Cerebral circulatory response coupling with visual stimulation during exercise [ PDF
4
0
0
全文
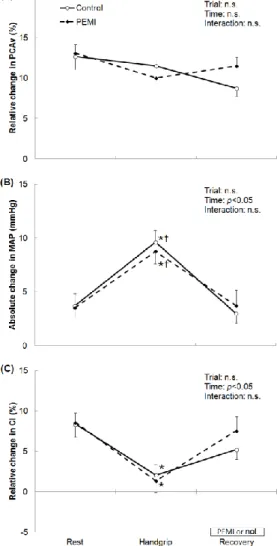
(2) 平均動脈圧(MAP, mean arterial. 静値を保ったが,視覚刺激に伴う昇圧応答が増加し,一. pressure)および PCAv を連続測定した.血流の流れ. 方で CI の増加は抑制された.以上のことから,視覚刺. やすさを示す脳血管コンダクタンス(CI, conductance. 激に伴う NVC は HG 中には保たれるものの,その調節. index)は,PCAv を MAP で除すことにより算出した.. 機序は安静時とは異なることが示唆された.. ⅲ)測定項目:. NVC の評価として,閉眼 20 秒間における PCAv の平. PEMI により HG 後も昇圧応答が継続したにもかかわ. 均血流速度に対する視覚刺激 40 秒間における最大応答. らず,PCAv は安静値に低下し,NVC も一定であった.. の相対変化を算出した.. PCAv の増加は筋代謝受容器反射の影響を受けないこ. ⅳ)統計解析: すべてのデータは平均値±標準誤差で. とが推察された.. 示した.経時変化に伴う差の検定には繰り返しのある 2 要 因分 散分析 を用い , 主 効果が 得られ た場 合 に は Bonferroni の検定を行った.ベースライン値との差の 検定には対応のある t 検定を用いた.有意水準はすべ て危険率 5%未満とした.なお,HG および回復 2 分間. 3.動的運動の強度が視覚刺激に伴う脳の血流応答に 及ぼす影響(実験②) 1)実験の背景および目的: 中等度の有酸素運動中に PCAv は増加し,NVC は保 14).一方,動的運動の強. においては,運動による血圧変動の影響を取り除くた. たれることが報告されている. め,それぞれ後半 1 分間のデータを解析対象とした.. 度が PCA 血流に及ぼす影響は不明である.脳血流の全 体量は疲労困憊に近い動的運動中には安静値程度まで. 3)結果:. 減少する. 16).この減少は,高強度運動中の過換気によ. HG により PCAv は安静時から有意に増加した.この. る動脈血二酸化炭素分圧( PaCO2 ,arterial partial. 際,MAP は増加し,CI は低下した.PEMI 中に PCAv. pressure of CO2)の低下が脳血管収縮を引き起こした. は安静値に低下したが,CI は安静値に回復しなかった. ことに起因すると推察されている.また,安静時にも. (図 1).NVC は HG および PEMI 中も安静値と差が. PCAv は PaCO2 低下による脳血管収縮により減少し,. なかった.安静時と比較して,HG により視覚刺激に伴 う MAP の応答は有意に増加し,CI の応答は有意に低 下した(図 2) . 4)考察: HG 中,CI が低下したにもかかわらず,PCAv は増加 した.したがって,PCAv の増加は血管拡張ではなく主 に昇圧によることが示唆された.HG 中にも NVC は安. 図 1. HG に伴う(A) PCAv および(B) CI の安静値からの相対変化. 図 2. 視覚刺激に伴う(A) PCAv, (B) MAP および(C) CI の変化.
(3) NVC は減少する 17).したがって,高強度の動的運動中. 評価として,閉眼 20 秒間における PCAv の平均血流速. には,過換気による PaCO2 の低下が脳血管収縮を介し. 度に対する視覚刺激 40 秒間における最大応答の相対変. て PCAv を減少させ,さらにベースラインの PCAv の. 化を算出した.換気量および PETCO2 から PaCO2 を推. 減少により NVC が減少するという仮説を立てた.. 定した 18).. 一方,視覚野の代謝要求は視覚処理の複雑さに影響さ. ⅳ)統計解析: 実験①と同様の解析を行った.. れることから,PCA の血流増加の程度は視覚刺激の複 雑さを反映する 4).したがって,高強度の動的運動によ. 3)結果:. り NVC が減少するならば,運動強度の増加に伴い PCA. PCAv および MAP はすべての強度の運動により安静. の血流増加の程度に対する視覚刺激の種類の影響は阻. 時から有意に増加した.PCAv は中等度運動により,コ. 害されるかもしれない.. ントロール条件よりも有意に増加した.CI は高強度運. そこで,異なる強度における動的運動中の PCAv を測. 動により,安静時から有意に低下した(図 3).すべて. 定し,NVC を評価した.また,視覚処理の複雑さを反. の条件において,PCAv はすべての視覚刺激により閉眼. 映する PCA の血流増加の程度に及ぼす運動強度の影響. 時から有意に増加し,PCA の血流増加の程度はチェッ. を検討した.. クボード,開眼およびフラッシュ光の順で大きく,その 順序に変動はなかった.したがって,NVC に対する運. 2)方法: ⅰ)被験者: 健常男性 14 名(年齢 24±1 歳)とした. 実験①と同様の手続きで実験を行った.. 動強度の影響は認められなかった(図 4) . 外耳温は高強度運動条件でのみ上昇し,PaCO2 は低強 度運動条件でのみ増加した.. ⅱ)実験手順: 実験はすべて 22°C に保たれた暗室に て行った.2 分間の安静時測定後,自転車エルゴメータ. 4)考察:. ー運動を 15 分間行わせた.心拍数 120・140・160 拍/. 動的運動中,PCAv の増加は CI の増加を伴わなかっ. 分に相当する 3 種類の運動強度を用いた(低強度,中. たことから,PCAv は主に血管拡張ではなく昇圧により. 等度,高強度と定義した).運動開始 3 分後から 12 分. 増加したことが示唆された.NVC は動的運動中にも安. 間,開眼,反転チェックボードおよびフラッシュ光の 3. 静値を保ち,それは主に視覚刺激に伴う昇圧応答による. 種類の視覚刺激を与えた.コントロール試行として,. ものであることが示唆された.また,異なる視覚刺激に. 安静を維持させた.. より誘発された PCA の血流増加の順序も安静時と変わ. ⅲ)測定項目: MAP および PCAv を連続測定した.. らず一定を保った.PCAv は運動時においても視覚野の. PCAv と MAP より CI を算出した.運動前後に外耳温. 代謝要求を反映し, 視覚処理の複雑さに応じて増加する. および呼気終末二酸化炭素分圧( PETCO2, end-tidal. ことが推察された.なお,本研究で用いた高強度運動条. CO2 partial pressure)をそれぞれ測定した.NVC の. 件において PCAv の減少が観察されなかったことは,. 図 3. 動的運動に伴う(A) PCAv および(B) CI の相対変化. 図 4. 視覚刺激に伴う(A)PCAv および(B)CI の変化.
(4) PaCO2 が低下しなかったことに起因すると推察される.. 6) Woods. visual function. Ophthalmic Physiol. Opt. 15:5–12.. 以上のことから,PCAv は動的運動により増加するに もかかわらず,視覚刺激に伴う NVC は動的運動中に保. & Thomson. (1995) Effects of exercise on aspects of. 7). Ishigaki. (1989) The influence of an exercise on the visual function; on the stricture of the visual field during exrcise.. たれることが示唆された.. Jpn. J. Phys. Educ. 34:245–253.. 4.まとめ. 8). PCA の血流は静的および動的運動による昇圧に伴い. cerebral arteries during the cold pressor test and handgrip. 増加した.こうしたベースラインの PCA 血流の増加に もかかわらず,NVC に伴う血流の増加は運動の影響を. exercise. Eur. J. Appl. Physiol. 112:641-646. 9). 運動時における NVC の調節機序は安静時とは異なる. Physiol. 111:2601–2606. 10). は NVC に影響せず,視覚刺激に伴う脳の血流増加の程. 3313–3318. 11). Physiol. 111:1217–1221.. 脳血流に影響を及ぼす要因として血圧,体温および. PaCO2 が挙げられる.実験②には,運動に伴う体温お よび PaCO2 の両方の影響が含まれることから,NVC. 12). 19)ことから,暑熱環境. 29:119–122. 13). 下ではベースラインの PCA 血流が減少し,視覚刺激に. 198:270–273. 14). る PaCO2 の低下が観察されなかった.したがって,疲. Ultrasound Med. Biol. 38:13–20. 15). NVC が減少する可能性は残されている.. 108:1701–1705. 16). 17). (fMRI) of the human brain. J. Neurosci. Meth. 54:171–187. 3) Aaslid. (1987) Visually evoked dynamic blood flow response. of the human cerebral circulation. Stroke 18:771–775. 4) Conrad. & Klingelhöfer. (1989) Dynamics of regional. cerebral blood-flow for various visual stimuli. Exp. Brain. Res. 77:437–441. 5). J. Neurol. Sci. 309:58–62.. (1999) The eye and the retina. In: Zigmond et al., eds.. et al. (1994) Functional magnetic-resonance-imaging. Willie et al. (2011) Utility of transcranial Doppler ultrasound for the integrative assessment of cerebrovascular function. J. Neurosci. Meth. 196:221–237.. Szabo et al. (2011) Hypocapnia induced vasoconstriction significantly inhibits the neurovascular coupling in humans.. 6.引用文献. pp. 821–851.. Ogoh & Ainslie. (2009) Cerebral blood flow during exercise: mechanisms of regulation. J. Appl. Physiol. 107:1370–1380.. ことが今後の課題として残された.. Fundamental Neuroscience, Academic Press, San Diego, CA,. Ogoh et al. (2010) Dynamic cerebral autoregulation during and after handgrip exercise in humans. J. Appl. Physiol.. 暑熱および疲労困憊に至る強度の動的運動が視覚刺 激に伴う脳の血流応答に与える影響について検討する. Fabjan et al. (2012) The effect of the cold pressor test on a visually evoked cerebral blood flow velocity response.. 労困憊に至るほどに高い強度の動的運動により PCA 血 流が減少し,ベースラインの PCA 血流の減少により. Willie et al. (2011) Neurovascular coupling and distribution of cerebral blood flow during exercise. Neurosci. Method.. 伴う NVC も減少することが予想される. また実験②で用いた高強度運動条件では,過換気によ. Hayashi et al. (2010) Effect of intensity of dynamic exercise on pupil diameter in humans. J. Physiol. Anthropol.. に対して体温が影響していた可能性は否定できない.. 2) DeYoe. Hayashi & Someya. (2011) Muscle metaboreflex activation by static exercise dilates pupil in humans. Eur. J. Appl.. 5.今後の研究課題. 1) Reid.. Ikemura & Hayashi. (2012) Ocular circulatory responses to exhaustive exercise in humans. Eur. J. Appl. Physiol. 112:. 度は運動に対して頑強であることが示唆された.. 高体温時には脳血流は減少する. Hayashi et al. (2011) Effects of dynamic exercise and its intensity on ocular blood flow in humans. Eur. J. Appl.. 受けず安静時とほぼ同程度の値を保った.一方,静的 ことが示唆された.以上のことから,運動に伴う昇圧. Ikemura et al. (2012) Autoregulation in the ocular and. 18). Jones et al. (1979) Difference between end-tidal and arterial. PCO2 in exercise. J. Appl. Physiol. 47:954–960. 19) Nelson. et al. (2011) Reductions in cerebral blood flow during. passive heat stress in humans: partitioning the mechanisms.. J. Physiol. 589, 16:4053–4064..
(5)
図

関連したドキュメント
In the separate vibration session, the effects of stimulation location (Achilles tendons, forefoot soles) and response direction (backward-lean, forward-lean) on postural
201 Tl distribution inhibiting the Sol antigravitational activation. However factors other than muscle activity may be involved in low blood perfusion seen in Sol of the U3w group.
Ser7 is the value of an American option computed using a 100,000 path Monte Carlo simulation taking 7 terms in series (1.3) as the exercise boundary.. LUBA is the LUBA
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
In section 3, we will prove Theorem 1.4 by the following steps: in subsections 3.1 and 3.2 we will find formal solutions of the coupling equations (Φ) and (Ψ): this shows the
Before the discussion in partial differential equations, let us give a brief survey on the coupling of two ordinary differential equations in [Section 4.1 of G´ erard-Tahara [1]]..
We have found that the model can account for (1) antigen recognition, (2) an innate immune response (neutrophils and macrophages), (3) an adaptive immune response (T cells), 4)
Other important features of the model are the regulation mechanisms, like autoregulation, CO 2 ¼ reactivity and NO reactivity, which regulate the cerebral blood flow under changes
