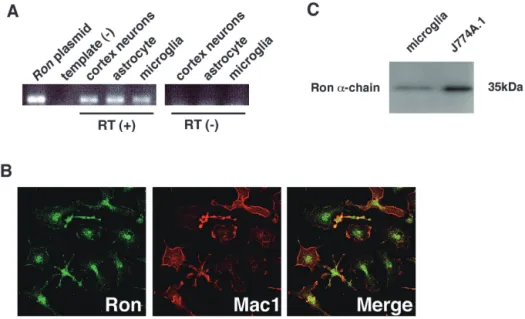
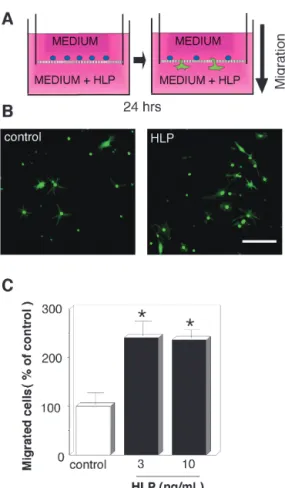
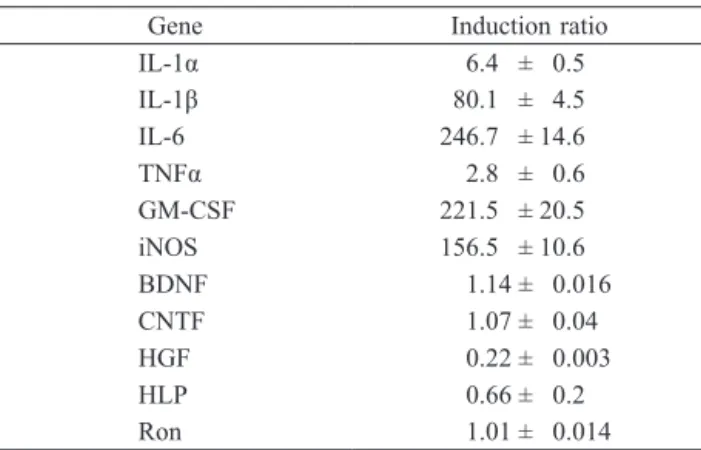
In this, the first ever in-depth study of the econometric practice of nonaca- demic economists, I analyse the way economists in business and government currently approach
He thereby extended his method to the investigation of boundary value problems of couple-stress elasticity, thermoelasticity and other generalized models of an elastic
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
This article concerns the behaviour of solutions to a coupled sys- tem of Schr¨ odinger equations that has applications in many physical problems, especially in nonlinear optics..
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
This paper presents an investigation into the mechanics of this specific problem and develops an analytical approach that accounts for the effects of geometrical and material data on
The proof of the existence theorem is based on the method of successive approximations, in which an iteration scheme, based on solving a linearized version of the equations, is
discrete ill-posed problems, Krylov projection methods, Tikhonov regularization, Lanczos bidiago- nalization, nonsymmetric Lanczos process, Arnoldi algorithm, discrepancy