熊本大学学術リポジトリ
Kumamoto University Repository System
Title 直腸肛門奇形に関する発生学的解析
Author(s) 須田, 博子 Citation
Issue date 2012‑03‑23
Type Thesis or Dissertation
URL http://hdl.handle.net/2298/25188 Right
目次
1. 要旨・・・p.3
2. 発表論文リスト・・・p.5
3. 謝辞・・・p.6
4. 略語一覧・・・p.7
5. 研究の背景と目的・・・p.9
6. 実験方法・・・p.20
1) Sd SktGt/+ SktGtマウスの作製
2) Sd SktGt/+ SktGtマウスの腎臓、直腸肛門部の表現型解析 3)直腸肛門形成期における組織学的解析
4)Skt遺伝子発現パターン解析
5) 遺伝子発現プロファイリング解析
①パスウエイ解析
②直腸肛門形成関連遺伝子発現解析 6) 統計学的解析
7. 実験結果・・・p.27
1) SktGt 変異を加えたSdマウスの表現型解析
2)直腸肛門奇形モデル [Sd SktGt/+ SktGt ]における直腸肛門発生期の表現型解析 3)直腸肛門周辺のSkt遺伝子の発現パターン解析
4)脊索と直腸肛門奇形
5) 遺伝子発現プロファイリング解析
① シグナルパスウェイ解析
② 直腸肛門形成関連遺伝子発現解析
8. 考察・・・p.42
9. 結語・・・p.49
10. 参考文献・・・p.50
1. 要旨
<目的>直腸肛門奇形(鎖肛)は、新生児外科の治療対象となる先天性疾患のうちで最も 頻度の高い疾患であるが、その発生機序に関する知見は少ない。また、ヒト鎖肛のモデル 動物になりえる100%の発生率を示す鎖肛モデルの報告はない。本研究は、高頻度に鎖肛 を発症するDanforth’s short tail(Sd)変異マウスにSickle tail遺伝子(Skt)の変異(SktGt)を加えた
(Sd SktGt/+ SktGt)マウスで鎖肛発生率が100%になることに注目した。本研究の目的は、総
排泄腔(cloaca)発生におけるSktと、鎖肛モデル(Sd SktGt/+ SktGt)胚の総排泄腔発生異常を検 討することで、S k tが直腸肛門発生と鎖肛発症に関連する時期及び組織を同定し、直腸肛 門形成に関与する分子メカニズムを解明することである。
<方法>コントロール胚、鎖肛モデル胚に対して直腸肛門形成期である胎生9.5日(E9.5)か
らE13.5においてHE染色を行った。また、(SktGt)のレポーター遺伝子を利用して総排泄腔周
辺のSkt発現をXgal染色により同定した。また、E10.5の臀部組織のRNAを用いてマイクロ アレイ解析によるパスウェイ解析を行った。加えて、直腸肛門形成に関連があると報告さ れている遺伝子についてIn situ hybridization (ISH)法にて検証を行った。
<結果>鎖肛モデル胚は、E10.5より背側部のcloaca plateの発生障害と、E11.5より尿直腸 中隔の低形成を示した。E13.5では、直腸肛門管は肛門窩に開口せず、その末端は尿生殖 洞と交通していた。Sktの発現は、正常発生ではE9.5よりcloaca plateで始まり、E11.5では
cloaca plateと背側の間葉細胞で認めるようになり、E13.5まで維持されていた。鎖肛モデル
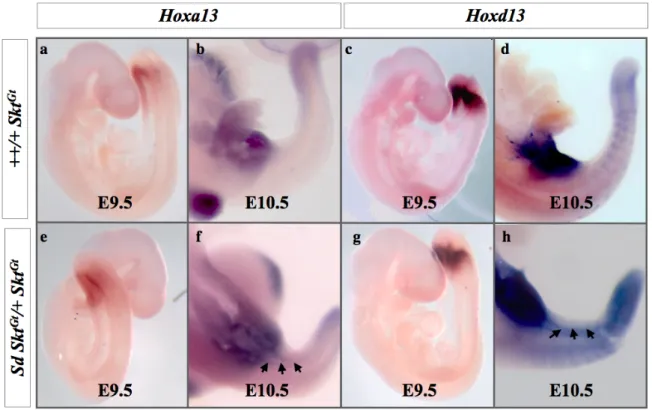
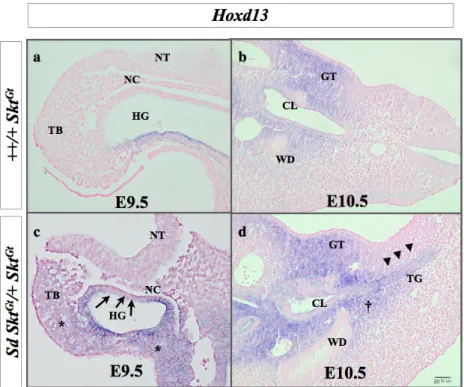
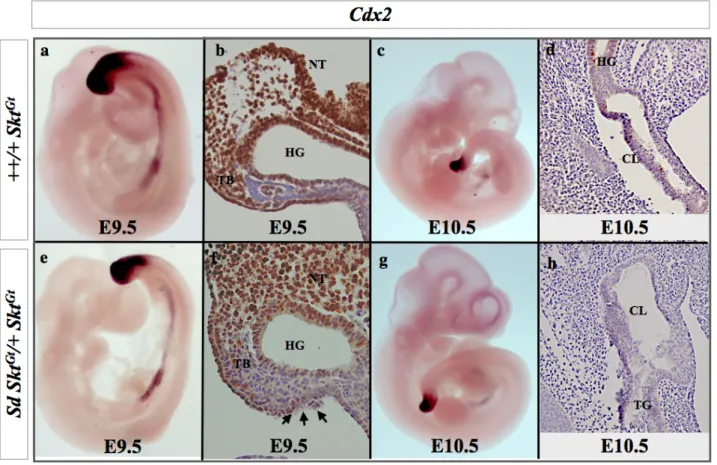
では短縮したcloaca plateとその背側に直腸肛門のかわりに増殖している間葉細胞で認め た。発現プロファイリング解析により、鎖肛モデルではCdx、5’Hox遺伝子の発現変化を示 しており、Hoxa13、Hoxd13は尾側腸管での異所性発現を示し、Cdx2はcloacal plateとその 周辺組織で発現が低下していた。
<考察>Skt遺伝子の発現は、正常直腸肛門の発生過程でcloaca plateで主に認め、鎖肛モデ ルの発生過程では、短縮したcloaca plateと増殖した間葉組織に認めた。このことは、Skt遺
伝子が、cloaca plateの分子マーカーになりえることを示しただけでなく、鎖肛発生の病態
にも関与していることを示している。マイクロアレイとI S H法解析の結果は、鎖肛発生に
Hoxa13、Hoxd13、Cdx2発現の変動が関与していることを示唆した。
<結論>高頻度に鎖肛を発症するモデルマウスを用いることによって、S k t遺伝子が、
cloaca plateの分子マーカーになり、鎖肛発生の病態にも関与していることを示した。更
に、Hoxa13、Hoxd13、Cdx2発現の変動も関与していることが示された。
2. 学位論文の骨格となる参考論文 (1) 関連論文および、参考論文
Hiroko Suda・Kwang-Jong Lee・Kei Semba・Fumie Kyushima・Takashi Ando・Masatake Araki・ Kimi Araki・Yukihiro Inomata・Ken-ichi Yamamura
The Skt gene, required for anorectal development, is a candidate for a molecular marker of the cloacal plate
Pediatr Surg Int. 2011;27(3):269-73.
(2) その他の論文のリスト
Takashi Ando, Kei Semba, Hiroko Suda, Akira Sei, Hiroshi Mizuta, Masatake Araki, Kuniya Abe, Kenji Imai, Naomi Nakagata, Kimi Araki, Ken-ichi Yamamura
The floor plate is sufficient for development of the sclerotome and spine without the notochord.
Mech Dev. 2011;128(1-2):129-40.
岡島英明, 須田博子, 猪股裕紀洋. 胆道閉鎖症における遺伝的素因と膵管胆道合流異常症. 小児外科. 2008;40:41-44.
3. 謝辞
熊本大学生命資源研究・支援センター・疾患モデル分野において熊本大学生命資源 研究・支援センター・疾患モデル分野・山村研一教授と熊本大学大学院医学研究科 小児外科学分野・猪股裕紀洋教授の御指導の下、本研究を行いました。多くの御指 導を頂き、深く感謝いたします。また、熊本大学生命資源研究・支援センター・疾 患モデル分野・荒木喜美准教授、熊本大学生命資源研究・支援センター・バイオ情 報分野・荒木正健准教授、熊本大学大学院医学研究科運動骨格病態学分野・仙波圭 先生には、日々の実験研究から論文作成まで幅広い御助言を頂きました。また、疾 患モデル分野、小児外科学分野の皆様には有形無形の多くの御協力、ご支援を受け ました。
心より感謝いたします。
4. 略語一覧
ABC法 : avidin-biotin complex bHLH:basic helix-loop-helix β-gal:β-galactosidase
β-geo:β-galactosidaseとneomycin耐性遺伝子の融合タンパクをコードした遺伝子 BMP:bone morphogenic protein
BSA :bovine serum albumin, ウシ血清アルブミン
CAG : cytomegarovirus enhancer, chicken beta actin promoter, rabbit beta-globin polyA cM:centimorgan, センチモルガン
Cdx : caudal type homeobox
DAPI : 4',6-diamidino-2-phenylindole
DNA:Deoxyribo nucleic acid, デオキシリボ核酸 E:Embryonic day, 胎生
Fgf : fibroblast growth factor
GTP : guanosine triphosphate, グアノシン三リン酸 GDP : Guanosine diphosphate, グアノシン二リン酸 Gapdh : Glyceraldehyde 3-phosphate dehydrogenase HE :Hematoxylin-Eosin
HH : Hamburger and Hamilton Hox : homeobox gene
IRES : Internal ribosome entry site.
n.s.:not significant
PBS:phosphate buffered saline
PBT:phosphate buffered saline containing 0.1% Tween-20 PCR:polymerase chain reaction, ポリメラーゼ連鎖反応 PFA:Paraformaldehyde
RT:room temperature, 室温 SD:Standard Deviation, 標準偏差 Sd:Danforth’s short tail マウス Skt:Sickle tail遺伝子
SktGt:B6;CB-SktGtAyu8021IMEG
Shh:Sonic hedgehog TBE:Tris-borate-EDTA
5. 研究の背景と目的 A. 直腸肛門奇形
直腸肛門奇形とは、直腸および肛門部の奇形を指し、一般的には鎖肛と呼ばれることが 多い。出生5,000に対して1例の割合で発症し、男女比は3:2と男児にやや多い。本奇形 は直腸閉鎖症のように正常肛門を有するものから、全く肛門形成のみられないもの、外表 や泌尿生殖器と瘻孔を形成するものまで幅広い病型がある。鎖肛は古くからよく知られた 疾患であり、肛門が閉鎖した患児の会陰部に開口部を作製するという治療が行われてい た。しかし、現在でいういわゆる ”低位” 鎖肛患児は救命することができたが、”高位” 鎖 肛患児は救命しえなかった。1835年にAmussatが直腸壁を皮膚に縫合したのが肛門形成術 の最初だといわれている。20世紀初期になり、低位鎖肛に対しては、会陰式肛門形成術 を、高位鎖肛に対しては新生児期に人工肛門造設を行い、成長を待って腹会陰式または仙 骨会陰式根治術を行うという二期的手術が行われるようになった。しかし、腹会陰式や仙 骨会陰式根治術では直腸肛門周囲筋群全体の把握が困難であるため術後排便機能障害を来 す例も多く1980年代にPenaによりposterior sagittal anorectoplastyが開発された [1]。近年で も直腸肛門周囲筋群の解剖を理解し、周辺の神経や筋を損傷せずに正確に直腸を筋束の中 心に引き下ろし排便機能や泌尿生殖機能を維持するために様々な工夫が考案されている。
また、泌尿生殖器、心臓、脊椎などの奇形を高頻度に合併するのも直腸肛門奇形の大きな 特徴であり、診断・治療の際には合併奇形の有無に留意することが必要である [2, 3]。直 腸肛門部については手術が行われれば生命予後の良い疾患であり、生命予後は合併奇形に 左右される。
B. 直腸肛門奇形の臨床的病型
直腸肛門奇形は前述したように病型が多様であり、閉鎖部位と泌尿生殖器系との間に存 在する瘻孔の有無・高さにより術式アプローチが異なるため病型の分類が重要となってく る。1934年に、Ladd & Grossが本症をI〜IV型に分類し、その後、Stephenらにより排便機 能において重要となる恥骨直腸筋を考慮した分類がメルボルン分類が提唱された。現在 は、これを単純化し整理したWingspread分類が汎用されている [1]。この分類では、男児と
女児で瘻孔形成部位など病型が異なるため性別による分類と、直腸盲端の高さによる高 位、中間位、低位による分類が行われている。高位は直腸盲端が恥骨直腸筋より頭則にと どまっているもの、中間位は恥骨直腸筋内にとどまるもの、低位は更に肛門側に伸長して いるものと定義され臨床的には倒立X線像や直腸肛門管の造影にて筋肉の存在の目安と成 る、恥骨・座骨の位置を基準に診断される。性差による分類では、泌尿生殖器と連続する 瘻孔の位置関係により、男児では尿道と交通した直腸前立腺部尿道瘻、直腸尿道球部瘻、
女児では生殖器と交通した直腸膣瘻、直腸前庭瘻、肛門前庭瘻、もしくは尿道・膣・直腸 が一孔となった総排泄腔奇形に分類される。
表1 Wingspread Classification (1984)
男児 女児
高位 高位 1. 肛門直腸無発生
a) 直腸前立腺部尿道瘻 b) 無瘻孔
2. 直腸閉鎖
1. 肛門直腸無発生 a) 直腸膣瘻 b) 無瘻孔 2. 直腸閉鎖 中間位
中間位 1. 直腸球部尿道瘻
2. 瘻孔を伴わない肛門無発生
1. 直腸前庭瘻 2. 直腸膣瘻
3. 瘻孔を伴わない肛門無発生 低位
低位 1. 肛門会陰部皮膚瘻
2. 肛門狭窄
1. 肛門前庭瘻 2. 肛門会陰部皮膚瘻 3. 肛門狭窄
総排泄腔奇形
C. 直腸肛門奇形と合併奇形
直腸肛門奇形は奇形症候群の一つとしてもよく知られており、他の臓器奇形を合併する ことが多い。合併奇形は約50%の患児でみられ、低位鎖肛よりも高位鎖肛に高率に合併す る。合併頻度の高い臓器は脊椎、腎臓であり、双方を合併する疾患は尾部退行症候群とし
てもよく知られている。その他、心臓、生殖器、食道気管、消化管奇形などを合併する
[4] [5] 。以下に直腸肛門奇形を合併する奇形症候群を示す(表2)。
表2 直腸肛門奇形を含む奇形症候群(文献1より引用)
VACTERL (Vertebral anomalies, anal atresia, cardiac malformations, tracheoesophageal fistula, renal anomalies, and limb anomalies)
MURCS (Mullerian duct aplasia, renal aplasia, and cervicothoracic somite dysplasia) OEIS (Omphalocele, exstrophy, imperforate anus, and spinal defects)
Axial mesodermal dysplasia Klippel-Feil syndrome
Sirenomelia-caudal regression Trisomy 21
Trisomy 13 Trisomy 18
Pallister-Killian syndrome Cat-eye syndrome
Parental unidisomy 16
Deletion 22q11 syndrome (del22q11.2) Currarino syndrome
Pallister-Hall syndrome Townes-Brock syndrome Ulnar-mammary syndrome Okihiro syndrome
Rieger syndrome
Thanatophoric dwarfism Hirschsprung disease Feingold syndrome Kabuki syndrome
Optitz BBB/G syndrome Johanson-Blizzard syndrome Spondylocostal dysostosis
Short rib – polydactyly syndrome Baller-Gerold syndrome
Ciliopathies Fraser syndrome Lowe syndrome Heterotaxia FG syndrome
X-linked mental retardation MIDAS syndrome
Christian syndrome
D. 直腸肛門奇形の病因
直腸肛門奇形の病因には、多くの先天奇形と同様、多因子が関与しているとはいわれて いるが、遺伝学的因子、環境因子も含めほとんど分かっていない。遺伝学的な見地から は、直腸肛門奇形患児の子孫で直腸肛門奇形発症率が出生100に対し1、と通常の出生5000 に対し1という発症率と比較し上昇するという報告や、直腸肛門奇形を含む奇形症候群患 児が複数存在する家系の報告がある [6] 。また、直腸肛門奇形を合併するTownes-Brocks syndrome [7, 8]、Currarino's syndrome [9, 10]や Pallister-Hall syndrome [11, 12]などは特定の遺 伝子の変異による常染色体優性遺伝疾患であることが知られており、ダウン症(21トリソ ミー)患児では通常の直腸肛門奇形と比較し瘻孔のない病型の占める割合が上昇するとの 報告もある[2, 5, 6]。これらより、直腸肛門奇形の病因に何らかの遺伝学的因子が絡んで いるということが示唆される。一方、疫学的解析では、母性因子として、妊娠時のアル コール摂取[13]、喫煙、カフェイン摂取 [14]、薬物摂取 [15]、ビタミン剤摂取、妊娠初期
の発熱 [16]、父性因子として喫煙、不妊、職業 [15] [16] などが直腸肛門奇形を合併する危
険因子として報告されている。
E. 直腸肛門発生
発生過程において直腸肛門は、ヒトでは胎生5週より8週、マウスでは胎生10.5日より 13.5日に形成される。後腸尾側端が膨らんで総排泄腔が形成され、この総排泄腔が尾側に 発達する尿直腸中隔により、腹側の尿生殖洞と背側の直腸肛門洞とに分離される。総排泄 腔の腹側、尿膜付着部尾側にはcloacal plateと呼ばれる中胚葉を欠いた内胚葉と外胚葉の2 層からなる組織が形成され、このcloacal plateも総排泄腔の分離にともない腹側の尿生殖膜 と背側の肛門膜に分離され、引き続きこれらが消失することによりそれぞれ尿道と肛門が
開口する[17, 18]。動物の直腸肛門奇形モデルでは、古くは鎖肛ブタ家系の解析にてcloacal
plate背側の形成障害が報告され[19, 20]、以後マウスやラットの自然発生、薬剤誘導性直腸
肛門奇形モデルでも同じくcloacal plateの形成が障害されていることが報告されている[21, 22]。すなわちcloacal plateが直腸肛門発生において重要な組織であることを示唆してい る。
F. 直腸肛門発生に関連する遺伝子
消化管の発生については、鶏を用いた解析が進められてきた。 Hamburger and Hamilton
(HH) stage 8に 前腸門、HH stage 13に後腸門が形成されそれぞれ後方、前方に伸長して
最終的にHH stage 24-28に卵黄嚢茎周囲で前腸と後腸が結合し、前腸、中腸、後腸が形成
される。この過程において、四肢、体節、神経管の形成を司るモルフォゲンとしてもよく
知られるSonic hedgehog (Shh) 遺伝子がまず前腸門、後腸門で発現し発生に従い腸管内胚葉
全体で発現するようになる。そして、このShhの発現が中胚葉でのBmp4、Hox遺伝子など の発現を誘導し、続いてこれらの遺伝子が内胚葉に作用するという内胚葉中胚葉相互作用 を通して、総排泄腔を含めた腸管の形成と領域化が行われるといわれている[23, 24]。以 後、直腸肛門発生においては、遺伝子改変マウスを用いた解析により直腸肛門奇形を表現 型とするマウスが報告されるようになり、まだ未解明な部分が多いが、直腸肛門発生に関 与する遺伝子制御の知見が蓄積されつつある。最初に報告された直腸肛門奇形を合併する 遺伝子改変マウスはShhシグナル経路改変マウスであり、Shh-/ - マウスは総排泄腔奇形、そ の下流の転写因子であるGli2の欠損では鎖肛、Gli3の欠損では肛門狭窄、Gli2とGli3の二重 変異マウスでは総排泄腔奇形の表現型となることが報告された [25, 26]。Shhシグナルが量 依存的に直腸肛門の形成を制御していると考えられる。H o x遺伝子は動物の分節的コン パートメントを決定する重要な遺伝子であるが、このHox遺伝子についても直腸肛門発生 への関与が報告されている。Hox遺伝子は脊椎動物では異なる染色体上に4つのクラスター を形成し、塩基配列の相同性から13のグループに分けられ、3' 側から番号がつけられてい る。発生過程ではこの順番に対応して前後軸に沿って入れ子的に発現し、領域特異的な形 態形成を支配している。Hox遺伝子の総排泄腔での発現は、内胚葉でHoxb-8、Hoxb-9、 Hoxc-9、Hoxa-13、Hoxd-13、中胚葉でHoxa-11、Hoxd-11、Hoxd-12、Hoxa-13、Hoxd-13が 発現している。この中で直腸肛門奇形をひきおこす遺伝子として報告されているのは、
Hoxd-12、Hoxa-13、Hoxd-13である[27]。 Hoxd-12 -/ -、 Hoxd-13 -/ - は肛門括約筋の発生が障 害されており、その結果、肛門直腸脱を引き起こす [28]。 Hoxa13 +/ - ;Hoxd13 -/ - は高度の 直腸上皮、平滑筋層の形成障害をきたし直腸低形成をきたす[29]。直腸肛門形成に関して はこれらの5‘Hox側遺伝子が肛門括約筋群も含めた直腸肛門発生に関与していることを示
唆している。また、Hox遺伝子と共通の祖先から分かれたとされるParaHox遺伝子の一つ
であるC d x遺伝子も直腸肛門形成に関与している。C d x 2は胎生初期に栄養外胚葉で発現
し、胎生8 . 5日ごろより腸管を含めた胚尾側で発現を認めるようになる遺伝子である
[30] 。Cdx2へテロ変異マウスの大腸では、食道・胃・小腸などのより口側の消化管組織を
含むポリープを形成することが知られており、Cdx2遺伝子は消化管上皮の運命決定に重要 であると考えられている[31] 。また、Cdx遺伝子はHox遺伝子と同様に前後軸形成において も重要な遺伝子で、後方(尾側)の形成に関わっており、Cdx変異マウスでは尾部の低形 成をきたす[32]。直腸肛門形成については、胎生9.5日より腸管内胚葉で特異的にCdx2の発 現をノックアウトすると鎖肛となることが報告されている[33]。ビタミンA活性化体であ るレチノイン酸は、発生における重要な因子の一つであり、妊娠初期のビタミンA過剰投 与により奇形合併率が上昇するということもよく知られている。動物実験でも、レチノイ ン酸を妊娠したラットやマウスに投与し直腸肛門奇形モデルを作製する方法がよく用いら
れている[21, 34]。発生過程において、胎生初期にレチノイン酸代謝酵素によりつくられる
前方優位のレチノイン酸濃度勾配が前後軸の決定に重要である。例えば、レチノイン酸分 解酵素であるCyp26a1は尾側で発現するが、Cyp26a1を欠損させたマウスでは尾側の形成 が障害され後腸が欠損することが知られている[35-37]。Fgf10は腸管においては、胎生10.5 日より後腸間葉組織で発現が始まり、以後口側に発現が拡大していく遺伝子である。大腸 上皮の細胞増殖への関与が知られている。 Fgf10 -/ - の直腸肛門発生は、胎生10.5日までは ほぼ正常に発生し、胎生10.5日以降になると、後腸の狭小化が始まり、胎生14.5日に大腸 が消失し鎖肛となると報告されている。このマウスでは鎖肛となるが、瘻孔は形成しな い。この報告ではcloacal plateについては触れられていないが、cloacal plateを介さない直腸 の発生障害単独の機序で直腸肛門奇形を引き起こしているとも考えられる[38][39]。胚発 生において重要なシグナル経路の一つであるWntシグナル経路の異常も直腸肛門発生障害 を引き起こす。Wnt5aは胎生10.5日の総排泄腔周囲では、生殖結節で発現している遺伝子 である。直腸肛門奇形モデルであるSdマウスとレチノイン酸誘導マウスの双方でこの発現 が低下していることが報告されており[21]、Wnt5a-/ - では瘻孔を形成する鎖肛と瘻孔を形成 しない鎖肛の表現型となる[40]。Wntシグナル経路の一つであるDact1遺伝子の変異マウス
も直腸肛門奇形となる。このマウスではDact1の原始線条での発現が消失し、Wntシグナル 経路の中でもWnt/βcatenin経路とPCP経路の異常が原因で直腸肛門発生が障害されると考 えられている[41]。直腸肛門発生にWntシグナルも重要な役割を担っていると考えられる。
S a l l遺伝子は亜鉛フィンガードメインをもつ転写因子として知られている遺伝子であり、
総排泄腔でも発現している。ヒトでは4つのSall遺伝子が知られているが、その中でもSall1 はTowens-Brocks syndromeの原因遺伝子として、Sall4はOkihiro sundromeの原因遺伝子で あるとされている。マウスではSall1-/ - では直腸肛門の表現型はないが[42] 、Sall4+/ - で肛門 狭窄、 Sall1+/ - Sall4+/ - で直腸肛門奇形となる。Sall1+/ - Sall4+/ - ではtruncated Sall1がSall4で 形成されたヘテロダイマーが、ドミナントネガティブに働き直腸肛門奇形を発症するとい うメカニズムが推定されている[43]。点突然変異誘発剤であるエチルニトロソウレアで誘
発されたVACTER症候群類似のマウスではプロタンパク質変換酵素であるPcsk5の突然変
異が報告されている。このマウスではPcsk5のターゲット遺伝子であるTGFβファミリー分 子Gdf1やHox遺伝子、また、ヒトのCaudal regression syndromeの原因遺伝子と推定されてい るMnx1遺伝子の発現異常を認めている。Gdf1 -/ - も直腸肛門奇形となる[44]。
このように、直腸肛門形成に関与しているであろう遺伝子の報告は非常に多岐に及ぶ が、直腸肛門発生を制御する一連の分子メカニズムを説明しうる知見は未だ得られていな い。また、これらの遺伝子改変マウスではほぼ100%に直腸肛門奇形を合併するが、直腸 肛門部以外の臓器でも重篤な奇形を合併しており、奇形症候群の範疇に該当しない一般的 なヒト鎖肛モデルとは異なっている。
表3 直腸肛門奇形を表現型とする遺伝子改変マウス
遺伝子型 直腸肛門奇形の表現型
(瘻孔の有無は記載のあったもののみ記載) 文献
Shh-/ - 総排泄腔奇形 [25, 26]
Gli-2-/ - 鎖肛 [25]
Gli-3-/ - 肛門狭窄 [25]
Gli-2-/ -; Gli-3+/ - 総排泄腔奇形 [25]
Gli-2+/ -; Gli-3-/ - 総排泄腔奇形 [25]
Gli-2-/ -;Gli-3-/ - 総排泄腔奇形 [25]
Hoxd13 -/ - 直腸脱 [28]
Hoxa13 +/ -; Hoxd13 -/ - 直腸低形成 [29]
Cdx2-/ - (conditional) 鎖肛 [33]
Cyp26a1-/ - 鎖肛(sirenomelia) [36]
Fgf10-/ - 鎖肛(瘻孔なし) [39]
Wnt5a-/ - 鎖肛(瘻孔あり25%, 瘻孔なし75%)) [40]
Dact1-/ - 鎖肛 [41]
Sall4+/ - 肛門狭窄 [43]
Sall1+/ - Sall4+/ - 鎖肛 [43]
Pcsk5 鎖肛 [44]
Gdf1 -/ - 鎖肛 [44]
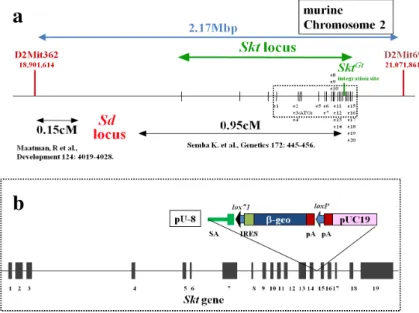
G. Danforth’s short tail (Sd) マウスとSickle tail (Skt) 遺伝子
Sdマウスは、1930年に Danforthにより発見された短尾が特徴的な半優性変異マウスであ る。脊椎の他に泌尿生殖器と直腸肛門の奇形も表現型とする[45, 46]。 Sdホモ変異体では 尾部形成障害が重篤であり、臀部はほぼ形成されない。 Sdホモ変異体は出生後すぐに死 亡し、Sdヘテロ変異体でも半数が死亡する。直腸肛門奇形は Sdホモ変異体では全ての個 体で、Sdヘテロ変異体になると約半数の個体で合併する。直腸肛門奇形の病型はSdホモ変 異体では鎖肛の中でも重篤な直腸膀胱瘻を合併する鎖肛もしくは総排泄腔奇形となり、外 生殖器の低形成も伴う。一方、Sdヘテロ変異体では Sdホモ変異体より表現型は軽度であ り、直腸尿道瘻を合併する鎖肛病型が多く、外生殖器もほぼ正常である。Sd遺伝子につ いては、Sd遺伝子座がマウス第2染色体に位置することは分かっているが遺伝子の同定に
は至っていない。そのため胎生早期の表現型が明らかでない時期では遺伝子型の決定が不 可能であり、Sdマウスを用いた発生学的解析は滞ってきた[47-49](図1)。
Cre-loxPシステムは部位特異的リコンビナーゼCreがloxpという特定の配列を認識し標的
変異を起こすシステムである。我々の研究室ではこのCre-loxPシステムを応用したトラッ プベクターを用いたランダムなマウスミュータジェネシスを行っている。使用しているト ラップベクターはスプライスアクセプター、IRES配列、lacZとneomycin耐性遺伝子の融合 タンパクであるβ-geo、polyAシグナルを持っており、遺伝子のエクソン間に挿入されると その部位より下流の遺伝子機能を破壊することができる[50]。我々のグループでは過去、
このトラップベクターpU-8をES細胞に導入するジーントラップ法により脊索でレポーター 遺伝子を発現するSktGt変異マウスラインを作製した [51]。この際同定したSkt (Sickle tail)は 新規遺伝子であり、マウス第2染色体Sd遺伝子座の近傍に位置している。そして、Sd変異 マウスとSktGt変異マウスを掛け合わせた結果、Sd変異遺伝子座の0.95cM遠位にSkt 遺伝子 が位置していることが明らかになった[51](図2)。この掛け合わせの際、2つの変異が cis -configurationで存在するダブル変異マウス(Sd SktGt /+ +)が得られ、この Skt locusに挿入
されたlacZ遺伝子をSd変異のgenotypingマーカーとして使用することが可能となった。 Skt
遺伝子は、成体では脊椎、椎間板随核、胎生期では脊索で発現し椎間板形成に影響を与え ている。SktGtヘテロ変異マウスは正常だがSkt ホモ変異マウスでは尾椎先端の椎間板形成 異常による尾先端の屈曲変形を認める[51]。直腸肛門部は SktGt ホモ変異マウスでも正常 な形態であり、また、成体まで成長し妊孕性も持つ。我々は SktGt 変異をSdヘテロ変異マ ウスに加えたダブル変異マウスの脊椎形成の解析では、[Sd +/+ +] では尾椎以下の欠損で あったが、 [Sd SktGt/+ +]、 [Sd SktGt/+ SktGt ]ではより上位の腰椎から欠損し脊椎形成が加 算的に重篤化することを見いだしSkt遺伝子が脊椎形成に影響を与えていることを示した [52](表4)。更に、 [Sd SktGt/+ SktGt ]では、生後早期にほぼ100%死亡しており、直腸肛門 の表現型を解析したところ100%の個体で直腸肛門奇形を合併していた。
これまで、100%の発症率を示す一般的なヒト鎖肛の病型に近い直腸肛門奇形モデルの 報告はなく、また、Sdマウスの胎生期の解析は遺伝子決定が不可能であったため困難で あったが、 [Sd SktGt/+ SktGt ] は、①Sdホモ変異マウス様の重篤な臀部奇形を含めその他の
重篤な臓器奇形を合併しない、②直腸肛門奇形を100%で合併する、③遺伝子型の決定が 可能であるという利点を備えている。
今回の研究の目的は、このマウスラインを直腸肛門奇形モデルとして、総排泄腔の発生 におけるSktと鎖肛モデル [Sd SktGt/+ SktGt ] 胚の総排泄腔発生異常を検討することで、Skt遺 伝子が直腸肛門発生と鎖肛発症に関連する時期及び組織を同定し、直腸肛門形成に関与す る分子メカニズムを解明することである。
図1. SdマウスとSkt変異マウスの表現型
(a)SdマウスとSkt変異マウスの表現型を示した。外観上、Sdへテロ変異マウスでは尾が短くなっており、
Sdホモ変異マウスになるとさらに尾部の形成障害が重篤になる。(b) Sktへテロ変異体では尾椎先端の表 現型はないが、Sktホモ変異体では尾椎先端部が屈曲する。
図2. Sd locusとSkt locusの位置関係とSktGtの構造
(a) Sd, Skt locusのPhysical / Genetic Map。点線枠は(b)の位置を表している。Sd locusとSktGtはマウス2番染色体 の近位に存在し、掛け合わせにより0.95cMの遺伝的距離にあることが報告されている。 (b) SktGtの構造。Skt 遺伝子の14、15番exon間にtrapベクターpU-8が挿入されている。 black box: exon
表 4. 脊椎奇形発生率 ( [52]より引用)
a
b
6. 実験方法
1) Sd SktG/+ SktGtマウスの作製
pU-8トラップベクターを用いたジーントラップ法は荒木等により報告されている[50]。 ジーントラップクローンのES細胞とICR (Charles River)の桑実胚を用いたアグリゲーション 法によりキメラマウスを作製し、C57BL/6 (CLEA)の雄と交配しF1ヘテロ接合体を得た。
Danforth’s short tail (Sd)マウスはジャクソン研究所 (Bar Harbor, ME, USA)より購入し、胚移 植により繁殖させた。Sd SktG/+ SktGtマウスはSd +/+ + マウス(C57BL/6 genetic background)と C57BL/6 genetic backgroundを持つSktGt/Gt マウスと交配させる事により得られた、Sd変異と SktGt変異が同一染色体に位置しているマウス[Sd SktGt /+ +; cis configuration]をSktGt/Gt マウス と交配もしくは胚移植することにより繁殖させた。遺伝子型はgenome PCR法にてSkt wild-
type alleleを検出することにより決定した。Skt遺伝子の14番目イントロンに位置するGTS
プライマー (5’- CCACCCCTACATGTGTCTTT -3’)とGTAプライマー(5’-
CGAGTAAGTAACATCCCTCC -3’)を使用し、これにより339bpのPCR産物を得ることで確
認することができた。また、この交配の際に得られる同胞である [+ +/+ SktGt ] マウスをコ ントロールとして使用した。
2) Sd SktG/+ SktGtマウスの表現型解析
Sd SktG/+ SktGtマウスの腎臓、直腸肛門部の表現型を明らかにするために胎生19.5日胚を
用いて解析を行った。標本は横隔膜より尾側を採取し、3.7% formaldehyde/PBSで3日間固 定後、全自動パラフィン置換装置 HISTOS5 (Leica Microsystems)を用いてパラフィン置換を 行い、包埋した後に4μmの厚さで薄切した。Hematoxylin-Eosin (HE)で染色後、正中矢状断 像にて正立顕微鏡を用いて観察した。
3) 直腸肛門形成期における組織学的解析
直腸肛門発生期である胎生9.5日から胎生13.5日の胚を用いて解析を行った。標本は採取 後、3.7% formaldehyde/PBSで一晩固定し、全自動パラフィン置換装置 HISTOS5 (Leica
Microsystems)を用いてパラフィン置換を行い、包埋した後に4μmの厚さで薄切した。
Hematoxylin-Eosin (HE)で染色後、正中矢状断像にて正立顕微鏡を用いて観察した。
4) Skt遺伝子発現パターン解析
Skt遺伝子単独の欠損では直腸肛門部に表現型をきたさないが、Skt遺伝子の変異が加わ ることにより、Sdマウスの直腸肛門奇形発生率を上昇させていた。Skt遺伝子の直腸肛門 奇形発生への関与を明らかにするため、直腸肛門形成期における総排腔周辺でのS k t遺伝 子発現パターンを解析した。また、S k t遺伝子が脊索でも発現している事を利用し[ 5 1 ]
[52] 、総排泄腔レベルの脊索の形態についても観察を行った。トラップベクター内に挿入
されているレポーター遺伝子(βガラクトシダーゼ遺伝子)を用いて、Whole-mount X-gal
染色法 (Allenら[53] )にて解析した。サンプルは固定液(1% formaldehyde, 0.2%
glutaraldehyde, and 0.02% NP-40 in phosphate-buffered saline (PBS))にて室温で0.5時間固定 後、PBSにて2回洗浄した。その後、X-gal染色液(5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 2 mM MgCl2, 0.5% X-gal in PBS)にて30℃にて一晩染色した。その 後、2回PBSで洗浄し、4% paraformaldehydeにて後固定を施行した。 Whole body 観察の際 には、エタノールステップ(25%、50%、70%、100% and 100%, それぞれ1 時間反応) にて脱
水後、benzylalcohol/benzylbenzoate (1:2)により検体を透明化し倒立顕微鏡にて観察した。
組織学的解析を行う際には X-gal 染色した胚を後固定後、パラフィン置換を行い、8μmの 厚さで薄切しNuclear Fast red で対比染色を行い正立顕微鏡にて観察した。
5) 遺伝子発現プロファイリング解析
S d遺伝子は同定されておらず、S dマウスの奇形発症メカニズムは分かっていない。ま た、Skt遺伝子についてもタンパク質を合成することは分かっているが、その蛋白の機能は 不明である。そこで、 [Sd SktGt/+ SktGt ] の奇形発症メカニズムを解明するため直腸肛門形 成初期の遺伝子発現プロファイリング解析を行った。そして、この遺伝子発現プロファイ ルを用いて ①パスウェイ解析と ②直腸肛門奇形関連遺伝子の中で変化を認めた遺伝子に ついての検証を行った。
遺伝子発現プロファイリング解析にはマイクロアレイ法を用いた。cloacal plateの形成障 害が始まる胎生10.5日胚(n=7)の下肢より尾側を切除採取し、凍結破砕後 RNAeasy mini kit(QIAGEN)を用いてtotal RNAを抽出し、解析はFilgen社に委託し全マウス36227遺伝子 について解析(CodeLinkTM Mouse Whole Genome Bioarray)し、コントロール [+ SktGt/+ +]
と直腸肛門奇形モデル [Sd SktGt/+ SktGt] で比較した。。
①パスウエイ解析
マイクロアレイ法により得られた遺伝子発現プロファイルより発現強度比 [Sd SktGt/+
SktGt] / [+ SktGt/+ +]が2.0以上の遺伝子、もしくは、0.5以下であった遺伝子を抽出し、抽出 した遺伝子をMicroarray Data Analysis toolを用いてパスウェイ解析を行った。データベース
はWikiPathwaysとNCBI BioSystemsを利用した。次に、有意差を得られたパスウェイにつ
いて以下の実験方法を用いて検証した。
Skt蛋白の細胞内局在
Skt蛋白の細胞内局在を明らかにするため培養細胞でSkt蛋白を強制発現させ蛍光二重免 疫染色を行った。まず、 CAGプロモーターでマウスSktを発現するコンストラクトを持っ たプラスミドをアフリカミドリザル腎細胞由来のBMT10細胞にヌクレオフェクションし
(Nucleofector™, Lonza) 、マウスSkt遺伝子をBMT10細胞内で強制発現させた。これらの細
胞をチェンバースライドで培養し、4%PFAにて室温で10分間固定後、PBTにて10分 浸透化処理を行い10%BSAでブロッキング処理後、一次抗体で常温で60分間反応させ た。続いて、Skt、Calnexineについては蛍光標識二次抗体にて常温で40分間反応させ た。最後にDAPIで1分間反応させ核染色を行い、封入後蛍光顕微鏡にて観察した。使用 した一次抗体と希釈率を以下に示す。アクチンマーカとしてPhalloidin、微小管マーカーと
してTubulin、小胞体マーカーとしてCalnexinを用いた。
rabbit anti-mouse EGFR antibody (Cell Signaling Technology, Inc. Beverly, MA), 1:1,000.
Phalloidin Green Fluorescent Conjugate, Acti-stain (Cytoskeleton, Inc. ), 100nM.
Monoclonal Anti-β-Tubulin−FITC antibody produced in mouse (Sigma–Aldrich Corp.), 1:100.
Purified Mouse Anti-Calnexin anti body (BD Transduction Laboratories), 1:50.
RhoGTP ase 活性測定
直腸肛門形成における細胞骨格調節経路について検証するためこの経路を制御するRho ファミリー分子の活性化測定を行った。RhoファミリーGタンパク質は他の低分子量Gタン パク質と同様に、GTP結合型が活性型であり、GDP結合型が不活性型である。胎生10.5日 胚(n=5)と胎生11.5日胚(n=5)のコントロール [+ SktGt/+ +]、直腸肛門奇形モデル [Sd SktGt/+ SktGt] 尾部を用いて、代表的なRhoファミリー分子であるRhoA、Cdc42の活性化状態 をそれぞれGTP結合型RhoA、GTP結合型Cdc42を G-LIZA Small G-Activation Kit
(Cytoskeleton, Inc)を用いて測定し評価した。胚を凍結破砕後、速やかにLysis Bufferにて
溶解し、各サンプル間で濃度を調整した。lysateをGTPアフィニティープレートに添加し
incubate後、検出用抗体を加えRhoAは蛍光法、Cdc42は比色法をを用いてマルチマイクロ
プレートリーダーで測定した。
②直腸肛門形成関連遺伝子発現解析
次に [Sd SktGt/+ SktGt] の直腸肛門形成に関与する遺伝子を明らかにするため、これまでに
報告されている約20個の直腸肛門形成に関連する遺伝子の中で変化の大きかったHoxa-13 遺伝子、 Hoxd-13遺伝子、Cdx2遺伝子についてReal time PCRによる遺伝子発現量解析とIn
situ hybridization法もしくは免疫染色法による遺伝子発現パターン解析を行った。それぞれ
の実験方法の詳細を以下に示す。
Real time PCR
サンプルは胎生9.5日は全身、胎生10.5日は尾側とした。マイクロアレイと同様にtotal RNAを抽出し、ランダムプライマーを用いて逆転写を行いcDNAを合成した。それぞれの 遺伝子についてTaqmanプローブとプライマー (Hoxa13 ; Mm00433967_m1、Hoxd13 ; Mm00433973_m1、 Cdx2 ; Mm01212280_m1 、Gapdh ; Mm99999915_g1、TaqMan® Gene
Expression Assays) を用いて測定した。内在性コントロールにはハウスキーピング遺伝子
であるGapdhを用いて補正し定量化した。
Whole mount in situ hybridization
<DIG labeled RNA probeの作製>
Cdx2、Hoxa13、Hoxd13についてはデータベースをもとにプライマーを作製し増幅した。
(Cdx2;Forward primer 5‘-aca gca gca gca gca aca ac-3‘、Reverse primer 5‘-tga ctc gaa cag cag caa ac- 3’ 、 Hoxa13; Forward primer 5‘- ctt ccc atg gaa agc tat c -3‘ 、Reverse primer 5‘- ttt ctc ttt gac cct cct g -3‘、 Hoxd13; Forward primer 5‘- tgg gct atg gct acc act tc -3‘ 、Reverse primer 5‘- gat gaa gac tca gtg gag ac -3)脊索の解析の際に用いたShhについてはすでに当教
室にShhcDNAをクローニングしたベクターが存在していたのでそれを用いた。PCRにて増
幅した配列をpGEM®-T Easy Vector(Promega)へ挿入し 大腸菌にて増幅させた後にプラ スミドをQIAGEN Plasmid Maxi Kit(QIAGEN)を用いて抽出、精製した。DIG-labelled RNA
probeはcDNAの5’側のMCSの制限酵素サイトで切断した(37℃、一晩)ベクターを、RNA
Polymerase (T3, T7 or Sp6プロモーター)(Roche Diagnostics)、DIG RNA Labeling Mix(Roche Diagnostics)とともに37℃、2時間incubateして作製した。in vitro transcription後のRNAプ ローブの精製にはマイクロバイオスピンカラム30 Tris RNase free (BioRad)を用いた。作製 したプローブは95℃、3分間denatureした後にな1×TBEバッファーで作製したアガロースゲ ルで100V、15分電気泳動して目的のサイズが得られているかを確認した。
< In situ hybridization>
次に作製したDIG labeled RNA probeを用いてRosenらの方法を一部改変したKokubuらの 方法によりWhole mount in situ hybridizationを行った。[54, 55]
・サンプルの剖出、固定、保存
氷上で胚を採取後、4% PFA中で一晩固定 (4℃) を行った。翌日、PBTで洗浄後、メタ ノールステップ(25%、50%、75%、100% 、 100% それぞれ30分反応)にて脱水し、-80℃
で保存した。
・Hybridization
メタノール中で保存している胚を氷上で7 5 %、5 0 %、2 5 %メタノール/ P B Sで1 0分ずつ incubateして再水和した後、6%過酸化水素/PBSで60分脱色した(氷上)。PBTで洗浄後 (氷 上)、RIPAバッファー(0.05% SDS, 0.15M NaCl, 1% NP40, 0.5% デオキシコール酸, 1mM EDTA, 5mM Tris-HCl pH8.0) で10分 incubate (氷上)し、PBTで洗浄 (氷上)後、4%PFA/0.2%
グルタールアルデヒド/PBTで20分間再固定した。hybe-buffer (50% formamide, 5×SSC, 500µg heparin, 0.02% Tween20) + tRNA (100µg/ml, invitrogen) で68℃、3時間incubateし
prehybridizationを行った後、RNAプローブを80℃, 3分間denatureした後に、hybe-buffer + tRNA (100µg/ml)に1:100になるように添加し(250ng/ml)、68℃で一晩 hybridizationさせた。
・余剰プローブの除去、DIG抗体反応
翌日、hybe-bufferで洗浄(65℃)し、RNase solution (0.5M NaCl, 10mM Tris-HCl pH7.5, 0.1% Tween20) にRNase A(100µg/ml)を加え37℃、60分incubateした。SSC/FA/Tween20 (50%
formamide, 2×SSC, 0.1% Tween20) で5分/2回、10分/3回、30分/5回洗浄(65℃)した後、TBST で洗浄(RT)した後に溶液をMABT(100mMマレイン酸, 150mM NaCl, pH7.5, 0.1% Tween-20) に置換しさらに洗浄(RT)した。次にblocking buffer (MABT, 1.5% blocking reagent [Roche Diagnostics])で1時間 blocking (RT)を行い、並行してDIG抗体(Anti-Digoxigenin-AP, Fab fragments [Roche Diagnostics])を別に用意したblocking bufferに加え(1:2000)、4℃, 1時間 preabsorptionした。続いてblocking bufferをDIG抗体を添加したblocking bufferに置換し、震 盪しながら一晩反応させた(4℃)。
・余剰抗体の洗浄、発色
TBSTで1時間ごとに8回洗浄 (RT)し、その後再びTBSTで震盪しながら4℃で3日間洗浄 後、溶液をNTMT[100mM NaCl, 100mM Tris-HCl (pH 9.5), 50mM MgCl2, 0.1% Tween20]に置 換しincubate (RT)した。NBT/BCIP Stock Solution (Roche Diagnostics)/ NTMT溶液を用いて一 晩発色させた(暗所、4℃)。発色させた胚は4% PFAで30分間再固定した(RT)。Whole mount で観察する際はエタノールステップ(25%、50%、75%、100% 、100% それぞれ30分反応) で脱水し、benzylalcohol/benzylbenzoate (1:2)で透明化した。組織学的解析の際には固定後 のサンプルをパラフィン置換、包埋後、8μm の厚さに薄切し、Nuclear Fast red で対比染色 を行い観察した。
免疫染色
胚を採取後、4%PFAにて一晩固定し、パラフィン置換後、4μmの切片を作製した。脱パ ラフィン後、5分間のマイクロウェーブ処理にて抗原賦活化処理を行い、0.3%過酸化水素/ PBSにて内因性ペルオキシダーゼ活性を阻止後、一次抗体で一晩反応させ、二次抗体はビ オチン化二次抗体を用いたABC法にて行った。対比染色にはHematoxylinを用いた。Cdx2 抗体 ( cell signaling #3977)は1/100希釈にて使用した。
(5) 統計学的解析
全データは平均値 ± 標準誤差 (SE) として表し、有意差はVARA関数で計算し、t検定で 測定した。p < 0.05 の場合に統計学的に有意差があると判定した。
7. 実験結果
1) SktGt 変異を加えたSdマウスの表現型解析
胎生19.5日胚を用いて、各遺伝子型について直腸肛門、腎臓の表現型を組織切片にて解
析した。[+ SktGt/+ SktGt]では全て正常に直腸肛門部は形成され肛門が肛門窩の位置に開口
していた。(n=7) [Sd +/+ +] では 従来の報告と同様に 50% (n=12) で直腸肛門奇形を合併 していたが、ダブル変異マウスでは、 [Sd SktGt/+ +] では80% (n=10)、[Sd SktGt/+ SktGt ]では
100% (n=8)と直腸肛門奇形発生率が加算的に上昇していた。更に、性差による検討は行っ
ていないが、直腸肛門奇形の瘻孔形成の有無について比較すると直腸肛門奇形を合併する 個体中、瘻孔を有するものは [Sd +/+ +] では33%で [Sd SktGt/+ +] では75%、 [Sd SktGt/+
SktGt ] で75% (うち1例は会陰皮膚瘻)でありダブル変異マウスでは瘻孔を形成する表現型が
多い傾向があった。(有意差はなし)腎奇形については、左右ともに腎臓を正常に形成し た個体は [Sd +/+ +] では33%(n=12)であったのに対し、 [Sd SktGt/+ +] では25% (n=10)、 [Sd SktGt/+ SktGt ] では13% (n=8)であり、腎奇形発生率も加算的に上昇していた。(data not shown)
2)直腸肛門奇形モデル [Sd SktGt/+ SktGt ]における直腸肛門発生期の表現型解析
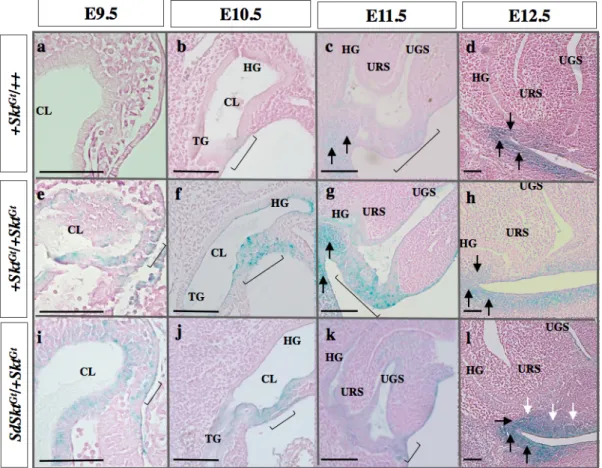
マウス直腸肛門形成期にあたるE9.5、E10.5、E11.5、E12.5、E13.5について総排泄腔の 発生を経時的に観察した。正常の直腸肛門発生([+ +/+ SktGt ] )では胎生9.5日に尿膜管茎 根部でcloacal plateとなる中胚葉を介した内胚葉と外胚葉の近接した部分は確認できたが内 胚葉と外胚葉が直接接している部分は認めなかった。胎生10.5日より中胚葉組織を介さな い内胚葉と外胚葉が接着したcloacal plateの構造が確認された。以後、胎生10.5日から胎生
12.5日にかけcloacal plateは生殖結節の発達にともない背腹軸方向へ伸長し、胎生13.5日に
なると尾側へ成長し下降してきた尿直腸中隔とcloacal plateが接触し、cloacal plateは腹側の 尿生殖膜、背側の肛門膜へと分離され、肛門膜の消失した部分が肛門として開口してい た。直腸肛門奇形モデル [Sd SktGt/+ SktGt ] では、胎生9.5日ではコントロール胚と明らかな 形態学的な違いは認めなかったが、胎生10.5日から11.5日にかけての、cloacal plateの尾側 の組織が欠如し、正常胚と比較してcloacal plateが短縮していた。胎生12.5日になると
cloacal plateを欠如した部分で間葉組織が増殖し、尿直腸中隔のcloacal plateへの接着が妨げ られていた。胎生13.5日になっても尿直腸中隔の下降が適切に起こっていないため、尿生 殖洞と肛門直腸洞との分離は不完全となり、両者を交通する瘻孔が形成されていた(図 3)。
図 3. 直腸肛門発生過程の比較
胎生9.5日から胎生13.5日総排腔発生過程を正中矢状断で示した。コントロールマウス [+ SktGt /+ +] では、
cloacal plateは増殖し尾根部まで伸長している。尿直腸中隔が下降し、腹側の尿生殖洞と背側の直腸肛門洞に
分離される。( a-e ) 胎生13.5日に肛門膜が消失し肛門が開口する。直腸肛門奇形モデル [Sd SktGt/+ SktGt] では
clocal plateの尾側へ伸長していない。尿直腸中隔は下降しているがcloacal plateと接触することなく、尿生殖
洞と直腸肛門洞は完全に分離されていない。 ( f-j ) 黒矢:瘻孔、白矢:増殖している間葉組織、括弧:
cloacal plate. Bars=100µm, HG: hind gut, TG: tail gut, URS: urorectal septum, UGS: urogenital sinus, GT: genital tubercle.
3)直腸肛門周辺のSkt遺伝子の発現パターン解析
胎生9.5日、10.5日、11.5日、12.5日に直腸肛門発生期におけるSkt遺伝子の発現パターン の解析をX-gal染色法にて行った。 [+ SktGt/+ SktGt] 胚は、[+ SktGt/+ +] 胚より強いシグナル を得られたので、 [+ SktGt/+ +] 胚 と同様に直腸肛門部に表現型がないことを確認し、 [+
SktGt/+ SktGt] もコントーロールとして用い、 [+ SktGt/+ +]、[+ SktGt/+ SktGt]、[Sd SktGt/+ SktGt] 胚の3種の遺伝子型について解析を行った。 [+ SktGt/+ +] 胚では、cloacal plateでのβガラク トシダーゼ活性は胎生10.5日以降でしか明確に確認できなかったが、 [+ SktGt/+ SktGt] 胚で は胎生9.5日より、cloacal plateを形成する内胚葉、外胚葉双方でβガラクトシダーゼ活性を 認めた。 [+ SktGt/+ +] 胚では、cloacal plateでのシグナル強度が低かったため、認識できな かったものと考えた。胎生11.5日、12.5日では、コントロール胚のシグナルはcloacal plate だけではなくcloacal plate後方の尾根部腹側の間葉組織でも認められた。一方、 [Sd SktGt/+
SktGt] 胚のβガラクトシダーゼ活性のシグナル発現パターンは、 胎生9.5、10.5日においては コントロール胚と同様に短縮したcloacal plateでのシグナルを認めた。胎生11.5、12.5 日で は、短縮しているcloacal plate尾根部腹側間葉組織のシグナルに加え、直腸肛門の代わりに 増殖している間葉組織でもシグナルを認めた(図4)。内在性の発現と比較するためにI n situ hybridization法と免疫染色を試みたが、cloacal plate周辺でのシグナルは得られなかっ
た。(data not shown) しかし、Skt遺伝子の発現が強い脊索ではいずれもシグナルが得られて
おり、cloacal plateでシグナルが得られなかったのは、cloacal plateでのSkt遺伝発現量が低 いためであり、Xgal染色の結果は内在性の発現を再現しているものと考える。
図 4. 総排腔周囲のXgal染色
胎生9.5日から胎生12.5日の[+ SktGt /+ +] ( a-d )、 [+ SktGt/+ SktGt] ( e-h ) 、 [Sd SktGt/+ SktGt] ( i-l )のXgal染色を 示す。βガラクトシダーゼ活性は [+ SktGt /+ +] では胎生10.5日から12.5日までcloacal plateで確認され、胎生 11.5日からはcloacal plateだけでなくcloacal plate周囲の間葉組織でも確認されるようになる。 [+ SktGt/+ SktGt]
では[+ SktGt /+ +]より強い発現が得られ、胎生9.5日のcloacal plateとなる内胚葉、外胚葉の部分でβガラクトシ
ダーゼ活性が確認できる。 [Sd SktGt/+ SktGt]のβガラクトシダーゼ活性のある組織は [+ SktGt /+ +] )、 [+ SktGt/+
SktGt]とほぼ同じである。胎生12.5日では、後腸の代わりに増殖している間葉組織でのβガラクトシダーゼ活 性が認められる。黒矢:βガラクトシダーゼ活性、白矢:増殖している間葉組織 括弧:cloacal plate Bars=100µm, HG: hind gut, TG: tail gut, URS: urorectal septum, UGS: urogenital sinus.
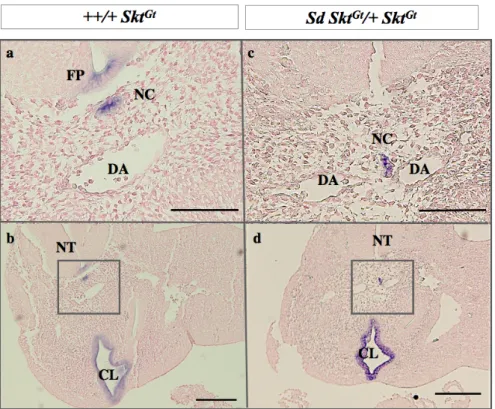
4)脊索と直腸肛門奇形
直腸肛門奇形モデルの脊索の形態について総排泄腔レベルの脊索に注目して解析した。
まず、全体像を把握するためWhole mount で観察を行った。[+ SktGt/+ +] 胚では 神経管腹 側に頭尾則に発達した脊索が観察される。[Sd SktGt/+ SktGt] 胚では、胎生9.5日の尾側の脊 索は正常に発達しているが、胸椎レベルでは脊索の分断化が認められた。胎生10.5日にな ると総排腔背側のレベルでも脊索形成が障害されており、分断化している部分に加え、腹 側に枝状に伸長する脊索が観察された。この異常組織を組織切片にて正中矢状断で観察す ると背側大動脈が一部欠損し、その欠損部を通り前方へ枝状に伸びる脊索組織が観察され た(図5)。脊索で発現する遺伝子としてよく知られてるShhの発現の有無を確認したとこ ろ、異常をきたした脊索においてもShhの発現は保たれていた。また、横断像で同部位を 観察したところ、血管の欠損部は背側大動脈の融合が障害された正中部分であった(図 5)。
図5. 総排腔周囲の脊索形成
Xgal染色により、胎生9. 5日から胎生12.5日の whole mount で観察した脊索の形態を [+ SktGt /+ +] ( a-c )、 [Sd SktGt/+ SktGt] ( d-f ) に示し、組織切片で観察した胎生10.5日の脊索の形態を、 [+ SktGt /+ +] ( i )、 [Sd SktGt/ + SktGt] ( j, k )に示した(白四角)。βガラクトシダーゼ活性は [+ SktGt /+ +] では胎生10.5日から12.5日まで神 経管腹側に伸びる脊索が確認される。 [Sd SktGt/+ SktGt] では胎生9.5日の尾部の脊索形成は[+ SktGt /+ +]と相違 ないが、胎生10.5日以降より尾部においても脊索が狭小化していた。腹側に枝状に伸長する脊索もみられ る。脊索が腹側に伸長する場所では背側大動脈の形成も障害されている(黒矢)。 ( j ) Bars=100µm, NC:
notochord, TG: tail gut, CL:cloaca, NT: neural tube, DA: dorsal aorta
図6. 脊索でのShhの発現
In situ hybridization法により、胎生10.5日胚脊索のShhの発現を横断面にて解析した。(a), (b) は [+ SktGt /+ +]
(c), (d)は [Sd SktGt/+ SktGt] を示す。(a), (c) は(b), (d) のそれぞれ拡大像を示す(黒四角)。 [Sd SktGt/+ SktGt] の 異常な形態を示す脊索でもShhは発現していた。神経底板が誘導されておらず、背側大動脈の融合が障害され ていた。Bars=100µm, NC: notochord, CL:cloaca, NT: neural tube, DA: dorsal aorta, FP: floor plate
5)遺伝子発現プロファイリング解析
①シグナルパスウェイ解析
マイクロアレイで得られた全マウス36227遺伝子の解析結果より、発現強度比 [Sd SktGt/+
SktGt] / [+ SktGt/+ +]が2.0以上の遺伝子もしくは、0.5以下であった遺伝子を抽出したところ 1291遺伝子が抽出された。これらの遺伝子を用いてパスウェイ解析を行ったところ9つの シグナルパスウェイで有意差が得られた(表5)。いずれも過去の報告で直腸肛門発生と の関連は指摘されていないシグナル経路であり、各々の変動していた遺伝子についてもこ れまで直腸肛門発生との関連は示唆されていない。我々は以前、ヒトSkt遺伝子の解析で、
S k t蛋白が細胞骨格の一つである微小管と共局在するという結果を得ている。そこで、
WikiPathways (Regulation of actin cytoskeleton)とBioSystems (G13 signaling pathway) の双方の 情報システムにおいて有意差を得られた細胞骨格調節経路に注目した。まずマウスS k t蛋 白の細胞内局在を確認後、細胞骨格調節経路を制御するRhoファミリーGタンパク質活性 について解析を行った。
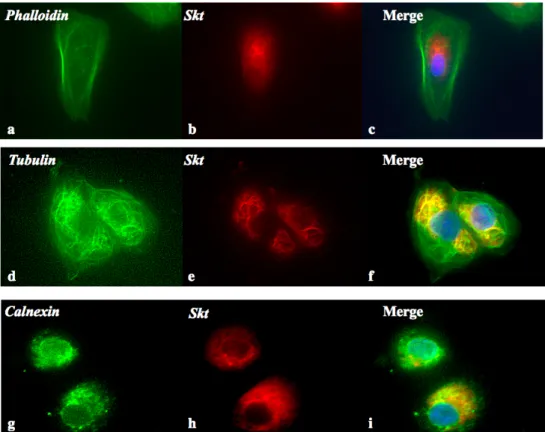
(a) マウスSkt蛋白細胞内局在
マウスSkt遺伝子を強制発現させたBMT10細胞で、細胞内小器官マーカーとマウスSkt蛋 白との蛍光免疫二重染色を行いSkt蛋白の局在を解析した。マウスSkt抗体のシグナルは、
核周辺でシグナルを強く認めた。そして、Tubulin抗体で得られたシグナルと一致してシグ ナルを認めており、Skt蛋白は微小管と共局在していた(図7)。その他の、Phalloidin、
Calnexin抗体とは一致するシグナルは得られなかった。
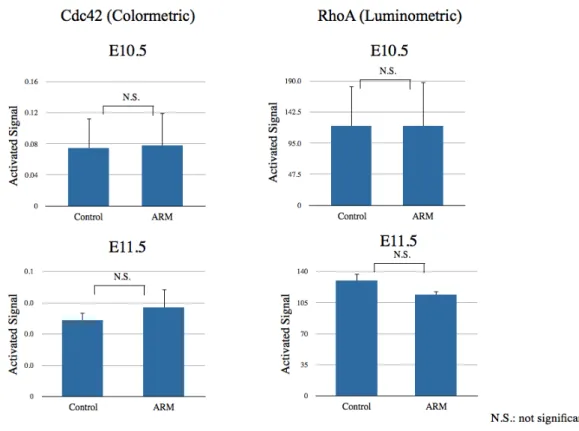
(b) RhoファミリーGタンパク質活性
胎生10.5日胚(n=5)と胎生11.5日胚(n=5)を用いてRhoファミリーGタンパク質である
RhoAとCdc42の活性化状態を評価した。胎生11.5日で直腸肛門奇形モデルにてRhoAの活性
が低下し、Cdc42活性が上昇している傾向はあったが、胎生10.5日、胎生11.5日のいずれに
おいてもRhoA、Cdc42の活性化状態の差は得られなかった(図8)。
表5. 有意差を認めたシグナル経路
Pathway name Changed
genes
Total genes
Z score ( >0)
P-value (≦0.05)
Oocyte meiosis(BioSystems) 13 104 3.106 0.00947
Regulation of actin cytoskeleton(WikiPathways) 16 139 2.952 0.00978
Ovarian infertility genes(WikiPathways) 6 30 3.378 0.01136
G13 signaling pathway(WikiPathways) 6 35 2.92 0.02099
Beta oxidation meta pathway(WikiPathways) 5 27 2.873 0.02647
Axon guidance(BioSystems) 13 123 2.442 0.03032
B cell receptor signaling pathway(WikiPathways) 15 146 2.382 0.03087 T cell receptor signaling pathway(BioSystems) 12 108 2.535 0.03376
FcγR-mediated phagocytosis(BioSystems) 10 90 2.314 0.03993
図7. Skt蛋白の細胞内局在
マウスSkt遺伝子をBMT10細胞に強制発現させ、細胞を固定後、二重染色を行い細胞内小器官(緑)(a, d,
g)とSkt蛋白(赤)(b, e, h) を検出した。DAPI(青)は核を示す。それぞれの右段に重ね合わせた像を示し た。Skt蛋白は微小管と核周辺で共局在していた。
図8. RhoGTPase活性化測定
胎生10.5日 (n=5)、11.5日胚 (n=5)の尾側を用いてCdc42、RhoAの活性化状態を評価した。Cdc42は比色 法、RhoAは発光法を用いて定量化した。いずれも有意差は得られなかった。
②直腸肛門形成関連遺伝子発現解析
これまでに報告されている直腸肛門形成に関連する遺伝子のマイクロアレイの結果を表 6に示した。これらの直腸肛門形成関連遺伝子の中で、直腸肛門奇形モデルで最も増加し ていた遺伝子は、Hoxa-13 遺伝子(モデル/コントロール ; 2.30613)、 Hoxd-13遺伝子(モ デル/コントロール ; 1.718743)であり、最も減少している遺伝子はCdx2遺伝子(モデル/コ ントロール ; 0.658666)であった。
この3つの遺伝子についてまず、発現量の差について胎生9.5、10.5日胚を用いて定量PCR にて解析したところ、胎生10.5日のHoxd-13遺伝子の発現は有意差を認めて上昇していた。
しかし、その他H o x a- 1 3、C d x 2遺伝子については発現量の差において有意差を得られな かった(図9)。