibody(Bio Academia)、rabbit anti-SpeB antibody(Toxin Technology)および rabbit anti-NADase antibody(Bio Academia)を用いた。 これら一次抗体全てはTTBS で 1:5000 に希釈して使用した。反応後、TTBS で 5 分間、3 回の洗浄を行なった。次に二次抗体 反応を 60 分間、37℃で行なった。二次抗体は biotinylated goat anti-rabbit IgG(BIO RAD)を用い、TTBS で 1:3000 に希釈して使用した。 反応後、TTBS で 5 分間、3 回の洗浄を行なった。その後、Amplified Alkaline Phosphatase Goat Anti-Rabbit Immun-Blot Assay Kit (BIO RAD)を用いてアルカリフォスファターゼ が複数標識されたビオチンとストレプトアビジンを結合させた混合液の作製を行なった。 この混合液にPVDF 膜を浸して 60 分間、37℃で反応させ、二次抗体へのアルカリフォス ファターゼの結合を増幅させた。反応後、TTBS で 5 分間、3 回の洗浄を行なった。その後、 AP Conjugate Substrate Kit(BIO RAD)を用いて 30 分間、アルカリフォスファターゼの発 色を行ない、目的タンパク質を検出した。
6
8. 電気泳動分析によるタンパク質バンドの光学積算値
SDS-PAGE および WB によって確認されたタンパク質バンドを画像解析ソフト(CS Analyzer ver3.0; ATTO)により光学積算値を算出した。
9. T-25、T-28 および T-B3264 型の増殖曲線
3 菌型を Todd-Hewitt broth 培養において 1、3、6、9 および 18 時間の 5 定点で増殖曲 線を検討した。McFarland の比色法(Jean F. MacFaddin,1985)に従って培養液の濁度を 600nm の波長で測定した。1.175%塩化バリウム二水和物水溶液と 1%硫酸水溶液を用いて 用事調製したMcFarland 濁度標準液により検量線を作成し、培養液中の菌数を求めた。
結果
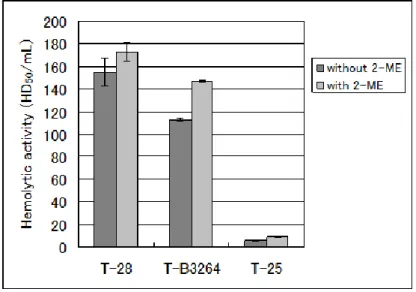
1. 2-ME による溶血活性の依存性
Fig.1 に示すとおり T-25 型の産生する溶血毒素は 2-ME 存在および非存在下でも溶血活 性が認められたが、存在下のほうが高い溶血率を示した。これはT-28 型および T-B3264 型 と同様の成績であった。また、Todd-Hewitt broth 培養における T-25 型の溶血力価は 9HD50/mL であり、T-28 型および T-B3264 型の 173HD50/mL および 147HD50/mL と比較 して著しく低値であった。Fig. 1 Dependence of hemolysis on 2-mercaptoethanol (2-ME).
Hemolytic activity was assayed by 50%-end-point titration with 2.5% rabbit erythrocytes. Hemolysin was measured using supernatants incubated for 6 h.
7
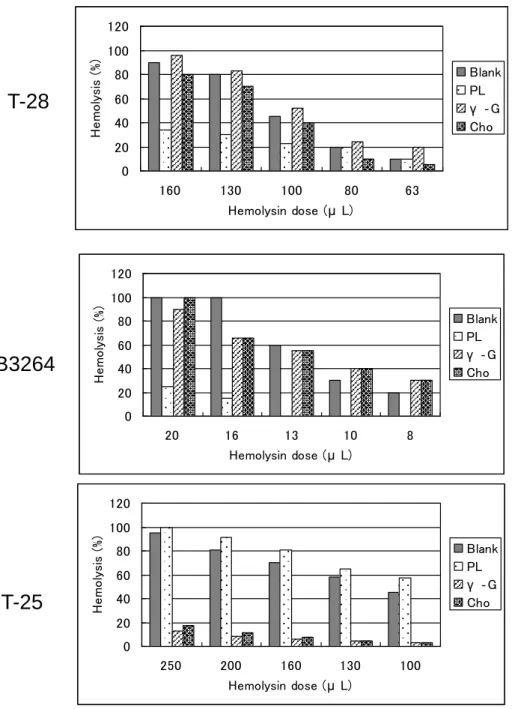
2. 溶血毒素の阻害態度
Fig.2 に示すとおり T-25 型の培養上清はコレステロールおよびγ-グロブリンによって溶 血が阻害され、リン脂質では阻害されなかったため、主要な溶血毒素はSLO であることが 確認された。T-28 型および T-B3264 型の主要毒素はリン脂質で阻害されたことよりストレ プトリジンS(SLS)であり、T-25 型と異なる毒素型であった。Fig. 2 Inhibition of hemolysin produced by Group A streptococcus.
Methods of analysis were the same as in Fig. 1. PL: phospholipid; -G: -globulin; Cho: cholesterol. 0 20 40 60 80 100 120 160 130 100 80 63 Hemolysin dose (μ L) H e mo ly si s (% ) Blank PL γ ‐G Cho 0 20 40 60 80 100 120 20 16 13 10 8 Hemolysin dose (μ L) H e mo ly si s (% ) Blank PL γ ‐G Cho 0 20 40 60 80 100 120 250 200 160 130 100 Hemolysin dose (μ L) H e mo ly si s (% ) Blank PL γ ‐G Cho
T-28
T-B3264
T-25
8
3. 代謝物質のプロファイル
Todd-Hewitt broth 培養における T-25 型の産生する病原因子である溶血活性、SpeB 活性 およびNADase 活性について T-28 型および T-B3264 型と比較を行なったが、Fig.3 のよう に各々菌型によって産生する病原因子の活性量は多様性を示した。なお、Fig.3 では SLO とSLS の目盛り単位が A と B で 10 倍異なっている。また、Table 1 に示すように食中毒 起因菌の産生する代謝物質の主要溶血毒素、溶血力価、溶血効率、SpeB 活性および NADase 活性について3 菌型間で比較したところ産生量および溶血毒素の性状に多様性が見られた。 3 菌型間で比較すると、T-25 型は NADase 活性、T-28 型は溶血活性、および T-B3264 型 はSpeB 活性値が最大であり、活性量は各々19.1Unit、173HD50/mL および 9.7U/mL であ
った。
Fig.3 Profile of metabolic substances produced by Group A streptococcus in food poisoning cases.
Streptolysin O (SLO), streptolysin S (SLS) and NAD glycohydrolase (NADase) were measured using the supernatants incubated for 6 h. Streptococcal pyrogenic exotoxin B (SpeB) was measured using the supernatants incubated for 18 h.
A
B
0 5 10 15 20 SLS(×10) SLO(×10) SpeB NADase T-28 T-B3264 0 5 10 15 20SLS SLO SpeB NADase T-259
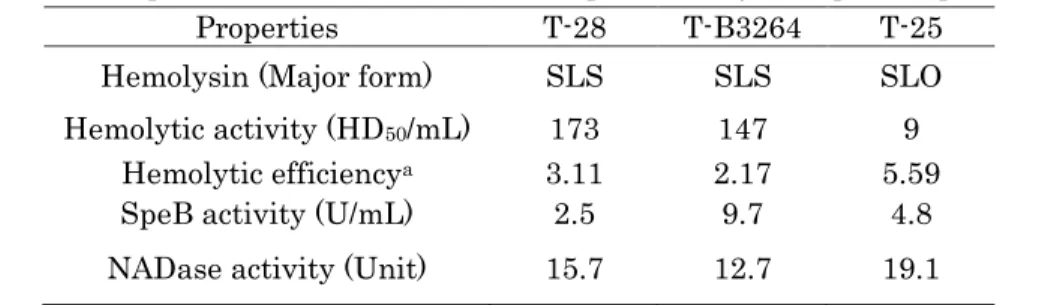
Table 1 Properties of metabolic substances produced by Group A streptococcus Properties T-28 T-B3264 T-25
Hemolysin (Major form) SLS SLS SLO Hemolytic activity (HD50/mL) 173 147 9
Hemolytic efficiencya 3.11 2.17 5.59
SpeB activity (U/mL) 2.5 9.7 4.8 NADase activity (Unit) 15.7 12.7 19.1 Hemolytic efficiency index of streptolysin O (SLO): 6.83±1.35 (N=50).
SLS: streptolysin S; HD50: 50% hemolytic dose of 2.5% rabbit erythrocytes 0.5mL; SpeB:
streptococcal pyrogenic exotoxin B; NADase: NAD glycohydrolase.
aHemolytic efficiency was calculated as the slope of the line according to the Materials
and Methods.
4. 代謝物質の SDS-PAGE 分析
Todd-Hewitt broth 培養における 3 菌型の培養開始 6 時間および 18 時間の培養上清のタ ンパク質分析を行なったところ、Fig.4 に示したように T-25 型は T-28 型および T-B3264 型と比較して6 時間で活性型 SpeB が高産生量であった。なお、3 菌型全てが 18 時間で活 性型SpeB に変換されていた。一方、SLO および NADase の産生量は微量なため CBB 染 色では確認することはできなかった。
Fig.4 SDS-PAGE patterns of various metabolic substances stained with Coomassie brilliant blue. Lane M: molecular weight marker
25 28 B3264 25 28 B3264 6h 18h M Culture time T serotypes SpeB SpeB precursor 200,000 116,250 97,400 66,200 45,000 31,000
10
5. T-25 型における SDS-PAGE 分析による各種代謝物質の発現時期
Fig.5 に示すとおり 44kDa に SpeB 前駆体、そして 30kDa に SpeB を確認することがで きた。また、時間経過とともに44kDa の前駆体から 30kDa の SpeB に変換されているこ とが確認できた。一方、SLO および NADase については量的変化まで CBB 染色で確認で きなかった。
Fig.5 SDS-PAGE patterns of various metabolic substances produced by S. pyogenes type T-25.
For analysis of metabolic substances, brain heart infusion broth was incubated with precultured broth for 2–30 h (with end points every 2 h) at 37℃. Lane M, molecular weight marker. MW, molecular weight.
6. T-25 型における WB 分析による各種病原因子の発現時期
WB 分析により、70 および 60kDa に SLO を確認できた。産生量のタイムコースでは 70kDa の SLO が培養開始 2 時間から 8 時間まで確認された。また、60kDa の SLO は 6 時 間から20 時間まで確認された(Fig.6-A)。NADase は 50kDa で確認され、10 時間で 48kDa 成分を確認した(Fig.6-B)。SpeB は SDS-PAGE 分析と同様に 44kDa に SpeB 前駆体、30kDa にSpeB を確認することができた(Fig.6-C)。 10 14 2 8 12 Culture time (h) 6 4 16 18 20 M 30kDa 116,250 97,400 66,200 45,000 31,000 200,000 MW 22 24 26 28 30 44kDa
11
Fig.6 Western blot (WB) analysis of streptolysin O (SLO) (A), NAD glycohydrolase (NADase) (B) and cysteine protease streptococcal pyrogenic exotoxin B (SpeB) (C) produced by Streptococcus pyogenes type T-25. Lane M, molecular weight marker; lane C, medium alone (brain heart infusion broth)
7. T-25 型における各種病原因子の活性量
溶血活性は培養開始4 時間以降に認められ、8 時間で 47.6HD50/mL と最も高値を示した。 8 時間をピークに減少し始め、14 時間後では 1/5 に減少した(Fig.7)。NADase 活性は 10 時 間で26.3Unit と最も高値を示し、その後、徐々に減少した (Fig.7)。SpeB 活性は 14 時間 から活性が認められ、20 時間でピークを示した。その後、20 時間以降の実験を行なったと ころ、24 時間で 2.04U/mL とピークを示した。しかし、それ以後 30 時間まで活性量はほ ぼ一定であった(Fig.8-B)。(A)
(B)
(C)
66,200 66,200 45,000 M M 2 4 6 8 10 12 14 16 18 20 C 70,000 60,000 2 4 6 8 10 12 14 16 18 20 C Culture time (h) Culture time (h) MW MW 44,000 30,000 50,000 45,000 31,000 M 2 4 6 8 10 12 14 16 18 20 C Culture time (h)12
Fig.7 Activity of SLO, SpeB and NADase produced by S. pyogenes type T-25.
8. T-25 型における各種病原因子の活性量と WB 分析の関係
光学積算値によるSLO のタンパク質発現ピークは培養開始 8 時間であり溶血活性のピー クと一致した(Fig.8-A)。SpeB 前駆体から活性型 SpeB へのタンパク質変換ピークは 14 時 間であり、SpeB タンパク質のピークは 24 時間であった。SpeB 活性が認められたのは 14 時間からであり、活性最大のピークは24 時間であったのでタンパク質 SDS-PAGE 分析と 一致した(Fig.8-B)。SpeB は前駆体から SpeB への変換後に活性が認められることが示され、 SLO 活性は SpeB に変換する直前にピークを迎えることが明らかとなった。NADase タン パク質発現ピークは12 時間であったが、活性ピークでは 10 時間であった(Fig.8-C)。なお、タンパク質発現量分析では SLO および NADase は免疫染色、SpeB は CBB 染色 によって検出されたバンドの光学積算値から測定した。 0 10 20 30 40 50 2 4 6 8 10 12 14 16 18 20 Culture time (h) H e mo ly tic a c tiv it y (H D50 / mL ) 0 0.4 0.8 1.2 1.6 2 S pe B a c tiv it y (U / mL ) N AD as e a c tiv it y (U n it )
13
Fig.8 Activity assay and electrophoretic analysis by SDS-PAGE of metabolic substances produced by S. pyogenes type T-25: (A) SLO, (B) SpeB, (C) NADase
0 5000 10000 15000 20000 25000 2 4 6 8 10 12 14 16 18 20 Culture time (h) S LO pr ot ei n (I nt eg ra ted va lue) 0 10 20 30 40 50 H em ol yt ic act iv it y (H D 50 /m L) 0 20000 40000 60000 80000 100000 120000 140000 160000 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 Culture time (h) 0 0.5 1 1.5 2 2.5 S pe B p ro te in ( In te gr at e d v al u e ) S pe B a c ti vi ty ( U / m L) S pe B p re c u rso r 0 2000 4000 6000 8000 10000 12000 2 4 6 8 10 12 14 16 18 20 Culture time (h) NA D as e p ro te in (I n te gr at e d va lu e ) 0 10 20 30 NA D as e ac ti vi ty (Un it )
(A)
(B)
(C)
14
9. T-25 型産生溶血毒素に対する EGCg による溶血阻害態度
溶血毒素の中和実験ではEGCg を 1mg 添加することで 9 割以上の溶血が阻害され、0.5mg 添加では弱い阻害を確認することができた(Fig.9)。
Fig.9 Inhibition of hemolysin by epigallocatechin gallate.
10. McFarland 比色法による T-25、T-28 および T-B3264 の増殖曲線
3 菌型全てが培養開始 3 時間で対数増殖期が確認され、菌数量は T-28、T-B3264 および T-25 の順で減少した(Fig.10)。
15
考察
国立感染症情報センターの発表によると、2005 年に流行した A 群レンサ球菌の T 血清型 の主要菌型はT-12,1,4 および 3 型の順であり、食中毒が発生した 7 月の主要菌型は T-12, 1,3 および 28 型の順であった。したがって、本食中毒を惹起した T-25 型は決して主要な 流行菌型ではなかった。しかし、T-25 型は 1999 年をピークとして急激な変動が見られる (Infectious Agents Surveillance Report, 2000)ため、今後の動向に注意する必要がある。な お、Suzuki and Nikkawa(2006)の報告によると本食中毒の患者、食材および調理従事者か らの分離菌株はT-25 型、発赤毒素speB遺伝子単独保有株であり、パルスフィールドゲル 電気泳動法によるDNA パターンは全て同一であった。 ここでは、T-25 型と他の食中毒患者由来株の病原因子の性状を比較する目的で T-25 型、 T-28 型および T-B3264 型の 3 菌型を対象とした。Fig.1 で示すように 3 菌型全てが 2-ME による還元剤存在下での力価が高値であった。また、T-25 型の産生する溶血毒素の溶血力 価はT-28 型および T-B3264 型と比較して著しく低値であった。T-25 型の溶血毒素の性状は Fig.2 で示すとおり SLO であると確認された。すなわち、SLO は免疫原性がありγ-グロブリンによって中和され、赤血球膜との結合部位がコレステロー ルであるためコレステロールによっても阻害されることが示された。一方、T-28 型および T-B3264 型の主要な溶血毒素である SLS は免疫原性がないため、in vivoではSLO より広 範囲に細胞障害を惹起すると考えられる。さらに、溶血力価もT-25 型と比較して著しく高 く、溶血毒素による生体への影響はT-28 型および T-B3264 型のほうが大きいと推察される。 食中毒由来株の産生する代謝物質のプロファイルは Table 1 に示すように菌株によって 病原因子の産生量および性状に多様性が認められた。T-25 型の溶血効率は 5.59 であり、T-28 型およびT-B3264 型は 3.11 および 2.17 であった。SLO の溶血効率は 6.83±1.35 であると 報告(Suzuki et al., 1988)されており、溶血効率の結果からも T-25 型の主要な溶血毒素は SLO であることが示され、阻害態度の結果と一致した。T-25 型の SpeB 活性は 4.8U/mL であり、T-B3264 型と比較して 1/2 倍であった。SpeB はシステインプロテアーゼであるが、 SpeB は補体成分 C3b を分解し,重症化に関与すると報告(Terao et al., 2008)されている。 また、微生物プロテアーゼは直接的な生体影響に加え、生体内のプロテアーゼインヒビタ ーを不活化させることにより内因性プロテアーゼを活性化させ、間接的に様々な病態発現 に関与する(前田, 1995)。このように SpeB の作用は多面的であり生体に与える影響は大き いと考えられる。NADase 活性は T-25 型が最も高く 19.1Unit であったが、NADase はin vivoにおいてSLO と相乗作用があることが報告(Bricker et al., 2005; Athanasios et al., 2006)されており、病態へ大きく関与すると考えられる。
16
代謝物質のSDS-PAGE 分析では Fig.4 に示すように 3 菌型で主に培養開始 6 時間から 18 時間にかけてSpeB 前駆体から活性型 SpeB への変換を確認することができた。S.pyogenes
においてSpeB遺伝子はPCR により 100%検出されると報告(Yu and Ferretti., 1991)され ているが、すべての菌株がSpeB を産生するとは限らない。しかし、用いた 3 菌型が SpeB を産生することを確認し、他の病原因子よりもはるかにタンパク質発現量が多いことが泳 動バンドより確認できた。培養開始6 時間において T-25 型は SpeB 前駆体から SpeB へ変 換され、T-28 型および T-B3264 型は変換されていないことが確認できた。T-25 型は活性型 SpeB への変換が T-28 型および T-B3264 型より早いことが確認された。なお、Fig.10 に示 すとおり、3 菌型の培養時における増殖曲線を検討した結果、T-25 型の増殖スピードはそ れほど速くはなかった。したがって、増殖スピードとSpeB の早期産生には関係がなかった。 SpeB の生成は自己触媒により N 末端側のタンパク質が離脱し、最終的に活性型 SpeB にな ると考えられるが、S.pyogenesのhigh-temperature requirement A (HtrA)が SpeB 前駆 体から活性型へ変換するのに間接的な役割を果たしているとの報告(Jason et al., 2007)も あり、HtrA が SpeB の活性化へ関与している可能性がある。また、活性型 SpeB の産生に よりSLO の溶血活性が低下することが報告(小林、浜田, 1981; 鈴木等., 1997)されており、 T-25 型の SLO は培養 6 時間で SpeB の影響によりタンパク質分解を受け、T-28 型および T-B3264 型と比較して著しく溶血力価が低かったと考えられる。培養 18 時間において 3 菌 型で6 時間と比較し、いずれも SpeB が高産生であり、最も SpeB 活性が低値であった T-28 型のSpeB タンパク質バンドが最も狭いことが確認できた。T-B3264 型は最も SpeB 活性が 高かったにも関わらず18 時間において活性型 SpeB のタンパク質バンドが T-25 型と同程 度であった。これはT-25 型が活性型 SpeB への変換が早いために自己分解によって失活し 始めていた可能性が考えられる。 これまでT-28 型および T-B3264 型の産生する溶血毒素およびプロテアーゼについて報告 (鈴木、坂口, 2009)されているが、今回、NADase を加え、T-25 型について併せて検討した。 食品を介して惹起した3 菌型による食中毒の臨床症状は通常の A 群レンサ球菌感染症の病 態と同様であり、主症状は咽頭痛、発熱および関節痛等である。3 菌型間の病原因子を比較 すると、溶血活性はT-28 型、SpeB 活性は T-B3264 型、NADase 活性は T-25 型が最も高 かった。各々病原因子間の相互作用により、病態へ複雑に関与していることが推察される。 食中毒の発症率はT-25 型、T-28 型および T-B3264 型で 44.6%、66.8%および 55.8%であっ たが、溶血活性が高いほど発症率が高くなっており、溶血毒素の細胞障害により上気道感 染が進行したと考えられる。これらの成績より、食中毒由来株の産生する病原因子および 性状には多様性があることが確認された。
17
T-25 型において主要な病原因子の発現時期を特定する目的で培養開始 2 時間から 20 時間 までの病原因子の発現量および活性量について検討した。Fig.5 に示すように代謝物質の SDS-PAGE 分析では時間とともに SpeB 前駆体から活性型 SpeB への変換過程を確認する ことができ、変換のピークは14 時間であった。一方、SLO および NADase については CBB 染色では確認できないため、WB 法を用いて分析した。Fig.6-A に示すように SLO は時間 とともに70kDa から 60kDa にシフトしていくことが確認できた。この分子量の異なる SLO については等電点の異なるSLO の存在が報告(Bhakdi et al., 1985; Suzuki, 2009)されてお り、本結果には示していないが、等電点既知のSLO を用いて同様の分子量を各々確認して おり、70kDa が等電点 Acidic type の SLO(pI6.0)であり、60kDa が Neutral type の SLO(pI7.5)と考えられる。これら 2 hemolytic form の SLO の特徴は溶血活性能に差がない との報告(Weller et al., 1996)もあるが、溶血効率において異なるとの報告(Suzuki et al., 1988; Suzuki 2009)もある。70kDa から 60kDa の SLO への変化はプロテアーゼによって 分解されると考えられる。Fig.6-B に示すように NADase は WB によって T-25 型で産生さ れることが確認できた。Stevens et al.は 1988 年以前に分離された全ての株は NADase 陰 性であるが、1988 年から 1997 年の間にアメリカで分離された M-1 型株の全ては NADase 陽性であると報告(Stevens et al., 2000)している。さらに、NADase は STSS を含む病態に 関係があることが報告されている(Bricker et al.,2005)。今後は食中毒由来と STSS の同一 菌型でNADase の性状が異なるのか否かの検討が課題である。 SLO、SpeB および NADase の活性量を測定し、タンパク質分析の結果と比較を行なっ た。Fig.8-A に示すように SLO による溶血活性はタンパク質分析の結果と一致することを 示し、ピークは 8 時間であった。また、Fig.8-B に示すように SpeB は前駆体から活性型 SpeB へ変換されるピークである培養開始 14 時間から活性が認められた。さらに、培養 20 時間以降にSpeB 活性が増加するのか減少するのか検証するため、SpeB 分析については 30 時間まで培養時間を延ばして活性測定およびタンパク質分析を実施した。その結果、24 時 間の時点で 2.04U/mL と最大活性が認められ、それ以降はほぼ一定であった。これはタン パク質分析の結果と一致した。これまでに活性量においてSLO と SpeB が反比例すること は報告(小林、浜田, 1981; 鈴木等., 1997)されているが、タンパク質分析によってもこれら の関係を明らかにすることができた。Fig.8-C に示すように NADase において活性のピーク は10 時間、タンパク質分析のピークは 12 時間であり、一致する結果が得られなかった。 NADase 分析については NADase がきわめて微量で活性測定、タンパク分析の精度の影響 も大きいと考えられる。 Fig.7 に示すように病原因子の発現時期と病態の関連性では、まず SLO が産生されるこ
18
とで上皮粘膜細胞障害によって咽頭痛が惹起したと考えられる。その後、NADase の産生 によりSLO との相乗作用(Bricker et al., 2005; Athanasios et al., 2006)、すなわち、細胞 障害および代謝障害を惹起し、さらにSpeB が産生されることにより主症状である発熱およ び関節痛を惹起したと推察される。なお、M タンパク質と宿主結合組織の共通抗原性によ る抗原抗体複合体形成やプロテアーゼによるキニン産生カスケードの活性化によるキニン 産生(Maeda, 1995)も関節痛の病態に影響したと考えられる。Fig.9 に示すように EGCg に よって用量依存的に溶血毒素が中和された。溶血毒素であるSLO と赤血球の結合には細胞 膜コレステロール 3β-OH 基と 17 位の疎水性側鎖が関与していると報告(D.Prigent and J .E.Alouf, 1976)されている。EGCg はガレート基をもっており、カテキン類の中でもより OH 基を有することから SLO と EGCg が結合することで赤血球に結合できなくなり溶血を 阻害すると考えられる。病原因子として最初に起因菌によって発現されるSLO を中和する ことによって、その後に産生されるNADase との相乗作用を抑制し、結果として病態を悪 化させないことが期待される。 今後、これら病原因子間の生体影響については in vivo実験、病原因子の同時あるいは時 間差投与による影響を検討することが課題である。
19
要旨
【目的】 近年、食品を介して起こる感染症は大きな社会問題となっており、食品媒介 A 群レンサ 球菌感染症の集団発生の報告が散見され、わが国においても1990 年代以降 7 例の食品媒介 レンサ球菌感染症の報告がある。しかしながら、疫学が中心であり、病態と病原因子の関 係性、菌型間における病原因子の産生量の比較についての報告は見受けられない。原因菌 の産生する病原因子の検討は病態解析にとって必要な課題であり、発現時期を知ることは 病原因子間の相互関係を理解するために重要である。著者は2005 年 7 月に神奈川県下の大 学オープンキャンパスで集団発生した食品媒介レンサ球菌感染症、いわゆる集団食中毒事 例を対象とし、病原因子の解析を行なった。これまでに報告のある T-28 型(東京)および T-B3264 型(千葉)の 2事例と比較する目的で起因菌Streptococcus pyogenes T-25型(神奈川) の産生する主要な病原因子である溶血毒素、cysteine protease streptococcal pyrogenic exotoxin B (SpeB)および NAD glycohydrolase (NADase)の活性量と溶血毒素の性状を検討 し、それら病原因子のSDS-PAGE 分析を行なった。さらに T-25 型の産生する主要な病原 因子であるstreptolysin O (SLO)、SpeB および NADase の活性量およびタンパク質量につ いて比較し、各種病原因子の産生時期について検討した。 【方法・結果】 1. まず、各種病原因子の活性測定を行なった。溶血毒素の溶血力価は T-25 型 9HD50/mL であり、T-28 型 173HD50/mL および T-B3264 型 147HD50/mL と比較して著しく低 値であった。3 菌型全てが産生する溶血毒素は還元剤による依存性を示したが、T-25 型の溶血毒素はリン脂質で阻害されず、γ-グロブリンおよびコレステロールで阻害 されることからSLO の性状が示され、T-28 型および T-B3264 型の streptolysin S の 性状とは異なった。T-25 型の SpeB 活性は 4.8U/mL であり、T-B3264 型より低値で あったが、NADase 活性は 3 菌型中で最も高く 19.1Unit であった。 2. 次に各種病原因子の SDS-PAGE によるタンパク質分析を行なった。培養 6 時間で T-25 型が SpeB 前駆体から活性型 SpeB に変換しており、T-28 型および T-B3264 型 では変換されていないことが確認された。培養18 時間において 3 菌型で SpeB が高 産生であった。3. T-25 型の産生する主要病原因子(SLO, SpeB, NADase)について 2~30 時間までの培 養上清を用いて活性とウェスタンブロット(WB)分析を行なった。WB 分析により
20
SLO、SpeB および NADase を免疫化学的に同定した。SLO は 70 および 60kDa に 2 hemolytic form を確認できた。SpeB は 44kDa に前駆体、30kDa に活性型 SpeB が確認された。また、NADase は 50kDa に確認された。病原因子発現のタイムコー スを見るとSLO タンパク質のピークは培養開始 8 時間であり、活性のピークと一致 した。SpeB タンパク質は前駆体から活性型へ変換されるピークが 14 時間であり、 活性が認められ始めた時期と一致した。また、活性型SpeB タンパク質のピークは 24 時間であり、活性のピークと一致したが、それ以降ほぼ一定の産生量であった。一方、 NADase タンパク質のピークは 12 時間であったが、活性のピークである 10 時間と 一致しなかった。 4. T-25 型の溶血毒素である SLO はエピガロカテキンガレート(EGCg)によって溶血活 性を阻害されることが明らかとなった。 【考察】 レンサ球菌食中毒由来株である T-25 型の主要溶血毒素は SLO であり、T-28 型および T-B3264 型の SLS とは異なっており、活性量も著しく低値であった。また、SpeB 活性、 NADase 活性ともにそれぞれの菌型で多様性があり、解析した 3 つの病原因子については 食中毒に特徴的なものは確認されなかった。SDS-PAGE 分析で T-25 型は他の 2 菌型と比 較して SpeB タンパク質の発現が早期であった。T-25 型の増殖スピードが T-28 型および T-B3264 型と比較して速いためと思われたが、3 菌型の培養時における増殖曲線を検討した 結果、T-25 型の増殖スピードは著しく速いわけではなかった。したがって、増殖スピード とSpeB の早期産生には関係がなかった。T-25 型は最も活性型 SpeB への変換が早いため、 SpeB のプロテアーゼ作用によって SLO タンパク質が分解を受け、溶血活性が著しく低か ったと考えられる。3 菌型の溶血毒素の性状および主要病原因子の活性量には多様性がある ことが明らかになり、病態には病原因子の相互作用が関与していると推察される。 次に T-25 型において病原因子のタンパク質発現と活性のタイムコースを見ると SLO が 培養8 時間でピークとなり、活性型 SpeB のピークは 24 時間であったが、それ以降ほぼ一 定であった。一方、NADase タンパク質のピークは 12 時間であったが、活性のピークであ る10 時間と一致しなかった。NADase はきわめて微量であるため測定法の精度を向上させ ること等を検討することでより正確な解析が可能となると考えられる。これまでに活性量 においてSLO と SpeB が反比例することは報告されているが、SDS-PAGE によるタンパク 質分析によってもこれらの関係を明らかにすることができた。
21
膜細胞障害によって咽頭痛が惹起したと考えられる。その後、NADase の産生により SLO との相乗作用によるSLO 病原性の増大を惹起し、さらに SpeB が産生されることにより主 症状である発熱および関節痛を惹起したと推察される。
EGCg は SLO と結合することで結果として SLO が赤血球に結合できなくなり溶血を阻 害すると考えられる。EGCg にはそもそも殺菌作用もあるが、病原因子として最初に発現 されると思われる細胞障害性毒素であるSLO を中和する能力は起因菌の生体への侵入抑制 効果やその後に産生されるNADase との相乗作用抑制等、結果として病態を悪化させない ことが期待される。
22
引用文献
1. Andreas Berge ,Lars Björck. (1995) Streptococcal cysteine proteinase releases biologically active fragments of streptococcal surface proteins. J Biol Chem. 28:9862-7.
2. Asteberg I, Andersson Y, Dotevall L, Ericsson M, Darenberg J, Henriques- Nordmark B, Söderström A. (2006) A food-borne streptococcal sore throat outbreak in a small community. Scandinavian journal of infectious diseases.38:988-94.
3. Athanasios Michos, Ioannis Gryllos, Anders Håkansson, Amit Srivastava, Efi Kokkotou, and Michael R. Wessels. (2006) Enhancement of Streptolysin O Activity and Intrinsic Cytotoxic Effects of the Group A Streptococcal Toxin, NAD-Glycohydrolase. J.Biol.Chem. 281:8216-23.
4. Bhakdi, S., J. Tranum-Jensen, and A. Sziegoleit. (1985) Mechanism of membrane damage by streptolysin O. Infect. Immun. 47:52-60.
5. Bricker,A.L., Carey,V.J., and Wessels,M.R. (2005) Role of NADase in Virulence in Experimental Invasive Group A Streptococcal Infection. Infect.Immun. 73:6562-6. 6. Dennis L. Stevens, Daniel B. Salmi, Eric R. McIndoo, and Amy E. Bryant. (2000)
Molecular Epidemiology of nga and NAD Glycohydrolase/ADP-Ribosyltransferase Activity among Streptococcus pyogenes Causing Streptococcal Toxic Shock Syndrome. J Infect Dis. 182:1117-28.
7. D. Prigent and J. E. Alouf. (1976) Interaction of streptolysin O with sterols. Biochim. Biophys. Acta. 443: 288-300.
8. Jason N. Cole, John A. Aquilina, Peter G. Hains, Anna Henningham, Kadaba S. Sriprakash, Michael G. Caparon, Victor Nizet, Malak Kotb, Stuart J. Cordwell, Steven P. Djordjevic and Mark J.Walker. (2007) Role of group A Streptococcus HtrA in the maturation of SpeB protease. Proteomics. 7:4488-98.
9. Kaluski DN, Barak E, Kaufman Z, Valinsky L, Marva E, Korenman Z, Gorodnitzki Z, Yishai R, Koltai D, Leventhal A, Levine S, Havkin O, Green MS.(2006) A Large Food-borne Outbreak of Group A Streptococcal Pharyngitis in an Industrial Plant Potential for Deliberate Contamination. The Israel Medical Association journal.8:618-21.
10. Kusama H, Ohashi M, Shimazaki H, Fukumi H, (1958) Studies on streptolysin O and antistreptolysin O. Spectrophotometric determination of the hemolytic activity of streptolysin O by the fifty per cent end-point titration. Jpn.J Med Sci Biol. 11:347-57.
23
11. Laemmli UK. (1970) Cleavage of structural protein during the assembly of the head bacteriophage T4. Nature. 227:680-5.
12. Pournaras S, Efstratiou A, Douboyas J, George RC, Tsakris A. (2000) Genetic relatedness of group A streptococci of the newly designated serotype M90 causing a food-borne outbreak and sporadic infections. Epidemiol Infect. 125:299-301.
13. Shiseki M, Miwa K, Nemoto Y, Kato H,Suzuki J, Sekiya K.,(1999) Comparison of pathogenic factors expressed by group A Streptococci isolated from patients with Streptococcal toxic shock syndrome and scarlet fever. Microbial Pathogenesis. 27:243-52.
14. Streptococcal infections in Japan, 1996-2000. (2000) Infectious Agents Surveillance Report. 21:240-1.
15. Suzuki J. (2009) Characterization of acidic and neutral streptolysin O. J Electrophoresis. 53:45-50.
16. Suzuki J, Kobayashi S, Kagaya K, Fukazawa Y. (1988) Heterogeneity of hemolytic efficievcy and isoelectric point of streptolysin O. Infect.Immun. 56:2474-8.
17. Suzuki R and Nikkawa T. (2006) A Food-borne epidemic of Group A Streptococcal Pharyngitis. Bull.Kanagawa Ins.of P.H. No.36:12-3.
18. Tanaka D, Shima T, Isobe J, Watahiki M, Matsumoto M, Endoh M, Okuno R, Ogata K, Nagai Y. Epidemiology and Molecular Analysis of Group A Streptococci from Patients Involved in Food-Borne Disease Outbreaks in Japan between 1996 and 2003. (2006) Japanese journal of infectious diseases. 59:202-3.
19. Tatsuno I, Sawai J, Okamoto A, Matsumoto M, Minami M, Isaka M, Ohta M and Hasegawa T. (2007) Characterization of the NAD-glycohydrolase in streptococcal strains. Microbiology. 153:4253–60.
20. Terao,Y.,Mori,Y.,Yamaguchi,M.,Shimizu,Y.,Ooe,K.,Hamada,S.,Kawabata,S. (2008) Group A streptococcal cysteine protease degrades C3(C3b) and contributes to evasion of innate immunity. J.Biol.Chem. 283:6253-60.
21. Wayne L. Hynes and John R. Tagg. (1985) A simple plate assay for detection of group A streptococcus proteinase. Journal of Microbiological Methods. 4:25-31.
22. Weller U, Müller L, Messner M, Palmer M, Valeva A, Tranum-Jensen J, Agrawal P, Biermann C, Döbereiner A, Kehoe MA, Bhakdi S. (1996) Expression of active streptolysin O in Escherichia coli as a maltose-binding-protein--streptolysin-O fusion protein. Eur J Biochem. 236:34-9.
24
analysis of a scarlet fever outbreak among adults through food-borne transmission in China. The journal of infection.55:419-24.
24. Yu CE and Ferretti JJ. (1991) Frequency of the erythrogenic toxin B and C genes (speB and speC) among clinical isolates of group A streptococci. Infect. Immun. 59:211-5. 25. Jean F. MacFaddin(1985).病原細菌の生化学的検査法 原著第 2 版.p. 425. 竹田美 文他監訳.医学書院. 26. 小林貞男、浜田暁子(1981).猩紅熱患者由来 A 群溶連菌の蛋白質分解酵素産生と M 抗 原量,ストレプトリジンO 価および菌型について.感染症学雑誌.56:20-5. 27. 前田浩(1995).微生物プロテアーゼの病原性に関する研究.日本細菌学雑誌. 50:921-36. 28. 鈴木潤、坂口和子(2009).集団発生した食品媒介レンサ球菌感染症原因菌の産生する病 原因子.感染症学雑誌.83:380-5. 29. 鈴木潤、吉原英児、小林貞男(1997).劇症型 A 群レンサ球菌感染症患者分離株の菌体外 産生毒素について.化学療法の領域.13:45-50.
25
第Ⅱ部
病原性酵母における抗真菌剤標的候補の探索・同定
緒論
今日の医療の目覚しい進歩は、難病患者の生命を長らえたばかりでなく、不治と思われ ていた病気をも克服して社会復帰さえも可能としている。しかしながら、その反面、抵抗 力の弱まった患者(易感染性宿主)が増加する結果となり、これらの患者への日和見真菌 感 染 症 が 医 療 上 の 深 刻 な 問 題 の 一 つ と な っ て き て い る (Laniado-Laborín and Cabrales-Vargas, 2009; Cassone and Cauda, 2012; Parize et al., 2012; Pfaller, 2012; Sipsas and Kontoyiannis, 2012)。Candidaは、ヒトの口腔や腸・膣・皮膚等に常在してい る酵母様真菌であるが、病原性酵母としても知られている。先に示した易感染性宿主にお いては、ヒト常在菌のCandida属が起因菌となるカンジダ症は深刻であり、医療現場では 早急な対策が切望されている (Ascioglu et al., 2002; Gupta and Tomas, 2003; Pfaller and Diekema, 2007; Andes et al., 2012; Gow and Hube, 2012; Testoni et al., 2012)。Candida 属酵母には様々な菌種が存在し、形態学的には出芽酵母 Saccharomyces cerevisiae に類似しているが、病原性 Candida 属酵母として知られているものは通常 C.albicansのように二倍体で増殖するが、C.glabrataのように一倍体で増殖するものも存 在する。Candida 属酵母の病原因子としては、感染の第一段階である上皮細胞への粘着・ 付着に関与する粘着因子、次に、Candida 感染の特徴である細胞内寄生性を担うカタラー ゼ、また、菌の組織への侵入を容易にするプロテアーゼや、環境条件により酵母形から菌 糸形へと形態を変化させる二形性(とくにC.albicansの場合)などが知られている(Mayer et al., 2013)。化学療法の観点からみると、これらの因子を抑制・不活化させることにより 予防や治療が可能になると考えられる。しかしながら、Candida の病原因子をたとえ抑制・ 不活化できたとしても菌の増殖・生存そのものが奪われるわけではなく、より根本的には 菌の増殖能そのものを担う分子を標的として選択・同定することが重要な課題となる。 分子生物学的領域の研究が最も進んでいる S.cerevisiae ではゲノム配列がすでに決定 され、その遺伝子の約10%が菌の生育に不可欠な、いわゆる必須遺伝子とされている。ま
た、C.albicans (Jones et al., 2004) とC.glabrata (Dujon et al., 2004) の両菌種において
はすでに全ゲノムシークエンスが決定され、ポストゲノム、すなわち遺伝子機能の解析が 研究の焦点となりつつある。
とくに、C.glabrata の一倍体としての性質は遺伝子解析が容易である点で大変貴重で
26
的候補を分子生物学的観点から効率よく探索するため、そのツールとしてC.glabrataの一 倍体としての性質に着目した。この一倍体 C.glabrata における温度感受性変異株 (temperature-sensitive mutant; 以下 TS-株)を用いて必須遺伝子の探索・同定を行なう ETS system は宮川らによって確立され、「TS 変異は通常、ゲノムにおける必須遺伝子の point mutaion によって起こる」という概念に基づいている(Miyakawa et al., 2009)。
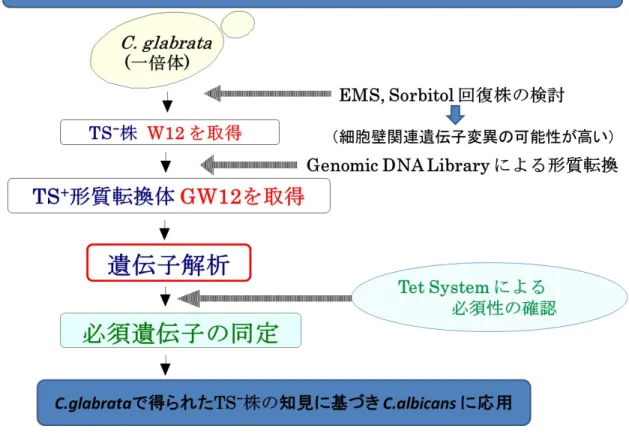
具体的な方法としては下記の戦略(図 1) に従い、本菌の生存に不可欠な必須遺伝子に変 異をもつ突然変異株をTS-株として分離した(第 1 章)。次に、各 TS-に対する相補活性(TS+)
を保有する遺伝子断片を分離するため、まず、親株由来のGenomic DNA Library を構築 した(第 2 章)。次にこれを Donor DNA(供与体)、各 TS-株をHost(宿主)として形質転 換を行なうことにより TS-を相補する TS+遺伝子断片を分離・同定 (第 3 章) し、さらに
得られた相補性遺伝子断片の詳しい解析や TS-株の TS 変異部位の解析等を試みた(第 4 章)。また、C.glabrata で得られた知見に基づいて、C.albicans における同様の point mutation の導入から TS-株の構築を試み、さらに、null mutant が取得できるか試みた(第 5 章)。新規抗真菌剤の標的候補探索の観点から選択毒性を考慮し、Cell wall integrity に 関与する遺伝子であるか検証した(第 6 章)。最後に、得られた TS-を相補するDNA 断片の 必須性を立証するため、テトラサイクリン (Tet) 応答性遺伝子発現制御系 (TET System) を用いた解析を行った(第 7, 8 章)。この戦略に従って、当研究室ですでに細胞周期制御遺 伝子TEM1 (Miyakawa et al., 2009) など多様な遺伝子が必須遺伝子として分離・同定さ れている (Miyakawa, 2013)。
本研究で着目する遺伝子は、本菌の生存に不可欠であるため、生物学的意味においてき わめて重要であるだけでなく、抗真菌剤開発の面において、この遺伝子(およびその遺伝 子産物)は薬剤の標的候補となることが期待されるものであり、医療の分野においても重 要な意義をもつものと考えられる。
27
28
第
1 章
【TS
-株の分離と性状解析】
緒言
本章では、緒 論で述べた基本構想の下に、一倍体で増殖することが知られている C.glabrataを親株として用いて,温度感受性変異株 (TS-株) の分離を試みた。必須遺伝子 に変異をもつ突然変異株は、条件致死変異株としてのみ分離可能である。現在までに条件 致死変異株を温度感受性変異株として分離した例として、原核生物 Bacillus subtilis のTS341 (Miyakawa and Komano, 1981)や真核生物Schizosaccharomyces pombeのcdc2 (Lee
and Nurse, 1987)およびS.cerevisiaeのCDC28 (Beach et al., 1982) 等、菌の生育に必須 の細胞分裂関連遺伝子の欠損した温度感受性変異株が分離されている(Lindahl et al., 1971; Miyakawa et al., 1982; Sherman et al., 1986; Sadaie et al., 1991; Takamatsu et al., 1992; Ben-Aroya et al., 2008)。 本研究においては現在までに病原性酵母C.glabrata由来のTS-株を約300 株 取得することができた。
本章では、これらのTS-株の取得方法および得られたTS-株の性状解析の結果について述
べる。
材料
実験に用いた菌株は C.glabrata 2001-TH (Kitada et al., 1995)(trp1 his3)(中外製薬 より分与)を用いた。保存方法としては、YPD を 5 ml で 16 hr、静置培養したものを遠心 沈殿させ、1 ml の YPD に懸濁し、グリセロールと 1:1 の割合で混合させ、-20℃、-80℃ に各々凍結保存した。
培養は YPD 液体培地(Sherman et al., 1986)、SA 寒天平板培地(Kagaya et al.,1989)、 SA+Sorbitol 寒天培地を以下の表 1 のように作製し用いた。
表1 YPD、SA 培地組成
培地組成 YPD Sabouraud (SA) SA+Sorbitol (Sorb+)
Yeast extract 10 g 5 g 5 g Polypepton 20 g 10 g 10 g Glucose 20 g 20 g 20 g Agar (栄研) ― 15 g 15 g Sorbitol ― ― 182.5g Distilled water (以下 D.W.) up to up to up to 1000 ml 1000 ml 1000 ml オートクレーブ 121℃ 20 min
29
実験方法
1) 生菌数測定と発育制限温度の設定
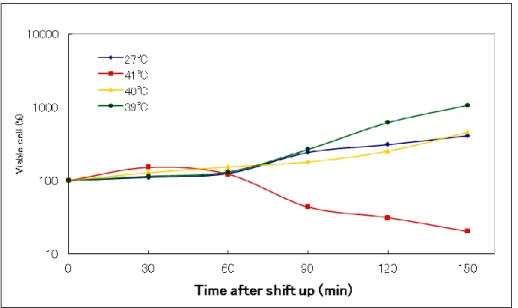
発育許容温度27℃における対数期の培養を OD600 0.1(Time 0)で各温度;27、39、40 および41℃にシフトした後、震とう培養し、Time 0、30、60、90、120 および 150 min で培養液を適当に希釈し、SA 寒天平板培地に Plating した。これを 27℃で一夜培養後に形 成されたコロニー数から生菌数をc.f.u./ ml として算定した。結果は、温度シフト後の c.f.u. をTime 0 における c.f.u.に対する割合として% c.f.u.で示した。また、各温度における培養で、親株の生育が可能な温度を基準として、発育制限温度を 設定した。
2) TS
-株の分離とその性状解析
1. 突然変異誘発剤 EMS 処理 まず親株をYPD 液体培地に植菌し、27℃にて一夜培養後、その一部を 5 ml の新鮮な YPD 液体培地を含む L 字管中で希釈し、同温度にて恒温槽中で振とう培養した。その後、対数 増殖期OD600 0.1 で突然変異誘発剤 ethyl methane sulfonate (EMS) により 0.2M, 60 分処理した。EMS は弱い変異発剤であり、1-methyl-1-nitro-nitrosoguanidine (MNNG) のよ うな他の強い変異誘発剤ではフレームシフトする可能性が高くなり目的としている TS-株
の分離が難しくなるため、本研究ではEMS を用いた(Strauss, 1961; Strauss et al., 1968; Sherman et al., 1986)。
3) TS
-株のスクリーニング
EMS 処理後、SA 平板培地に Plating し、菌の残存生存率が 50~20 %になる条件で得ら れたコロニーから、各コロニーをSA 寒天平板培地にレプリカ、27℃および発育制限温度で 一夜培養した。その結果、27℃においてのみコロニー形成能が認められたものについて、 Single Colony Isolation を行い、各コロニーについて、先と同様にレプリカ、その TS 性を 再度検討した。これによりTS 性が確認されたものを TS-株として保存し、実験に用いた。
結果と考察
C.glabrata からの温度感受性変異株の分離にあたり、発育制限温度を設定することが必 要となる。まず、親株であるC.glabrata 2001-TH 株の発育に対する温度の影響を検討した。 2001-TH 株の 27℃における対数増殖期の培養を各温度;27、39、40、41℃、にシフト後、 生菌数変化を測定した(図 1-1)。30
図1-1 親株の高温シフト後の生菌数変化
C.glabrata 2001-TH の 27℃で対数増殖させ、OD600 0.1 (Time 0) で図に示す各温度にシ
フト後、経時的に各々の一部を採取、稀釈後、寒天平板倍地に塗布し、27℃で一夜培養し た。Time 0 における c.f.u. に対する割合(%)としてシフト後の c.f.u. を算定した。
その結果、39℃および、40℃にシフトした場合には 27℃と同様かそれ以上の% c.f.u.の増 加が認められた。一方、41℃にシフトした場合は培養経過にともない% c.f.u.が、始めの約 30 分までは 27℃のときよりも急速に増加したが、その後は徐々に減少していくことが明ら かになった。このことは、もし、変異株の分離にあたって発育制限温度を 41℃に設定した 場合、この温度では親株でさえ正常に発育できないことを意味する。また、40℃の場合は、 この図に示すように親株は確かに39℃の場合と同様に c.f.u.の増加が認められるが、一夜培 養後のコロニーが 39℃に比べ小さかったこと、また、インキュベーターの庫内温度が設定 温度に+/- 約 0.5℃の誤差があること等の問題が考えられた。したがって、これらの結果か ら、変異株を分離するにあたっての発育制限温度を39℃に設定した。 TS-株のスクリーニ ングは、EMS 処理後、発育許容温度である 27℃で SA 寒天平板培地に培養して出現したコ ロニーを27℃と 39℃にレプリカした。その結果、39℃において発育が見られなかったコロ ニーにつき、27℃のコロニーを Single Colony Isolation して、その約 50 コロニーを再び 27℃と 39℃にレプリカした。27℃では全てのコロニー形成が認められ、39℃ではいずれも コロニー形成されなかったものをTS-株として分離した。この方法で、これまでに約300 株
31
また、温度感受性試験において、39℃でコロニー形成が認められる株は、自然復帰突然 変異株 (Spontaneous revertant)が出現していることが考えられる。そのため各変異株につ いて復帰突然変異 (Reversion) の頻度を検討し、発育制限温度 39℃での Revertant への復 帰突然変異率がきわめて低い(10-5以下)ものを用いた。これにより、Genomic DNA Library
(第 2 章) を用いた形質転換実験により TS-を相補する(TS-をTS+に復帰させる)DNA 断片を
32
第
2 章
【
C.glabrata
Genomic DNA Library の構築】
緒言
第1 章より、C.glabrata を EMS によって突然変異誘発処理し、発育制限温度 39℃にお けるコロニー形成能を失ったTS-株(温度感受性変異株)を取得することができた。取得し
た TS-株は Reversion の頻度が十分低いことから,Host (宿主) として有用であり、
C.glabrata Genomic DNA Library からの相補性 (TS+) 遺伝子のスクリーニングが可能で
あると考えられる。したがって,本章では TS-株から発育制限温度 39℃におけるコロニー
形成能を回復したTS+株をスクリーニングするためのC.glabrata Genomic DNA Library
の構築を試みた。そのために,C.glabrataよりGenomic DNA を調製し、それを制限酵素
Sau3AI で Partial Digest することにより、TS-株の欠損遺伝子を相補する目的DNA 断片
(3〜12 kbp)を含む様々な DNA 断片を作製した。これら DNA 断片を Insert とし、Plasmid ベクター, pCgACH-3 (Kitada et al., 1996) にLigation し,大腸菌へ Transformation した。 得られた数千コロニーを集めて培養した後、Plasmid 抽出を行い、C.glabrata Genomic DNA Library を構築した。 こうして得られた DNA Library を Donor として TS-株(宿主)
を形質転換させ、発育制限温度39℃におけるコロニー形成能を回復した TS+株が得られれ
ば、その株は相補性 (TS+) 遺伝子の断片を保有している可能性が高いと考えられる。
材料
Library用にC.glabrata 2001-TH を、Library 確認用にC.glabrata 2001-THU (Nakayama et al., 1998) (trp1 his3 ura3)を使用した。
酵母の培養時にはYPD 液体培地と SA 寒天平板培地 (第 1 章)を用い、大腸菌の培養時に はLB 液体培地、LA 寒天平板培地を用いた (表 2-1)。また,Yeast transformation には形 質転換体と非形質転換体の区別をするためにYNB 寒天平板培地を用いた (表 2-2)。 表2-1 LB、LA 培地組成 Yeast extract 5 g Tryptone (bacto) 10 g NaCl 10 g D.W. up to 1000 ml pH 7.0 (with 1N NaOH ) 1.0 ml Agar (bacto) 15 g (LA のみ)
33 表2-2 YNB 培地組成 Difco-Agar 10 g Glucose 5 g D.W. up to 900 ml オートクレーブ 121℃ 20 min その後、10×YNB (濾過滅菌) を 100 ml 添加
実験方法
分子生物学上の基本的手法は定法 (Sambrook et al., 1989)に従った。1) C.glabrata Genomic DNA の調製
C.glabrata Genomic DNA Library 構築のため、以下の試薬 (表 2-3~2-6)を用い、酵母
菌体から定法 (Miyakawa, 2000)に従い全 DNA を調製した。 表2-3 0.5M EDTA (pH 7.5) EDTA-2Na・H2O 55.8 g NaOH 6.5 g D.W 245 ml Total 300 ml 表2-4 0.5M EDTA (pH 9.0) EDTA-2Na・H2O 9.3 g NaOH 1.5 g D.W 44.3 ml Total 50 ml
表2-5 Sol-E (1M sorbitol-0.1M EDTA)
Sorbitol 72.8 g
0.5M EDTA (pH7.5) 80 ml
D.W 270 ml
Total 400 ml
表2-6 Saline- EDTA (0.15M NaCl - 0.01M EDTA・2Na)(pH 8.0)
NaCl 17.5 g
0.5M EDTA・2Na (pH 8.0) 40 ml
D.W 1960 ml
34
2) C.glabrata Genomic DNA Library の調製
1.
C.glabrata
Genomic DNA の Partial DigestionC.glabrata Genomic DNA を Partial Digest した。TS-を相補する目的DNA 断片を網羅
するために3〜12 kbp の断片が最も多くなるように制限酵素濃度、反応時間を検討した。 まず,予備実験として、Library の構築に通常用いる制限酵素Sau3AI による Partial Digest の至適濃度,時間を検討した。この結果から得たデータをもとにし、至適酵素濃度、至適 反応時間を決定し、大量のDNA を用いて至適条件下で Partial Digest の後、アガロースゲ ル電気泳動により目的とする3〜12 kbp の断片を上 (9~12 kbp 付近)、中 (6~9 kbp 付近)、 下 (3~6 kbp 付近)の3領域に分けて抽出、次項で調製した Plasmid ベクターとの Ligation 反応に用いた。
2. Plasmid ベクターの調製
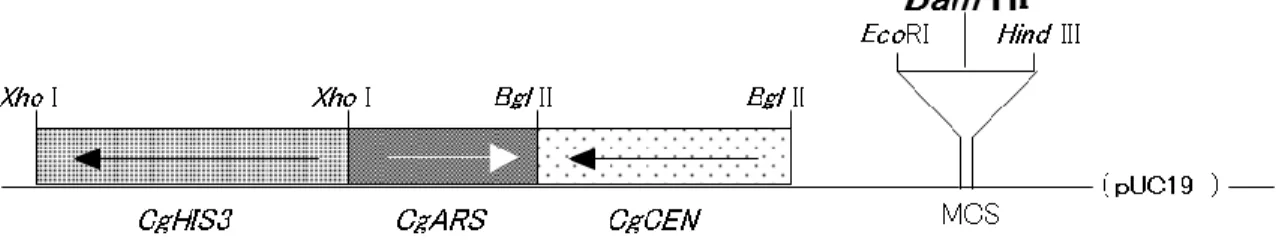
抽出したDNA 断片 3〜12 kbp を Insert (Donor DNA 断片) とし、ベクターとして使用 するpCgACH-3(Kitada et al, 1996) (図 2-1)に組み込む。この pCgACH-3 は、大腸菌用の Plasmid ベクターである pUC19 株にC.glabrata由来の3 つの遺伝子断片、CgHIS3 (His+
マーカー)、CgARS (Autonomous Replicating Sequence;自律的複製の塩基配列)、および
CgCEN (Centromere) が組み込まれた Shuttle ベクターであり、C.glabrata由来の遺伝子
を組み込んだ形質転換体の分離にきわめて有用なものである。
図2-1 Genomic DNA Library 構築に用いた Plasmid ベクター, pCgACH-3 の制限酵素地図
CgHIS3、CgARS および CgCEN は各々C.glabrata 由来の HIS3、ARS (Autonomous Replicating Sequence)およびCEN (Centromere) を、 MCS はベクター pUC19 のマ ル チ ク ロ ー ニ ン グ サ イ ト を 示 す 。 ま た こ の 図 に 記 載 さ れ て い な い が α -Complementation 用にlac Z が MCS を含んで構築されている。
35
たPlasmid ベクター(pCgACH-3)の MCS 内のBamHI サイトを制限酵素で切断 (表 2-7) した後、脱リン酸化した。Plasmid の分離・精製 (アルカリ SDS 法) (試薬:表 2-9, 10, 11)、 脱リン酸化は定法に従った (Sambrook et al., 1989)。
3. Ligation, 大腸菌 Transformation は定法(Sambrook et al., 1989) に従って行った。
表2-8 Ligation 反応液
insert + ベクター ベクターのみ ベクター (pCgACH-3) 4.5 kbp (5 ng/μl) 1 μl 1 μl
Insert 3~12 kbp (50 ng/μl) 1 μl - Ligation kit (TAKARA) 2 μl 1 μl
Total 4 μl 2 μl
Plasmid の精製における試薬の調製
表2-9 A 液 (Tris Glucose Buffer)
D.W. 7.7 ml Glucose 0.072 ml 0.5 M EDTA (pH8.0) 0.16 ml 2 M Tris (pH8.0) 0.1 ml Total 8.0 ml リゾチーム (0.8 mg/A 液 0.25ml )加え、攪拌する 表2-10 B 液 (Alkali-SDS) D.W. 3.7 ml 1 N NaOH 0.8 ml 10% SDS 0.5 ml Total 5.0 ml 表2-7 pCgACH-3 BamH1 制限酵 素処理 D.W. 60 μl BamHI 10 μl 10×[BamHI] buffer 10 μl DNA(pCgACH-3) (500 ng/μl) 20 μl Total 100 μl