複 写 ・可 ㈲ 隣 し.・学 内の み)/不 可
※ 年 月 日以 降 許 諾 す る
リポ ジ トリ:公 開/非 公 開
※ 年 月 日以 降許 諾 す る
生 命 科 学1464
首 都 大 学 東 京 博 士(理 学)学 位 論 文(課 程 博 士)
論 文 名 有牲 生殖 種 ハ チ ジ ョゥベ ニ シ ダ との 交雑 に よる 無 配 生 殖 種 ベ ニ シ ダ の遺 伝 的 多 様 化(英 文)
著 者 山本 薫
審査担 当者 主 査 委 員 委 員
赫 上 暫 明
z魚L
罧 嫁
上記 の論文 を合格 と判 定す る 平成2夕 年 ヒ月 λ夕日
首都 大学東京大学 院理 工学研 究科教授会
研究科長ra脅 老
DISSERTATION FOR A DEGREE OF DOCTOR OF PHILOSOPHY IN SCIENCE
TOKYO METROPOLITAN UNIVERSITY
TITLE : Genetic diversification of an apogamous fern species, Dryopteris erythrosora through hybridization with
its related sexual species, D. caudipinna
AUTHOR : Kaoru YAMAMOTO
EXAMINED BY
Examiner in chief Oi
Examiner Q k ,p-e- 10ThalAt-4')--a
Examiner
7C'----A /7 c--
QUALIFIED BY THE GRADUATE SCHOOL
OF SCIENCE AND ENGINEERING
TOKYO METROPOLITAN UNIVERSITY Dean ti
Date 1rclA, 2--1 2_0 / 3
Contents
Summary
Chapter 1
Chapter 2
Chapter 3
Chapter 4
General Introduction
The distribution of L
1
6
The distribution of Dryopteris caudipinna (Dryopteridaceae), a sexually reproducing close relative of apogamous D. erythrosora, in Japan...24
Cytological and genetic analyses in mixed populations of apogamously reproducing Dryopteris erythrosora and their sexual relative
D. caudipinna, on Izu-Oshima sland...39
Mapping and genetic analyses of mature apogamous Dryopteris
erythrosora and sexual D. caudipinna individuals in mixed populations.
Chapter 5
Dryopt
Chapter 6 Genera 1
Acknowledgements
Literature Cited....
Artificial crossing experiments between the triploid apogamous Dryopteris erythrosora and the diploid sexual D. caudipinna...
Discussion...
64
86
Summary in Japanese
108
115
117
124
Summary
Apogamy in ferns is a type of asexual reproduction by which unreduced spores are formed, and the resultant gametophytes produce the sporophytes of the next generation without fertilization. Thus, all offspring from a parent are expected to be clonal. However, apogamous species frequently show a large amount of morphological variations and often some genetic variations. Walker (1962) hypothesized that this occurs because apogamous species acquire genetic variation from related sexually reproducing species through hybridization. When triploid apogamous species hybridize with diploid sexual species, tetraploid hybrids are expected to be formed. Walker documented the formation of tetraploid apogamous hybrids between triploid apogamous and diploid sexual races of Pteris cretica in artificial crossing experiments. However, most apogamous fern species are triploids, and tetraploids are very rare. Subsequently, Lin et al (1992, 1995) hypothesized that recurrent ploidy reductions from triploid to diploid occurred in apogamous ferns, which increased their ploidy from diploid to triploid by crossing with related sexual diploid species. This hypothesis is called the
"hybridization cycle hypothesis ."
I attempted to verify the "hybridization cycle hypothesis," which explains how apogamous ferns acquire genetic variation from sexual relatives without polyploidization. This hypothesis consists of two processes: (1) hybridization between
an apogamous fern species and its closely related sexual species and (2) ploidy reduction from the triploid to diploid cytotype of the apogamous species through unequal meiosis. Although each of these two important processes has been separately documented using different apogamous fern species, the hypothesis is relevant only when the two processes occur at a time. Therefore, I considered the necessity to document the two processes in one group of ferns.
I selected apogamous Dryopteris erythrosora and its closely related sexual species, D. caudipinna, for my study and attempted to obtain evidence supporting that both hybridization and ploidy reduction occurred in apogamous fern species in nature. I also attempted to reproduce these two processes under experimental conditions and estimate the frequency of the two processes in apogamous fern species to evaluate how easily the processes that comprise the hybridization cycle hypothesis could occur in nature.
First, I examined in detail the geographical distribution of the sexual species D.
caudipinna to identify the mixed populations of the two closely related species with different reproductive modes. Dryopteris caudipinna sporadically occurs along the coastal areas of Japan, but is common and dominant in the Izu Islands. Moreover, it was suggested that these two species grow in competing frequencies on Izu-Oshima Island, Tokyo Prefecture, which lies on the northernmost part of the Izu Islands (Chapter 2).
Second, I searched for mixed populations of the two closely related species with
different reproductive modes as well as for pure apogamous populations consisting only of the apogamous species on Izu-Oshima Island. I compared the amount of genetic variation in the apogamous species between the mixed and pure apogamous populations.
The comparison showed that the genetic variation in the apogamous species was significantly higher in the former than in the latter (Chapter 3).
Third, I set up quadrats within the mixed populations on Izu-Oshima Island as well as in the forests of Jinmuji Temple, Kanagawa prefecture, and examined all the mature sporophytes growing in the quadrats cytologically and genetically to obtain additional evidence supporting the hybridization cycle hypothesis. In addition to the major triploid cytotype, diploid apogamous cytotypes of D. erythrosora were found both in the quadrats. Allozyme alleles were shared between the apogamous and sexual species in each population. Therefore, cytotypes and allelic distribution patterns predicted by the hybridization cycle hypothesis were observed in nature. Moreover, tetraploid apogamous cytotypes, which could represent hybrids between triploid apogamous D. erythrosora and diploid sexual D. caudipinna, were also observed in the two quadrats (Chapter 4).
Finally, to get more direct evidence supporting the hybridization cycle hypothesis, I performed artificially crossing experiments between the triploid apogamous D.
erythrosora and diploid sexual D. caudipinna. Triploid and tetraploid hybrids between the two species were formed at relatively high frequencies. Therefore, I could
demonstrate that the apogamous fern has a high level of crossing ability. The triploid apogamous cytotypes, which had genetic markers of both the species, could be considered to have originated through hybridization with the sexual species following ploidy reduction from triploid to diploid of the apogamous species. Thus, the processes supposed in the hybridization cycle hypothesis could be reproduced in the artificial crossing experiments. Moreover, I also demonstrated that antheridia were induced on the gametophytes of the apogamous species only when the sexual species was growing nearby. Therefore, apogamous D. erythrosora was shown to have a preference for
crossing with its sexual relatives (Chapter 5).
In this study, I could reproduce the two key processes predicted by the hybridization cycle hypothesis under experimental conditions because triploid hybrids between the triploid apogamous D. erythrosora and diploid sexual D. caudipinna were obtained. I also demonstrated that these two processes could occur relatively frequently.
Moreover, diploid apogamous cytotypes were observed in both the natural populations examined. Therefore, these results supported the hybridization cycle hypothesis.
Moreover, in the artificial crossing experiments, tetraploid hybrids were formed in even higher frequencies than the triploid hybrids. Tetraploid apogamous individuals were also found in the two natural populations investigated in this study. This suggested that tetraploid hybrids may have important roles in the processes of increasing the amount of genetic variation within the apogamous species. Accordingly, I revised the
hybridization cycle hypothesis to include the formation of
segregation of diploid apogamous cytotypes from the hybrids.
tetraploid hybrids and
Chapter 1. General Introduction
Most species of land plants, including ferns, reproduce sexually. Ferns are characterized by a life cycle that includes independent diploid and haploid phases. The haploid (n) phase is represented by hermaphroditic gametophytes, which can produce both male and female gametes (sperms and eggs) by mitotic division. The diploid (2n) phase is characterized by sporophytes, which are the conspicuous plants recognized as ferns. The life cycle of typical ferns consists of the following steps: (1) the diploid sporophyte produces haploid spores by meiosis; (2) a spore mitotically grows into a haploid gametophyte, which typically consists of a photosynthetic prothallus; (3) the gametophyte produces gametes (usually both sperm and eggs on the same prothallus) by mitosis; (4) a mobile, flagellate sperm fertilizes an egg that remains attached to the prothallus; (5) the fertilized egg is now a diploid zygote and mitotically grows into a diploid sporophyte of the next generation (Figure 1-1).
In case of sexual reproduction in most diploid ferns, motile sperms from the prothallium must swim through water to reach the eggs in the mature archegonia of a nearby, different prothallium because fertilization usually cannot occur within a gametophyte (intra-gametophytic selfing) because of severe inbreeding depression after selfing (Soltis and Soltis 1990, Watano and Iwatsuki 1988). The inability of the sperms to swim long distances makes fertilization in ferns a high-risk and costly process.
Although majority of ferns reproduce sexually, as described above, some ferns have developed the ability to reproduce asexually in the course of evolution. Apogamy or agamospory is a type of asexual reproduction observed in ferns, in which unreduced spores are formed, and the resultant gametophytes produce sporophytes of the next generation without fertilization (Manton 1950) (Figure 1-2). Apogamous reproduction is relatively common in ferns. Approximately 10% of all ferns (Lovis 1977) and 13% of Japanese fern taxa are reported to reproduce apogamously (Takamiya 1996). Unlike other asexual reproductive modes, including vegetative reproduction through gemma, apogamous reproduction involves the production of spores that can be dormant and survive for long periods, even in dry conditions, such as in the air. Therefore, apogamous reproduction is advantageous, particularly for long-distance dispersal and the formation of new populations from a small number of spores. Moreover, because it bypasses the costly process of fertilization, apogamous reproduction is considerably more effective than sexual reproduction, especially for propagation.
In contrast, apogamous reproduction is considered to have a major drawback in the production of genetic variation within species and within populations. In sexually reproducing ferns, the process of sporogenesis, via meiosis and fertilization, produces and maintains intra-specific genetic variation. Such genetic variation in local populations, as well as in each species, is important both for long-term evolution and short-term survival (Maynard Smith 1978). In case of apogamous reproduction, these
two variation-generating processes, i.e., meiosis and fertilization are absent. Only genetic clones of the parent individual are produced, leading to the decrease and eventual disappearance of genetic variation from the population. In such a situation, if there is extreme environmental change or an epidemic of infectious diseases the risk of extinction of the populations and/or species will be high.
Thus, apogamous fern species are expected to demonstrate a limited amount of genetic variation. However, such a situation is not common. In spite of the clonal nature of apogamous reproduction, several apogamous fern species demonstrate large amounts of morphological and at least some genetic variation (Watano and Iwatsuki 1988, Suzuki and Iwatsuki 1990, Lin et al. 1995, Takamiya et al. 2001, Ootsuki et al. 2011).
Previous studies analyzing genetic variation in an apogamous fern species using enzyme electrophoresis have reported 45, 4, 14, and 8 different clones from Dryopteris nipponensis Koidz. (Ishikawa et al. 2003a), Asplenium unilaterale Lam. (Watano and Iwatsuki 1988), Dryopteris bissetiana (Baker) C. Chr. (Lin et al.1995), and Cyrtomium fortunei J. Sm. (Ootsuki et al. 2011), respectively. Furthermore, genetic variation in combination with cytological variation has been reported in apogamous ferns. Six different triploid and five diploid clones have been reported in Pteris cretica L. (Suzuki and Iwatsuki 1990), and one tetraploid and four triploid clones have been reported in Diplazium doederleinii (Luerss.) Makino (Takamiya et al. 2001). Reports of apogamous fern species without intra-specific genetic variation are very rare, though Darnaedi et al.
(1990) reported that Dryopteris yakusilvicola Kurata, a triploid apogamous species endemic to Yaku Island, did not demonstrate any allozyme variation. Apogamous fern species commonly display some degree of genetic variation, even though apogamous reproduction produces only clones of parental types, and the amount of genetic variation within each apogamous species is expected to be very small.
Several hypotheses have been proposed to explain the observed levels of genetic variation within apogamous fern species: (1) recurrent origin of apogamous races from sexual species (Gastony and Gottlieb, 1985); (2) genetic segregation by homoeologous chromosome pairing (Klekowski 1973); (3) hybridization with closely related sexual species (Walker 1962, Suzuki and Iwatsuki 1990).
Gastony and Gottlieb (1985) proposed hypothesis (1); recurrent origin of apogamous species from related sexual species. Apogamous species would acquire genetic variation from their sexual relatives, if the former originated repeatedly from the latter. However, the process of sporogenesis that produces unreduced spores in apogamous ferns is extremely complicated but precise. In most sexual ferns, the archesporial cell undergoes four mitotic divisions giving rise to 16 spore mother cells in each sporangium. These subsequently undergo meiosis forming 64 spores. In contrast, in most apogamous ferns, sporogenesis follows the Dopp-Manton scheme (Manton 1950), in which the fourth mitosis is terminated in the early anaphase and a restitution nucleus is formed. The resultant eight spore mother cells possess doubled sets of
chromosomes. Subsequently, the spore mother cells undergo meiosis or two sequential cell divisions and produce 32 unreduced spores. It is unlikely mutation caused the occurrence of sporogenesis, a complicated process, originated independently several times, in several apogamous species belonging to different fern families.
Klekowski (1973) first proposed hypothesis (2); the pairing of homoeologous chromosomes. This process may be an important mechanism for generating genetic variation in apogamous ferns. For example, if the genotype of the parental triploid apogamous sporophyte is abc, the genotype of the spore.mother cells of the apogamous fern should be aabbcc. In the usual process of apogamous reproduction, the genotype of all offsprings from such a sporophyte would also be abc because only homoeologous
chromosome pairing, which leads to pairing of the same genes (a-a, b-b, c-c), occurs in the spore mother cells. In contrast, if homoeologous chromosome pairing (for example, a-b, a-b, c-c) occurs, gametophytic progeny with aac and bbc genotypes will be produced in addition to those with the abc genotype. Similarly, if another type of homoeologous chromosome pairing (a-c, a-c, b-b) occurs, the progeny with aab and bcc genotypes will also be produced. Thus, genetic segregation through homoeologous chromosome paring can generate genetic variation within apogamous fern species or even in local populations.
Ishikawa et al. (2003b) first reported positive evidence supporting genetic segregation through homoeologous chromosome pairing in the apogamous fern species
D. nipponensis. They compared genotypes of the parent sporophyte and its progeny using enzyme electrophoresis. Among 250 gametophyte progenies developed from spores of a parental sporophyte of D. nipponensis with the Pgi-2 genotype abc, four demonstrated genotypes different from that of the parent (three aac and one bbc gametophytes were detected). Similarly, among 34 sporophyte offsprings of the parent sporophyte with abc genotype, one showed bcc genotype, whereas the other 33 showed abc. Thus, Ishikawa et al. (2003b) clearly demonstrated the existence of occasional genetic segregation possibly through homoeologous chromosome paring in ferns.
Subsequently, Ootsuki et al. (2012) reported more precise data supporting genetic segregation in apogamous ferns through homoeologous chromosome paring using C.
fortunei as plant materials and nucleotide sequence information from pgiC as genetic markers. They examined a total of 732 progenies (250 gametophytes and 482 sporophytes) obtained from a parental sporophyte whose pgiC genotype was elucidated as aab. Their results indicated that 11 (4.4%) gametophytes and 8 (1.7%) sporophytes showed a different genotype (aaa) from that of the parent sporophyte. Because they analyzed a large number of progenies and detected sufficient numbers of segregated offspring, they were able to demonstrate that genetic segregation occurs in apogamous C. fortunei at a relatively high frequency. Moreover, they were able to show that the segregation frequency in gametophytes is significantly higher than that in sporophytes of the next generation (x2 = 4.90, P = 0.027). They hypothesized that these data might
suggest that apogamous fern species harbor deleterious genes and these genes might be expressed in greater numbers in sporophytes, with a more complicated morphology than in gametophytes.
Although it is now evident that genetic segregation through homoeologous chromosome paring is an important mechanism for generating genetic variation in apogamous ferns, this process is not sufficient to maintain the levels of genetic diversity that have been observed in most apogamous species. If genetic, segregation occurs at a high frequency in apogamous ferns, heterozygosity will quickly be lost within individuals and eventually within species, because of the random genetic drift.
Therefore, apogamous species must possess mechanisms that increase heterozygosity, even within individuals.
Hypothesis (3) states that hybridization with related sexual species can explain the observed levels of heterozygosity in a species or even within individuals of an apogamous fern species. The ability of apogamous ferns to cross with closely related sexual species was first demonstrated by Walker (1962), who performed artificial crossing experiments between apogamous and sexual races of Pteris cretica. He reported that haploid eggs of the sexual diploid race can be fertilized by unreduced sperms of the apogamous triploid race to produce tetraploid hybrids, which can reproduce apogamously (Figure 1-3). Suzuki and Iwatsuki (1990) supported this hypothesis by suggesting that wild populations of apogamous P. cretica in Japan were
derived through hybridization between the apogamous race of the species and its closely related sexual species P. kidoi Sa. Kurata.
However, crossing between sexual and apogamous species is still not sufficient to explain the existence of genetic variation in apogamous fern species. If an apogamous species crosses with a sexual species in the manner suggested by Walker (1962), the resultant apogamous species would become higher polyploides because the genome of the sexual species is added to that of the apogamous species with every cross. Most of the apogamous fern species are triploid; tetraploids or higher polyploides are extremely rare in apogamous ferns. Thus, the apogamous species should also have mechanisms that decrease ploidy levels.
Unequal meiosis, as reported by Lin et al. (1992, 1995), can cause a ploidy reduction in apogamous ferns. Lin et al. performed cytological and genetic studies using Dryopteris pacifica Hieron., from which no sexual races, but both diploid and triploid apogamous races have been recorded in Japan, to elucidate the origin of intra-specific cytological variation in apogamous species. They reported that among the 150 gametophyte progenies grown from the spores of a triploid parent cytologically examined by them, two (1.3%) were diploid. Among the 110 sporophyte offsprings examined, one (0.9%) was diploid. In addition, they observed unequal meiosis during sporogenesis in the triploid apogamous sporophytes, which produced diploid spores.
Thus, they demonstrated that both diploid spores and diploid gametophytes can arise
from a triploid apogamous parent. Furthermore, they demonstrated that parental triploid
apogamous sporophytes can at least occasionally form diploid apogamous sporophyte
offspring.
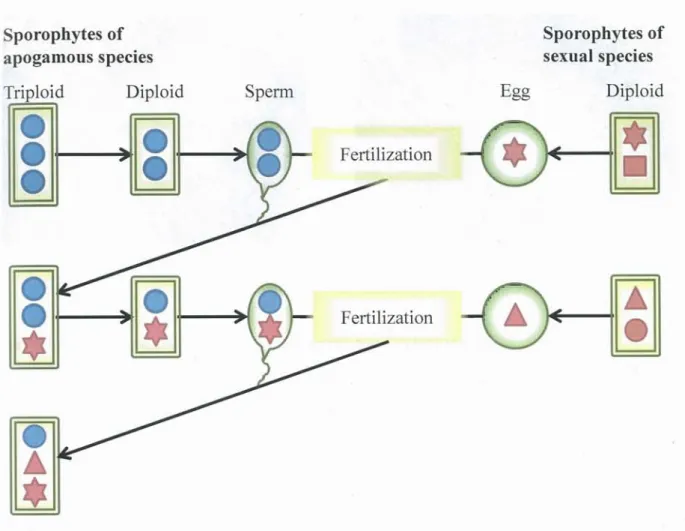
In addition, Lin et al. (1992, 1995) proposed that a recurrent cycle with ploidy reduction (oligoploidization) from triploid to diploid apogamous plants and polyploidization from diploid to triploid by crossing with related sexual diploid species (from triploid apogamous to diploid apogamous and from diploid apogamous back to triploid apogamous), might explain the existence of genetic variation in triploid apogamous fern species (Figure 1-4). According to their hypothesis, triploid apogamous fern species can incorporate genetic variation from related diploid sexual species without polyploidization. They named their proposed processes the "hybridization cycle hypothesis". According to this hypothesis, some individuals (gametophytes and/or sporophytes) of the apogamous species decrease their ploidy by unequal meiosis as the first step. In the second step, the apogamous individuals with decreased ploidy hybridize with sexual species that can normally perform meiosis and produce haploid
eggs.
The hybridization cycle hypothesis has not yet been verified because each step in the hypothesis has been separately documented in different groups of apogamous ferns
under experimental conditions. This hypothesis can be justified only when all the steps
occur together in nature. Therefore, to verify the cycle hypothesis, I proposed to
reproduce all of its steps in a particular apogamous fern species and its sexual relatives under experimental conditions. It will also be necessary to examine natural mixed populations of closely related sexual and apogamous fern species to document the evidence of the processes of the hybridization cycle hypothesis at work in natural conditions.
However, it is rare that both apogamous fern species and it sexual relatives are distributed in Japan (Takamiya 1996). It is common that only apogamous species are observed and their related sexual species are not observed in Japan. Therefore, it is difficult to find relevant fern materials, from which both closely related apogamous and sexual species can be examined. This may be the reason why the hybridization cycle hypothesis has not been tested so far. Moreover, it is even more difficult to find natural mixed populations of closely related apogamous and sexual fern species. Thus, it will be increasingly difficult to document the hybridization cycle in nature.
To verify the hybridization cycle hypothesis, I selected a triploid apogamous species, Dryopteris erythrosora (Eaton) 0. Kuntze., and its closely related diploid sexual species, Dryopteris caudipinna Nakai as the plant materials of my study (Figure 1-5). To verify the hypothesis, I considered it most important to select species from which I could obtain sporophytes and/or spores of both closely related apogamous and sexual species in Japan. The pair, apogamous D. erythrosora and sexual D. caudipinna, appeared to be the best study materials.
reproduce all of its steps in a particular apogamous fern species and its sexual relatives under experimental conditions. It will also be necessary to examine natural mixed populations of closely related sexual and apogamous fern species to document the evidence of the processes of the hybridization cycle hypothesis at work in natural conditions.
However, it is rare that both apogamous fern species and it sexual relatives are distributed in Japan (Takamiya 1996). It is common that only apogamous species are observed and their related sexual species are not observed in Japan. Therefore, it is difficult to find relevant fern materials, from which both closely related apogamous and sexual species can be examined. This may be the reason why the hybridization cycle hypothesis has not been tested so far. Moreover, it is even more difficult to find natural mixed populations of closely related apogamous and sexual fern species. Thus, it will be
increasingly difficult to document the hybridization cycle in nature.
To verify the hybridization cycle hypothesis, I selected a triploid apogamous species, Dryopteris erythrosora (Eaton) 0. Kuntze., and its closely related diploid sexual species, Dryopteris caudipinna Nakai as the plant materials of my study (Figure 1-5). To verify the hypothesis, I considered it most important to select species from which I could obtain sporophytes and/or spores of both closely related apogamous and sexual species in Japan. The pair, apogamous D. erythrosora and sexual D. caudipinna, appeared to be the best study materials.
Dryopteris erythrosora is a species of dryopteroid fern (Dryopteridaceae) characterized by the following morphological features: (1) triangular and bipinnate fronds, (2) costal or subcostal sori, and (3) bullate scales present on the abaxial side of the pinna rachis and midribs of segments. D. caudipinna is a close relative of D.
erythrosora and is extremely similar to it in gross morphology, but tends to have a narrower pinnula than D. erythrosora. However, its morphology-based recognition of D.
caudipinna from apogamous D. erythrosora is not always successful (Yamamoto and Nakaike 1983) because individuals of the apogamous species sometimes have a narrow pinnula and look quite similar to D. caudipinna. I have referred to the two closely related species with different reproductive modes, D. erythrosora and D. caudipinna,
together as the D. erythrosora complex from here on in this thesis. These two species belong to sect. Polystichodrys according to the classification system of the genus Dryopteris by Ito (1939) and to sect. Erythrovariae of subg. Erythrovariae according to that by Frazer-Jenkins (1986).
In this study, I have attempted to carefully investigate natural mixed populations of the apogamous fern species and its related sexual species to find the traces of hybridization and ploidy reduction indicated by the hybridization cycle hypothesis in nature. For this purpose, it is important to locate relevant mixed populations in the field.
However, it was expected that verifying such a complicated hypothesis in nature would be extremely difficult. In addition, I attempted to reproduce the following two important
steps of the cycle hypothesis under culture conditions: (1) hybridization between the triploid apogamous D. erythrosora and the diploid sexual D. caudipinna and (2) ploidy reduction from the triploid apogamous type to the diploid apogamous type D.
erythrosora, by unequal meiosis. Furthermore, I attempted to clarify possible frequencies of the two steps under experimental conditions to evaluate the ease with which the processes of the hybridization cycle hypothesis can occur in nature.
. This thesis is structured as follows: In the following chapter (Chapter 2), I will describe the results of my study, which attempt to clarify the geographical distribution of Dryopteris caudipinna, a sexually reproducing fern species, which is expected to be closely related to the apogamous D. erythrosora, in Japan. The current section was intended for selecting mixed populations of the sexual and apogamous species. It was well known that apogamous D. erythrosora is widely distributed in Japan, from Hokkaido to Kyushu, from the lowlands to the mountains having altitudes of 1,000 m or higher (Kurata and Nakaike 1985). In contrast, sexual D. caudipinna was considered extremely rare, and its geographical distribution was not clear. I hypothesized that mixed populations of apogamous D. erythrosora and sexual D. caudipinna would be easily found when the distribution of the latter species was clarified.
Furthermore, in Chapters 3 and 4, I have investigated natural mixed populations of apogamous D. erythrosora and sexual D. caudipinna, which were selected on the basis of the distribution data obtained in Chapter 2, using cytological and genetic
methods. The aim of Chapters 3 and 4 was to obtain evidences that support the
hypothesis that the apogamous fern incorporates genetic variation of its related sexual
species in nature through the processes indicated by the hybridization cycle hypothesis.
In Chapter 3, I have surveyed several mixed populations of the two closely
related species with different reproductive modes as well as pure apogamous
populations consisting of only apogamous D. erythrosora on Izu-Oshima Island, Tokyo
Prefecture, where coexistence of the two species in competing frequencies was
suggested by the results of Chapter 2. I subsequently compared the amount of genetic
variation observed in the pure apogamous population with that in the mixed population
to obtain evidence indicating that apogamous ferns incorporate genetic variation from
their sexual relatives.
In Chapter 4, I have set quadrats within the mixed populations on Izu-Oshima Island and in Jinmuji Temple, Kanagawa Prefecture for comparison. I examined all mature sporophytes growing in the quadrats both cytologically and genetically to obtain additional evidence that may support that the processes proposed in the hybridization cycle hypothesis are operating in nature.
In Chapter 5 of this thesis, I performed artificial crossing experiments between triploid apogamous D. erythrosora (male) and diploid sexual D. caudipinna (female) to obtain more direct evidence supporting the hybridization cycle hypothesis, and
quantitatively examined the crossing ability of the apogamous fern species.
A.
Ycng sporophyte
(2n)
Fertilization
Archegonium
Meiosis
+,,,,,,mpGametop ( n)
•
Egg
(n) Sperm
(n)
• 4:5
Antheridium
Figure 1-1. Life cycle of a fern (sexual reproduction).
In the life cycle of a typical sexually reproducing fern, first, a diploid sporophyte produces haploid spores by meiosis. Next, a spore grows into a haploid gametophyte by mitosis. The
gametophyte then produces gametes (sperm and eggs) again by mitosis.
The sporophyte of the next generation is produced through fertilization of the egg by a sperm.
Adult sporophyte (2n)
Young sporophyte
(2n)
Fertilization
Meiosis
Spore (2n) Gametophyte
(2n)
Figure 1-2. Life cycle of a fern (apogamous reproduction).
In the life cycle of an apogamously reproducing fern, unreduced spores are formed without normal meiosis. The spores grow into gametophytes, which produce the sporophytes of the next generation without fertilization.
Apogamous species
Triploid Sperm
(4))14:
Fertilization• •
•
Sexual species
Egg Diploid
Tetraploid hybrid (Apogamous cytotype)
Figure 1-3. Hybridization between closely related triploid apogamous and diploid sexual species of fern.
The reduced egg of the diploid sexual species is fertilized by unreduced sperm of the triploid apogamous species to produce a tetraploid hybrid. The hybrid possesses the genomes of both parental species and can reproduce apogamously. The blue solid circle indicates the genome of the apogamous species; the red solid star and the red solid square indicate the genome of the sexual species.
Sporophytes of apogamous species
Triploid Diploid Sperm
Fertilization
Sporophytes of sexual species
Egg Diploid
•
• • Fertilization
2. c.
• 0
Figure 1-4. Hybridization cycle hypothesis proposed by Lin et al. (1992, 1995).
In this hypothesis, recurrent ploidy reduction (oligoploidization) occurs from triploid apogamous sporophytes to diploid apogamous ones, and polyploidization occurs from diploid to triploid through fertilization of a haploid egg of the sexual diploid species by diploid sperm of the related apogamous species. Through these processes, the triploid apogamous species (originally consisting only of the genome indicated by the blue circle) acquires genetic variation from the sexual species (indicated by the red solid star and the red solid triangle).
Figure 1-5. Photographs of the two fern species used in this study.
a. Dryopteris caudipinna Nakai (sexual species)
b. Dryopteris erythrosora (D.C. Eaton) Kuntze (apogamous species).
Chapter 2. The distribution of Dryopteris caudipinna (Dryopteridaceae), a sexually reproducing close relative of apogamous D. erythrosora, in Japan
2.1. Introduction
The autumn fern, Dryopteris erythrosora (D. C. Eaton) Kuntze is one of the most commonly observed fern species in Japan, showing very large morphological
diversity (Iwatsuki 1995). It was reported as a triploid apogamous species (Takamiya 1996). Therefore, its closely related sexual species may contribute to the formation of genetic diversity of the apogamous fern species. Only two sexual diploid species, which may have been involved in the formation of the apogamous species complex, are recognized in Japan: D. caudipinna Nakai, which was originally described on Hachijojima Island, and D. koidzumiana Tagawa from the Ryukyu Islands. Information on their distribution, especially of the former species, has only been documented sporadically or is unpublished. The number of spores per sporangium can be an indicator of the reproductive mode of most leptosporangiate ferns (Takamiya 1996); the presence of 64 spores usually suggests sexual reproduction, while 32 spores suggest apogamy. In this chapter, I updated information related to their geographical distribution mainly by counting the number of spores per sporangium using herbarium specimens.
Originally, Dryopteris caudipinna was not recognized as a sexual counterpart of D. erythrosora, but was thought to be a relative with a narrow pinnula when Nakai (1931) described it. Succeeding studies revealed that D. erythrosora is an apogamous triploid (Kurita 1961), and that plants morphologically corresponding to D. caudipinna are sexual diploids (Hirabayashi 1970). As emotional in Chapter 1, D. caudipinna tends to have a narrower pinnula than D. erythrosora, but its morphology-based recognition from apogamous D. erythrosora is not always successful (Yamamoto and Nakaike 1983) because apogamous individuals sometimes have a narrow pinnula and look quite similar to D. caudipinna. Therefore, the reliable identification of D. caudipinna should be based on counting the spore number per sporangium, counting chromosomes, or determining the genome size. In other words, the taxon is defined by its reproductive mode, not by morphological characters.
Previously known localities confirmed by spore or chromosome counting included Tsukuba-shrine in Ibaraki Prefecture (Fujimoto 1998), Jinmuji in Kanagawa Prefecture (Yamamoto and Omori 1991), Izu-Oshima Island (Nakaike and Yamamoto 1984) and Hachijojima Island (Hirabayashi 1970, Yamamoto and Nakaike 1983) in Tokyo Prefecture, Kanmuri Island in Kyoto Prefecture (Mitsuda 1990), the Shimane Peninsula in Shimane Prefecture (Miyamoto and Nakamura 1990), and Kirishima in Miyazaki and Kagoshima Prefectures (Miyamoto and Nakamura 1990). According to Iwatsuki (1995), its distribution is confined to the Izu Islands, Miura Peninsula, and Izu
Peninsula; however, it is distributed over a much wider range. Although both Hirabayashi (1974) and Miyamoto and Nakamura (1990) published distribution maps of D. caudipinna, it is unclear whether they examined spore or chromosome numbers, as they did not state the sources of their data.
2.2. Materials and Methods
Candidate specimens of D. caudipinna based on frond shape were selected from approximately 3,500 sheets of D. erythrosora and D. caudipinna deposited in the Makino Herbarium of Tokyo Metropolitan University (MAK) and the National Museum of Nature and Science (TNS). The spore number per sporangium was counted under a stereomicroscope for all of the candidates, unless they were uncountable. For specimens from the Izu Islands, except Izu-Oshima and Hachijojima Islands, for which comprehensive spore counts had already been made, all of the specimens were selected for spore examination to determine the general ratio of sexual to apogamous individuals on the islands.
2.3. Results
The spore number per sporangium was counted for approximately 500 sheets of the selected specimens. Among them, 241 specimens bear 64 spores and about 50 specimens bear 32 spores per sporangium (Figure 2-1). The spore numbers of the rest specimens were uncountable. Sporangia taken from the following specimens contained 64 spores and their morphological characters matched those of typical D. caudipinna (Figure 2-2). The specimens from previously known localities are excluded from the list.
Distribution map of D. caudipinna is shown in Figure 2-3.
JAPAN:
Ibaraki Pref.:
Nishiibaraki-gun, Iwama-machi, Yamane (1981-11-08, M. Yasu s.n., TNS-VS-934523).
[Makabe-gun,] Makabe-machi, Sakayori -- Shiio Yakuoin (1973-12-02, H. Ito s.n., TNS-VS-1038281).
Chiba Pref.:
Chosei-gun, Ichinomiya-machi, Gundari-yama (2010-03-08, K. Yamamoto and A.
Ebihara 100308G1-8,14,15,18,24,25,27, MAK).
Kanagawa Pref.:
[Fujisawa-shi,] Enoshima (1979-09-18, M. Minemura s.n., TNS-VS-461080).
Tokyo Pref.:
Toshima Isl. (1933-07-30, Y. Jotani, 8202, TNS-VS-815690; 1933-07-31, Y. Jotani s.n., 121073; Y. Jotani 8207 TNS-VS-815683; Y. Jotani 8210, TNS-VS-815979; 1957-07-18, Y. Jotani 26210, TNS-VS-815688).
Niijima Isl., Hon-mura (1981-05-04, H. Endo s.n., TNS-VS-601629; 1981-05-??, H.
Endo s.n., TNS-VS-931539).
Kozushima Isl., Tenjo-san (1936-07-24, Y. Jotani 6312, TNS-VS-815697; Y. Jotani 6313, TNS-VS-815698); precise locality unknown (1936-07-26, Y. Jotani 6311, TNS-VS-815695); precise locality unknown (1966-07-26 to 1966-08-03, H. Ohba 3018,
TNS-VS-173140, 222713).
Mikurajima Isl., Sato (1934-07-26, Y. Jotani 10816, TNS-VS-121076; Y. Jotani 10818, TNS-VS-815794; 1934-07-27, Y. Jotani 10353, TNS-VS-815792); Oyama (1934-07-26, Y. Jotani 10817, TNS-VS-815973); precise locality unknown (1967-07-29, Y. Jotani et al. 1279, TNS-VS-815788; 1967-07-31, T. Matsuoka 1104, TNS-VS-815787;
1967-08-02, K. Inoue s.n., TNS-VS-1017491).
Miyakejima Isl., Oyama (1932-08-17, Y. Jotani 5034, TNS-VS-815708); Oyama, alt.
300m (1980-10-10, H. Hatta and S. Matsumoto, living collection of Tsukuba Botanical Garden 13490, TNS-VS-9508224, 9508226); precise locality unknown (1936-07-11, K.
Hayashi •s.n., TNS-VS-56244; 1961-04-02, S. Otaki 3058, TNS-VS-1038468;
1964-07-??, collector unknown, TNS-VS-815713; 1964-07-10, collector unknown, TNS-V S-816077, 816078).
Aogashima Isl., Ikenosawa (1958-07-20, Y. Jotani 27607, TNS-VS-815715, 815716, 815967, 815968; 1977-10-31, M. Kato and E. Miki 112, TNS-VS-369811);
Todaisyo-jinjya (1988-12-21, T. Shimakawa s.n., TNS-VS-522201).
Aichi Pref.:
Atsumi-gun, Atsumi-cho, Yamada (1980-11-23, K. Inukai 4201, TNS-VS-550286).
Tottori Pref.:
[Iwami-gun, Fukube-mura] Mani-san (1931-11-08, N. Hiratsuka s.n., TNS-VS-512216).
Shimane Pref.:
Masuda-shi, Takashima (1980-10-10, N. Miake s.n., TNS-VS-449584).
Oki Islands, Nishinoshima Isl., Oki-gun, Nishinoshima-cho, Mita, Takuhi-yama (2008-07-01, Y. Kazumi s.n., TNS-VS-1105395).
Yamaguchi Pref.:
Yamaguchi-shi, Niho, Asaji River (1993-02-20, H. Masaki s.n., TNS-VS-745192;
1993-08-12, H. Masaki s.n., TNS-VS-745190).
Toyoura-gun. [Toyota-cho,] Toyotanaka-mura (1947-08-23, K. Oka 1229, TNS-VS-276102).
Toyoura-gun, Hohoku-cho, Awano, midstream of Futanoi River (1993-08-30, H. Masaki s.n., TNS-VS-745191).
Otsu-gun, Yuya-cho, Sakane-ue (1992-08-22, H. Masaki s.n., TNS-VS-745177, 745178).
Kochi Pref.:
Sukumo-shi, Okinoshima Isl. (1971-07-23, T. Inobe s.n., TNS-VS-1038280).
Nagasaki Pref.:
Tsushima Isl., Kamiagata-gun, Kamitsushima-cho, Izumi, alt. 40--50m (1979-07-21, S.
Tsutsui 17479, TNS-VS-573603);
Kamiagata-gun, Mine-mura, Kamizato, along Shishinouchi-kawa alt. 60m (1979-07-19, S. Tsutsui 17322, TNS-VS-573599);
Kamiagata-gun, Kamitsushima-cho, Eko, alt. 20--30m (1979-07-20, S. Tsutsui 17432, TNS-VS-573602);
Kamiagata-gun, Kamiagata-cho, Mitake, alt. 80--90m (1979-07-19, S. Tsutsui 17349, TNS-VS-573601);
Kamiagata-gun, Kamiagata-cho, Mitake, alt. 200--400m (1968-09-28, S. Tsutsui s.n., TNS-VS-204608);
Shimoagata-gun, Izauhara-cho, Ariake-yama, (1968-09-23, H. Koyama 2819, TNS-VS-204386; 1978-02-26, S. Tsutsui 15131, TNS-VS-573596).
Tsushima-shi, Mitsushima-cho, Kanedajo (2007-10-09, K. Fuse 292, TNS-VS-768795);
Tsushima-shi, Izuhara-cho, Nainkawa (2007-10-06, K. Fuse 182, TNS-VS-768813).
Fukuoka Pref.:
[Tagawa-gun, Kawara-machi,] Kawara-dake (1937-08-26, M. Irie s.n., TNS-VS-513221; 1951-8-26, H. Ouchi s.n., TNS-VS-100314).
Kagoshima Pref.:
Okuchi-shi, Manegahira (1959-08-22, T. Yamanaka s.n., TNS-VS-734596).
Aira-gun, Kamo-cho, Yadome-dake (1983-07-19, M. Kawabata s.n., TNS-VS-742738, 742740, 742741).
[Izumi-shi,] Jonodan -- Shibi -- Miyanojo (1957-08-15, S. Kurata 882, TNS-VS-724546).
SOUTH KOREA:
Cheju-do Isl. (1962-07-28, S.-Y. Oh. 2673, TNS-VS-1038337).
Soheuksan-do Isl. (1949-08-05, O. Yongsok 8098, TNS-VS-151180).
2.4. Discussion
I confirmed the occurrence of Dryoptteris caudipinna in 13 Prefectures of Japan and in two islands of Korea, including seven Prefectures and korea as new records (Figure 2-3). I also confirmed its almost complete distribution on the major Izu Islands, except Shikinejima Island, which lacked specimens with countable spores. Our investigation widened its distribution range north to Iwama in Ibaraki Prefecture, east to Gundari-yama in Chiba Prefecture, south to Yadome-dake in Kagoshima Prefecture, and west to Soheuksan-do Island, Korea. In contrast to the wide distribution of D.
erythrosora which completely covers the warm temperature region in Japan (Kurata and Nakaike 1985), Dryopteris caudipinna tends to be distributed on islands and in coastal areas under the influence of warm currents (the Kuroshio and Tsushima Currents), although it also occurs in mountainous areas in Kyushu, as its southern limit of distribution. This sexual fern species usually occurs under Castanopsis forests, and its localities in the Kanto district (northern limit) are mostly well-preserved precinct groves.
However, the distribution of D. caudipinna does not seem to be simply delimited by the climatic condition, because the species has not yet been found on the Izu and Kii Peninsulas, which probably share similar meteorological conditions with the above localities (for the Izu Peninsula, see Nakaike et al. 1984). Dryopteris caudipinna usally forms mixed populations with apogamous D. erythrosora, although its pure populations lacking apogamous sheets of the specimens were checked for individuals occur on
Hachijojima (Yamamoto and Nakaike 1983), Toshima, Niijima, Kozushima, Mikurajima, and Aogashima Islands. Ecological and historical factors which determines the ratio of the sexual to the apogamous forms is still unclear.
Another problem concerns the relationship between the two sexual diploid taxa D. caudipinna and D. koidzumiana. The latter is usually treated as a species endemic to the Ryukyu Islands, and is characterized by abruptly narrowing pinnae apices. Their distributions apparently do not overlap each other, but the difference in their frond morphological characters is quite subtle. At present, I have little evidence for the presence of two independent biological units corresponding to the two sexual diploids.
Additional analyses with ample sampling should be performed in mixed populations of apogamous Dryopteris erythrosora and sexual D. caudipinna. Future analysis should be also made to elucidate relationship between the two diploid sexual
species, D. caudipinna and D. koizumiana.
a IP
•
b
100 Et m 100 )t m
Figure 2-1. Pictures of spores in sporangia.
a. The sporangium contains 64 spores, indicating sexual reproduction (S. Tsutsui 17349, TNS-VS-573601) .
b. The sporangium contains 32 spores, indicating apogamous reproduction (S. Kobayashi
2993, TNS-VS-573459).
C
4
I
d i
1
,.,„,
Figure 2-2. Representative specimens of Dryopteris caudipinna.
a. Ibaraki Prefecture, Tsukuba-shi, Tsukuba Shrine (1987-08-23, S. Matsumoto s.n., living collection of the Tsukuba Botanical Garden 77398, TNS-VS-9508257)
b. Aichi Prefecture, Atsumi-gun, Atsumi-cho, Yamada (1980-11-23, K. Inukai 4201, TNS- VS-550286)
N
W
S
•
•
„int
l
• 0
•
y
0 400 km
Figure 2-3. Distribution map of Dryopteris caudipinna.
Open triangles, previously known localities; Closed circles, study.
new localities reported in this
N
W E
S
Kozu Isl.
Mikura Isl.
Hachij o Isl.
Aogashima Isl.
Oshima Isl.
Toshima Isl.
Ni] ima Isl.
Miyake Isl.
0 50 km
Figure 2-4. Pie charts showing the approximate ratio of sexual Dryopteris caudipinna (white) to apogamous Dryopteris erythrosora (black).
The counts are based on Nakaike and Yamamoto (1984) for Oshima Island, on Yamamoto and Nakaike (1983) for Hachijojima Island and on spore observations made in this study for the remaining islands.
Chapter 3. Cytological and genetic analyses in mixed populations of apogamously reproducing Dryopteris erythrosora and their sexual relative D. caudipinna, on Izu-Oshima Island
3.1. Introduction
In Chapter 2, the geographical distribution of Dryopteris caudipinna, a sexually reproducing close relative of the apogamously reproducing Dryopteris erythrosora, was clarified. According to the results obtained in the main islands of Japan, apogamous D.
erythrosora is widely distributed and extremely common, whereas sexual D. caudipinna is rare and its distribution is confined to coastal areas. In contrast, in the Izu Islands, Tokyo Prefecture, Japan, sexual D. caudipinna is dominant and common, whereas apogamous D. erythrosora is relatively rare there. In general, sexual species appear to have difficulty in growing together with closely related apogamous species because the fertility and propagation of sexual species has disadvantages compared with their apogamous relatives, which can reproduce without costly fertilization. This should be one of the major reasons for the dominant distribution of either apogamous D.
erythrosora or sexual D. caudipinna in a particular locality in Japan. However, these two fern species with different reproductive modes are observed growing together with competitive frequencies on Izu -Oshima Island, which lies in the northern most part of the Izu Islands and is closest to Honshu, the largest main island of Japan. Therefore,
Izu-Oshima Island may provide a rare opportunity to examine mixed populations of closely related sexual and apogamous fern species.
Moreover, putative hybrids between D. erythrosora and D. caudipinna have been reported on the Izu-Oshima Island (Nakaike and Yamoto 1984). As already mentioned in the General Introduction (Chapter 1), Walker (1962) documented the hybridization between related apogamous and sexual fern species in artificial crossing experiments using Pteris cretica. He attempted to hybridize sperms developed on the gametophytes of its triploid apogamous type and eggs developed on those of the diploid sexual type and succeeded in producing a tetraploid hybrid with the ability of apogamous reproduction. Thus, it appeared likely that similar hybridization events may take place in natural populations of the Dryopteris species growing on Izu-Oshima Island.
I selected the Izu-Oshima Island as my study site to examine mixed populations of apogamous D. erythrosora and sexual D. caudipinna. I considered the need to clarify the geographical distribution of the apogamous and the sexual species of Dryopteris and to determine exact localities of mixed populations on Izu-Oshima Island. Such a detailed distribution within the island was not elucidated by the investigation of Chapter 2 based only on herbarium specimens because the specimens were sporadically collected and did not cover the entire distribution within the island. Therefore, I should examine the possibility of hybridization between the apogamous and the sexual species in their natural mixed populations.