生物間相互作用に基づくマツ材線虫病発病メカニズ ムの統合的理解
山口, 莉未
https://doi.org/10.15017/4060237
生物間相互作用に基づく
マツ材線虫病発病メカニズムの統合的理解
山口 莉未
目次
第1章 序論 ... 1
第1節 研究の背景 ... 1
1.1. マツ材線虫病の発生とその被害 ... 1
1.2. マツ材線虫病の枯死メカニズム ... 2
1.3. マツ材線虫病における宿主の要因 ... 3
1.4. マツ材線虫病における病原体の要因 ... 4
1.5. マツ材線虫病の発生と病徴進展に関わる環境要因 ... 5
第2節 研究の目的 ... 8
第2章 マツノザイセンチュウの動態とクロマツの防御反応との関連性の解明 ... 10
第1節 クロマツ樹体内におけるマツノザイセンチュウ頭数測定法の確立 ... 10
1.1. はじめに ... 10
1.2. 材料と方法 ... 11
1.3. 結果 ... 12
1.4. 考察 ... 13
第2節 マツノザイセンチュウの動態とクロマツの防御反応の時空間的解析 ... 19
2.1. はじめに ... 19
2.2. 材料と方法 ... 20
2.3. 結果 ... 23
2.4. 考察 ... 24
第3節 小括 ... 35
第3章 クロマツ抵抗性および感受性クローンにおける宿主−病原体間相互作用の解明 ... 36
第1節 抵抗性および感受性クロマツにおける宿主−病原体トランスクリプトーム同時解析 ... 36
1.1. はじめに ... 36
1.2. 材料と方法 ... 39
1.3. 結果 ... 42
1.4. 考察 ... 47
第2節 小括 ... 75
1.4. 考察 ... 82
第2節 小括 ... 96
第5章 総合考察 ... 97
5.1. 感染初期におけるマツノザイセンチュウの動態とクロマツの防御反応 ... 97
5.2. クロマツの抵抗性反応 ... 99
5.3. マツ材線虫病の発病メカニズムと今後の展望 ... 101
摘要 ... 104
謝辞 ... 106
引用文献 ... 107
第 1 章 序論
第1節 研究の背景
1.1. マツ材線虫病の発生とその被害
我が国の代表的なマツ類であるクロマツ(Pinus thunbergii Parl.)およびアカマツ(P. densiflora)は,
乾燥地や貧栄養な土壌といった厳しい環境への適応性が高く,青森から鹿児島まで広範に分布する。特
にクロマツは耐潮性に優れるため,江戸時代より海岸林造成の主要樹種として用いられ,防風林および
防砂林として機能を果たしてきた。両樹種は古くから白砂青松といわれるような海岸や湖沼の美しい景
観を構成することに加え,詩歌,伝統芸能,美術,造園,信仰など日本の文化とも深く結びついている
(梅津,2016)。さらに,薪炭材や用材として用いられるなど利用価値も高い。しかし,これらのマツ
類は,マツ材線虫病の病原体であるマツノザイセンチュウ(Bursaphelenchus xylophilus (Steiner & Buhrer)
Nickle; pinewood nematode; 以下,PWN)に対し強い感受性を示し(図1-1),本病によって我が国のマ
ツ林は衰退の一途を辿っている。
マツ材線虫病に関する最初の記録は,1905年頃に発生した長崎市内におけるマツの集団枯損にまで遡
ることができる(矢野,1913)。1940年代には九州全域と瀬戸内海沿岸部を中心に甚大な被害が認めら
れ,1970年代に入ると中国地方の日本海側,中部地方の各県や関東北部など全国に拡大した(藤本ら,
第1章
は,1979年度の243万㎥を最大値として減少傾向にはあるものの,2017年度の被害材積は40万㎥であ
り,依然として我が国最大の森林病害である(林野庁,2018)。
マツ材線虫病は,我が国だけではなく世界的な脅威とされている。東アジアでは,1980年代に中国
(Cheng et al., 1986),台湾(Tzean and Jan, 1985),韓国(Yi et al., 1989)において本病の被害が確認さ
れている。さらに,1999年にはポルトガル(Burgermeister et al., 1999),2008年にはスペイン
(Abelleira et al., 2011)といった西ヨーロッパにまで被害は拡大し,現在PWNはEU諸国を始めとする
40カ国以上で検疫対象に指定されている(竹内,2019)。
1.2. マツ材線虫病の枯死メカニズム
PWNがマツ材線虫病の病原体である(清原・徳重,1971)ことが特定されて以降,これまで40年以
上に亘りマツ材線虫病による枯死メカニズムを明らかにするための研究が精力的に行われてきた。日本
では,PWNは主にマツノマダラカミキリ(Monochamus alternatus)によって媒介され,カミキリがマツ
類の若枝をかじる後食時に傷口から侵入する(Mamiya and Enda, 1972)。PWNの樹体内侵入後,マツ樹
体内では様々な組織学的・細胞学的変化が生じる。Fukuda(1997)は,本病の病徴進展を感染初期と病
徴進展期の2段階に分類した。感染初期では,PWNの侵入後,樹脂道における移動,摂食に伴い,マ
ツの防御反応としてエピセリウム細胞および柔細胞における二次代謝産物の産生,仮道管への脂質代謝
病の感染初期における病徴は非親和性の場合でも発現する一方で,病徴進展期における病徴は親和性の
場合にのみ発現し,この段階に移行すると宿主は枯死を免れることができないとされる。
1.3. マツ材線虫病における宿主の要因
植物の病気が発生するためには,植物側が病原体を受け入れる性質を備えていることが必要である。
このような性質を感受性といい,逆に受け入れない性質を抵抗性という(大木,2007)。清原・徳重
(1971)は,マツ属9種に対する接種実験により,PWNに対する抵抗性は樹種間で差があることを報
告している。さらに,二井・古野(1979)はマツ属30種に対する接種実験を行い,マツ属内で明らか
に抵抗性樹種と感受性樹種が併存することに加え,種内においても抵抗性の個体間差が存在することを
示唆している。抵抗性に樹種間での差異(堂園ら,1973; Bedker et al., 1987; Oku et al., 1989; Kuroda et al.,
1991)および樹種内での差異(Kuroda, 2004; 川口,2006; Mori et al., 2008)が認められることは複数の
研究によって支持されており,それらの研究結果から抵抗性個体ではPWNの移動・増殖を阻害する因
子があるものと推察される。さらに,抵抗性のリギダマツ(P. rigida)とテーダマツ(P. taeda)の雑種
であるリギテーダマツ(P. rigi x taeda)の主軸を加熱処理した場合にPWNの移動が阻害されなかった
こと(Oku et al., 1989)や,テーダマツの切枝ではPWNが顕著に増殖したこと(真宮,1982)などか
ら,抵抗性因子として動的抵抗性の関与が考えられる。しかし,この動的抵抗性に関与すると考えられ
る物質については,リギテーダマツで認められた水溶性のPWN不動化物質(Oku et al., 1989)や,スト
第1章
1.4. マツ材線虫病における病原体の要因
植物病理学において広く用いられているように,秋庭(2006)は,マツ材線虫病における病原性をあ
る樹種を枯死させることができるかという質的なものとし,病原力をその樹種をどの程度枯死させるこ
とができるかという量的なものとして定義している。PWNは東アジアに分布するシロマツ(P.
bungeana),アカマツ,リュウキュウマツ(P. luchuensis),タイワンアカマツ(P. massoniana),クロ
マツ,およびヨーロッパに分布するヨーロッパクロマツ(P. nigra),ヨーロッパアカマツ(P.
sylvestris),フランスカイガンショウ(P. pinaster)には病原性を示す(Vicente et al., 2012)。その一方
で,原産国である北米のマツ類に対しては病原性を持たないものと考えられている(秋庭,2006)。こ
の北米におけるマツ類とPWNとの関係性は,同じくBursaphelenchus属であり北半球に広く分布するニ
セマツノザイセンチュウ(B. mucronatus)が分布域のマツ類には病原性を示さない(Pereira et al.,
2013)関係性と類似している。PWNやニセマツノザイセンチュウの場合,野外における枯損木あるい
はカミキリから分離された個体群を培養したものをアイソレイトと呼ぶ。これらの線虫は有性生殖によ
って繁殖するため,作出されたアイソレイトは遺伝的にはある程度多様な個体を含む(竹本,2012)。
PWN種内においては,このアイソレイト間で病原力に強弱があることが報告されている(Kiyohara and
Bolla, 1990)。清原(1977)は,全国各地から採取されたPWNアイソレイトの培地上における増殖力の
(Odani et al., 1985; Fukuda et al., 1992; Ikeda and Kiyohara, 1995; Ichihara et al., 2000),あるいは接種箇所
から離れた部位でも検出されるが増殖は認められないこと(Futai, 1980b; Son and Moon, 2013b)が報告
されている。したがって,PWNの病原性および病原力は,樹体内における増殖力と強く関連すると考
えられる。
1.5. マツ材線虫病の発生と病徴進展に関わる環境要因
植物の病気の発生には感受性のある宿主と病原性のある病原体の他に,好適な環境が必要であり,温
度,湿度,降雨などの物理的環境が含まれる(Scholthof, 2007)。温度や湿度などの非生物的な因子は,
病原体に対する宿主の感受性や病原体の成長・生殖・感染に影響を及ぼす(Sturrock et al., 2011)ため,
気候変動による宿主と病原体の相互作用の変化は,病害発生の最も大きな原動力となる可能性があると
考えられる。マツ材線虫病における発病あるいは病徴進展の程度は,温度(清原,1973; Sikora and
Malek, 1991; Melakeberhan et al., 1992),光条件(金子・陳野,1986; Kaneko, 1989; 川口・玉泉,2006;
松永ら,2015),水分条件(大山ら,1975; 大山ら,1976; 岩崎ら,1999; Braasch, 2000)などの環境因
子によって影響を受けることが報告されている。これらの因子のうち,光条件と水分条件は宿主の光合
成などの生理機能に関わるため,PWNに対するマツの抵抗力を変化させるものであると考えられる。
実際に,川口・玉泉(2006)は,クロマツに対する庇蔭処理は,PWNの移動に対しては著しい影響を
及ぼさなかったが,PWNの増殖を早め,爆発的な増殖と同時に針葉の変色が生じたことを報告してい
第1章
なること(清原,1973)や,移動・増殖の両方が活発になること(Ichihara et al., 2000)が示され,結果
としてより病徴が進展したことが報告されている。
図1-1 マツ材線虫病の被害を受けたアカマツ(佐賀県嬉野市嬉野町,2019年8月)
第1章
第2節 研究の目的
これまでに実施されたマツ材線虫病を対象とした研究によって得られた知見から,感染初期から病徴
進展期に移行する際の現象であるPWNの顕著な増殖がマツを枯死に至らしめる要因であると考えられ
る。しかし,樹体内におけるPWN増殖の可否を決定するマツ側とPWN側それぞれの要因についての
情報は不足しており,結果としてなぜマツは発病に至るのかは明確でない。一般的に,侵入した病原体
に対し,宿主の防御反応が生じても時期や量が不適切であったり,その病原体に対する有効な反応が抑
制されたりすると発病に至る(山田,2006)。PWNに感受性を示すマツ類の場合も,PWNの活動抑制
に効果的な防御反応ができないために発病し,枯死に至ると推測される。一方,PWNは北米原産の線
虫であり,北米のマツ類には激害をもたらさない(秋庭,2006)が,アジアやヨーロッパのマツ類の多
くに病原性を示すことを考慮すれば,原産地の宿主との共進化によって獲得された病原性が,耐性を持
たない感受性マツ類の樹体内で発揮される,あるいはそれらマツ類の防御反応に対する耐性を付与して
いる可能性がある。したがって,マツ材線虫病の発病メカニズムを理解するためには,PWNの感染
後,樹体内におけるPWNの移動・増殖といった動態とその病原性,それに対する宿主の防御反応と病
徴進展について,両者の詳細な情報を得る必要があると考えられる。
本研究では,PWNに強い感受性を示すクロマツを対象として,宿主と病原体間の相互作用,および
相互作用の発現に関わる環境条件を視野に入れた基礎研究を分子生物学的手法により行った。第2章で
の移動・増殖能に影響を及ぼす環境要因である温度が異なる条件下で接種実験を行うことで,マツ樹体
内におけるPWNの増殖過程と病徴進展との関連性について明確にした。第5章では,第2章から第4
章までに得られた宿主・病原体・環境の3つの要因に関する知見をもとに,マツ材線虫病の発病メカニ
ズムについて総合考察を行った。
第2章
第 2 章 マツノザイセンチュウの動態とクロマツの防御反応との関連性の解明
第1節 クロマツ樹体内におけるマツノザイセンチュウ頭数測定法の確立
1.1. はじめに
植物体内や土壌中の線虫頭数を測定する際には,慣習的に線虫を分離後,顕微鏡下において目視で数
える手法が用いられてきた。しかし,本手法はサンプルの保存状態や分離条件,人為的過誤によって測
定結果にばらつきが生じることが報告されている(Yan et al., 2013)。PWNを木片から分離する際に用
いられるベールマン法は,2日間の分離時間を設けた場合でも全PWN数の70–80%程度しか分離できな
いとされる(真宮,1975; Son and Moon, 2013a)。そのため,本手法は少数のPWNを検出することが特
に難しいとされ,測定対象のサンプルを一定期間室温で放置し,十分に増殖させた後に定量する手法が
用いられた例もある(Kuroda et al., 1988; 黒田・伊藤,1992)。
近年,植物寄生性線虫の定量検出を目的として,real-time polymerase chain reaction(PCR)法が用いら
れている(Takeuchi and Futai, 2009; Sato et al., 2010; Baidoo et al., 2017)。本手法は,植物体内もしくは
土壌中の線虫を分離することなくDNAを抽出し,測定対象のDNA配列をマーカーとして特異的に検出
する手法であるため,前述した従来の手法における分離と測定の際の問題点を克服することが可能であ
る。マツ材線虫病に関する研究では,real-time PCR法は主に感染木の診断を目的として,線虫集団もし
発されてきた(Cardoso et al., 2012)。樹体内におけるPWNの定量検出について,Takeuchi and Futai
(2009)は,インターカレーション法により2種類のDNAマーカー(ITS regionsとβ-tubulin)を用い
ることで,樹体内のPWNを1頭から検出し,定量する手法を確立している。インターカレーション法
とは,二本鎖DNAに入り込んだ蛍光物質が発した蛍光強度を測定することで,PCR産物量を測定する
手法である。しかし,本手法は,目的産物ではない非特異的増幅産物を測定する恐れがあるため,正確
な定量を妨げてしまう可能性が指摘されている(Suarez et al., 2005)。さらに,Takeuchi and Futai
(2009)の研究では,使用されたDNAマーカー間で検量線の精度が異なっていたことから,マーカー
についても更なる検討が必要であると考えられる。
本節では,ハイブリダイゼーション法であるTaqMan quantitative real-time PCR(qPCR)法を利用し,
樹体内のPWN頭数をより高い精度で測定する手法を確立した。TaqMan qPCR法では,PCRプライマー
に加え,目的配列に結合するDNAプローブを使用する。このDNAプローブは蛍光物質で標識されてい
るため,目的産物のみを検出することができる。したがって,前述のインターカレーション法と比較し
てより特異的に目的産物を定量することができるとされている。本研究では,目的領域のコピー数が定
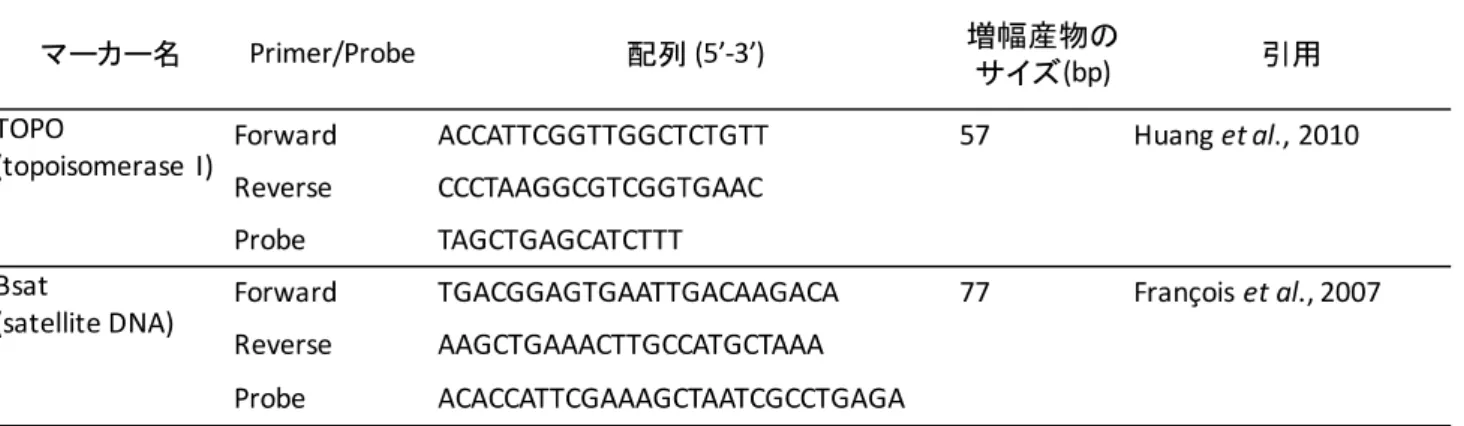
量結果に与える影響についても検討するために,topoisomerase I(TOPO,シングルコピー)とsatellite
DNA(Bsat,マルチコピー)の2つのマーカーを使用した。
1.2. 材料と方法
第2章
ルは液体窒素中で凍結させ,オートミル(Tokken, Inc.)を用いて粉砕し,isolation buffer[10% (w/v)
polyethylene glycol, 0.35 M sorbitol, 0.1 M Tris-HCl, 0.5% (w/v) spermidine, 0.5% (w/v) spermine, and 5% (v/v)
mercaptoethanol]で洗浄した。サンプルは遠心分離することで上清を除去し,DNeasy Plant Mini Kit
(Qiagen)のプロトコルに従ってDNAを抽出した。DNAは50 µLの超純水に溶出した。
1.2.2. TaqMan qPCR分析
反応は,StepOnePlus™ Real-time PCR System(Applied Biosystems)を用いて行った。反応溶液とし
て,16 µL溶液[TaqMan® Universal Master Mix II, no UNG (Applied Biosystems) 10 µL, a mixture of primers
and probes (250 nM) 1 µL, water 5 µL]に,5倍希釈したDNA 4 µLを加え,総量を20 µLに調整した。
PCRプライマーとプローブは,TOPO(Huang et al., 2010)とBsat(François et al., 2007)を使用した(表
2-1)。反応溶液は,95˚Cで10分加温後,40サイクル(95˚C 15秒,55˚C 1分)の条件で増幅させた。
分析ではサンプルあたり3反復を設けた。
1.3. 結果
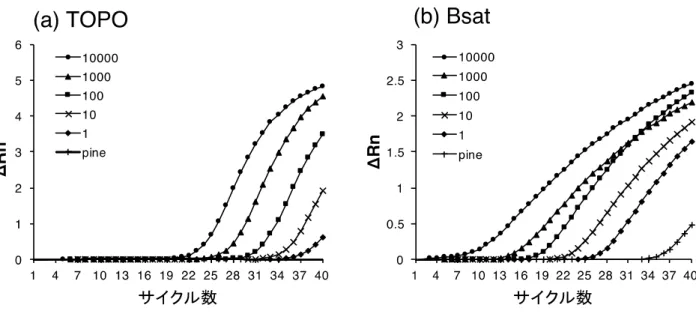
クロマツ木片と1頭のPWNを混合したサンプルにおけるthreshold cycle(Ct)値は,各マーカーで
35.05(TOPO)と27.39(Bsat)であった(表2-2)。木片のみのサンプルにおけるCt値は,Bsatマーカ
ーを使用した場合には36.37であったのに対し,TOPOマーカーを使用した場合にはPCR産物の増幅は
1.4. 考察
DNA topoisomerase Iは,全ての真核生物のゲノム上に存在し(Huang et al., 2010),その配列はシング
ルコピーである。Huang et al.(2010)は,TaqMan qPCR法でtopoisomerase Iをターゲットとしたマーカ
ーを用いることで,0.01 ngのgenomic DNAもしくは1頭のPWNを検出可能であったことを報告してい
る。Satellite DNAも同様に,真核生物の多くが有している特徴的な配列であるが,ゲノム上に複数配列
が存在する(Grenier et al., 1997)。François et al.(2007)は,satellite DNAをターゲットとしたマーカー
を用いることで,1 pgのgenomic DNAもしくは1頭のPWNを検出可能であったことを報告している。
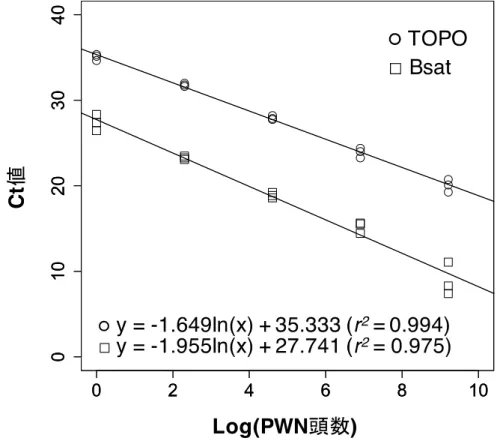
本研究では,マーカーとしてこれらTOPO(Huang et al., 2010)とBsat(François et al., 2007)を用いた
結果,両マーカーにおいてマツ木片中に存在するPWN 1頭分のDNAを検出することができた(表2-
2,図2-1)。この結果は,これらの配列がクロマツとPWN間で異なっており,TaqMan qPCR法によっ
てクロマツDNA中のPWNのDNAを特異的に検出可能であることを示している。クロマツ木片中の
PWN 1頭に対するCt値は,それぞれ35.05(TOPO)と27.39(Bsat)であった(表2-2)ことから,
Bsatマーカーはより検出感度が高いことが示された。これは,satellite DNAはゲノム上に複数配列存在
することから,Bsatマーカーを使用した場合,シングルコピーのTOPOマーカーを使用した場合と比較
して検出可能な蛍光強度に至るまでのサイクル数が少なくなるためであると考えられる。しかしなが
ら,シングルコピーであるTOPOマーカーにおいてもマツ木片中の1頭のPWNを検出できたことか
第2章
のコピー数がPWN個体間で異なっていたためであると考えられる。実際に,PWN 1頭に対するCt値は
それぞれ35.05 ± 0.37(TOPO)と27.39 ± 0.95(Bsat)(表2-2)であり,標準偏差はBsatマーカーで大
きかった。したがって,TaqMan qPCR法でTOPOマーカーを用いることにより,マツ樹体内における
PWN頭数を高い精度で定量できると考えられる。
表2-1 TaqMan qPCR分析に使用したプライマーとプローブ
第2章
表2-2 スタンダードサンプルのthreshold cycle (Ct) 値
図2-1 TaqMan qPCR分析による増幅曲線
(a)topoisomerase I(TOPO),(b)satellite DNA(Bsat)の結果をそれぞれ示す。
第2章
図2-2 TaqMan qPCR分析によって作成された検量線
topoisomerase I(TOPO),satellite DNA(Bsat)の結果をそれぞれ示す。
第2節 マツノザイセンチュウの動態とクロマツの防御反応の時空間的解析
2.1. はじめに
病原体が天然分布域以外から侵入した場合,侵入地の生物はその病原体に対する抵抗性を備えていな
いために壊滅的な被害を受ける可能性がある(Hansen, 2008)。PWNに感受性を示すマツ類は,菌の感
染や昆虫の食害といった生物的ストレスに対しては克することができるが,PWNに対しては感染初期
からテルペノイドやタンニン様物質といった防御物質を産生するにも関わらず,その活動を抑制するこ
とができない(Kuroda, 2008)。むしろ,マツは自らの過剰な防御反応の結果,枯死に至るとする説が
提唱されている(二井,1984; Myers, 1988; Iwahori and Futai, 1993; 二井,2003; Hara et al., 2006)。Hirao
et al.(2012)は,クロマツ抵抗性および感受性クローンにPWNを接種した結果,感受性クローンにお
いて感染特異的(pathogenesis-related, PR)タンパク質などの抗菌タンパク質様遺伝子群が速やかに高い
レベルで発現誘導されることを報告した。同様の現象がクロマツの他クローン(Nose and Shiraishi,
2011)の他に,同じくPWNに対し感受性を示すタイワンアカマツ(Xu et al., 2013)においても認めら
れている。PRタンパク質は,病原体の侵入や非生物的ストレス,過敏感反応,全身獲得抵抗性に応答
して蓄積する(Veluthakkal and Dasgupta, 2010)ため,感受性マツ類の遺伝子発現レベルにおける防御反
応に関する知見は,マツは自らの防御反応によって枯死に至るとする仮説を支持するものである。しか
し,第1節で示したように従来の手法では樹体内のPWNを定量的に検出することに難があることか
ら,それらマツの防御反応と樹体内におけるPWNの移動・増殖との関連性については正確に評価され
第2章
本節では,クロマツ樹体内におけるPWNの動態とマツの防御反応との関連性を明らかにすることを
目的として,クロマツ苗にPWNを接種後,1・3・7・14日目(days post-inoculation, DPI)に主軸3箇所
におけるPWN頭数と,1・3・7 DPIにおけるクロマツPR遺伝子群(PR-2,PR-5,PR-6)の発現量を評
価した。樹体内のPWN頭数測定には,本章第1節で確立したTaqMan qPCR法を用いた。PR-2,PR-5,
PR-6は,先行研究(Hirao et al., 2012)においてクロマツ感受性クローンにおいて高発現していた遺伝子
であり,一般的に,PR-2とPR-5の発現はサリチル酸(salicylic acid, SA)シグナル経路活性のマーカー
として,PR-6の発現はジャスモン酸(jasmonic acid, JA)およびエチレン(ethylene, ET)シグナル経路
活性のマーカーとして使用されている(Derksen et al., 2013)。
2.2. 材料と方法
2.2.1. 供試材料とマツノザイセンチュウの接種
九州大学箱崎キャンパス内のファイトトロンにおいて,非抵抗性クロマツ2年生実生苗32個体を1
週間順化させた。試験期間中の平均気温は29.8˚C(26.8°C–37.5°C)であった。灌水は試験期間中毎日十
分に行った。
2017年8月2日に,クロマツ苗20個体の地際部分に強病原性であるPWN(Ka4アイソレイト)
10,000頭(/50 µL)を剥皮法にて接種した。対照として,12個体には蒸留水を同様の手法で接種した。
ルは,速やかに液体窒素中で凍結させ,−80°Cで保存した。供試苗についての情報は,表2-3に示し
た。
2.2.2. DNA分析とTaqMan qPCR
木片(0.1 g)を本章第1節で述べた手法に従って,DNA分析とTaqMan qPCR分析に供試した。PCR
プライマーとプローブはtopoisomerase I(Huang et al., 2010)を使用した。分析ではサンプルあたり3反
復を設けた。
2.2.3. RNA分析とqRT-PCR
木片(0.1 g)をDNA分析と同様の手法で粉砕後,PureLink® Plant RNA Reagent(Invitrogen)のプロト
コルに従ってRNAを抽出した。RNAはPureLink RNA Mini Kit(Invitrogen)とRNase-free DNase I
(Qiagen)を用いて精製した。RNA濃度はNanoDrop spectrophotometer(Thermo Scientific)で測定し
た。7 DPIの1個体の中部と14 DPIの全てのサンプルからはRNAが抽出できなかったため,以降の分
析から除外した。RNA(500 ng in a final volume of 20 µL)は,SuperScript® IV First-Strand Synthesis
System(Invitrogen)のプロトコルに従ってFirst-strand cDNAに合成された。
Quantitative real-time reverse transcription (qRT)-PCRの反応溶液として,16 µL溶液[Fast SYBR® Green
Master Mix (Applied Biosystems) 10 µL, primers (300 nM) 1 µL, water 5 µL]に,10倍希釈したcDNA(10
ng)4 µLを加え,総量を20 µLに調整した。PCRプライマーは,Hirao et al. (2012) のPR-2,PR-5,PR-
6を使用し,同じく内在性コントロールとしてelongation factor 1-alphaを使用した(表2-4)。反応溶液
第2章
2.2.4. データ解析
全ての統計解析は,R version 3.4.3(R Core Team, 2017)を用いて行った。Real-time PCR分析によって
得られたPWN頭数とPR遺伝子群の相対発現量のデータ(それぞれ3反復の平均)は,log10 (x + 1) に
変換した。
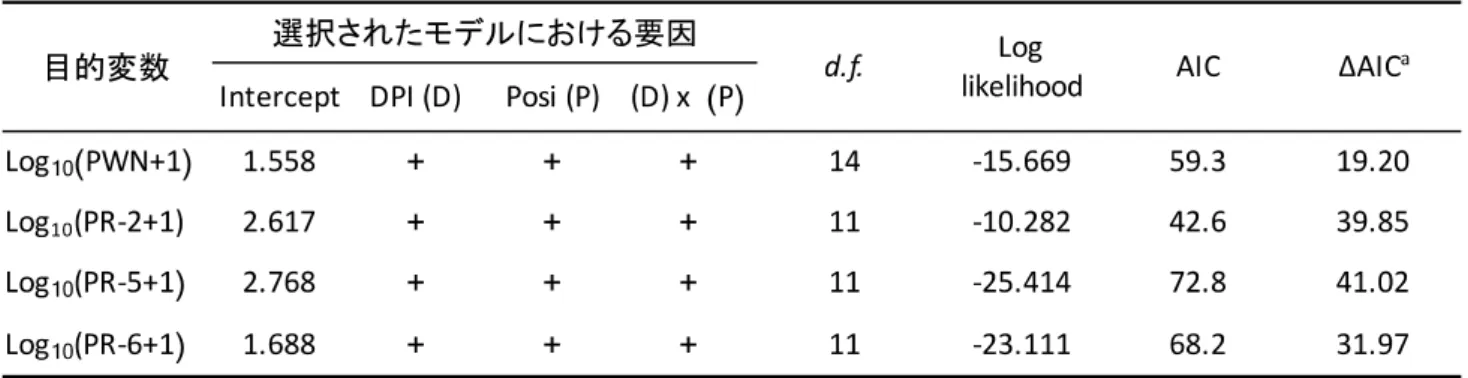
PWN頭数とPR遺伝子群の相対発現量に対する,時点(DPI),部位(Posi),およびそれらの交互
作用(DPI × Posi)の効果をlme4 package(Bates et al., 2015)の“lmer” functionを用いて線形混合モデル
により解析した。DPI,Posi,およびDPI × Posiはそれぞれ固定効果とし,苗個体を変量効果として扱っ
た。モデル選択は,MuMIn package(Bartoń, 2018)の“dredge” functionを用いて赤池情報量基準
(Akaike’s information criterion, AIC)により行った。PWN頭数とPR遺伝子群の相対発現量のベストモ
デルに交互作用項が含まれていたため,データをDPI別に分割し,再度モデル選択を行った。各DPIの
データセットに対しPosi間での多重比較をmultcomp package(Hothorn et al., 2008)の“glht” functionを
用いてTukey’s honest significant difference (HSD) multiple comparison testsにより行った。
各クロマツ苗におけるPWN頭数とPR遺伝子群の相対発現量について3部位の平均を時点間で比較
するために,one-way analysis of variance(ANOVA)とTukey’s HSD multiple comparison testを行った。
各時点と各部位におけるPWN頭数とPR遺伝子群の相対発現量間で,ピアソンの積率相関係数を求
めた。
2.3. 結果
2.3.1. 植物体内におけるマツノザイセンチュウの頭数
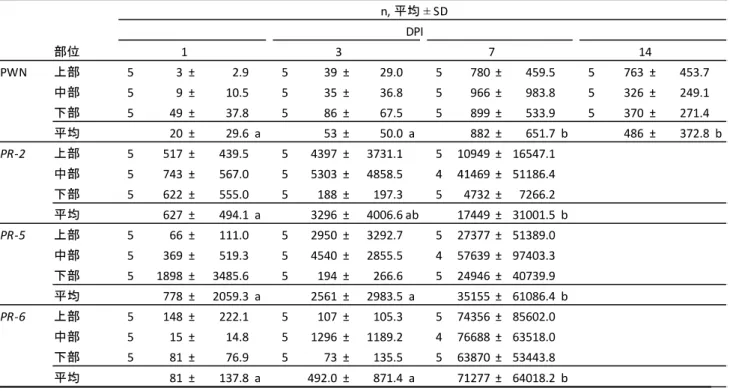
1・3・7・14 DPIのクロマツ主軸内におけるPWN頭数をTaqMan qPCR法により測定した結果を表2-
5に示す。モデル選択の結果,DPI,Posi,DPI × Posiを含むモデルがベストモデルとして選択された
(表2-6)。各DPIのデータセットについて解析した結果,1 DPIと14 DPIのデータにおいて,Posiの
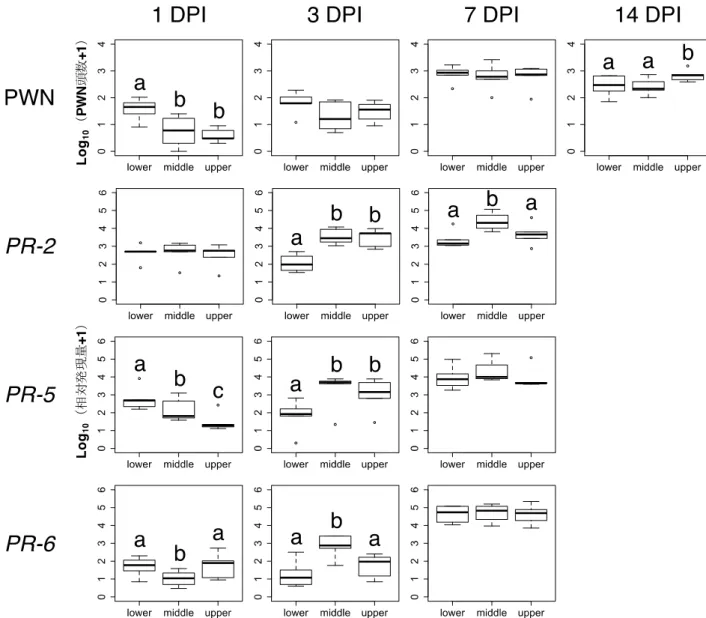
効果を含むモデルの方がAICは小さかった。1 DPIでは,PWNは主軸3箇所全てで検出されたが,下部
において検出されたPWN頭数が最も多かった(図2-4)。一方,14 DPIでは,上部においてPWN頭数
は最も多い結果となった。
One-way ANOVAとTukey’s HSD multiple comparison testの結果,苗あたりの平均PWN頭数はDPI間
で有意差があり,PWN頭数は1・3 DPIと比較して7・14 DPIに有意に増加していた(表2-5)。
2.3.2. クロマツ防御関連遺伝子群の発現
1・3・7 DPIのクロマツ主軸内におけるPR-2,PR-5,PR-6の相対発現量をqRT-PCR法により評価し
た結果を表2-5に示す。遺伝子発現解析は,各時点と各部位において,水接種したサンプルに対する
PWN接種したサンプルの相対発現量を算出することで行った。モデル選択の結果,DPI,Posi,DPI ×
Posiを含むモデルがベストモデルとして選択された(表2-6)。各DPIのデータセットについて解析し
た結果,PR-2の3 DPIと7 DPI,PR-5とPR-6の1 DPIと3 DPIのデータにおいて,Posiの効果を含むモ
デルの方がAICは小さかった。PWN接種後,これらPR遺伝子群の発現量は全ての部位で顕著に増加
第2章
量は中部で最も高くなっていたが,PR-5とPR-6の相対発現量については部位間で有意差は認められな
かった。
One-way ANOVAとTukey’s HSD multiple comparison testの結果,苗あたりのPR遺伝子群の平均発現
量はDPI間で有意差があり,相対発現量は1・3 DPIと比較して7 DPIに有意に増加していた(表2-
5)。
2.3.3. マツノザイセンチュウの頭数とクロマツ防御関連遺伝子群の発現との関連性
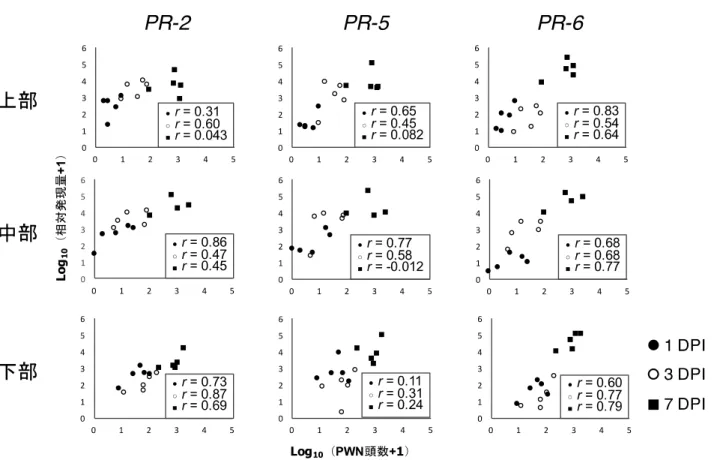
Log10 (x + 1)に変換した各時点および各部位でのPWN頭数とPR遺伝子群の相対発現量間には,有意
な相関は認められなかった(p > 0.05)。しかし,各時点の各部位,および全体として両者間に正の相関
がある傾向が認められた(図2-5)。
2.4. 考察
2.4.1. マツノザイセンチュウの移動とクロマツの防御反応との関連性
感染初期のクロマツ樹体内におけるPWNの分布と組織の応答については,複数の研究が実施されて
いる。Hara et al.(2006)は,放射柔細胞における急激な細胞学的変化が,接種箇所近傍だけではなく,
樹体全体でPWNが増殖する以前に観察されたことを報告した。一方,Ichihara et al.(2000)は,感染初
期での病徴進展や組織学的変化がマツ組織におけるPWNの移動と関連することを報告しており,この
箇所近傍に分布しており,ごく少数のPWNが接種箇所から離れた部位でも検出された(表2-5,図2-
4)。しかし,PR遺伝子群の相対発現量は苗の全ての部位で顕著に増加していた(表2-5)。一方で,
各時点と各部位においてPWN頭数とPR遺伝子群の相対発現量には正の相関がある可能性が示唆され
た(図2-5)。以上のことから,感染初期では,PWNの迅速な移動によって,マツ苗では全身において
防御反応が誘導され,さらに,その反応の強度は近傍に存在するPWN頭数に比例する可能性があると
考えられる。
本研究では,クロマツ苗の防御反応のマーカーとして,SA応答のPR-2およびPR-5とJA/ET応答の
PR-6を使用した。SAやJA,ETなどの植物ホルモンを介したシグナル応答は,植物–病原体相互作用に
おいて重要なプロセスであり,通常,SAシグナル経路はJA/ETシグナル経路と拮抗する(Yang et al.,
2015)。PR遺伝子群の空間的な発現挙動については,シロイヌナズナ(Arabidopsis thaliana)にテンサ
イシストセンチュウ(Heterodera schachtii)を感染させた際,SA応答のPR-1,PR-2,PR-5は根と葉の
両方で発現誘導された一方で,JA応答のPR-3,PR-4は根では発現レベルに変化はなく,葉ではPR-4
が発現減少したことが報告されている(Hamamouch et al., 2011)。その一方で,A. thalianaにサツマイ
モネコブセンチュウ(Meloidogyne incognita)を感染させた際には,SAとJA応答双方のシグナル経路は
根で活性化したことが報告されている(Hamamouch et al., 2011)。本研究では,1 DPIにおいてPWNの
大部分が接種箇所近傍で検出され,3 DPIでは検出されたPWN頭数は部位間で差が認められなかった。
しかし,PWNの分布に対してPR遺伝子群の発現パターンは遺伝子によって異なっており,SA応答で
第2章
の現象には,SAやJA/ETなどの植物ホルモンによって媒介されるシグナル経路が関与している可能性
がある。
2.4.2. マツノザイセンチュウに対するクロマツの過剰な防御反応
マツ材線虫病において,感染初期から病徴進展期に移行するとマツは枯死を免れることができないと
される(Fukuda, 1997)。Hirao et al.(2012)は,抵抗性クロマツと比較して,感受性クロマツにおいて
PRタンパク質などの抗菌タンパク質に関連する遺伝子群が感染初期から高いレベルで発現誘導され,
その強度は7 DPIで顕著に増加することを報告している。本研究では,感受性クロマツ苗の主軸3箇所
において,PR遺伝子群の発現とPWN頭数を経時的に評価した。1 DPIと3 DPIでは,PR遺伝子群は各
部位において高発現していたが,7 DPIにPWN頭数とPR遺伝子群の相対発現量は顕著に増加していた
(表2-5,図2-4)。さらに,外部病徴は全ての苗で14 DPIに発現していた(表2-3)。これらの結果
は,感染初期には植物体全体で防御反応を行っているにも関わらず,この反応はPWNの増殖抑制には
効果的でないことを示唆する。
PWNは北米を除く地域に分布するマツ類に対しては外来種であるため,これら地域に分布する感受
性マツ類のPWNに対する防御反応は,これらのマツ類が頻繁に遭遇する菌類などの病原体のために用
意されたものであると考えられる(二井,2003)。そのため,PWNはそれらの標準的な防御反応を克
する感染メカニズムを有していると考えられる。複数の研究が,PWNは移動経路における障害物質を
れらの分子が,感染初期における感受性マツ樹体内でのPWNの生存に関連している可能性がある。し
たがって,クロマツ苗では,PWNの迅速な移動に伴って全身で防御反応が誘導されるが,この反応は
PWNが移動や防御能を有しているためにその活動を抑制することができないと推察される。その結
果,植物体全体でPWNの爆発的な増殖による顕著な防御反応が誘導され,枯死に至ると考えられる。
第2章
表2-3 供試材料の苗高と外部病徴
表2-4 qRT-PCR分析に使用したプライマー(Hirao et al., 2012)
第2章
表2-5 PWN頭数とPR遺伝子群の発現量
表2-6 AICに基づいた線形混合モデルの選択
第2章
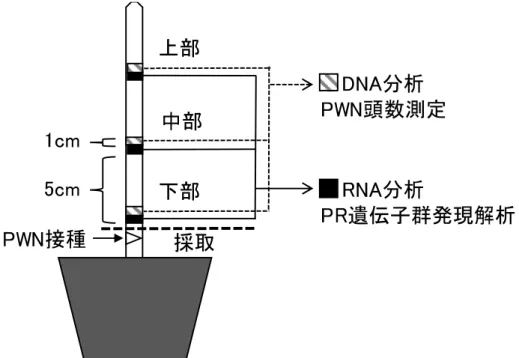
図2-3 サンプル採取箇所
図2-4 各時点および各部位におけるPWN頭数とPR遺伝子群の発現量
各接種後経過日数(days post inoculations,DPI)における結果をそれぞれ示す。異なるアルファベ
ットはTukey’s HSD multiple comparison tests(p < 0.05)によって有意差が認められたことを示す。
第2章
図2-5 PWN頭数とPR遺伝子群の関係性
第3節 小括
本章では,TaqMan qPCR法を利用することで,樹体内のPWNを分離することなく直接的に定量する
手法を確立した。ゲノム上にシングルコピーで存在するTOPOとマルチコピーで存在するBsatの2マー
カーを使用した結果,両マーカーにおいてマツ木片中に存在する1頭のPWNを検出することが可能で
あった。両マーカーで作成した検量線を比較した結果,TOPOマーカーでより高い決定係数が得られた
ことから,本マーカーを使用することで,高い精度でマツ樹体内のPWN頭数を推定することが可能で
あると考えられた。本手法を利用して,クロマツ実生苗3部位におけるPWN頭数と植物の防御関連遺
伝子であるPR遺伝子群の発現量を経時的に評価することで,両者の関連性を明らかにした。感染初期
では,PWNの大部分は接種箇所近傍に分布しており,ごく少数のPWNが接種箇所から離れた部位で検
出されたが,PR遺伝子群は全身で強く発現上昇していた。その後,PWN頭数とPR遺伝子群の発現量
は苗全体で顕著に増加し,全ての苗が外部病徴を示した。したがって,PWNの迅速な移動によって苗
全体で防御反応が誘導されるものの,この防御反応はPWNの増殖抑制に効果的ではなく,結果として
枯死に至ると考えられる。
第3章
第 3 章 クロマツ抵抗性および感受性クローンにおける宿主−病原体間相互作用の解明
第1節 抵抗性および感受性クロマツにおける宿主−病原体トランスクリプトーム同時解析
1.1. はじめに
マツ材線虫病の被害拡大抑制のために,1973年から農林水産技術会議の特別研究「マツ類材線虫の防
除に関する研究」が実施され,本病に対する抵抗性育種の可能性と重要性が明らかにされた(大庭ら,
1983)。これを契機として実施された多くの研究開発を背景として,1978年から西日本地域において
「マツノザイセンチュウ抵抗性育種事業」が開始された(戸田,2004)。この育種事業では,本病の激
害地における生存木の中から抵抗性クロマツ候補木14,620個体が選抜され,人工的なPWN接種により
抵抗性を検定することで,最終的に16個体が選抜された。選抜されたマツの抵抗性は,自然受粉家系
への接種検定における生存率に従って,抵抗性の強い方から順に5–1の5段階でランク付けされた(戸
田,2004; Hirao et al., 2012)。しかし,これら複数品種存在するクロマツ抵抗性クローンの質的な差異
については不明である。
大庭ら(1983)は,マツの抵抗性要因のうち,PWNとマツとの関係性について,抗性作用(樹体内
に侵入したPWN増殖を抑制する)と耐性(PWNによる傷害を修復する,あるいはそれに耐える)を想
定している。Kusumoto et al.(2014)は,PWNを接種したクロマツ抵抗性クローン(波方73,ランク
hydrolase family proteinが高発現していた一方で,感受性クローンではPR-1b,PR-2,PR-3や
leucoanthocyanidin dioxygenaseが高発現しており,抵抗性には活性酸素を除去する能力が関連する可能性
があることを述べている。Hirao et al.(2012)は,抵抗性クローン(波方73)では7・14 DPIにPR-9,
PR-10,細胞壁関連遺伝子群が高発現していた一方で,感受性クローンではPRタンパク(PR-1b,2,
3,4,5,6)質などの抗菌タンパク質関連遺伝子群が1・3・7 DPIに高発現したことを報告している。
この結果から,彼らは感受性クロマツでは過剰な防御反応が誘導される一方で,抵抗性クロマツでは細
胞壁関連遺伝子群が誘導されており,これら防御反応の違いが鍵となることを示唆している。これらの
研究から,クロマツの抵抗性メカニズムには,抵抗性のうち動的な「耐性」が関与する可能性があると
考えられる。しかし,クロマツ抵抗性クローンを用いた研究は極めて少ないうえに,PWNの病原性を
加味したクロマツの抵抗性および感受性因子についての議論は不足している。
Hogenhout et al.(2009)は,エフェクターを宿主細胞の構造と機能を変える全ての病原体タンパク
質および小分子として包括的に定義している。植物寄生性線虫は,植物体に侵入後,宿主細胞の分解や
植物の防御反応の抑制,植物の防御シグナル経路への干渉といった様々な機能をもつエフェクターを分
泌する(Haegeman et al., 2012)。PWNの遺伝子基盤に関しては,2007年にexpressed sequence tagsの解
析が行われ(Kikuchi et al., 2007),2011年にドラフトゲノム配列が決定されている(Kikuchi et al.,
2011)。以降,PWNの病原性について,トランスクリプトーム解析やプロテオーム解析などから得ら
れた生体分子情報が多数報告されている。Shinya et al.(2013b)は,PWNの病原性について,マツ樹体
第3章
については,PWNは活性酸素や様々な化学物質に対する防御に関わる分子を有する可能性が示されて
いる(Shinya et al., 2010; Shinya et al., 2013a; Qiu et al., 2013; Cardoso et al., 2016; Espada et al., 2016; Tsai et
al., 2016)。擬態分子や,他の植物寄生性線虫において植物の防御反応を抑制する分子に類似する配列
が検出されたとする報告(Shinya et al., 2013a; Espada et al., 2016)は,PWNがマツの防御反応を阻害す
る可能性を示唆するものである。さらに,Hu et al.(2019)は,PWNのトランスクリプトーム解析から
特定されたエフェクター候補BxSapB1がベンサミアナタバコ(Nicotiana benthamiana)の細胞死を引き
起こすことを明らかにし,BxSapB1がPWNの病原性に寄与する可能性を示した。しかし,これらの知
見は,樹体から分離した,あるいは木片を浸した液中でのPWNの反応から推定されたものである
ため,これら病原性候補分子のマツ樹体内における発現を実際に検証する必要がある。
Dual RNA-sequencing(RNA-seq)解析は,宿主細胞と病原体を物理的に分離することなくRNAを抽
出し,in silicoで双方のトランスクリプトームを推定する手法である。そのため,測定された遺伝子発
現データは,巨視的な相互作用による変化を直接反映しており,分離過程でのバイアスを避けることが
できる(Schulze et al., 2016)。近年,このdual RNA-seq解析は,植物病害における宿主−病原体間の相
互作用を理解するために様々な種間で使用され,その有効性が認められている(Kawahara et al., 2012;
Hayden et al., 2014; Meyer et al., 2016; Petitot et al., 2016; Chen et al., 2017; Shukla et al., 2018; Kovalchu et al.,
2019)。
往の研究(Nose and Shiraishi, 2011; Hirao et al., 2012; Kusumoto et al., 2014)から,感染初期にクロマツ抵
抗性および感受性クローン間において組織・遺伝子発現レベルで反応に違いが認められたため,1・3
DPIのサンプルを対象に解析を行った。クロマツ抵抗性3クローン(波方37,ランク4; 三崎90,ラン
ク4; 田辺54,ランク2)の交配実験より,抵抗性に関する遺伝要因数は1.96個と推定されている(倉
本ら,2007)ことから,抵抗性因子は複数存在すると考えられる。そこで,本研究では,供試材料とし
てクロマツ抵抗性2クローン(波方37,三崎90)を用いることで,複数の抵抗性因子についての知見
を得ることも試みた。
1.2. 材料と方法
1.2.1. 供試材料とマツノザイセンチュウの接種
クロマツ抵抗性クローンは波方37と三崎90(双方ともランク4)を使用し,感受性クローンは県福
岡2号を使用した。供試木48個体(平均苗高47.8 ± 9.6 cm)は全て2016年3月頃に森林総合研究所林
木育種センター九州育種場内で接ぎ木された。これら供試木は,2017年3月から九州大学箱崎キャンパ
スの野外において育成し,試験の1週間前に九州大学箱崎キャンパス内のファイトトロンに移動させ
た。試験期間中の平均気温は29.8°C(26.8°C–37.5°C)であった。灌水は試験期間中毎日十分に行った。
2017年8月2日に,各クローン12個体の接ぎ木部分の約5 cm上部に強病原性であるPWN(Ka4ア
イソレイト)10,000頭(/50 µL)を剥皮法にて接種した。対照として,各クローン4個体には蒸留水を
第3章
に供試し,DNA分析を行った部分の上部(接種箇所上部0.1 g)をRNA分析に供試した。水接種した個
体は,PWNを接種した個体と同じ部分をRNA分析に供試した。
1.2.2. DNA分析とTaqMan qPCR
DNA分析およびTaqMan qPCR分析は,第2章第1節に記述した手法に従って行った。PCRプライマ
ーとプローブはtopoisomerase I(Huang et al., 2010)を使用した。TaqMan qPCR分析ではサンプルあたり
3反復を設けた。
1.2.3. RNA分析とシーケンシング
RNA分析は,第2章第2節に記述した手法に従って行った。抽出したRNAのうち,PWN接種した
1・3 DPIのサンプルを供試した。RNAはAgilent 2100 Bioanalyzer(Agilent)を用いて品質を確認した。
RNAの生物学的反復(n = 3)はそれぞれ混合し,6サンプル(2時点 × 3クローン)に調整した(5
µg)。
ライブラリ調整とシーケンシングは北海道システムサイエンス社に依託した。シーケンシングは
HiSeq 2000(Illumina)を使用し,ペアエンド100 bp読み取りにより塩基配列データを取得した。得られ
たリードのうち,アダプター配列と低品質のリードをTrimmomatic software(Bolger et al., 2014)により
除去した。
1.2.4. データ解析(マツ側)
これらアセンブルされたde novoトランスクリプトームに対し,各ライブラリのフィルタリングされ
たリードをBWA(Li and Durbin, 2009)によってマッピングした。マッピングされたリード数は,
Samtools(Li et al., 2009)を用いて算出した。これらのリード数から各遺伝子でnumber of counts per
kilobase per million reads(CPKM)を算出し,平均CPKMが1以上の遺伝子を以降の解析に使用した。
主成分分析は,対数(log2)変換したデータセットに対しR version 3.5.1(R Core Team 2018)の
“prcomp” functionを用いて行った。
抵抗性クロマツと感受性クロマツ間の発現変動遺伝子(differentially expressed genes, DEGs)の検出
は,edge R package(Robinson et al., 2010)を用いて行った。リード数の正規化は,edge R package内の
Trimmed mean of M values正規化法を用いて行った。本実験ではRNA-seqに供試したサンプルに反復が
ないため,edge Rのガイドラインに従って“estimateGLMCommonDisp” functionを用いて遺伝子間の分
散を予測した。False discovery rate(FDR)の閾値は0.001に設定した。
各クローンにおける平均CPKMが10以上のDEGsについて,Blast2GO v5.2.5(Götz et al. 2008)を用
いてBlastxによる相同性検索を行った。Blastx検索は,Arabidopsis thaliana protein sequencesに対し,E-
valueの閾値を1.0E-5とすることで行った。アノテーションが付与されなかった遺伝子については,non-
redundant protein databaseに対し,Blastx検索を行った。
ベン図はVENNY 2.1(Oliveros, 2007)によって,ヒートマップはgplots package(Warnes et al., 2016)
の“heatmap.2” functionを用いて図化した。
第3章
グした。マッピングされたリード数から,Cufflinks(Trapnell et al., 2010)を用いてfragments per kilobase
of exon per million fragments mapped(FPKM)を算出した。全ライブラリでFPKMが1以上の遺伝子を以
降の解析に使用した。
PWNのエフェクター候補分子を特定するために,PWNのタンパク質配列に対してSignalP 5.0
(Almagro et al., 2019)を用いたシグナル配列の探索と,TMHMM 2.0(Krogh et al., 2001)を用いた膜貫
通領域の探索を行った。シグナル配列を含み,かつ膜貫通領域を含まない遺伝子について,Blast2GOを
用いてBlastpによる相同性検索を行った。Blastp検索は,non-redundant protein databaseに対し,E-value
の閾値を1.0E-5とすることで行った。
細胞壁分解酵素様遺伝子を特定するために,Nematode’s Cell Wall Degrading Enzyme database
(https://www.depts.ttu.edu/pss/mendulab/NCWDE/)(Rai et al., 2015)に記載されているB. xylophilusの
Gene IDを参照した。ただし,細胞壁分解酵素の作用を促進するエクスパンシンに関しては,Blastp検
索の結果をもとに特定した。
ペプチダーゼおよびペプチダーゼインヒビター様遺伝子を特定するために,MEROPSデータベースに
対してBlastpによる相同性検索を行った。E-valueの閾値は1.0E-10とした。
1.3. 結果
あった。7 DPIでは,検出されたPWNは複数個体の上部・下部において100頭を超えていた。14 DPI
では,抵抗性である波方37と三崎90では数百頭程度のPWNが検出されたが,感受性である県福岡2
号では2個体の上部・下部において1,500頭を超えるPWNが検出された。
外部病徴である主軸の下垂は,7 DPIにおいて感受性である県福岡2号の1個体で初めて認められ,
14 DPIでは県福岡2号の3個体,抵抗性である波方37の2個体,同じく抵抗性である三崎90の1個体
で認められた。
1.3.2. シーケンシングデータの概要
1・3 DPIにおけるクロマツ抵抗性2クローンと感受性1クローンのcDNAライブラリをRNA-seq解
析に供試した結果,計277,503,322リードが得られた(表3-1)。得られたリード数の範囲は,各ライブ
ラリにおいて4400万–4800万リードであった。これらのリードをフィルタリングした結果,計
214,645,032リード(Rawリードの77.4%)が得られた。以降の解析の流れと絞り込まれた遺伝子数は図
3-2に示した。
1.3.3. De novo assemblyおよびマッピング
フィルタリングされたリードをアセンブルした結果,201,317コンティグ(平均長846.8 bp,N50長
1,482 bp)が得られた(表3-2)。1ローカスあたり最も長いコンティグを遺伝子(unigene)とした結
果,110,495遺伝子(平均長672.1 bp,N50長1,224 bp)が得られた。
各クローンから得られたリードをマッピング後,各遺伝子についてCPKMを算出した結果,26,250遺
第3章
感受性である県福岡2号は,抵抗性である波方37および三崎90と第一主成分・第二主成分で区別され
た一方で,波方37と三崎90は第一主成分で区別された。
1.3.4. クロマツ抵抗性および感受性クローン間で発現差のあった遺伝子
クロマツ抵抗性および感受性クローン間におけるDEGsを明らかにするために,感受性である県福岡
2号のデータセットを抵抗性である波方37および三崎90のデータセットと比較した。その結果,1,940
DEGsが得られた(図3-4)。各組み合わせにおけるDEGの分布(M-A plot)は図3-5に示す。
PWNへの抵抗性あるいは感受性に関連すると考えられる候補遺伝子を特定するために,ある一定値
以上(発現上昇したクローンで平均CPKM ≥ 10)の発現量が認められた400 DEGsに絞り込んだ。これ
らのDEGsを用いてヒートマップを作成した結果,各クローンで1・3 DPIの発現パターンは類似する傾
向にあった(図3-6)。ベン図を作成した結果,感受性と比較して抵抗性で発現上昇していた遺伝子に
ついては,波方37および三崎90ではそれぞれ113 DEGsと51 DEGsが特異的であり,37 DEGsは両ク
ローンで共通していた(図3-7)。抵抗性と比較して感受性で発現上昇していた遺伝子については,波
方37および三崎90ではそれぞれ80 DEGsと56 DEGsが特異的であり,60 DEGsは両クローンで共通し
ていた。
400 DEGsをBlastx検索に供試した結果,181 DEGsがArabidopsis thaliana protein sequencesに対して,
34 DEGsが他の植物のprotein sequencesに対してアノテーションが付与された。これらのDEGsを機能
れた(表3-3,図3-8)。LRR-RLK関連遺伝子については,全てのクローンで複数の遺伝子が発現上昇
していたが,感受性である県福岡2号で発現上昇した遺伝子は多い傾向にあった(図3-8)。NBS-LRR
関連遺伝子についても,全てのクローンで複数の遺伝子が発現上昇していたが,抵抗性である波方37
において発現上昇した遺伝子は多い傾向にあった。波方37で発現上昇したDEGsのうち,1 DEG
(TRINITY_DN42529_c0_g1_i1)がCC-NBS-LRRクラス,4 DEGs(TRINITY_DN44841_c0_g1_i7,
TRINITY_DN46717_c0_g3_i8,TRINITY_DN46717_c0_g4_i1,TRINITY_DN47008_c3_g1_i12)がTIR-
NBS-LRRクラスに分類された。
細胞壁の形成と生合成に関連するアノテーションが付与されたDEGsのうち,1 DEGがcellulose
synthase family protein,3 DEGsがlaccase遺伝子群,2 DEGsがxyloglucan endotransglucosylase/hydrolase
(XTH)遺伝子群,3 DEGsがHXXXD-type acyl-transferase family protein,1 DEGがglycosyl hydrolase
family protein,3 DEGsがpectin lyase-like superfamily proteinに分類された(表3-4,図3-8)。これらの
DEGsのうち,波方37では,cellulose synthase family protein,laccase遺伝子群,XTH 9
(TRINITY_DN38633_c0_g1_i1)が発現上昇する傾向にあった。三崎90では,laccase遺伝子群,
HXXXD-type acyl-transferase family protein,XTH 32(TRINITY_DN43228_c0_g3_i2)が発現上昇する傾向
にあった(図3-8)。一方で,glycosyl hydrolase family proteinとpectin lyase-like superfamily proteinは,三
崎90と県福岡2号で発現上昇する傾向にあった。
抗菌タンパク質に関連するアノテーションが付与されたDEGsのうち,12 DEGsがPR proteins,2
第3章
1.3.5. クロマツ抵抗性および感受性クローン共通で高発現していた遺伝子
DEGsでない遺伝子のうち,CPKMが1,000以上の遺伝子を抽出した結果,104遺伝子となった。これ
らの遺伝子のうち,21遺伝子がPR proteinに分類された(表3-6)。
1.3.6. マツノザイセンチュウゲノムへのマッピングとエフェクター候補遺伝子の樹体内における発現
各ライブラリにおいてフィルタリングされたリードをB. xylophilusリファレンスゲノムにマッピング
した結果,平均0.237%のリードがマッピングされた(表3-1)。
全ライブラリでFPKMが1以上の遺伝子を抽出した結果,937遺伝子となった。それらのうち,
SignalPによってシグナル配列を含むことが予測され,かつTMHMMによって膜貫通領域を含まないこ
とが予測された遺伝子を抽出した結果,125遺伝子となった。これらの遺伝子をBlastp検索に供試した
結果,51遺伝子が植物の細胞壁分解,ペプチダーゼとペプチダーゼインヒビター,抗酸化や植物の防御
反応の操作と関連すると考えられる遺伝子群に分類された。
植物の細胞壁分解に関与する14遺伝子のうち,1遺伝子がglycoside hydrolases(GH)25,1遺伝子が
GH27,3遺伝子がGH45(cellulase),8遺伝子がpolysaccharide lyases(pectate lyase),1遺伝子が
expansinのファミリーに分類された(表3-7)。
ペプチダーゼとペプチダーゼインヒビターに関連するアノテーションが付与された25遺伝子のう
ち,22遺伝子がペプチダーゼ(9 Cysteine,7 Serine,3 Metallo,2 Aspartic,1 Unknown)に分類され,3
dependent monooxygenase domain proteinに分類された(表3-9)。1遺伝子は,アノテーションは付与さ
れなかったが,先行研究(Hu et al., 2019)でベンサミアナタバコの細胞死を引き起こし,PWNの病原
力に寄与することが示唆されたBxSapB1(BUX.s00139.62)と一致していた。
1.4. 考察
1.4.1. マツ材線虫病におけるdual RNA-seq解析の有効性
網羅的遺伝子発現解析を利用してマツ材線虫病における生体分子レベルの情報を取得する試みは,宿
主であるマツ(Shin et al., 2009; dos Santos and de Vasconcelos, 2011; Nose and Shiraishi, 2011; Hirao et al.,
2012; Santos et al., 2012; Xu et al., 2013; Gaspar et al., 2017; Liu et al., 2017; Lee et al., 2019; Xie et al.,
2020),もしくは宿主から分離した病原体であるPWN(Qiu et al., 2013; Espada et al., 2016; Tsai et al.,
2016; Hu et al., 2019)のいずれかを対象として実施されてきた。特に,PWNを対象とした網羅的遺伝子
発現解析では,宿主から数時間かけて分離したPWNが供試されているため,樹体内における病原力が
直接的に反映されているとは言い難い。本研究では,感染初期におけるクロマツ抵抗性2クローンおよ
び感受性1クローンを対象としてdual RNA-seq解析を行うことで,直接的に獲得された宿主と病原体双
方の遺伝子発現情報から,PWNに対するクロマツの抵抗性もしくは感受性に関連する要因を特定する
ことを目的とした。
本研究では,RNA-seqによって獲得された全リードをde novo assemblyに供試し,マッピングの結果
第3章
て獲得された全リードのうちB. xylophilusリファレンスゲノムにマッピングされたリードについて,全
てのライブラリで発現が認められ,かつ分泌タンパク質をコードしていると考えられる遺伝子を抽出し
た結果,125遺伝子となった。Blast検索の結果,これらの遺伝子は,先行研究でPWNの感染時に発現
が認められている,植物の細胞壁分解,ペプチダーゼとペプチダーゼインヒビター,抗酸化や植物の防
御反応の操作と関連するファミリーに分類された。さらに,これらの遺伝子はPWNのプロテオーム解
析(Shinya et al., 2013a)によって報告された分泌タンパク質の多くと一致しており,一部報告されてい
ない遺伝子も認められた。したがって,マツ材線虫病にdual RNA-seq解析を適用することによって,宿
主と病原体のトランスクリプトームを同時に獲得し,それぞれの発現遺伝子を明らかにできることが示
された。
本研究では,1・3 DPIのマツ木片から抽出したRNAをシーケンシングに供試し,得られたリードを
B. xylophilusリファレンスゲノムにマッピングした結果,平均0.237%のリードがマッピングされた(表
3-1)。通常の発現レベルのmRNAを解析対象とする場合,発現量を算出する場合には最低2,000万リ
ードが必要であると考えられている(今村・鈴木,2016)が,本研究では,PWN由来と考えられるリ
ード数は平均105,151リードであった。病原菌Phytophthora ramorumに感染したブナ科の樹木であるタ
ンオーク(Notholithocarpus densiflorus)を対象としてdual RNA-seq解析をした場合,1 DPIでは0.1%の
リードがP. ramorumのゲノムにマッピングされたのに対し,5 DPIでは1.1%のリードがマッピングされ
(Shukla et al., 2018)。したがって,dual RNA-seq解析は宿主と病原体の発現遺伝子を網羅的に取得す
ることが可能であるが,感染初期を解析対象とした場合,病原体由来のRNA量は宿主由来と比較して
極めて少なくなると考えられる。そのため,dual RNA-seq解析を行う際には,シークエンス深度やサン
プリング法を考慮して実施する必要があると考えられる。
1.4.2. クロマツの病原体認識
植物が病原体に感染した際,植物は細胞膜内のパターン認識受容体(pattern recognition receptors,
PRR)によって病原体の構成成分である病原体関連分子パターン(pathogen-associated molecular patterns,
PAMPs),あるいは宿主が傷害を受けた場合に放出される損傷関連分子パターン(damage-associated
molecular patterns, DAMPs)を検出し,PAMPs誘導免疫と呼ばれる防御反応を開始する(Malinovsky et
al., 2014; Nejat et al., 2016; Andersen et al., 2018; Ramirez-Prado et al., 2018)。PRRは,受容体キナーゼも
しくは受容体様タンパク質であり,受容体キナーゼタイプのPRRはleucine-rich repeat(LRR)などのリ
ガンド認識ドメインを含む(Zipfel, 2014)。病原体はこのPAMPs誘導免疫の抑制や,その他感染に有
利な働きをもつエフェクターを植物細胞内に送り込むことによって感染を成立させようとする。それに
対し,植物はこれらのエフェクターをRタンパク質と呼ばれる細胞内受容体によって認識し,PAMPs
誘導免疫よりもさらに強力なエフェクター誘導免疫が誘導される(Malinovsky et al., 2014; Nejat et al.,
2016; Andersen et al., 2018; Ramirez-Prado et al., 2018)。多くのRタンパク質は,ヌクレオチド結合部位
(nucleotide-binding site, NBS)とLRRを有しており,N末端部分にToll-likeドメインを含むTIR-NBS-
第3章
本研究では,クロマツ抵抗性および感受性クローン間でDEGsとして検出された遺伝子のうち,
PAMPs誘導免疫に関連すると考えられるRLK,PK,LRR-RLKをコードする遺伝子群と,エフェクタ
ー誘導免疫に関連するNBS-LRRをコードする遺伝子群について,全てのクローンで複数の遺伝子の発
現が認められた(図3-8)。特に,LRR-RLKに関連する遺伝子群は感受性である県福岡2号において,
NBS-LRRに関連する遺伝子群は抵抗性である波方37において発現する傾向にあった。マツ材線虫病に
おいて,病原体に対する植物の防御反応の最も上流に位置する病原体認識に関連する遺伝子発現につい
ての情報は,本研究によって初めて獲得された。この結果は,病原体認識に関する遺伝子発現が抵抗性
個体と感受性個体で異なることで,下流の防御反応に両者間で差異が生じる可能性を示唆するものであ
る。NBS-LRR遺伝子は植物ゲノム上でファミリーを形成し,これまでにパパイアやキュウリの約50か
らイネの653までNBS-LRR遺伝子をコードする配列が多数特定されている(Marone et al., 2013)。ネ
コブセンチュウやシストセンチュウに対する抵抗性植物では,耐性を付与するR遺伝子の多くはNBS-
LRR遺伝子であることが明らかにされており,それらのうちジャガイモ(Solanum tuberosum)のGro1-4
とプルーン(Prunus domestica)のMaはTIR-NBS-LRRをコードし,トマトのMi-1.2,Mi-9,Hero-Aな
どはCC-NBS-LRRをコードしている(Sato et al., 2019)。他の植物で抵抗性遺伝子とされているNBS-
LRR遺伝子群が1つのクロマツ抵抗性クローンで発現上昇する傾向にあることは,本研究によって初め
て明らかにされ,興味深い結果といえる。クロマツにおいても,病原体の認識に関わるこれらNBS-