持久的走行トレーニング初期段階における骨格筋の脂質代謝関連酵素活性の変化過程
7
0
0
全文
(2) 冬季スポーツ研究 第 7 巻 1 号 9-14, 2004 (北海道教育大学冬季スポーツ教育研究センター紀要). 持久的走行トレーニング初期段階における骨格筋の脂質代謝関連酵素活性の 変化過程 鈴木淳一 1)、平塚勇介 2)、東浦拓郎 3) 北海道教育大学冬季スポーツ教育研究センター1)、北海道教育大学教育学研究科 2)、筑波大 学大学院博士課程人間総合科学研究科 3) Time-course changes in citrate synthase and 3-hydroxyacyl-CoA dehydrogenase activities in soleus muscle of young treadmill-trained rats Research and Education Center for Winter Sports1), Graduate School of Education2) Hokkaido University of Education, 5-3 Ainosato, Kita-ku, Sapporo, Hokkido 002-8502, Japan. Graduate School of Comprehensive Human Sciences, University of Tsukuba, Tennodai, Ibaraki 305-8577, Japan Abstract This study was designed to examine the time-course changes in citrate synthase (CS) and 3hydroxyacyl-CoA dehydrogenase (HAD) activities during endurance training by running in young female Wistar rats. Exercise training by running started at the age of 5 weeks and lasted for 5 weeks at 25 m/min on a 25% grade, 10-60 min/day. Enzyme activities were observed in the soleus muscle. The soleus muscle weight and its weight-to-body weight ratio were significantly greater in the training group than in the sedentary control group at Week 5 and Weeks 3 and 5, respectively. Training significantly increased the HAD activity by 30%, 29% and 37%, respectively, at Weeks 3, 4, 5. The CS activity was significantly increased in training groups by 35%, 47% and 47%, respectively, at Weeks 3, 4, 5. The HAD-to-CS ratio (HAD/CS) remained unchanged throughout the experimental period. These results suggest that it takes at least 3 weeks of intensive training to cause adaptive changes in fatty acid utilization in slow twitch muscle. Similar time-course changes in CS and HAD may contribute to effective fatty acid utilization in muscle cells. Key words: citrate synthase; endurance training; 3-hydroxyacyl-CoA dehydrogenase; skeletal muscle. 緒言 有酸素運動時では、エネルギー基質として 主に血中グルコース、筋グリコーゲン及び脂肪 酸が利用される。近年、その優れた効率性や有 用性の高さから、脂質代謝能力が持久力の指標 としてが注目されている。ヒトの研究では、脂 肪酸のエネルギー貢献率は低強度運動時では約 90 %、中等度運動時では約 65 %であること が知られている。また、長期間の持久的トレー ニング後に、エネルギー基質として脂肪酸の動 員が増大することが知られている [3,6,9-11]。 脂肪酸のβ酸化はミトコンドリア内で行われ、 その後有酸素的に ATP が生成される。この脂. 肪酸β酸化系の律速酵素である 3-ヒドロキシア シル-CoA 脱水素酵素(HAD)の活性は、長期 間の持久的トレーニング後に増大することがこ れまで多くの研究で報告されている [3,6,9-11]。 しかし、トレーニング初期段階における HAD 活性の変化過程に関してはほとんど研究 されていない。Cheng ら[4]は、Long-Evans 系雄 ラットを用いて高脂肪食摂取と自発走運動を 6 週間行い、ヒラメ筋の HAD 活性の変化過程を 観察した。その結果、高脂肪食摂取群では HAD 活性が漸増したが、通常食摂取群では HAD 活 性にほとんど変化がみられなかったと報告して いる。Henriksson ら [7]は、New Zealand White. 9.
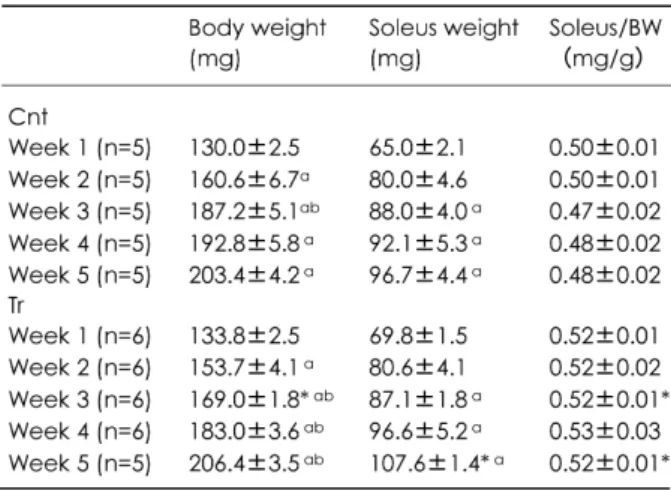
(3) 鈴木淳一、平塚勇介、東浦拓郎. Rabbit を用いて速筋である前脛骨筋に 10Hz、 24h/day の条件で電気刺激を 10 週間行い、HAD 活性の変化過程を観察した。電気刺激開始 1、 2、3、5、8、10 週後、それぞれ HAD 活性の値 を測定した結果、いずれも 0 週時(電気刺激開 始前)より高い値を示し、3 週後が最高値であ ったと報告している。 しかしながら、高強度の持久的走行トレー ニングを行うと、その初期段階でどのように脂 質代謝能力が改善されていくのか、その変化過 程を観察した研究はこれまで報告されていない。 そこで本研究では、ラットに高強度の持久 的走行トレーニングを 5 週間負荷し、1 週間毎 の骨格筋 HAD 活性の変化過程について明らか にすることを目的とした。 実験方法 1.実験動物 実験には 3 週齢の Wistar 系雌ラット 54 匹 を用いた。2 週間飼育室の環境に慣らした後、 5 週齢時に安静コントロール(Cnt)群 25 匹、 トレーニング(Tr)群 29 匹の 2 グループに分 けた。すべてのラットは室温 24±1 ℃、相対 湿度約 50 %、12 時間の明暗周期の環境下で飼 育し、飼料(日本クレア/CE-2)と水道水を共 に自由摂取させた。 2.運動負荷 Tr 群のラットには、ラット用トレッドミ ル(KN-73/夏目製作所)による持久的走運動 を負荷した。トレーニングは 5 週齢時に 25 m/min、10 min、勾配 25 %の条件で開始し、60 min になるまで 3 min/day の割合で運動時間を 漸増させ、週 5 日の頻度で 5 週間負荷した。 3.筋標本 トレーニング開始 1、2、3、4、5 週後、そ れぞれペントバルビタール麻酔下(50 mg/kg) のラットからヒラメ筋を摘出し、湿重量を秤量 した。筋のサンプルは、液体窒素で瞬間凍結後、 −80 ℃で冷凍保存した。 4.酵素溶液の調製 筋のサンプルは−25 ℃下で微粉状にし、5 mM β-mercaptoethanol、0.5 mM EDTA、0.02 % bovine serum albumin(BSA)を含む 20 mM phos-. 10. phate buffer(pH7.4)を用い、ポリトロン型ホ モジナイザーで氷冷下でホモジネート(15,000 rpm、15 sec×2)した。そして、0 ℃、13,000 g で 15 分間遠心分離した後、その上澄み液を酵 素溶液として、酵素活性の測定まで−80 ℃で 冷凍保存した。 5.酵素活性の測定 3-ヒドロキシアシル-CoA脱水素酵素 (HAD, EC 1.1.1.35)活性の測定には Bass ら[1]の方法 を用いた。酵素溶液に、5 mM EDTA、0.45 mM NADH を含む 100 mM triethanolamine-HCl buffer (pH 7.0)を加え、25 ℃の温水中に 10 分間静 置し温度平衡に達した後、0.1 mM acetoacetylCoA を添加して反応を開始させた。反応開始後 5 分間にわたり分光光度計を用いて 340 nm に て吸光度の変化を測定した。 Citrate Synthase(CS, EC 4.1.3.7)活性の測 定には Srere [13]の方法を用いた。酵素溶液に 1 mM DTNB、10 mM acetyl-CoA を含む 0.1 M Tris-HCl buffer(pH8.1)を加え、25 ℃の温水 中に 10 分間静置し温度平衡に達した後、10 mM oxaloacetate を添加して反応を開始させた。反 応開始後 5 分間にわたり分光光度計を用いて 412 nm にて吸光度の変化を測定した。 6.統計処理 すべての値は平均値±標準誤差で示した。 すべてのデータはまず、Kormogorov-Smirnov 検 定によって正規性の検定を行ない、正規分布し ていることを確認した。2 群間の比較にはStudent の t 検定を用いた。また、各群の経時変化 の 検 定 に は 一 元 配 置 分 散 分 析 及 び Fisher の post-hoc テストを用いた。 検定結果は危険率 5 % 未満を有意水準とした。 実験結果 表 1 にトレーニング期間中の体重、ヒラメ 筋重量及びヒラメ筋重量/体重比の値を示した。 体重は、第 3 週において Tr 群の値が Cnt 群よ り有意に低い値を示したが(P<0.05)、他の週 では変化がみられなかった。経時変化では、両 群ともに週毎に増加が認められた。ヒラメ筋重 量は、第 5 週において Tr 群の値が Cnt 群より 有意に高い値を示した(P<0.05)。経時変化で.
(4) トレーニングによる HAD 活性の変化過程. 高い値を示した(P<0.05) 。 考察. は、両群ともに漸増していた。ヒラメ筋重量/ 体重比は、第 3、5 週において TR 群は Cnt 群 より有意に高い値を示した(P<0.05)。経時変 化では、両群ともに 5 週間通してほぼ一定の値 を示した。 図 1 に、HAD 活性(U/g tissue)の変化過 程を示した。HAD 活性は、第 3、第 4、第 5 週 において、 Tr 群が Cnt 群よりもそれぞれ約 30 %、 29 %、37 %有意に高い値を示した(P<0.05) 。 TR 群の経時変化では、第 1 週に対して第 3 週 以降有意に高い値を示した(P<0.05)。Cnt 群で は、第 1 週から第 5 週まで緩やかに漸増したが、 有意差は認められなかった。 CS 活性は Tr 群が Cnt 群より第 3 週で約 35 %、第 4 週で 47 %、第 5 週で 47 %有意に 高い値を示した(P<0.05;図 2)。Tr 群の経時 変化では、第 1 週に対して第 3、4、5 週、第 2 週に対して第 3 週が有意に高い値を示した (P<0.05)。Cnt 群では、5 週間通して変化がみ られなかった。 図 3 に HAD/CS 比の変化過程を示した。 HAD/CS 比は脂肪酸のエネルギー貢献率を示す ものである。すべての週において、両群間に有 意差は認められなかった。Tr 群の経時変化では、 5 週間通してほぼ一定の値を示した。Cnt 群で は、第 1 週に対して第 2、3、4、5 週が有意に. 本研究では、5 週齢の Wistar 系雌ラットを 用いて、5 週間の持久的走行トレーニング期間 中、ヒラメ筋における HAD 活性がどのように 変化していくのか、その適応過程を観察した。 トレーニング期間中、HAD 活性は第 3 週 目以降に有意に増加していた(P<0.05;図 1) 。 これは、第 3 週目以降に脂肪酸のβ酸化が亢進 され、脂質からのエネルギー供給量が増大した ことを示唆している。エネルギー基質として脂 肪酸の利用が増大すると、筋や肝臓のグリコー ゲンが節約されるものと考えられている [8,11,12] 。Geor ら[5] は、持久的走行トレーニ ングを負荷した馬に高強度の持久的走運動を実 施させたところ、筋グリコーゲン代謝率が減少 し、トレーニング開始前よりも運動終了時の血 中や筋中乳酸濃度が減少することを観察した。 その結果、VO2max が増加し、疲労困憊に至る までの時間が延長したことを報告している。こ. 11.
(5) 鈴木淳一、平塚勇介、東浦拓郎. の時間が延長したことを報告している。これら のことから、有酸素的運動能力が向上する際に、 脂質代謝系の適応が密接に関与しているものと 考えられる。 本研究において、Tr 群の CS 活性は、HAD 活性と同様に第 3 週目以降に有意に増加してい た(P<0.05;図 2) 。この結果は、クエン酸回路 の活発化と脂肪酸のβ酸化亢進が同時に起こっ たことを示唆している。β酸化によって生成さ れたアセチル-CoA は、クエン酸回路に入り、 その後有酸素的にエネルギーが産生される。β 酸化だけが過剰に進行すると、アセチル-CoA の蓄積が生じ、細胞質からミトコンドリア内へ の脂肪酸の輸送が抑制されることから、脂質代 謝の亢進には、クエン酸回路の活性化が不可欠 であると考えられる。また、多くの先行研究で、 持久的トレーニング後、HAD 活性と CS 活性が 共に有意に増加していたことが観察されている [3,6,9-11]。. 12. Suzuki ら[14]は、持久的走行トレーニン グを行うと、組織の酸素需要増大に対する適応 として、まず毛細血管密度や C:F 比(筋線維 1 本当たりの毛細血管数)の増加が起こり、そ の後、酸化酵素活性(コハク酸脱水素酵素)が 増加すると報告している。本研究においても、 持久的走行トレーニングによって、第 1、第 2 週目に微小循環系の適応が起こり、その後第 3 週目以降に酸化酵素活性(HAD、CS)が増加 したものと推察される。このことから、脂質代 謝系の適応は、微小循環系の改善後に起こるも のと考えられる。運動時には、血中から取り込 まれた脂肪酸が内因性のものよりもより酸化さ れることから、毛細血管網の増加が脂質代謝の 亢進に深く関与していると考えられる。 Tremblay [15]らは、ヒトを対象に、持続的 走行トレーニングを負荷した群と高強度の間欠 的走行トレーニングを負荷した群に分け、トレ ーニングが脂質代謝系に及ぼす効果を比較した ところ、持続走群の運動中の総エネルギー消費 量は間欠走群の約 2 倍であったにも関わらず、 間欠走群の方が HAD 活性が増加し、皮下脂肪 が顕著に減少していたことを観察した。この結.
(6) トレーニングによる HAD 活性の変化過程. 果から、持久的トレーニングでは、運動強度が 高いほど脂質代謝系の適応を促進すると報告し ている。5 週齢の Wistar 系雄ラットに約 60 % VO2max 強度の持久的走行トレーニングを 6 週 間負荷した先行研究では、ラスト 2 週間の運動 時間は 120 分にも及んだが、トレーニング後、 HAD 活性の有意な増加は観察されなかった[16]。 本研究では、第 1 週から第 3 週までの 1 日当た りの運動時間は 10∼52 分であった。また、本 研究とほぼ同週齢で同系統のラットを用いた Brooks ら[2]の研究から、本研究の運動強度を 推測すると、最大酸素摂取量の約 82 %以上、 最大心拍数の約 90 %以上になるものと考えら れる。これらのことから、運動時間は短くても 高い運動強度であれば脂質代謝系の適応を引き 起こすと考えられる。 本研究の安静対象群では、有意差は認めら れなかったが、5 週間の実験期間中 HAD 活性 は緩やかに漸増していた(図 1) 。しかし、CS 活 性にはほとんど変化はみられなかった(図 2) 。 また、HAD/CS 比は第2週目以降、第 1 週目に 対して有意に高い値を示していた(P<0.05;図 3)。これらの結果は、通常の成長過程では、有 酸素的エネルギー代謝量はほとんど変化しない が、脂質のエネルギー貢献率は成長とともに増 大することを示唆するものである。 本研究と同週齢で同系統のラットを用いた 先行研究で、持久的トレーニング後、type IIa fiber の横断面積が増大することが観察されてい る[14]。この研究では、本研究と同じ運動強度 のトレーニングを負荷したことから、高強度の 持久的トレーニングが type IIa fiber の肥大を促 進したものと考えられる。本研究では、Tr 群の ヒラメ筋重量が第 5 週目に有意に増加していた。 (P<0.05;表 1)また、ヒラメ筋重量/体重比も 第 3、第 5 週目に有意に増加していた(P<0.05; 表 1)。この結果は、本研究の持久的走行トレ ーニングによって、type IIa fiber 組成が約 20 % であるヒラメ筋が肥大したことを示しており、 上述した報告を支持するものと考えられる。一 方、安静対象群では、5 週間通してヒラメ筋重 量は緩やかに漸増していたが、ヒラメ筋重量/ 体重比には変化がみられなかった(表 1)。こ. の結果から、対象群のヒラメ筋重量の増加は、 成長過程に伴う体重の増加に比例して起こった ものと考えられる。 本研究では、持久的走行トレーニングの初 期段階において、HAD 活性がどのように変化 するのか、その適応過程を明らかにすることを 目的とした。その結果、第 3 週目以降に HAD 活性が有意に増加したことを観察した。このこ とから、本研究で用いた高強度の持久的トレー ニングでは、第 3 週目以降に脂質からのエネル ギー供給量が増大することが示唆された。また、 β酸化の亢進とクエン酸回路の活発化が同時に 観察され、これはエネルギー基質としてより多 くの脂肪酸を動員するために不可欠な適応性変 化と考えられる。 参考文献 1. Bass A, Brdiczka D, Eyer P, Hofer S & Pette D. 1969. Metabolic differentiation of distinct muscle types at the level of enzymatic organization. Europian J Biochem, 10, 198-206. 2. Brooks GA & White TP. 1978. Determination of metabolic and heart rate responses of rats to treadmill exercise. J Appl Physiol. , 45(6), 1009-1015. 3. Carter SL, Rennie CD, Hamilton SJ & Tarnopolsky. 2001. Changes in skeletal muscle in males and females following endurance training. Can J Physiol Pharmacol, 79(5), 386-392. 4. Cheng B, Karamizrak O, Noakes TD, Dennis SC & Lambert EV. 1997. Time course of the effects of a high-fat diet and voluntary exercise on muscle enzyme activity in long-evans rats. Physiology & Behavior, 61(5), 701-705. 5. Geor RJ, McCutcheon LJ & Shen H. 1999. Muscular and metabolic responses to moderateintensity short-term training. Equine Vet Suppl, 30, 311-317. 6. Grinton S, Powers SK, Lawler J, Criswell D, Dodd S & Edwards W. 1992. Endurance training-induced increases in expiratory muscle oxidative capacity. Med Sci Sports Exerc, 24(5), 551-555. 7. Henriksson J, Chi MMY, Hintz CS, Young DA, Kenneth KK, Salmons S & Lowry OH. 1986. Chronic stimulation of mammalian muscle:. 13.
(7) 鈴木淳一、平塚勇介、東浦拓郎. changes inenzymes of six metabolic pathways. Am.J.Physiol, 251(Cell Physiol.20), C614C632. 8. Hurley BF, Nemeth PM, Martin WH, Hagberg JM, Dalsky GP & Holloszy JO. 1986. Muscle triglyceride utilization during exercise:effect of training. J Appl Physiol, 60(2), 562-567. 9. Powers SK, Lawler J, Criswell D, Lieu FK & Martin D. 1992. Aging and respiratory muscle metabolic plasticity: effects of endurance training. J Appl Physiol, 72(3), 1068-1073. 10. Schantz P, Henriksson J & Jansson E. 1983. Adaptation of human skeletal muscle to endurance training of long duration. Clin Physiol, 3(2), 141-151. 11. Serrano AL, Quiroz-Rothe E, Rivero JL. 2000. Early and long-term changes of equine skeletal muscle in response to endurance training and detraining. Pflugers Arch, 441(2-3), 263-274 12. Simi B, Sempore B, Mayet MH, Favier RJ. 1991. Additive effects of training and high-fat diet on energy metabolism during exercise. J Appl Physiol, 71(1), 197-203. 13. Srere PA. 1969. Citrate synthase. Methods in Enzymology, 13, 3-11. 14. Suzuki J, Kobayashi T, Uruma T & Koyama T. 2001. Time-course changes in arteriolar and venular portions lf capillary in young treadmilltrained rats. Acta Physiol Scand, 171, 77-86. 15. Tremblay A, Simoneau JA & Bouchard C. 1994. Impact of exercise intensity on body muscle metabolism. Metabolism, 43(7), 814-818. 16. Zonderland ML, Bar PR & Reijneveld JC, Spruijt BM, Keizer HA, Glatz JF. 1999. Different metabolic adaptation of heart and skeletal muscles to moderate-intensity treadmill training in the rat. Eur J Appl Physiol Occup Physiol, 79(5), 391-396.. 14.
(8)
図
関連したドキュメント
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
French case system has a case called tonic in addition to nominative, accusative and dative, and all French nominal SFs appear in tonic forms, regardless of what case their
The purpose of the Graduate School of Humanities program in Japanese Humanities is to help students acquire expertise in the field of humanities, including sufficient
Amount of Remuneration, etc. The Company does not pay to Directors who concurrently serve as Executive Officer the remuneration paid to Directors. Therefore, “Number of Persons”
いられる。ボディメカニクスとは、人間の骨格や
一方、Fig.4には、下腿部前面及び後面におけ る筋厚の変化を各年齢でプロットした。下腿部で は、前面及び後面ともに中学生期における変化が Fig.3 Longitudinal changes
小国町 飛び込み型 一次産業型 ひっそり型 現在登録居住者。将来再度移住者と して他地域へ移住する可能性あり TH 17.〈Q 氏〉 福岡→米国→小国町
