近畿大学学術情報リポジトリ
53
0
0
全文
(2) Mechanism of Wood Decay by Brown-Rot Fungi Shohei Kaneko Graduate School, Kinki University Division of Agricultural Science (Advisor: Prof. Akio Enoki) September, 2004. lliiKt*¥*¥~a¥1iJfJi:l4 ptffl ~~{t¥W:r:z ~ -T l§ lJZ (fl:I~. : ;fI~N~~t5D. Submitted to the Graduate School, Kinki University, to fulfill the requirement for the Doctrate Degree..
(3) I wish to express my sincere gratitude to Dr. Akio Enoki, Professor of Faculty of Agriculture, Kinki University, for his kind guidance, valuable advice, stimulating discussion and critical review throughout the work including the manuscript of this thesis. Thanks are due to Dr. Tadashi Okamoto and Dr. Takao Terashita, Professor of Faculty of Agriculture, Kinki University, for reading the entire text in its original form. I greatly thanks to Dr. Hiromi Tanaka and Dr. Shuji Itakura, Professor of Faculty of Agriculture and Assistant proffesor of Faculty of Agriculture, Kinki University, for their kind advice, and valuable comments and discussion. Finally, Thanks are also due to all the past and present members of the laboratory of the Forest Products Chemistry, Department of Agricultural Chemistry, Faculty of Agriculture, Kinki University, for their kind cooperation and help..
(4) CONTENTS. CHAPTER. I. INTRODUCTION ............................................................................................................................................. 3 CHAPTER. n. HYDROXYL RADICAL PRODUCTION, OXALIC ACID CONCENTRATION, AND DEGRADATION OF WOOD, CRYSTALLINE CELLULOSE, AND A LIGNIN-RELATED COMPOUND IN CULTURES OF BROWN-ROT FUNGI......................................................................... 6 BACKGROUND .................................................................................................................................................. 6. MATERIALS AND METHODS .............................................................................................................................. 8. Fungal strains ............................................................................................................................................ 8 Media ......................................................................................................................................................... 8 Degradation of wood, cellulose, and a lignin model compound ............................................................. 8 Accumulation of oxalic acid ...................................................................................................................... 9 CMCase and Avicelase assays ................................................................................................................ 10 Hydroxyl radical production ................................................................................................................... 10 RESULTS ........................................................................................................................................................ 11. Degradation of Japanese beech wood ..................................................................................................... 11 Degradation of cellulose in glucose and wood cultures ........................................................................ 12 Degradation of a lignin model compound in glucose and wood cultures ............................................. 14 Oxalic acid concentrations in cultures ................................................................................................... 16 Hydroxyl radical production in glucose and wood cultures .................................................................. 17 DISCUSSION .................................................................................................................................................... CHAPTER. 19. m. PHYSICAL AND CHEMICAL PROPERTIES OF AN EXTRACELLULAR LOW-MOLECULARWEIGHT SUBSTANCE FROM THE BROWN-ROT BASIDIOMYCETE TYROMYCES PALUSTRlS ...................................................................................................................................................... 23 BACKGROUND ................................................................................................................................................ 23. MATERIALS AND METHODS ........................................................................................................................... 25. Organism ................................................................................................................................................. 25. 1.
(5) Culture conditions ................................................................................................................................... 25 Isolation and partial purification of the extracellular substance .......................................................... 25 Ethylene generation from KTBA ............................................................................................................. 26 Electrophoresis ........................................................................................................................................ 26 Electrotransfer ......................................................................................................................................... 27 Hydroxyl radical production ................................................................................................................... 27 Protein, amino acid, carbohydrate, and Fe analysis ............................................................................. 27 Fe( IlI)- and Cu( IJ)-reducing activity ...................................................................................................... 28 Determination of carbonyl groups .......................................................................................................... 28 Determination of aldehyde groups .......................................................................................................... 29 Determination of protein glycosylation .................................................................................................. 29 RESULTS AND DISCUSSION ............................................................................................................................. 30. Partial purification of the extracellular substance ................................................................................ 30 Composition and molecular weight ................................................................................. ....................... 30 Hydroxyl radical generation ................................................................................................................... 32 Amino acid, monosaccharide, and Fe content ....................................................................................... 34 Fe( lll)- and Cu( IJ)-reducing activity ...................................................................................................... 37 Carbonyls ................................................................................................................................................. 39 Glycosylation ........................................................................................................................................... 39 Proposed mechanism for wood decay by brown-rot fungi ..................................................................... 41. CHAPTER. N. CONCLUSION ................................................................................................................................................ 43 REFERENCES ................................................................................................................................................ 45. 2.
(6) CHAPTER I Introduction White- and brown-rot fungi are the wood destroying fungi and a group of basidiomycetes. Morphological aspects of wood degradation by white-rot fungi are different from those of brown-rot fungi. White-rot fungi are able to degrade cell wall components (cellulose, hemicellulose, and lignin). The hyphae of white-rot fungi grow inside the cell lumens and produce a gradual thining of the cell walls from the lumina toward the middle lamella as decay advances (Liese. 1970) (Fig. 1). White-rot fungi have a cellulolytic system that consists of a endo- f3 -1 ,4-glucanase, a exo- f3 -1,4glucanase, and a f3 -glucosidase. Endo- f3 -l,4-glucanase randomly attacks the cellulose. f3 -l,4-glycosidic linkages. Exo- f3 -l,4-glucanase attacks and hydrolyzes the crystalline region of the cellulose polymer, and f3 -glucosidase chain and splits. hydrolyze degradation products of cellulose to glucose. Eriksson suggested that the endo- f3 -1 ,4-glucanase act randomly the cellulose chain in wood, and open up chain ends where the exo- f3 -1,4-glucanase can act (Reese et al. 1950). White-rot fungi remove both the lignin and carbohydrate simultaneously (Kirk and Highley. 1973) or preferentially remove the lignin component (Kirk and Moor. 1972). Two extracellular heme peroxidases of white-rot fungi, lignin peroxidase and manganese peroxidase, are important in the degradation of the lignin component in wood (Tien and Kirk. 1983; Glen et al. 1983; Glen and Gold. 1985) because they can oxidatively degrade a variety of lignin-related components (Miki et al. 1987; Wariishi et al. 1989). However, these peroxidases have phenol oxidase activity, and polymerize lignin substrates rather than decomposing them (Haemmerli et al. 1986; Odier et al. 1988). Thus, other system to prevent or reverse the porimerization reaction are necessary for the actual depolymerization of lignin polymers in wood. However, the process of removal from cell wall is not completely understood.. Brown-rot fungi are known to cause the most serious damage to wood in service.. 3.
(7) Relatively little attention has been given to identify the mechanism of wood-degradation by brown-rot fungi and the mechanism are not fully understood. Brown-rot fungi can degrade crystalline cellulose in wood even though they lack exo1,4-.B -glucanase activity (Kirk and Highley. 1973). The fungi also degrade the lignin component in wood to a significant extent although the fungi preferentially remove both the polysaccharide components from the wood, leaving an amorphous, brown, crumbly residue that is composed largely of lignin (Enoki et al. 1988; Highley. 1987). Brown-rot fungi degrade crystalline cellulose under the conditions in which the fungi transform lignin-related compounds and brown-rot fungi are unable to degrade the cellulose substrate under the conditions in which the fungi are unable to transform. the. lignin-related compounds (Enoki et al. 1988). This indicates that brown-rot fungi may generate a unique wood-component-degrading system that can split cellulose as well as lignin into fragments. The extracellular one-electron oxidation activity of brown-rot fungi is proportional to degradation activities of the fungi against wood, crystalline cellulose and lignin related compound (Enoki et al. 1988). Some brown-rot fungi can degrade three 1,2-diarylethane lignin model compounds, yielding the same products in relatively high yields as those found in high yields in the degradation of the lignin model compounds by hydroxyl radicals (Espejo et al. 1990). Hydroxyl radicals are detected in cultures of brown-rot fungi in surprisingly high yields (Backa et al. 1992).. Brown-rot fungi cause rapid depolymerization of the cellulose in wood before losses in total wood substance are detected. The pattern of progressive change in the mean degree of polymerization of the holocellulose in wood caused by Fenton's reagent is similar to that caused by brown-rot fungi (Cowling. 1961; Koenigs. 1974). In the early stages of wood degradation, it is often observed that the S2 layer of the cell wall is extensively degraded while the S3 layer adjacent to the lumen remains relatively unaffected by hyphae from the lumen (Highley and Murmanis. 1985. Kuo. 1988). Ovalbumin (Mw 45,000) could not penetrate the cell walls of the wood decayed to 70% weight loss by the brown-rot fungus (Srebotnic and Messner. 1991). The maximum pore diameter in the cell walls of the wood severely decayed by brown-rot fungi never exceeded 38A (Mw 6,000) (Flournoy et al. 1991). Enzymes such as cellulase and peroxidase are still 4.
(8) too large to penetrate cell walls in the early stages of wood degradation by brown-rot fungi (Flournoy et al. 1991). These findings above mentioned suggest that brown-rot fungi extracellularly produce an one-electron oxidation system that is capable of penetrating the cell walls of wood, modifying the walls and that this one electron oxidation system includes hydroxyl radicals.. The aim of this study is to investigates the mechanjsm of wood degradation by brownrot fungi . I studied the following to provide important findings on wood degradation by brown-rot fungi; 1) Relationship between degradation of wood, cellulose or lignin-related compounds and production of hydroxyl radical or accumulation of oxalic acid in cultures of brown-rot fungi. 2) The chemical and physical properties of extracellular substance from the brown-rot Basidiomycete Tyromyces paiustris. 3) The possible role of a one-electron oxidation system involving hydroxyl radical in the degradation of wood, crystalline cellulose, and lignin by T paiustris.. Middle lamell l. ~~(~ L~~~~--~~. Primary wall. •. Hyphae. White-rot 1\mai. Brown-rot 1\mai. Fig. 1 Model for wood cell wall degradation mechanism by white-rot and brown-rot fungi.. 5.
(9) CHAPTER. n. Hydroxyl Radical Production, Oxalic Acid Concentration, and Degradation of Wood, Crystalline Cellulose, and a Lignin-Related Compound in Cultures of Brown-Rot Fungi Background. Brown-rot fungi preferentially degrade the cellulose in wood (Enoki et al. 1988), although they lack exo-l,4-/3 -glucanase activity for hydrolyzing crystalline or natural cellulose (Highley. 1988). However they degrade and metabolize cellulose only when their lignolytic systems are active (Enoki et al. 1988; Enoki et al. 1985). The oneelectron-oxidizing activity in cultures of brown-rot fungi is proportional to the degradation of lignin-related model compounds, natural cellulose substrates, and wood in the cultures (Enoki et al. 1989). These facts suggest that brown-rot fungi generate a unique system that is capable of modifying lignin and fully degrading natural cellulose.. Brown-rot fungi rapidly depolymerize the cellulose in wood before any loss in total wood mass is detectable (Cowling. 1961). Furthermore, fungal enzymes such as cellulases and peroxidases are too large to penetrate cell walls during the early stages of wood degradation by brown-rot fungi (Flournoy et al. 1991; Kuo et al. 1988). This suggests that a component of the brown-rot wood-degrading system is small enough to diffuse into the sound wood cell wall.. The pattern of holocellulose depolymerization in brown-rotted wood is similar to that caused by Fenton reagent (Cowling. 1961; Koenigs. 1974), and natural cellulose depolymerized by hydroxyl radical (·OH) is similar in chemical structure to brownrotted cellulose (Kirk et al. 1991). Furthermore some brown-rot fungi degrade ligninrelated model compounds to yield relatively large amounts of the same products that are obtained via ·OH-attack on the lignin model compounds (Espejo et al. 1990; Zohar et al. 1998). These findings suggest that ·OH may be a component of the brown-rot wooddegrading system.. 6.
(10) An extracellular low-molecular-weight substance that catalyzes a redox reaction between O 2 and an electron donor to produce ·OH via O 2- and H 20 2 has been isolated from wood-degrading cultures of the brown-rot fungus, T. palustris (Hirano et al_ 1995). Most of the extracellular ·OH produced in intact cultures of T. palustris are generated by this low-molecular-weight substance (Hirano et al. 1997). During early stages of wood degradation by T. palustris, the low-molecular-weight substance is found in the fungal cytoplasm and cell Wall, in the extracellular sheath surrounding the fungal cell wall, and throughout the wood cell wall, suggesting that it diffuses through the S3 layer into the S2 layer and the middle lamella (Hirano et al. 2000). In the early stages of wood decay by the brown-rot fungi, T. palustris and Coniophora puteana, H20 2 is detected in both the fungal cell wall and the wood cell wall (Kim et al. 2002). Thus, there is growing evidence that brown-rot fungi produce extracellular ·OH as part of their wooddegrading system.. However it is not known whether the production of ·OH is a universal characteristic of wood degradation by brown-rot fungi. Some brown-rot fungi exhibit high one-electronoxidation activity in cultures containing wood but little activity in cultures containing glucose as a carbon source. This one-electron-oxidation activity appears to result from. ·OH (Enoki et al. 1989). Early findings suggest that oxalic acid may be involved in wood decay by brown-rot fungi. Cultures of brown-rot fungi often contain significant amounts of oxalic acid; whereas cultures of white-rot fungi usually have little or no oxalic acid (Shimazono. 1955; Takao. 1965). Oxalic acid hydrolyzes the hemicellulose in wood, making the cellulose fibers more accessible to cellulases (Beck-Anderson. 1987). Oxalic acid also may initiate the depolymerization of amorphous cellulose directly (Shimada et al. 1991). Postia placenta produces sufficient oxalic acid to decrease the pH of the decaying wood, thereby promoting brown-rot degradation of the cellulose (Green et al. 1992).. Herein I examine the possible roles of ·OH and oxalic acid in the degradation of wood, crystalline cellulose, and a lignin-substructure model by seven fungi that are usually regarded as brown rots.. 7.
(11) Materials and methods. Fungal strains. The brown-rot fungi used for this study were Gloeophyllum trabeum (Pers. Ex Fr.) MUff. IFO 6268 (LZT), Tyromyces palustris (Berk. Et Curt.) MUff. FRI0507 (TYP), Laetiporus sulphhureus (Bull. Ex Fr.) Bond. Et Sting. IFO 6497 (LAS), Postia placenta (Mad.-698-R) (PPL), Daedalea dickinsii Yasuda FRI T 4b (DAD), Coniophora puteana (Schumacher ex Fr.) IFO 8764 (COP), and Lentinus lepideus (Fr.) IFO 8719 (LEL). Cultures were maintained through periodic transfer on slants as previously described (Takahashi. 1976).. Media. Basal agar medium was prepared as previously described (Enoki et al. 1988), except that ten-fold concentrated trace elements were used. Glucose cultures contained 2% glucose and wood-containing cultures contained 0.2% glucose.. Degradation of wood, cellulose, and a lignin model compound. The degradation of Japanese beech (Fagus crenata Blume) by each fungus was determined as previously described (Enoki et al. 1988), except that the basal agar medium of wood cultures contained 0.2% glucose instead of 1% glucose.. Cellulose degradation in glucose cultures of each fungus was determined as previously described (Enoki et al. 1988), except that the filter papers were of 5-cm diameter.. "Sandwich" cultures were used to measure cellulose degradation in wood-containing cultures. Sawdust of Japanese beech was extracted twice with acetone and dried. The sterilized sawdust (1.5 g), containing 60% distilled water, was sprinkled evenly over the surface of basal agar medium (50 ml) containing 0.2% glucose in 300-ml Erlenmeyer flasks. One filter paper (5 cm) was placed over the sawdust and another 1.5 g of sawdust was spread over the filter. Cultures were inoculated with a small piece of fungal. 8.
(12) mat and incubated at 28°C in air. After the indicated periods, the mycelia and sawdust were removed carefully from the filter paper with running water. The decayed filter papers were extracted once with acetone, once with distilled water, and again with acetone, dried, and weighed. Degradation of the lignin model compound, 4-ethoxy-3-methoxyphenyl- f3 -guaiacyl ether, in glucose cultures, was determined as described previously (Enoki et al. 1988).. For determining the degradation of a lignin model compound, 4-ethoxy-3methoxyphenylglycerol- f3 -guaiacyl ether in wood cultures, 2 g of sawdust, was sprinkled evenly over the surface of basal agar medium (50 ml) containing 0.2% glucose in 300-ml Erlenmeyer flasks. The lignin-model compound (2 mg in 200 /..11 acetone) was added to the surface. After two days, cultures were inoculated with a small piece of fungal mat and incubated at 28°C in air. Degradation was determined as previously described (Enoki et al. 1988). Each measurement was the average of triplicate cultures.. Accumulation of oxalic acid. Basal agar medium (50 ml in 300-ml Erlenmeyer flasks) contained either 2% or 0.2% glucose. Sawdust of Japanese beech (3g) was sprinkled evenly over the surface of the 0.2%-glucose cultures. The cultures were inoculated with a small piece of fungal mat and incubated at 28°C in air. At the incubated intervals, 100 ml of ethyl acetate and 0.2 ml of 1M HCl were added to each culture. The mixtures were stirred with a glass rod, and shaken for 30 min. Then the mixtures were centrifuged at 10,000 X g for 20 min. and filtered through filter paper. Each of the ethyl acetate layers was washed with distilled water (50 ml, two times). Ethyl acetate solution (50 ml) was taken out of each ethyl acetate layer and evaporated under reduced pressure. Trimethylsilanation of the residues was carried out by adding 1,(0) /..11 of bis-(N,O-trimethylsilyl)trifluoracetamidepyridene (1: 1) to each of the dry residues and heating at 80°C for 5 min. Gas chromatography was carried out with a glass column packed with 3% OV-101 on chromsorb WAW DMCS. The oven temperature was programmed from 100°C to 230°C. 9.
(13) at lO"C/min. Triplicate cultures were measured.. CMCase and Avicelase assays. Basal agar medium (50 ml in 300-ml Erlenmeyer flasks) contained either 2% or 0.2% glucose. Sawdust of Japanese beech (3g) was sprinkled over the surface of the 0.2% glucose-cultures. The cultures were inoculated with a small piece of fungal mat and incubated at 28°C in air. After 10 to 60 days of incubation, 30 ml of distilled water was added aseptically to ten flasks of each culture. The mixtures were stirred with a glass rod, and centrifuged at 10,000 X g at 4°C for 20 min. The supernatants collected from each of the ten cultures were combined and lyophilized. The residue was dissolved in 100 ml of distilled water. Acetone (-10°C) was added to the solution to 70% (V/V). The mixtures were allowed to stand overnight at 4°C, and were centrifuged. The pellet obtained from each was dissolved in 10 ml distilled water and centrifuged to remove insoluble materials and used for CMCase and Avicelase assays. CMCase and Avicelase were assayed by measurement of reducing sugars by the method of Somogyi-Nelson (Somogyi. 1951). Each of the reaction mixtures contained 5 mg of carboxymethy1cellulose sodium salt (CMC, Nacalai Tesque, Inc.) or Avicel (Funacel SF, Funakoshi Co., Ltd.) in 5 ml of Na-acetate buffer (O.IM, pH 5.0). Two ml of each of the solutions was added to each of the reaction mixtures. The reaction mixtures were incubated at 37°C for 0,0.5, 1, 2 and 3 h without agitation for CMC assay and 0, 6, 24 and 48 h with agitation for Avicelase assay. The reaction mixtures for Avicelase were centrifuged to remove insoluble Avicel, and the supernatants were assayed.. Hydroxyl radical production. Basal agar medium (50 ml in 300-ml Erlenmeyer flasks) contained either 2% or 0.2% glucose. Sawdust (3 g), was sprinkled evenly over the surface of the 0.2%-glucose cultures. The cultures were inoculated with a small piece of fungal mat and incubated at 28°C in air. At the indicated intervals, 20 ml of 10% (v/v) dimethyl sulfoxide (DMSO). were added to the cultures. After an additional 24-hour incubation at 28°C, 50 ml of distilled water was added and the cultures were shaken vigorously and centrifuged at 10.
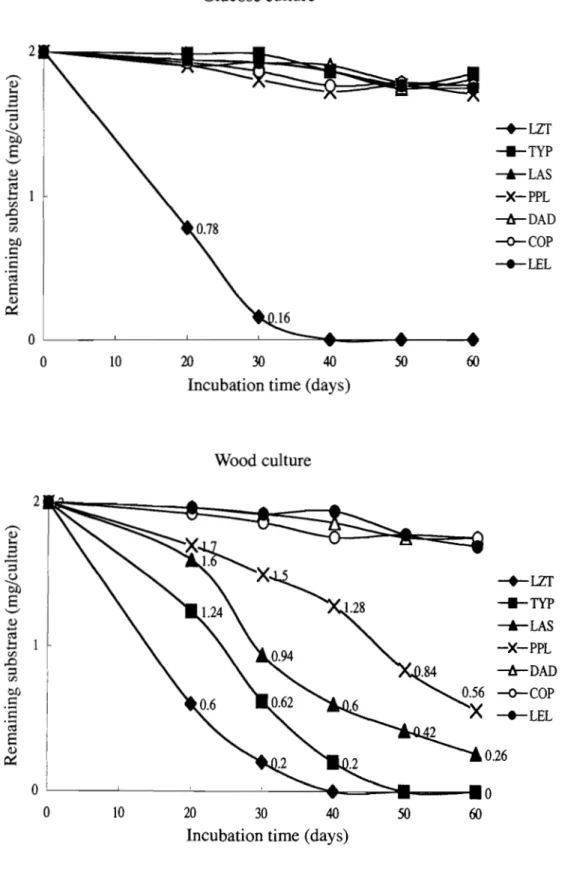
(14) 10,000 X g. The supernatants were filtered through filter paper. Fast Yellow salt solution was freshly prepared by dissolving 1.5 g of the salt in 100 ml of distilled water. The salt solution was filtered and 10 ml were added to each of the culture filtrates. The solutions were shaken for 30 min, transferred to separatory funnels, and extracted with ethyl acetate (100 ml). After the ethyl acetate layers were washed three times with distilled water (50 ml), 50 ml was removed and evaporated under reduced pressure. Each residue was dissolved in ethyl acetate (1 ml), filtered through a Millipore filter (pore size 0.2 !1 m), and analyzed by high performance liquid chromatography as previously. described (Fukui et al. 1993). Triplicate cultures were measured.. Results. Degradation of Japanese beech wood. As shown in Figure 1, LZT and TYP both degraded Japanese beech wood at high rates. Over the 60-day incubation period, LZT and TYP caused similar weight losses in the wood (17% and 16%, respectively). However LZT cultures exhibited a 13% weight loss during the initial 30-day incubation period, compared with only a 9 % weight loss in TYP cultures during the same period. During the second 30 days of incubation, LZT cultures exhibited only 56% of the weight loss observed in TYP cultures during the same period.. Although the weight losses of the wood in LAS and PPL cultures over 60 days of incubation (10% and 8%, respectively) were only 47,..,,63% of the weight losses observed in LZT and TYP cultures, they were still significant (Fig. 1). However unlike LZT and TYP cultures, LAS and PPL cultures degraded more beech wood during the final 30 days of incubation than during the initial 30-day period. The other three fungi, DAD, COP, and LEL, did not significantly degrade beech wood in culture.. 11.
(15) 700 659 630. ,.-., (!) 1-0. 600. ~. 500. -=' .a. ~. -.-LZT. 8. '-". "Cj. 0 0 ~ 4-;. 0. _TYP. 400. 367. -Ir-DAD. 300. -o-COP. rJ:l rJ:l. ..s..... ...c:. --+-LEL. 200. ~. ~. ---'-LAS -X-PPL. 156. 100 51 0 15. 30. 45. 60. Incubation time (days) Fig. 1 Weight loss of Japanese beech wood blocks in cultures of seven fungi.. Degradation of cellulose in glucose and wood cultures. As shown in Figure 2, only two of the brown-rot fungi, COP and LZT, significantly degraded filter-paper cellulose in glucose cultures, with weight losses of 59% and 19%, respectively, over the 60-day incubation period. COP degraded the cellulose at the highest rate in both glucose and wood cultures, with a weight loss of 61 % in wood cultures after 30 days of incubation. As after 45 days, filter papers were seriously destroyed and become very fragile, I failed to recover the remaining residues of the filter papers without any significant loss of the residues on the 45th or 60th day of incubation. LZT exhibited a filter-paper weight loss of 17% during the initial 30 days of incubation in wood cultures and an additional 12% weight loss during the subsequent 30 days. Wood cultures of TYP exhibited a filter-paper weight loss of 12% during the initial 30 days of incubation in wood cultures and an additional 9% weight loss during the subsequent 30 days.. Over the 60-day incubation period in wood cultures, LAS and PPL degraded very similar amounts of filter paper (15% and 14%, respectively).Whereas PPL degraded the 12.
(16) filter paper at a steady rate throughout the incubation period, LAS cultures exhibited a 19-mg weight loss during the first 30 days and a 29-mg weight loss during the second 30 days. In total, LAS and PPL each degraded about 49% as much cellulose as LZT and about 67% as much as TYP in wood cultures. There was little or no degradation of filter-paper cellulose in either glucose or wood cultures of LEL (Fig. 2).. Filter-paper cellulose degradation could not be measured with DAD since the thick mycelia covering the paper could not be removed completely without loss of the substrate. However the filter papers in both wood and glucose cultures of DAD changed very little in thickness or stiffness during the 60-day incubation, indicating that they were not significantly degraded.. Glucose culture 250 ----....<l). .a. "5. ~ 00. a. 200. 195. '-'. -+-LZT. ~. ~ 150. _TYP. .... .....<l). -'-LAS. 0... c.;::: 4-<. 0. ~PPL. 100. ~COP. ___ LEL. <n <n. 0. ...... ...c::. 62. 50. 00. ~. 0 15. 30 45 Incubation time (days). 13. 60.
(17) Wood culture 250 ~ ----;.., ,3. I i. 219. '3. :- 1 ~. 0£). a. '-'. -+--LZT. ;..,. ~. 0.. ro 0... _TYP. I. --'-LAS. ;..,. .... ~. <;:::. 4-c. -*-PPL. 100. 80. 0. rJ) rJ). -.... .-~. ~:. 0. ..c:: 0£). :f. 93. 15. 30 45 Incubation time (days). -o-cop -.-LEL. 60. Fig. 2 Degradation of cellulose as filter paper in glucose and wood cultures of six fungi.. Degradation of a lignin model compound in glucose and wood cultures. As shown in Figure 3, only LZT degraded 4-ethoxy-3-methoxyphenyl- /3 -guaiacyl ether in glucose culture. In these cultures the substrate was undetectable by 40 days of incubation, whereas the other six fungi had degraded less than 0.3 mg after 60 days in glucose cultures.. In wood cultures, LZT degraded all of the substrate within 40 days and TYP degraded it all within 50 days. LAS and PPL had degraded 87% and 72% of the substrate, respectively, after 60 days of incubation. The other three fungi showed little or no activity against the lignin model after 60 days in wood culture (Fig. 3).. 14.
(18) Glucose culture 2 ,.-.... Q). I-<. B. := -2. --+-LZT ____e__Typ. OJJ. S. '-" Q) ...... cd .01. ---'-LAS -X-PPL. [/J. ..0. :=. ---tr-DAD. OJJ. -o-COP. [/J. t:: ........ -+-LEL. t:: .@. S Q) ~. 0 0. 10. 20. 30. 40. 50. 60. Incubation time (days). Wood culture 2 ,.-.... Q). I-<. --S := ...... := u. --+-LZT ____e__TYP. OJJ. '-". 2cd. ---'-LAS -X-PPL. .0 [/J ..0. :=. ---tr-DAD. [/J. -o-cop. OJJ. .;::t::. -+-LEL. .@. E Q). 0.26. ~. 0. 0 0. 10. 20. 30. 40. 50. 60. Incubation time (days) Fig. 3 Degradation of the lignin model compound, 4-ethoxy-3-methoxyphenyl- /3 guaiacyl ether, in glucose and wood cultures of seven fungi.. 15.
(19) Oxalic acid concentrations in cultures. Figure 4 shows the oxalic acid concentrations in glucose and wood cultures during the 60-day incubation period. Glucose cultures of TYp, LAS, and PPL produced significant amounts of oxalic acid throughout the incubation period. Oxalic acid concentrations peaked in TYP and LAS cultures on day 20 at 30 mg and 21 mg per culture, respectively. In glucose cultures of COP, some amounts of oxalic acid (1-7 mg) were detected throughout the incubation period. In glucose cultures of LZT, DAD, and LEL, only very small amounts (less than 1 mg) of oxalic acid were detected.. Much smaller amounts of oxalic acid were detected in wood cultures as compared to glucose cultures. TYp, LAS, and PPL cultures had small amounts of oxalic acid (3-9 mg) throughout the incubation period. Less than 1 mg of oxalic acid was detectable in wood cultures of the other fungi (Fig. 4).. Glucose culture. ~r ,..-.., C!). 1-<. -+-LZT _TYP. B '3 20 ~. b.I). -Ir-LAS. 8 '-". -X-PPL. '"d .~. ~DAD. u. --<r-COP. ~ x. 10. -+--LEL. 0. 10. 20. 30. 40. Incubation time (days). 16. 50. 60.
(20) Wood culture 30. ---....=. -+-LZT. ~. I-;. 3. """-TYP .t-LAS. 20. -S2 01) 8 '--'. -X-PPL. '" .~. .~ ~. -1s-DAD 10. i<. 0. X. X- -X-X. 30. 40. -o-COP _LEL. 0. 10. 20. 50. 60. Incubation time (days) Fig. 4 Oxalic acid concentrations in glucose and wood cultures of seven fungi.. Hydroxyl radical production in glucose and wood cultures. The assay for -OH, based on the transformation of DMSO to methanesulphinic acid, is specific for -OH (Hirano et al. 1997). In glucose culture, only LZT produced significant -OH (Fig. 5). Production in LZT glucose cultures was highest on day 10 and had dropped by more than 90% by day 30.. In wood cultures, only LZT and TYP exhibited high levels of -OH (Fig. 5). The production of -OH in LZT wood cultures was somewhat higher than in LZT glucose cultures, peaking on day 20 of incubation and then decreasing sharply to less than 4% of the maximum by day 60. The production of -OH in wood cultures of TYP peaked on day 20 of incubation and decreased gradually to about 16% of the maximum by day 60. Over the entire 60-day incubation period, the ratio of the integrals of the -OH production in LZT and TYP cultures was 1.02. However over the last 30 days of incubation, the ratio of the integrals of-OH production in LZT and TYP cultures was 17.
(21) 0.76. Wood cultures of LAS and PPL exhibited much lower -OH production than LZT and TYP cultures (Fig. 5). Over the 60-day incubation period, the integrals of the-OH production by LAS or PPL cultures was 33-35% those of the LZT or TYP cultures. Over the 60-day incubation period, the ratio of the integrals of the -OH production in wood cultures of LAS and PPL was approximately 1.2. In LAS and PPL cultures, -OH production increased gradually, peaking on day 45 and then decreasing. During the initial 30-day incubation period, the integrals of the -OH production in LAS and PPL wood cultures were about 30% of the integrals for the second 30-day incubation period. There was little or no -OH production in wood cultures of DAD, COP, and LEL (Fig. 5).. Glucose culture. ..c: ----. '"<t N. -'"3 --E ~. ;:l ....... 500. u. "0 400 -+-LZT. r--. b. _TYP. X 300 '-'. -'-LAS -X-PPL. c::. .9 ...... u. ;:l. ""0 0 I-< 0... -..... -x ro. u. -lr-DAD. 200. -o--COP -""-LEL. 100. ""0. ro. I-<. :>... 0 ""0. I-<. :>... ::t:. 0 10. 20. 45. 30. Incubation time (days). 18. -. QO.
(22) Wood culture. ,.-... ..c: '<:t N. --..... '3 --S "0 ~. 600. ::l. (,). r-. 500 -+-LZT. 400. _TYP. b...... X '-" c 300. ~LAS. -X-PPL. .s ...... -tr-DAD. (,). ::l "0 0. -o--COP. 200. ____ LEL. ~. 0-. ~. (,) .-<. 100. -g. ~. >-.. >< 0. 0. ~. 10. 20. 45. 30. 60. "0. >-.. l::. Incubation time (days). Fig. 5 Hydroxyl radical production in glucose and wood cultures of seven fungi.. Discussion. White-rot and soft-rot fungi preferentially degrade and eliminate the lignin in wood cell walls via the activities of ·OH and phenol oxidases, including laccase, manganese peroxidase, and lignin peroxidase (Tanaka et al. 1999; Tanaka et al. 2000). In addition, white-rot and soft-rot fungi generally produce the full cellulolytic enzyme complement (endo-l,4- J3 -glucanase, exo-l,4- J3 -glucanase, and 1,4- J3 -glucosidase). Therefore they can hydrolyze highly-crystalline cellulose substrates (Highley. 1988; Tanaka et al. 1988). But a complete cellulolytic system alone is insufficient for degrading the cellulose in wood cell walls because the lignin covering the cellulose prevents the enzymes from reaching the substrate (Tanaka et al. 1999). Some deuteromycetes, such as Trichoderma sp. and Fusarium sp., produce a complete cellulolytic system and efficiently hydrolyze highly-crystalline cellulose to glucose, but can degrade neither wood nor the cellulose component of wood because they have no system for removing the lignin (Tanaka et al. 1999; Highley. 1973).. 19.
(23) The brown-rot fungi, LZT, TYP, LAS, and PPL, preferentially degrade and eliminate the polysaccharides in wood, leaving a lignin-rich residue (Enoki et al. 1988; Highley. 1987). Wood-containing cultures of TYP have high CMCase activity but negligible Avicelase activity. Both activities are negligible in glucose cultures ofTYP (Hirano et al. 1997). Likewise, Avicelase activity is negligible throughout the incubation period in both glucose and wood cultures of LZT, LAS, and PPL (data not shown).. Fungal enzymes such as cellulases are too large to diffuse into wood cell walls during the early stages of wood degradation by brown-rot fungi (Flournoy et al. 1991; Kuo et al. 1988). Although the S2 layer of the wood cell wall is degraded extensively during the early stages of brown-rot degradation, the S3 layer adjacent to the cell lumen is less affected when it is attacked by fungal hyphae from the lumen (Kuo et al. 1988; Highley and Murmanis. 1985). However ·OH can depolymerize crystalline cellulose and destroy its crystalline structure (Kirk et al. 1991). Furthermore, ·OH can attack the aromatic rings in lignin, causing a variety of reactions including hydroxylation and ring-opening (Gierer et al. 1992). However ·OH alone does not delignify wood significantly (Chirat and Lachenal. 1997). Endoglucanases isolated from cultures of a brown-rot fungus and a cellulolytic fungus little hydrolyze crystalline cellulose or the cellulose in wood unless the crystalline cellulose or the wood has been pretreated with ·OH (Ratto et al. 1997).. These findings suggest that ·OH produced in cultures of brown-rot fungi is primarily responsible for destroying the crystalline structure of the cellulose and attacking the lignin in wood, thereby opening canals through the S3 layers for endo-l ,4-/3 -glucanase diffusion. In the present study, I have assessed the possible involvement of ·OH and oxalic acid in the degradation of wood, filter-paper cellulose, and a lignin-substructure model by seven fungi usually regarded as brown-rot fungi.. Over the 60-day incubation period of wood-containing cultures of LZT and TYP, declining levels of ·OH production coincide with declining rates of wood and filterpaper cellulose degradation, as well as the degradation rate of the lignin-substructure model compound, 4-ethoxy-3-methoxyphenyl-/3 -guaiacyl ether (Figs. 1,2,3, and 5). Furthermore, in wood-containing cultures of LAS and PPL, increasing rates of ·OH 20.
(24) production coincide with increasing rates of wood, cellulose, and lignin model compound degradation. Wood-containing cultures of DAD, COP, and LEL do not produce significant amounts of ·OH; nor do they cause significant weight loss in beech wood or the disappearance of the lignin model compound. Cultures of LEL do not result in significant filter-paper weight loss and, although not measurable, DAD also does not appear to significantly degrade filter paper.. Although unable to degrade either wood or the lignin model compound, COP extensively degrades filter-paper cellulose in both glucose and wood-containing cultures (Fig. 2). Among the fungi tested, only COP and LZT significantly degrade filter paper in glucose cultures. In both wood and glucose cultures of COP, the activities of Avicelase and CMCase are significant, whereas phenol oxidase activity is negligible (data not shown). In addition, two extracellular exo-cellobiohydrolases have been purified to homogeneity from culture filtrates of COP (Schmidhalter and Canevascini. 1993). Thus the COP strain used in the present work has a pattern of wood degradation that is cellulolytic but nonlignolytic, as in fungi such as Trichoderma sp. and Fusarium sp., rather than a brown-rot pattern of wood degradation.. The generation of ·OH in wood-degrading cultures of LZT, TYP, LAS, and PPL is inversely proportional to the amount of oxalic acid present in the cultures (Figs. 4 and 5). Thus the amounts of oxalic acid in cultures of the four fungi are in inverse proportion to the rates of degradation of wood, filter-paper cellulose and the ligninrelated compound.. These results indicate that ·OH could be readily decomposing the oxalic acid in cultures. For example, when cultures of LZT are actively depolymerizing holocellulose, exogenously-added oxalic acid is rapidly converted to CO2 (Espejo et al. 1990). LZT significantly degrades wood (Fig. 1). It also significantly degrades filter-paper cellulose and a lignin-substructure model in both glucose and wood cultures (Figs. 2 and 3). LZT generates large amounts of ·OH in both glucose and wood cultures (Fig. 5) and accumulates only traces of oxalic acid (Fig. 4). TYp, LAS, and PPL degrade wood, and filter-paper cellulose and the lignin model compound in wood-containing cultures, to a significant extent. They accumulate only very small amounts of oxalic acid in wood 21.
(25) cultures. However in glucose cultures of TYp, LAS, and PPL, degradation of filter paper or the lignin model compound is negligible, as is production of -OH; whereas large amounts of oxalic acid accumulate. Of all the fungi tested, COP has the highest degradation rate for filter-paper cellulose, in both glucose and wood cultures, but does not degrade wood or the lignin model compound (Figs. 1,2, and 3). Although oxalic acid is present in glucose cultures of COP throughout the incubation period, only very small amounts are detected in wood cultures (Fig. 4). DAD and LEL do not degrade any of the substrates, produce little or no -OH, and accumulate only very small amounts of oxalic acid in glucose and wood cultures (Figs. 1-5). These results indicate that oxalic acid is not directly involved in the degradation of wood, filter-paper cellulose, or the lignin model compound by brown-rot fungi. However oxalic acid may be involved in -OH production by brown-rot fungi that have substantial wood-degrading activities. In cultures of brown-rot fungi, -OR is generated by the Fenton reaction: (Enoki et al. 2003). Oxidation of oxalic acid to CO2 can reduce Fe( III) to Fe( II ) (Schmidt 1981). Some oxidants oxidize oxalic acid with the simultaneous reduction of O2 to superoxide anion, O 2- (Barr et al. 1992). Superoxide anion reduces Fe(III) to Fe( II) and forms H20 2 in a disproportionation reaction. In this manner, oxalic acid may be involved in producing both Fe(II) and H 20 2 for hydroxyl radical generation.. 22.
(26) CHAPTER. ill. Physical and Chemical Properties of an Extracellular Low-MolecularWeight Substance from the Brown-Rot Basidiomycete Tyromyces palustris Background Although brown-rot fungi significantly degrade the lignin in wood, they preferentially degrade and eliminate the polysaccharide components, leaving an amorphous, brown, crumbly residue composed largely of lignin (Highley. 1987~. Filley et al. 2002). Brown-. rot fungi degrade cellulose in wood even though they lack exo-1 ,4-glucanase activity for hydrolyzing crystalline or natural cellulose. However they will degrade and metabolize cellulose only when their ligninolytic systems are active (Enoki et al.. 1988~. Hirano et al. 1997). This indicates that brown-rot fungi may possess a unique wooddegrading system that is capable of fully degrading natural cellulose and modifying lignin.. In the early stages of wood degradation by brown-rot fungi, the S2 layer of the wood cell wall is degraded extensively. However the S3 layer adjacent to the cell lumen is less affected when it is attacked by fungal hyphae from the lumen (Enoki et al. 1988; Kuo et al. 1988). Fungal enzymes, such as cellulase and peroxidase, are too large to penetrate wood cell walls at early stages of degradation (Flournoy et al. 1991; Srebotnik and Messner 1991). The one-electron-oxidizing activity in cultures of brown-rot fungi is proportional to the degradation of lignin-related model compounds, natural cellulose substrates, and wood in the cultures. This one-electron-oxidizing activity appears to result from hydroxyl radicals (Enoki et al. 1989). Hydroxyl radical (-OH) is a strong oxidant that severely degrades the cellulose in wood, but does not delignify wood significantly (Chirat and Lachenal 1997). However -OH attacks lignin through a variety of reactions (Filley et al. 2002; Gierer et al. 1992). Some brown-rot fungi degrade lignin-related model compounds to yield relatively large amounts of the same products that are obtained when the lignin model compounds are attacked by -OH (Espejo et al. 1990; Zohar et al. 1998). Furthermore, endoglucanases isolated from cultures of a 23.
(27) brown-rot fungus hydrolyze very little of the cellulose in wood, unless the wood has been pretreated with ·OH (Ratto et al. 1997). These results suggest that brown-rot fungi have a unique non-enzymatic degradative system that may involve the diffusion of ·OH into secondary cell walls, with the degradation occurring at some distance from the fungal hyphae in the lumen.. An extracellular metabolite, 2,5-dimethoxy-l ,4-benzoquinone, has been isolated from liquid cultures of the brown-rot fungus, Gloeophyllum trabeum. This benzoquinone can reduce Fe(III) to Fe (II) and O 2 to H 20 2 , thereby promoting a Fenton reaction between Fe(II) and H 20 2 (Zohar et al. 1998). A low-molecular-weight peptide «5,000) that is a high-affinity Fe(III) chelator and that can reduce Fe(III) to Fe(II) also has been isolated from culture extracts of G. trabeum (Wang and Gao 2003).. Extracellular low-molecular-weight substances that catalyze a redox reaction between O 2 and an electron donor to produce ·OH have been isolated from wood-degrading cultures of the brown-rot fungi G. trabeum and Tyromyces palustris. (Hirano et al. 1997; Enoki et al. 1992; Hirano et al. 1995). The low-molecular-weight substance from T. palustris cultures reduces Fe(III) to Fe(II) and strongly chelates Fe(II) (Hirano et al. 1995). Most of the extracellular ·OH produced in intact cultures of T. palustris is generated by a redox reaction between O 2 and an electron donor that is catalyzed by this low-molecular-weight substance (Hirano et al. 1995). During the early stages of wood degradation by T. palustris, the low-molecular-weight substance is localized in the fungal cytoplasm, the fungal cell Wall, the extracellular sheath surrounding the fungal cell Wall, and throughout the wood cell wall, suggesting that it diffuses from the S3 layer into the S2 layer and the middle lamella (Hirano et al. 2000). Herein I report on the physical and chemical properties of this extracellular low-molecular-weight substance from cultures of T. palustris.. 24.
(28) Materials and methods. Organism. The brown-rot fungus, T. palustris (Berk. Et Curt) Murr. FRI 0507, was used throughout this study.. Culture conditions. Basal agar medium was prepared as previously described (Enoki et al. 1988) except that the medium contained 0.2% glucose and ten-fold concentrated trace elements. Wood meal from Japanese beech (Fagus crenata Blume) was extracted twice with acetone and dried. The \}'ood meal (2 g), wetted with 1.2 ml water, was sterilized and sprinkled evenly over the surface of the basal medium (30 ml) in 300-ml Erlenmeyer flasks. The cultures were inoculated with a small piece of fungal mat and incubated at 28°C. Isolation and partial purification of the extracellular substance. After 30 days of incubation, 30 ml of distilled water was added to the cultures. The mixtures were stirred with a glass rod and centrifuged at 10,000 x g at 4°C for 20 min. The supernatants were filtered. The extracellular medium from 100 flasks was acetoneprecipitated (70% v/v), as described previously (Hirano et al. 1997; Hirano et al. 1995). The precipitate was dissolved in 100 ml of distilled water and centrifuged at 10,000 x g for 20 min. Aliquots (10 ml) of the resulting supernatant were loaded on a distilledwater-equilibrated Sephadex G-50 column (3.8 X 60 cm) and the column was eluted with distilled water. Fractions were collected, the absorbance at 280 nm was measured, and the ethylene generation from 2-keto-4-thiomethylbutyric acid (KTBA) of the fractions was assayed as described below. Fractions comprising the major peak of ethylene-generating activity were pooled and lyophilized. The lyophilized material was combined and dissolved in 50 ml of distilled water. Aliquots (10 ml) were loaded on a distilled-water-equilibrated Sephadex G-25 column and eluted with distilled water. Fractions were collected and assayed for ethylene generation from KTBA. Fractions comprising the major peak of ethylene-generating activity were pooled and lyophilized. 25.
(29) The lyophilized material was combined and dissolved in distilled water (20 ml). Aliquots (l ml) of the solution was applied to a column of Hiprep 16/10 DEAE FF (Amersham Biosciences, Sweden), previously equilibrated with 0.02 M Tris-HCl buffer (pH 8.0) and eluted with a linear gradient of 0 to 2 M NaCl by Fast-Protein Liquid Chromatography (Amersham Biosciences, Sweden). Fractions containing the major peak of one-electron oxidizing activity, as measured by ethylene generation from KTBA, were pooled and desalted on a column of HiTrapTMDesalting (Amersham Biosciences, Sweden). The desalted solution was lyophilized. A final yield of 50 mg of the extracellular one-electron-oxidizing substance was obtained from 100 culture flasks.. Ethylene generation from KTBA. Ethylene generation from KTBA was measured as described previously (Enoki et al. 1989). Reaction mixtures contained 300 Jll of eluate, 3001'1 of 0.01 M H2 0 2 , 300 1'1 of 0.16 M N a-acetate buffer (pH 4.5), and 501'1 of 0.1 M KTBA. After 1h at 30°C, 1 ml of gas was removed from the headspace and the ethylene concentration was measured directly by gas chromatography as described previously (Enoki et al. 1989).. Electrophoresis. Sample preparation and Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoreses (Tricine-SDS-PAGE) were as described by Schagger and Jagow. 1987. The slab gels (0.75 mm thick) consisted of three sections: a 15-cm separation gel (16.5% T, 3% C), a 3-cm spacer gel (10% T, 3% C), and a 2-cm stacking gel (4% T, 3% C). T denotes the total percentage concentration of both (acrylamide and bisacrylamide). C denotes the percentage concentration of the cross linker relative to the total concentration T. The mounted gels were allowed to polymerize overnight at room temperature. Electrophoresis of the samples was performed at room temperature. The initial running conditions were 30 V for 1-2 h. After the samples had entered the stacking gel, electrophoreses was at 80 V. Proteins were visualized by staining with silver nitrate.. 26.
(30) ElectrotransJer Following electrophoresis the gel was equilibrated in transfer buffer (10mM 3[cyclohexylamino-1-propanesulfonic acid] (CAPS), pH 11, including 20% methanol) for 10 min. Proteins were blotted onto the PVDF membrane (Immobilon-pSQ transfer membrane, pore size 0.1/.1 m, Millipore) equilibrated in transfer buffer before use, using a semidry electroblotting assembly. Electrotransfer was performed for 3h at 0.2mA!cm2 • The PVDF membrane was stained 0.1 % (w/v) Coomassie Brilliant Blue R-250 in 50% (v/v) methanol for 1 min and then destained 40% (v/v) methanol including 10% (v/v) acetic acid. Protein containing regions detected by Coomassie Brilliant Blue stain were cut out and were rinsed in 50% (v/v) methanol before protein sequence analysis (Ploug et al. 1989).. Hydroxyl radical production. Hydroxyl radicals were measured by the method of Fukui et al. 1993. The basic reaction mixture contained 1 mg of the partially-purified preparation, 3 X 10-4 mol of dimethyl sulfoxide (DMSO), and Na-acetate buffer (40 mM, pH 4.5) in a total volume of 2 ml in a 33-ml test tube. Hydrogen peroxide (3 X 10-6 mol), NADH (3 X 1O-6 mol), NADH plus superoxide dismutase (SOD) (0.3 mg, 2,670units mg- I ), NADH plus catalase (0.3 mg, 42,000units mg- I ), NADH plus N,N' -dimethyl-4-nitrosoaniline (DMNA) (6 X 1O-6mol), or ascorbic acid (3 X 1O-6mol) were added to the basic reaction mixture. After 1.5 h of incubation at 30°C under 100% O 2 or N2 , 1 ml of 2% (w/v) Fast Yellow GC Salt reagent (Sigma Chemical Co., St. USA) was added to the reaction mixture. The mixture was shaken and allowed to stand for 30 min. Ethyl acetate (10 ml) was added and the mixture was shaken well. After the ethyl-acetate layer was washed with distilled water (10 ml, 3 times), 8 ml of the ethyl-acetate layer was removed and evaporated off under reduced pressure. The residue was dissolved in ethyl acetate (500 }ll), filtered through a Millipore filter (pore size 0.2 }lm), and analyzed by high-performance liquid chromatography as previously described (Lowry et al. 1951).. Protein, amino acid, carbohydrate, and Fe analysis. 27.
(31) The protein concentration of the partially-purified preparation was determined by a modification of the method of Lowry et al. 1951. The amino acid composition was determined with an amino acid autoanalyzer (HITACHI, L-8500, Japan), following hydrolysis of the sample with 6 N HCl. The neutral carbohydrate content was determined by the phenol-sulfuric acid assay (Dubois et al. 1956). Monosaccharide composition was measured with a saccharide composition analysis kit (Honen Corporation, Japan) according to the manufacturer's instructions. Following hydrolysis of the sample, the released monosaccharides were coupled with p-arninobenzoic ethyl ester at the reducing end by reductive amination, according to the method ofYasuno et al. 1997. The p-aminobenzoic ethyl ester-derivatized monosaccharides were injected into a Honenpak C18 column (75 X4.6 mm i.d., Honen Corp., Tokyo, Japan) and detected by fluorescence monitoring according to the method ofYasuno et al. 1997. Ferrous iron content was measured with ferrozine by the procedure of S0rensen. 1982. Primary sequence analysis was performed by automated Edoman degradation with a model 477A sequencer from Applied Biosystems using the manufacture's standard programming and chemicals.. Fe(III)- and Cu(II)-reducing activity. The reduction of Fe(III) to Fe(II) by the preparation was measured using ferrozine, according to the procedure of S0rensen. 1982. Distilled water (500 'pI), with or without 10 ppm Fe(III), was added to an aqueous solution of the partially-purified preparation (35 'pg in 500 'pI). For the control, an aqueous solutions (500 .pI) containing 10 ppm Fe(II) or Fe(III) was added to distilled water (500 'pI). After 24 h of incubation at 30°C, 500 'pI of 1% ferrozine in distilled water were added. After standing for 24 h at 30°C, the absorption spectra of the solutions were measured from 400 nm to 750 nm with a spectrophotometer. The Cu(II) reducing ability of the preparation was measured by the method of Somogyi-Nelson (Somogyi. 1951).. Determination of carbonyl groups. The carbonyl groups, other than those in the peptide bonds, were assayed by the method described in Uchida et al. 1998. Distilled water (500 'pI), with or without 3.5 mg of 28.
(32) sample, was added to 0.1 % 2,4-dinitrophenylhydrazine in 2 N HCl. After 1 h at room temperature, 500 ]ll of 20% trichloroacetic acid (TCA) was added to each assay. Following centrifugation at 9,500 x g for lOmin, the supernatants were scanned from 800 nm to 300 nm. The number of carbonyl groups was calculated based on the rate of decrease in absorbance at 365 nm.. Determination of aldehyde groups. Aldehyde groups in the preparation were assayed using methone (dimethyldihydroresorcin). Distilled water (1 ml), with or without 10 mg of sample, was mixed with 100 ]ll of 0.1 M methone in methanol and shaken vigorously for 30 min. Controls contained 100]l1 of 0.1 M methone and 1 ml aqueous solution of 0.012 M formaldehyde or 0.012 M 2,4-dihydroxybenzaldehyde. The mixtures were extracted with 1 ml ethyl acetate. Methone in the ethyl acetate fraction was measured directly by gas chromatography at 160°C in the isothermal mode using a glass column packed with 3% OV-101 on chromsorb Q 1001120 (Applied Science, Japan). The number of aldehyde groups was calculated based on the loss of methone.. Determination of protein glycosylation. Glycosylation of N-terminal and side-chain amino acids in the glycopeptide was measured by the thiobarbituric acid (TBA) colorimetric technique of Fluckiger and Gallop. 1984. Following the addition of 500 ]ll of 0.5 M oxalic acid to 1-ml aqueous solutions containing 0, 1 mg, or 2 mg of sample, or 25 ]lmol of cellobiose (8.6mg), glucose (4.5mg), galactose (4.5mg), or mannotriose (12.6mg), the tubes were mixed and capped with rubber stoppers vented with 26-gauge needles. The tubes were placed in a heating block at 100°C and the needles were removed after a lO-min equilibration period. After 5 h, the tubes were cooled in an ice bath. The volumes of the solutions were adjusted with water to 1.5 ml. Cold 40% TCA (500 ]ll) was added to each tube and the tubes were mixed and centrifuged at 9,500 x g for 4 min. A 500 ]l1 aliquot of the supernatant was removed and 500 ]ll of a 0.05 M aqueous 2-thiobarbituric acid solution were added. The solutions were incubated at 37°C for 15 min and at room temperature for 20 min. The absorption spectra were measured from 400 nm to 700 nm. The 29.
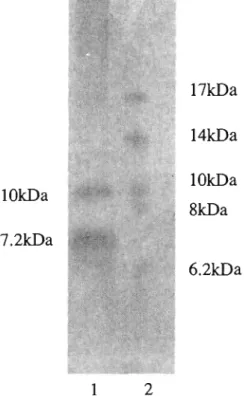
(33) standard curves were prepared using }lmol range of 5-hydroxymethylfurfural (5-HMF) and 500 }ll of a 0.05 M aqueous TBA.. Results and discussion Partial purification of the extracellular substance. The Sephadex 0-50 gel-filtration chromatography of the acetone precipitate of the extracellular fluid from 30-day-old T. palustris cultures was carried out. Only one major peak of one-electron-oxidation activity, as measured by ethylene generation from KTBA, was observed in the low-molecular-weight fractions. (1000~5000).. This peak. corresponded to the maximum absorbance peak at 280 nm. The major peak was pooled and lyophilized. Then the Sephadex 0-25 gel-filtration chromatography of this pooled material was carried out. Low molecular-weight fractions. (1000~5000),. constituting the. major peak of one-electron-oxidation activity, were pooled, lyophilized, and subjected to anion exchange chromatography on HiPrep 16/10 DEAE FF. Fractions, constituting the major peak of one-electron-oxidation activity were pooled, desalted, and lyophilized. The yield of this partially-purified preparation was 50 mg from 100 flasks of 30-ml, 30day-old cultures of T. palustris.. Composition and molecular weight. The molecular weight of the partially-purified preparation, as determined by gelfiltration on Sephadex 0-25 or Sephadex 0-50, was in a range of 1,000 ~ 5,000 (data not shown). Tricine-SDS-PAOE of the partially-purified preparation yielded two bands of7.2 kDa and 10.0 kDa (Fig. 6). The partially purified preparation was 42% neutral carbohydrate by weight and 54% protein by weight. The true molecular weight of the preparation could be lower than 7.2 kDa and 10.0 kDa because molecular weights of glycopeptides determined by SDS-PAOE are higher than true molecular weights (Leach et al. 1980). The N-terminal amino acid sequence of the separated substances (7.2 kDa, 10.0 kDa) by Tricine-SDS-PAOE is shown in Fig. 7. These two sequences were same sequence from N-terminal amino acid to seventh amino acid (Fig. 7). The BLAST search was performed about these sequences but protein have a homology with these 30.
(34) substances was not in the BLAST database.. 17kDa 14kDa l OkDa. lOkDa. 8kDa. 7.2kDa 6.2kDa. 1. 2. Fig.6 Tricine SDS-PAGE of the partially purified extracellular substance from T. paiustris. Lane 1: Partially purified substance. Lane 2: Protein standard. Gel was stained with silver nitrate.. 5. 10.0 kDa. 10. NHAAQQ. 7.2kDa Fig. 7 The N-terminal amino acid seque,nce of one-electron oxidation substance from T. paiustris. The sequence of N-terminal amino acid of one-electron oxidation substances were determined with a model 477 A automated gas phase sequencer. Shaded area shows homologies.. 31.
(35) Hydroxyl radical generation. The transformation of DMSO to methane sulfuric acid is a specific assay for ·OH (Hirano et al. 1997). The generation of ·OH by the partially-purified glycopeptide in the -6. presence of the electron donor NADH (3 X 10 mol) under a 100% O 2 atmosphere (9.2 8. 1. X 10- mol mfl 1.5h- ) was given a relative value of 100% (Table 1). In the absence of H20 2 or an electron donor such as NADH or ascorbic acid, only a very small amount of ·OH was generated under 100% O 2 and almost no ·OH was generated under an N2 atmosphere. In the presence of NADH or ascorbic acid under 100% O 2, ·OH generation was 15-fold and 47-fold higher, respectively, than in the absence of an electron donor. In the presence of NADH, under 100% O 2, catalase almost completely blocked ·OH generation and the ·OH-scavenging agent DMNA suppressed ·OH generation. Likewise, SOD inhibited ·OH generation in the presence of NADH by about 40% (Table 1). These results indicate that the extracellular glycopeptide from T. palustris catalyzes redox reactions between electron donors such as NADH or ascorbic acid and O 2 to produce H20 2 via superoxide and further reduces H20 2 to ·OH. We previously demonstrated that the production of ·OH in T. palustris cultures is related to the degradation rates of wood, crystalline cellulose, and lignin-model compounds in the cultures (Hirano et al. 1997).. 32.
(36) Table 1 Generation of hydroxyl radical by the partially-purified glycopeptide isolated from wood-degrading cultures of T. palustris. a. Atomosphereb. Addition to reaction mixture. X 10-8 mol mrl 1.5h-. Ie. Relative value (%). None. 100% O2. 0.6. 6. None. 100% N2. 0.3. 3. 3 X 10-6 mol H2 0 2. 100% N2. 4.0. 44. 3 X 10-6 mol NADH. 100% O2. 9.2. 100. 3 X 10-6 mol NADH, 0.3mg SODd. 100% O2. 3.6. 39. 3 X 10-6 mol NADH, 0.3mg catalase. 100% O 2. 0.9. 10. 3 X 10-6 mol Ascorbic acid. 100% O2. 28.2. 307. 100% O2. 3.3. 36. 6. 3 X 10-6 mol NADH, 6X 10- mol DMNA. aReaction mixtures contained 1.0mg of the preparation, 3 X 1O-4mol DMSO, and acetate buffer (40mM, pH4.5) in a total volume of 2 ml. Reactions and detection of methane sulfuric acid were as described in the text. bReaction mixtures were incubated under 100% O 2 or N 2 , as described in the text. cEach value represents the average of triplicate assays. dSuperoxide dismutase, 2,670unitimg.. 33.
(37) Amino acid, monosaccharide, and Fe content. Amino acid analysis of the partially-purified preparation indicates that it contains 10.4% aspartic acid, 9.1 % glutamic acid, 1.3% lysine, and 25.1 % NR3 (Table 2). Thus the substance contains an abundance of amino acids with side-chain amino groups. The glycopeptide contains unusually high levels of galactose (47%), glucose (22%), and xylose (18%); whereas there is less than 1% N-acetyl-D-glucosamine and the Nacetyl-D-galactosamine is negligible (Table 3). In glycopeptides, N-acetyl-Dglucosamine usually combines with the side-chain amino group of asparagine and Nacetyl-D-galactosamine combines with the OR side chains of serine or threonine. These results suggest that carbohydrates other than N-acetyl-D-glucosamine and N-acetyl-Dgalactosamine are attached to side-chain amino groups in this glycopeptide. The partially-purified glycopeptide contained 0.06% ferrous iron by weight, as determined with ferrozine.. 34.
(38) Table 2 Amino acid composition of the partially-purified glycopeptide isolated from wood-degrading cultures of T. palustris. a. Amino acids. %. Amino acids. %. Asparagine. NDb. Tryptophan. ND. Aspartic acid. 10.4. Threonine. 8.9. Alanine. 7.8. Valine. 3.6. Arginine. 1.1. Histidine. 0.4. Isoleucine. 3.2. Phenylalanine. 3. Glycine. 7.1. Proline. ND. Glutamine. ND. Methionine. ND. Glutamic acid. 9.1. Lysine. 1.3. Cysteine. ND. Leucine. 5.4. Serine. 9.6. NH3. 25.1. Tyrosine. 3.7. aDetermined with an amino acid autoanalyzer, following hydrolysis of the sample with 6 N HCl. bND: Not detectable.. 35.
(39) Table 3 Monosaccharide composition of the partially-purified glycopeptide isolated from wood-degrading cultures of T. paiustris. a. Monosaccharide. %b. Galactose. 47. Mannose. 8. Glucose. 22. Arabinose. 4. Xylose. 18. N-acetyl-D-glucosamine. 1. N-acetyl-D-galactosamine. 0. aMonosaccharide composition was determined using a saccharide composition analysis kit, as described in the text. bpercentage of total monosaccharides.. 36.
(40) Fe(IJl)- and Cu(IJ)-reducing activity. The ferric-iron-reducing activity of the partially-purified glycopeptide was measured using ferrozine (Fig. 8, Table 4). Fig. 8 shows that 1 mg of the glycopeptide reduced about 1.7}tmol of Fe(Ill) to Fe(ll); that is, a single mole of the glycopeptide reduced about 17 moles of Fe(III) to Fe(II), assuming that the molecular weight is 10,000. The Cu(II)-reducing activity of the glycopeptide, measured by the method of Somogyi-Nelson, was 1.3 }tmol/mg (Table 4). 2.5. 2.0. 1.5. 1.0. 0.5. 400. 500. 700. 600. 800. Wavelength (nm) Fig. 8 Fe(III) reduction by the partially-purified extracellular glycopeptide from. T. palustris cultures. Absorption spectra were determined from 400 t0750 nm. Fe(III) reduction was measured using 1% ferrozine, according to the procedure of S~rensen (1982), as described in the text. A: 10/3 ppm Fe(II); B: 10/3 ppm Fe(Ill), 35 }tg glycopeptide sample; C: 10/3 ppm Fe(Ill); D: 35 }tg glycopeptide sample. (. ): absorbance at 560nm. 37.
(41) Table 4 Fe(III)- and Cu(II)-reducing activities and carbonyl and aldehyde content of the partially-purified glycopeptide isolated from wood-degrading cultures of T. palustris.. jtmol mg-!. moI10 4 g-!. Fe (III) reductiona Cu(II)-reductionb. 1.7. 17. 1.3. 13. Carbonyl groupsc. 0.6. 6. Aldehyde groupsd. negligible. negligible. 0.5. 5. 1-amino-2-ketosese. aDetermined using 1% ferrozine, according to the procedure of SiZlrensen (1982), as described in the text. bDetermined by the method of Somogyi-Nelson. CDetermined using 2,4-dinitrophenylhydrazine method of Uchida et al. (1998) as described in the text. dAssayed using methone, as described in the text. e1-amino-2-ketoses produced by the condensation of the N-terminal or side-chain amino groups and carbohydrate aldehydes.. 38.
(42) Carbonyls The carbonyl groups in the glycopeptide, excepting those in the peptide bonds, were determined with 2,4-dinitrophenylhydrazine, which combines with carbonyl groups, with the loss of water, to form 2,4-dinitrophenylhydrazone, which is insoluble in water. The number of carbonyl groups is calculated based on the decrease in absorbance at 365 nm. Carbonyl groups were present 0.6/.1 mol mg- 1 in the glycopeptide (Table 4). The aldehyde groups in the glycopeptide were determined using methone, which readily condenses stoichiometrically with aldehyde groups but not with ketones. 1. Aldehyde groups were present in the glycopeptide at less than 3 X 1O-9}lmol mg- or 0.04 1. mol mg- (Table 4). This suggests that most of the carbonyl groups in the glycopeptide are ketone or endiol groups rather than aldehydes. However 0.6 }lmol of ahydroxyketone per mg of glycopeptide could reduce, at most, only 1.2 }lmol of Fe(Ill), less than the measured Fe(IlI) reduction of 1.7 }lmol per mg of glycoprotein (Table 4).. Glycosylation. Glycosylation of the N-terminal and the side-chain amino groups in the glycopeptide was measured using TBA method. In the TBA colorimetric method, 1-amino-2deoxyketoses are dehydrated in boiling oxalic acid and released as 5-HMF. The sugarfree protein is removed by TCA precipitation and the 5-HMF concentration is determined colorimetrically after condensation with TBA (Fruth. 1988). One mg of the glycopeptide produced about 0.5 }lmol of 5-HMF, whereas 25 }lmol of cellobiose (8.6mg) produced 0.11 /.1 mol of 5-HMF (Fig. 9). Glucose and galactose (25 }lmole, 4.5mg), each formed 0.06 }lmol of 5-HMF. Mannotriose (25 }lmol, 12.6 mg) produced 0.15 }lmol of 5-HMF (data not shown). These results suggest that under the acidic conditions polysaccharides were dehydrated in very low yields and were hydrolyzed in high yields (or were hydrolyzed in high yields and were hydrated in very low yield) to 5-HMF in less than 0.3% yields. Thus, most of the 5-HMF produced from the partiallypurified preparation came from the carbohydrates condensing with the N-terminal and side-chain amino group in the glycopeptide. The mole number of side-chain amino groups condensing with carbohydrates to form the glycopeptide could be larger than 0.5 39.
(43) ]lmol mg-!, since 5-HMF formation from N- and O-glycosides also is small under acidic conditions (Fluckiger and Gallop. 1984). Side-chain amino groups in proteins c()ndense with carbohydrate aldehyde groups to yield glycosylamines (Schiff bases), which undergo nucleophile-catalyzed rearrangements to form 1-amino-1-deoxy-D-fructose derivatives (Amadori compounds). These compounds reduce O 2 to H 20 2 and. Fe (Ill) to Fe(ll) (Oak et al. 2000). Thus, ironcontaining glycosylated peptides could reduce O 2 to -OH via H2 0 2 as shown in Fig. 10.. 2.5. 2.0. 8. 1.5. §. ~ '". ..0. <. 1.0. C (0.94) D (0.47). 0.5 E (0.41. F (0.34 300. 400. 500. 600. 700. 800. Wavelength (nm) Fig. 9 Absorption spectra of 5-HMF prodl.lced from TBA and the carbohydrate released from the partially-purified extracell-ular glycopeptide from T. palustris cultures. Experimental procedures were as described in the text. A: 0.5 mg of the glycopeptide per ml; B: 0.25 mg of the glycopeptide per mI; C: 0.125 ]lmol of 5 -HMF per ml; D: 0.125/2 ]lmol of 5i-hydroxymethylfurfural per mI; E: 8.6 mg (25 ]lmol) cellobiose per ml; F: 4.5 mg (25 ]lmol) of glucose per ml; G: 4.5 mg (25 ]lmol) of galactose per mI. (. ): absorbance at 443nm. 40.
(44) Proposed mechanismfor wood decay by brown-rotfungi. On the basis of earlier findings and the results presented here, I propose the following mechanism for wood decay by brown-rot fungi: Fungal hyphae in the wood cell lumen secrete a glycosylated peptide of lower than 10,000 Da. This effector is able to diffuse through the S3 layer into the S2 layer and middle lamella of the cell wall, where it reduces Fe (III) to Fe(II) and chelates Fe(II). Alternatively it may reduce Fe(III) and chelate Fe(II) prior to diffusion into the S3 layer. The glycopeptide with Fe(II) catalyzes redox reactions between O 2 and an electron donor to produce -OR via R 20 2 as shown in Fig. 10. The -OR attacks cell wall constituents, causing the depolymerization of both crystalline and noncrystalline cellulose. The -OR also attacks the lignin in the cell Wall, causing a variety of reactions. These processes transform the cell wall by opening chanals through the S3 layer for enzyme diffusion. Enzymes, including endoglucanases, then can penetrate the cell wall and act on hemicellulose and noncrystalline regions of the cellulose that have been attacked by -OR.. Reactions caused by the attack of -OR on lignin, including the hydroxylation of. -... aromatic rings, demethylation of methoxyl groups on aromatic rings, and {3 -0-4 alkyl aryl ether-cleavage, produce a significant number of dihydroxy aromatic structures (Filley et al. 2002; Renriksson et al. 2000). These dihydroxy aromatic structures could reduce O 2 to R 20 2, Fe (III) to Fe(II), and the oxidized form of the glycopeptide to the reduced form (Paszczynski et al. 1999; Jensen et al. 2001).. The reaction of -OR with saccharides is initiated by hydrogen subtraction, followed, under aerated conditions, by oxidative C-C cleavage, which can cause depolymerization of saccharides and the formation of new aldehyde compounds (Uchida and Kawakishi. 1988). This series of reactions produces superoxide, which reduces Fe(III) to Fe(II) or generates R 20 2 by either non-enzymatic or SOD-catalyzed dismutation. Some of the newly-produced aldehydes also may reduce Fe(III).. The ongoing production of -OR by the glycopeptide in wood cell walls requires a constant supply of O 2 and an electron donor for the reduction of the oxidized form of 41.
(45) the glycopeptide. The identification of the electron donor will be the subject of further investigation.. ,. RI N. "'H2 H", /:0. RI. RI. R. N. NH. NH. ,. C:/. ,. ,I. ,. H2O". ,. ,. f: HH-~ H- -H r ---~""'----... H-C-OH--~. C-OH--~. C-=O , , ,. 1. H-r-OH_ HO-C-H. , HO-C-H ,. HO-C-H. H-C-O-R2. ,. HO-C-H. ,. HO-C-H. ,. R3. R3. R3. 1. bwR Fe( II ). RI , NH , H-C-H. ·OH+"OH. Fe(1II ). , C-=O , C-=O , R3. H202 2Fe( II ). ~. r. 02. RI ,. 2Fe(lII). ~. NH , H-C-H. ,. C-OH. ). ",. C-OH. electron donor. electron acceptor. (NADH). (NAD+). R3. Fig. 10 Proposed mechanism for the generation of ·OH and the reduction of Fe(IIl) to Fe(ll) by the extracellular glycosylated peptide from T. palustris.. 42.
(46) CHAPTER. N. Conclusion The rate of hydroxyl radical generation in cultures of brown-rot fungi is directly proportional to the rates of degradation of wood crystalline cellulose and lignin related compounds in the cultures. Thus hydroxyl radicals are involved in wood decay by brown-rot fungi. Most of the hydroxyl radicals produced in cultures of T. palustris are generated by the redox reaction between O2 and a certain electron donor catalyzed by a low-molecular-weight. substance.. The. low-molecular-weight. substance. was. a. glycopeptide composed of 54% protein and 42% neutral carbohydrate, containing 0.06% Fe( IT) by weight. The molecular weight as estimated by Tricine-SDS-PAGE is 7,200~1O,000,. whereas the molecular weight as determined by size-exclusion on gel-. filtration chromatography is. 1,000~5,000.. The glycopeptide reduces O2 to H20 2 and. Fe(III) to Fe( IT). Thus the glycopeptide generate hydroxyl radical via a Fenton reaction. The glycopeptide contains at least 0.5 I-L mol/mg of. a -hydroxyketone or endiol groups.. Most of the a -hydroxyketone groups are l-amino-2-ketoses produced by the Nterminal or side chain groups and carbohydrate aldehydes. I propose the following mechanism of wood decay by brown-rot fungi (Fig. 11). The hyphae in the cell lumina secrete low molecular weight substance. (1.0~5.0. kDa). The. substance diffuse into S2 layer of the cell wall, reduces Fe(III) present there to Fe( IT) and chelates Fe( IT), (or reduces Fe(III) to Fe( IT), chelates Fe( IT), and diffuses into S2 layer) (Fig. 11). The substances with Fe( IT) catalyze a redox reaction between O2 and a certain electron donor to produce ·OH via superoxide anion and H20 2; the .OH transforms the cell wall layer by cutting canals through the S3 layer for endo-cellulase diffusion, degrading the cellulose in collaboration with endo-cellulase, and modifying the lignin in the layers.. 43.
(47) Middle lamella Primary wall. · Fe( II )-glycopeptide. 'f:' elec~~n donor. 1. IIaInrJirm ~ Fe( III )-gl. 'de \. Fe( III). ·OR. S3 layer. S2 1ayer. H2Ch Decay zone. ·OH Hyphae. SI layer. .. ·OM. .:. ,~,-~. Fe( III). H~(Rr ';'$. /o~ e 0 2, electron dono. Fe( II )-glycopeptide. IIaInrJirm. C J \: elec~n donor peptide ~. H:tOz Hyphae. 0 2, electron donor. Fig. 11 Schematic representation of the process of wood degradation by brown-rot fungi. [A] Initial stage of wood cell wall degradation; [B] Advanced stage of wood cell wall degradation. Endo-cellulase can penetrate the cell wall layers of the wood .. 44.
(48) REFERENCES Backa, S., Gierer, J., Reitberger, T. and Nilsson, T. (1992) Hydroxyl radical activity in brown-rot fungi studied by a new chemiluminescence method. Holzforschung, 46,61-67. Barr, D. P., Shah, M. M., Grover, T. A. and Aust, S. D. (1992) Production of hydroxyl radical by lignin peroxidase from Phanerochaete chrysosporium. Arch. Biochem. Biophys. 298, 480-485. Beck-Anderson, 1. (1987) Production, function, and neutralization of oxalic acid produced by the dry rot fungus and other brown-rot fungi. Internat. Res. Group on Wood Preserv. Doc. No. IRGIWP/1330. Chirat, C. and Lachenal, D. (1997) Effect of hydroxyl radicals on cellulose. and pulp. and their occurrence during ozone bleaching. Holzforschung, 51, 147-154. Cowling, E. B. (1961) Comparative biochemistry of the decay of sweetgum sapwood by white-rot and brown-rot fungi. U. S. Dep. Agric. Tech. Bull. No.1, 258,1-75. Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A. and Smith, F. (1956) Colorimetric method for determination of sugars and related substances. Anal. Chern. 28, 350-356.. '". Enoki, A., Takahashi, M., Tanaka, H. and Fuse G. (1985) Degradation of lignin-related compounds and wood components by white-rot and brown-rot fungi. Mokuzai Gakkaishi, 31, 397-408. Enoki, A., Tanaka, H. and Fuse, G. (1988) Degradation of lignin-related compounds, pure cellulose, and wood components by white-rot and brown-rot fungi. Horzforschung, 42,85-93. Enoki, A., Tanaka, H. and Fuse, G. (1989) Relationship between degradation of wood and production of H 20 2-producing or one-electron oxidases by brown-rot fungi. Wood Sci. Technol. 23, 1-12. Enoki, A., Tanaka, H. and Itakura, S. (2003) Physical and chemical characteristics of glycopeptide from wood decay fungi. In: Goodell, B., Nicholas, D. D. and Schultz, T. P. Wood deterioration and preservation. American Chemical Society Press, Washington D. C. 140-152. Espejo, E., Agosin, E. and Vicuna, R. (1990) Catabolism of 1,2-diarylethane lignin model compounds by two brown-rot fungi. Arch. Microbiol. 154,370-374. 45.
図



+7





Outline
関連したドキュメント
Degradation mechanism of lignin model compound by ozonolysis l: veratrole, 2: guaiacol, 3: catechol, 4: quinone, 5: muconic acid dimethylester, 6: muconic acid monomethylester,
Furthermore, the relationships between mechanical properties of cores taken from various portions of bridge pier and ASR degradation ranks classified by the various inspection
Thecriminalcaseswithstimulantssucllasmethamphetamine,designersdrugsand
東京大学 大学院情報理工学系研究科 数理情報学専攻. [email protected]
情報理工学研究科 情報・通信工学専攻. 2012/7/12
A class of pairs of weights related to the boundedness of the Fractional Integral Operator between L
In 1997, Harboure, Salinas and Viviani in [HSV], gave necessary and sufficient conditions on the weights for the boundedness of the fractional integral operator I γ from weighted
Related to this, we examine the modular theory for positive projections from a von Neumann algebra onto a Jordan image of another von Neumann alge- bra, and use such projections
As an important consequence of Theorem 1, we deduce in Corollary 3.11 the following prime-to-p version of Uchida’s Theorem on isomorphisms between absolute Galois groups of
