2 試料と方法
2013年 3月11日OB-B地点 (図 1) で採取したサンプ ルからA. miii を顕微鏡下で拾いだし,2013 年 3 月 12 日 から4月13日まで飼育を行った。飼育容器には12ウェ ル マ ル チ プ レ ー ト (1 つ の 穴 の 直 径 が25mm, 深 さ1 はじめに
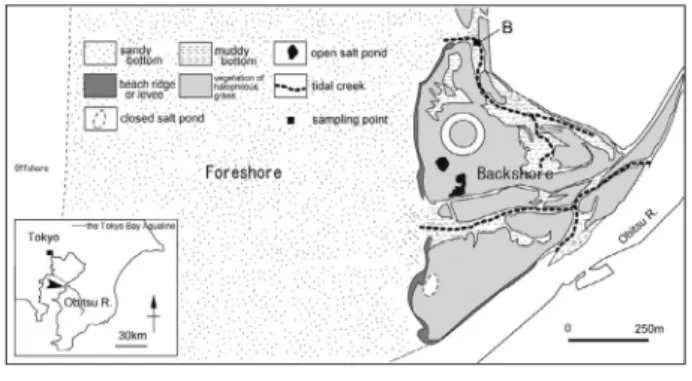
Angulicytherura miii は日本の汽水域に生息するポド コーパ目の貝形虫類 (甲殻亜門) である。その個体群動 態は小櫃川の河口に形成された干潟(盤州干潟)におい て明らかにされており,水温が低くなる12月に現れ, 水温が上昇する4月に個体数を減少させ,夏季は完全に 姿を消すことが報告されている (中尾・塚越,2000,2008; 中尾,2010)。後背湿地と砂質前浜干潟の境界付近の砂 泥底に現れるが (図 1),その生息場所は数年を経過して も大きく移動することはない (Nakao & Tsukagoshi, 2002;中尾・塚越,2000,2008)。これまで,本種の夏越しにつ いての情報はまったく得られておらず,堆積物が不安定 な干潟上で,なぜ毎年同じ場所に現れる事ができるのか という謎が残されていた。本研究では飼育実験下での観 察を行い,A. miii の生活史を明らかにする事を目的と する。
中尾 有利子
In this study, the life history of Angulicytherura miii (Ishizaki, 1969) was clarified under culture conditions, and eggs deposited on sediment particles were found for the first time. The species, which produce single generation per year, sur-vive summer as “oversummering” eggs. The eggs were deposited in tiny dents on the surface of only mineral particles that had a slightly depressed form. No eggs were founded on any particle that had smooth surface. The adhesion of the egg to the particle was strong enough to prevent the egg from being detached from the particles. On the contrary, eggs of Loxoconcha kosugii, sympatric species, were almost spherical and were deposited on the surface of not only mineral parti-cles but also organic partiparti-cles, including carapace of other individuals. The adhesion of the egg to the partiparti-cles was weak. These observations revealed that A. miii selected mineral particles, which have tiny dents on the surface and range from fine sand to coarse sand particles, as ovipositional substances in order to remain in their habitat in the estuary until next winter. This behavior is considered a seasonal adaptation and caused A. miii to be considered a winter-ephemeral species and migrate the temperate zone from high latitude area in the Pacific.
Keywords: Ostracoda, brackish-water, ovipositional substances, seasonal adaptation, Oversummering
Angulicytherura miii
(甲殻亜門:貝形虫綱)
にみられる
夏越しの戦略としての産卵基質の選択
The Selection of Ovipositional Substances for Oversummering of
Angulicytherura miii (Crustacea: Ostracoda)
Yuriko NAKAO
*(Received November 16, 2013)
Department of Geosystem Sciences, Nihon University: 3-25-40 Sakurajousui, Setagaya-ku, Tokyo 156-8550, Japan 日本大学文理学部地球システム科学科:
〒156-8550 東京都世田谷区桜上水3-25-40
20mm)を使用し 1 週間おきに新しい容器に交換した。 飼育には塩分約17‰に調整した人工海水を用いて 3,4 日おきに交換し,保冷庫で水温を5 ℃から 7 ℃に保っ た。堆積物 (敷石) として,実験個体の採取地点である OB-B地点で採取された堆積物粒子,同地点付近で採取 されたより粗めの粒子を含む堆積物粒子,熱帯魚飼育用 の底砂 (以降リーフサンドとする),ガラスビーズの 4 種類を底面に敷いた。試料採取地点とその付近から得ら れた堆積物粒子は飼育で使用した塩水で洗浄し,顕微鏡 下でデトリタスを取り除いてから使用した。餌は OB-B 地点で採取されたデトリタスを用いた。堆積物の種類, デトリタスの有無によって飼育条件Aから飼育条件Hの 8 つの条件を用意した (表 1,図 2)。飼育個体はOB-B 地点で採取し,それぞれの飼育条件につき雄20個体, 雌20個体の計40個体を用意した。飼育個体が死んだ場 合は取り除き,新しい個体の補充は行わなかった。飼育 中は,生殖行動と堆積物粒子の観察を毎日行った。全飼 育個体の死亡後,顕微鏡下で堆積物を確認し,卵の有 無,堆積物粒子1 粒あたりに産み付けられた卵の数,す べての堆積物粒子の大きさを計測した。粒子の大きさ は,顕微鏡写真を撮影し,PCに取り込んだ画像を用い て,フリーウェアソフトのimage-Jを使用して,各粒子 の外接長方形で計測した。数値は小数第一位を四捨五入 して整数で示した。また,自然状態との比較のため, OB-B地点で採取した堆積物および Loxoconcha kosugiiの 行動を顕微鏡下で観察した。
3 結果
3-1 産卵基質と卵の形 Angulicytherura miii の卵が初めて確認された (図 3 )。 確認された産卵基質はすべて鉱物粒子で,植物片や生物 の遺骸など有機物粒子に産下された卵は確認されなかっ た。卵は球形ではなく,鉱物粒子表面の僅かなくぼみに はめ込むように産み付けられているため,上側の丸みは 弱く平らで,輪郭はくぼみに沿う形を呈していた。ま た,卵同士が隣接する場合は,お互いに隙間をうめるよ うな相補的な輪郭をとり,産卵基質に押しつけたような 形を呈していた。卵と産卵基質との固着は強いと考えら れ,産卵基質から外れて単体で存在する卵は観察されな かった。これらの事は,野外で採取された堆積物でも確 表1 設定された8 つの飼育条件 飼育条件 堆積物(敷石) デトリタス 飼育個体数 塩濃度(‰) 水温(℃) A OB−B から採取(細) 無 ♂20,♀20 16−18 5−7 B OB−B の周辺から採取(粗) 無 ♂20,♀20 16−18 5−7 C リーフサンド 無 ♂20,♀20 16−18 5−7 D ガラスビーズ 無 ♂20,♀20 16−18 5−7 E OB−B から採取(細) 有 ♂20,♀20 16−18 5−7 F OB−B の周辺から採取(粗) 有 ♂20,♀20 16−18 5−7 G リーフサンド 有 ♂20,♀20 16−18 5−7 H ガラスビーズ 有 ♂20,♀20 16−18 5−7 図2 設定された8 つの飼育条件の写真。EからHには餌としてデトリタスを入れた。スケールバーは 1mm。認された。 野外で採取された堆積物から,A. miii と同所的に生 息するLoxoconcha kosugii のものと思われる卵が確認さ れた (図 4)。L. kosugiiの卵は歪みのない球形で,鉱物粒 子だけでなく有機物粒子や生きている貝形虫類の背甲も 産卵基質として利用されていた。また,産卵基質に付着 せず,単体で存在する卵も観察され,卵と産卵基質の固 着が弱く簡単にはなれてしまう事が明らかにされた。実 験室内での数日間の飼育と観察の結果,卵はL. kosugii のものである事,また,産下後1日から長くとも3日の 間に孵化する事が確認された。 3-2 デトリタスと卵数 表1 で示したAからHの各飼育環境において,堆積物 粒子の数と卵が産み付けられた堆積物粒子の数,卵の数 図3 鉱物粒子に産下されたAngulicytherura miii の卵。1 ,3:鉱物粒子表面の凹みに産み付けられた卵。2:互いに隙間をうめ るような形で密着して鉱物粒子に産み付けられた卵。1 の矢印,4:鉱物粒子に押しつけられたような形を呈する卵。出っ 張らず平になっているのがわかる。 図4 堆積物中に見いだされたLoxoconcha kosugii の卵。1: ほぼ球形で,細かい堆積物粒子にも産み付けられてい る卵。2:基質からはずれた卵。産卵基質との固着が 弱いことが分かる。3:卵から孵化したばかりの幼体 (矢印)。4:幼体の背甲に産み付けられた卵。
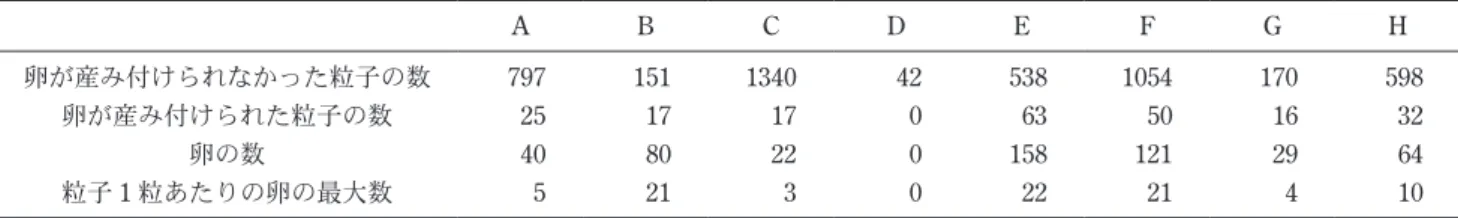
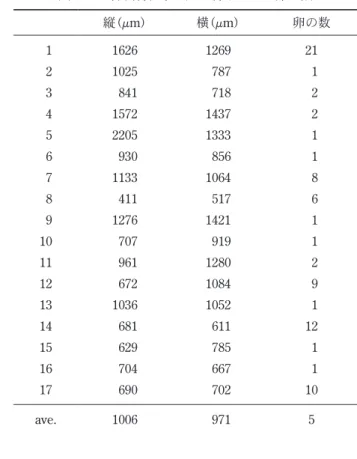
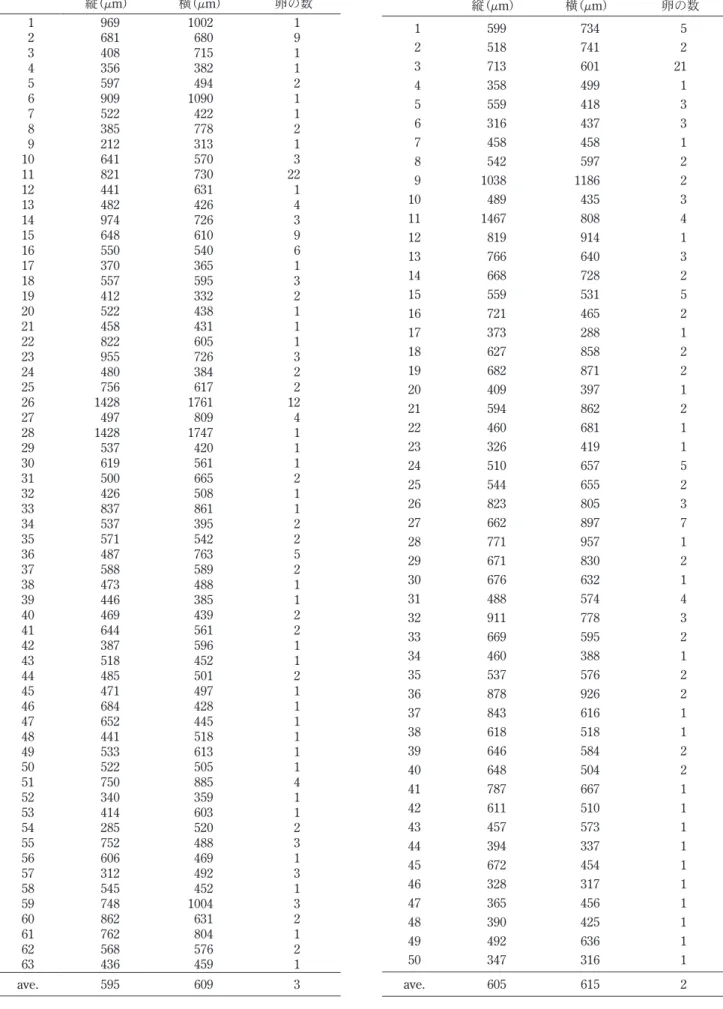
表4 から表10に示す。飼育条件E,G,Hにおいて,一辺 の長さが250 μm以下の産卵基質が数個あったものの, 他の産卵基質はすべて二辺の長さがともに250 μm以上 であった。 表面形態に着目すると,産卵基質に利用された鉱物粒 子の表面はすべて凹凸があった。反対に,表面の形態が なめらかで凹凸がないガラスビーズに産み付けられた卵 は全く確認されなかった。飼育条件Hで確認された卵 は,すべてデトリタスに含まれている鉱物粒子に産み付 けられたものであった。飼育条件Dにおいても,交尾 行動は観察されたので,適当な産卵基質がなかったため に産卵せずに死亡した事になる。A. miii は表面に凹凸 を表2 に示す。産み付けられた卵の数は,それぞれ飼育 条 件Aで は40個,Bで は80個,Cで は22個,Dで は 0 個,Eでは158個,Fでは121個,Gでは29個,Hでは64 個であった。同じ堆積物の条件で比較すると,デトリタ スを入れなかった飼育条件AからDより,入れた飼育条 件EからHの産下卵数が多いという結果が得られた。 3-3 産卵基質の大きさと形状 飼育実験に用いられた堆積物粒子のサイズとA. miii に産卵基質として利用された鉱物粒子の大きさに関する データを表3 に,産卵基質として利用された鉱物粒子の 大きさとそれぞれの鉱物粒子に産み付けられた卵の数を 表2 8 つの各飼育条件で使用された堆積物粒子と卵の数 A B C D E F G H 卵が産み付けられなかった粒子の数 797 151 1340 42 538 1054 170 598 卵が産み付けられた粒子の数 25 17 17 0 63 50 16 32 卵の数 40 80 22 0 158 121 29 64 粒子1 粒あたりの卵の最大数 5 21 3 0 22 21 4 10 表3 堆積物粒子全体のサイズデータと卵を産み付けられた粒子のサイズデータ A B C D 堆積物粒子全体 縦 横 平均 縦 横 平均 縦 横 平均 縦 横 平均 標本数 822 822 822 168 168 168 1357 1357 1357 42 42 1185 最大値 1358 1217 1263 3049 2430 2739 1089 1176 1016 1171 1199 856 最小値 7 13 10 8 14 14 8 10 12 863 846 1017 平均値 440 436 438 683 669 676 407 409 408 1015 1019 1010 中央値 420 417 422 686 716 708 385 388 392 1006 1010 5167 標準偏差 212 203 199 469 406 427 135 136 125 71 74 72 産卵基質 標本数 25 25 25 17 17 17 17 17 17 0 0 0 最大値 802 879 824 2205 1437 1769 1058 705 792 − − − 最小値 335 403 374 411 517 464 295 260 282 − − − 平均値 536 598 567 1006 971 988 465 472 469 − − − 中央値 497 603 586 930 919 893 394 439 404 − − − 標準偏差 127 144 124 451 297 354 200 138 137 − − − E F G H 堆積物粒子全体 縦 横 平均 縦 横 平均 縦 横 平均 縦 横 平均 標本数 601 601 601 1104 1104 1104 186 186 186 630 630 630 最大値 2637 1825 2201 2282 3154 2647 855 831 785 1195 1188 1191 最小値 10 17 13 14 8 11 25 31 28 52 59 60 平均値 438 427 432 404 396 400 354 348 351 345 339 342 中央値 417 416 420 319 318 320 332 339 335 235 227 232 標準偏差 238 225 224 298 298 289 166 162 157 299 296 296 産卵基質 標本数 63 63 63 50 50 50 16 16 16 32 32 32 最大値 1428 1761 1594 1467 1186 1137 666 720 630 894 636 752 最小値 212 313 262 316 288 323 297 227 272 168 208 218 平均値 595 609 602 605 615 610 462 455 459 399 393 396 中央値 537 542 548 597 596 580 469 454 451 386 387 372 標準偏差 229 268 237 210 197 186 109 115 97 139 115 119 縦,横は外接長方形の2辺をしめし,平均は縦と横の値の平均。最大値,最小値,平均値,中央値の単位はμm。
表4 飼育条件Aで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 584 626 5 2 765 654 5 3 428 466 1 4 393 455 3 5 542 606 1 6 412 497 1 7 679 520 1 8 512 607 2 9 483 688 2 10 614 609 1 11 424 497 1 12 444 403 1 13 778 761 1 14 802 846 3 15 611 745 1 16 335 414 1 17 772 760 2 18 516 879 1 19 351 547 1 20 431 754 1 21 462 449 1 22 329 400 1 23 391 492 1 24 472 584 1 25 433 484 1 ave. 519 590 2 表5 飼育条件Bで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 1626 1269 21 2 1025 787 1 3 841 718 2 4 1572 1437 2 5 2205 1333 1 6 930 856 1 7 1133 1064 8 8 411 517 6 9 1276 1421 1 10 707 919 1 11 961 1280 2 12 672 1084 9 13 1036 1052 1 14 681 611 12 15 629 785 1 16 704 667 1 17 690 702 10 ave. 1006 971 5 表6 飼育条件Cで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 515 498 1 2 657 469 1 3 735 454 1 4 305 260 1 5 394 705 1 6 561 690 1 7 364 644 3 8 332 329 2 9 301 397 1 10 364 369 1 11 338 322 1 12 373 349 1 13 370 362 1 14 401 650 1 15 1058 525 2 16 295 456 2 17 542 550 1 ave. 465 472 1
表7 飼育条件Eで産卵基質として利用された鉱物粒子の 大きさ鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 969 1002 1 2 681 680 9 3 408 715 1 4 356 382 1 5 597 494 2 6 909 1090 1 7 522 422 1 8 385 778 2 9 212 313 1 10 641 570 3 11 821 730 22 12 441 631 1 13 482 426 4 14 974 726 3 15 648 610 9 16 550 540 6 17 370 365 1 18 557 595 3 19 412 332 2 20 522 438 1 21 458 431 1 22 822 605 1 23 955 726 3 24 480 384 2 25 756 617 2 26 1428 1761 12 27 497 809 4 28 1428 1747 1 29 537 420 1 30 619 561 1 31 500 665 2 32 426 508 1 33 837 861 1 34 537 395 2 35 571 542 2 36 487 763 5 37 588 589 2 38 473 488 1 39 446 385 1 40 469 439 2 41 644 561 2 42 387 596 1 43 518 452 1 44 485 501 2 45 471 497 1 46 684 428 1 47 652 445 1 48 441 518 1 49 533 613 1 50 522 505 1 51 750 885 4 52 340 359 1 53 414 603 1 54 285 520 2 55 752 488 3 56 606 469 1 57 312 492 3 58 545 452 1 59 748 1004 3 60 862 631 2 61 762 804 1 62 568 576 2 63 436 459 1 ave. 595 609 3 表8 飼育条件Fで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 599 734 5 2 518 741 2 3 713 601 21 4 358 499 1 5 559 418 3 6 316 437 3 7 458 458 1 8 542 597 2 9 1038 1186 2 10 489 435 3 11 1467 808 4 12 819 914 1 13 766 640 3 14 668 728 2 15 559 531 5 16 721 465 2 17 373 288 1 18 627 858 2 19 682 871 2 20 409 397 1 21 594 862 2 22 460 681 1 23 326 419 1 24 510 657 5 25 544 655 2 26 823 805 3 27 662 897 7 28 771 957 1 29 671 830 2 30 676 632 1 31 488 574 4 32 911 778 3 33 669 595 2 34 460 388 1 35 537 576 2 36 878 926 2 37 843 616 1 38 618 518 1 39 646 584 2 40 648 504 2 41 787 667 1 42 611 510 1 43 457 573 1 44 394 337 1 45 672 454 1 46 328 317 1 47 365 456 1 48 390 425 1 49 492 636 1 50 347 316 1 ave. 605 615 2
がある粒子を産卵基質として選択し,なめらかな粒子に は産下しないことが示された。
4 考察
今回,Angulicytherura miii の卵が初めて観察され,成 体個体が生殖後に死滅したことから,本種が卵態で冬越 しならぬ 夏越し をしている事が明らかにされた。これ まで,卵態で冬越しをする種は報告されているが (例:Cytherois fischeri : Horne, 1983),夏越しをする種の報告
は初めてとなる。A. miii の近縁種は日本海の大陸沿岸 やアラスカに多く分布し,本種は寒冷な北太平洋に起源 をもつと考えられており (中尾・塚越,2000),卵態での 夏越しは,本来寒冷な環境に起源をもつ種による,温暖 な環境に対する適応の一つであると考える事ができる。 トガリアメンボの分布域の拡大に見られるように,休眠 卵を持つことが,時として受動的分散をもたらし,その 分類群の分布域拡大に大きい役割をもつことがある (中 尾他,2010)。本種は,産卵基質の受動的分散と卵態夏越 しという季節適応の結果,分布範囲を温帯域まで拡大す る事が可能になったと考えられる。 Athersuch et al. (1989) によると,卵態で冬越しをす るCytherois fischeri は,春に越冬した卵が孵化し, これを 第1 世代とすると,春から夏にかけて第 1 世代が成長し て第2 世代となる卵を産下し,第 2 世代は直後に孵化, 成長して第3 世代となる卵を産下する。越冬をするのは この第3 世代か,夏から秋の間に成長をした第 3 世代が 産下した第4 世代にあたる卵で,一年の間に少なくとも 3 世代が入れ替わる。これに対し,今回の飼育実験で産 下された卵はA. miii の出現期間がおわる 4 月までの間に 孵化しなかった。これにより,A. miii は第 1 世代が卵 下した第2 世代が夏越しをするという 1 年 1 世代の生活 史を持つ事が明らかになった。第1 世代が死滅する 4月 から第2 世代が現れる12月の間を卵態で過ごすとする と,卵期間は少なくとも8ヶ月あると考えられる。海域 から汽水域に生息する貝形虫類では耐性卵を持たないの が一般的で,Elofsonia baltica のみ耐性卵をもつことが
知られている (Athersuch et al., 1989)。Theisen (1966)
によると,E. baltica の耐性卵は,低温 (凍結を含む), 高 塩分,低塩分,低酸素に耐える。また,淡水生の貝形虫 類には,30年乾燥に耐えた卵も報告されている(Kesling, 1951)。今回発見されたA. miiiの卵が耐性卵なのか,卵 期間が数年に及ぶものがあるのかは今回の実験では明ら かにできなかった。いずれにせよ,産卵基質の流出・運 搬は生物の分布拡大に果たす役割が大きいと考えられる ため,本種の卵態夏越しの方法は貝形虫類の分布拡大, 表9 飼育条件Gで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 325 478 1 2 428 350 4 3 436 382 1 4 533 421 1 5 318 227 1 6 599 505 1 7 540 720 3 8 411 372 3 9 529 520 1 10 319 351 1 11 567 488 3 12 666 588 2 13 297 565 2 14 462 474 1 15 487 408 2 16 477 434 2 ave. 462 455 2 表10 飼育条件Hで産卵基質として利用された鉱物粒子の 大きさと各鉱物粒子に産み付けられた卵の数 縦(μm) 横(μm) 卵の数 1 315 347 1 2 336 345 1 3 435 291 1 4 420 404 5 5 379 429 1 6 448 408 2 7 301 389 3 8 393 385 2 9 317 338 2 10 676 636 3 11 523 566 2 12 534 544 1 13 399 523 2 14 306 312 1 15 326 246 1 16 337 236 1 17 291 388 1 18 544 463 4 19 485 355 3 20 324 258 1 21 894 609 3 22 520 502 10 23 168 268 1 24 398 471 2 25 212 418 1 26 443 439 1 27 406 558 2 28 350 315 2 29 397 364 1 30 234 220 1 31 290 208 1 32 362 325 1 ave. 399 393 2
ビーズは産卵基質として選択されない。反対に,L. kosugii で観察された卵と基質の固着の弱さは,卵が産下後数日 の内に孵化をするので卵が基質から離れたとしても生息 範囲外への流出の可能性が低いため大きな問題とならな いということで説明される。また,L. kosugii は産卵基 質の選択を行わない事も同じ理由と考えられる。 様々な形態をもつ堆積物粒子の粒径は容易に扱う事が できないため,今回は外接長方形の縦と横の長さの平均 値をその堆積物粒子の粒径として代表させた。A. miii の産卵基質となった鉱物粒子は,ほぼ中粒砂から粗粒砂 であることが示され,底質の粒度分布に関わらず,A. miii が産卵基質として,中粒砂から粗粒砂の大きさをもつ粒 子を選択しているといえる (図 5,図 6)。「ルーべーの 種分化を考える上でも重要になる。 次に産卵基質に着目する。A. miii は産卵基質として 表面に卵がはめ込めるような凹凸がある鉱物粒子を選択 していた。鉱物粒子の選択は,基質そのものが時間経過 により分解し,卵が基質から外れる事をふせぐためと考 えられる。また,Loxoconcha kosugii でみられた産卵基 質との固着が弱い球形の卵と比較すると,A. miii の卵 はとなりあう卵が密着するような形になり,産卵基質に しっかりと固着されていることが分かる。基質表面の凹 凸を利用するとともに,卵の固着をより強固なものに し,孵化時期 (次の低温期) までの約 8ヶ月の間,卵が 堆積物粒子と共に生息範囲にとどまる事に成功させてい ると考えられる。そのため,表面がなめらかなガラス 0 500 1000 1500 2000 2500 3000 3500 0 500 1000 1500 2000 2500 3000 3500 0 500 1000 1500 2000 2500 3000 3500 0 500 1000 1500 2000 2500 3000 3500
a
b
図5 飼育に用いられた堆積物粒子の大きさ (a) と産卵基質として選択された鉱物粒子の大きさ (b)。X軸は表 4 から表10の横の 長さ,Y軸は縦の長さを示す。単位は μm。Angulicytherura miiiが250 μm以下の鉱物粒子を産卵基質として選択しないこ とがわかる。図6 8 つの飼育条件で使用された堆積物粒子のサイズ分布と,各階級に産下された卵の個数。粒子の大きさは,外接長方形の
観察が必要とされる。
5 まとめ
A. miii は卵態で夏越しをしている事が初めて確認さ れた。本種は,生息場所からの卵の流失を防ぐため,一 定の条件を満たす堆積物粒子を産卵基質として選択して いる事が明らかにされた。すなわち,1 . 表面に凹凸の ある,2 . 中粒砂から粗粒砂の大きさをもつ,3 . 鉱物粒 子を選択している事がわかった。またこの性質をもつ A. miii の分布は,堆積物の粒径に左右される。一方, A. miii のように卵態で長期間をすごす生物では,能動 的分散よりも,産卵基質の移動といった受動的分散がそ の生物の分布の拡大に大きな役割をはたすと考えられ る。 本種の他地点での生活史,近縁種の分布と生活史 の解明が適応放散の研究に新しい知見を示すであろう。 謝辞 本研究をすすめるにあたり,静岡大学理学部地球科学科の 塚越哲教授には粗稿を読んで頂き,有益な助言,議論をして 頂いた。日本大学文理学部地球システム科学科の遠藤邦彦名 誉教授には研究環境を長年にわたり与えて頂き,様々なご助 力を頂いた。また,日本大学生物資源科学部の小沢広和助教 には様々な助言をして頂いた。その他大勢の方々のご協力に 感謝し,お礼を申し上げる。 本研究の費用の一部に平成24年度日本大学文理学部個人 研究費(研究題目:貝形虫類の温度指標としての有効性の検 証)を使用した。 沈降曲線」 (Rubey, 1933) から,一般に水中では中粒砂 より大きい粒子は沈降速度が速くなることが知られてい る。A. miii は生息地点の堆積物粒子の中で相対的に沈 降速度が早い粒径の鉱物粒子を産卵基質として選択して おり,それによって生息場所からの卵の流失の可能性を 低めていると考えられる。 貝形虫類の分布を制限する要因としては,塩分,水 温,有機物量,堆積物の粒度など様々なものが挙げられ る。これまでの調査からA. miii は後背湿地と砂質前浜 干潟の境界付近にのみ生息していることが明らかにされ てきたが,その分布を制限する要因については明らかで はなかった。盤州干潟の底質の中央粒径は190 μm程度 と報告されており (内山,2005),A. miiiが産卵基質とし て選択するには小さすぎる。これに対し, A. miiiの生息 地点の底質の粒径の中央値はそれより大きく (表 3), 産卵基質として選択できる較的大きな堆積物粒子が分布 していることがわかる。これは,産卵基質を選択し,卵 態で夏越しをするという生活史をもつA. miii の分布 は,底質の粒径に大きく制限される事を意味する。 また,今回の観察では,1 個体あたりの産卵数は明ら かにされなかった。しかしながら,同じ堆積物粒子の条 件ではデトリタスをいれた方が産み付けられた卵が多 かったことから,A. miii の栄養源がデトリタスにある 可能性が示された一方で,1 個体の産卵数は成体の栄養 条件によって変化する可能性が残され,結論には一層のAthersuch, J., Horne, D. and Whittaker, E. (1989) : Marine and Brackish water ostracods. In: Kermack, D. M. and Barnes, R. S. K. (eds.) Synopses of the British Fauna (New Se-ries). E. J. Brill, 343pp.
Horne, D. J. (1983) : Life-cycles of Podocopid Ostracoda –a re-view (with particular reference to marine and brackish water species). In: Maddocks, R. F. (ed.), Application of Ostracoda. Dept of Geosciences, Univ. of Houston, p.581-590.
Kesling, R. V. (1951) : The morphology of ostracod molt stages. Illinois Biological Monographs, 21, 1-114.
中尾史朗・前田輝久・山尾あゆみ(2010).日本に定着した トガリアメンボの周年経過と卵休眠.京都府立大学学術 報告「生命科学」,62,1-12. 中 尾 有 利 子(2010): 小 櫃 川 河 口 干 潟 に お け る Angulicy-therura miii(貝形虫,甲殻類)の個体群動態.日本大 学部文理学部自然科学研究所研究紀要,45,133-141. 引用文献 中尾有利子・塚越 哲(2000):貝形虫類の群集組成とその 季節変化―小櫃川河口干潟を例として―.池谷仙之・棚 部一成(編),古生物の科学3 古生物の生活史,朝倉 書店,p.239-245.
Nakao, Y. and A. Tsukagoshi (2002) : Brackish-water ostracoda (Crustacea) from Obitsu River estuary, Central Japan. Species Diversity, 7, 1, 67-115.
中尾有利子・塚越 哲(2008):干潟域に生息する貝形虫群 集の季節変化と経年変化.日本大学部文理学部自然科学 研究所研究紀要,43,249-275.
Rubey, W. W. (1933) : Settling velocities of gravel, sand, and silt particles. American Journal of Science, 5th Series, 25, 325-338.
Theisen, B. F. (1966) : The life history of seven species ostra-cods from a Danish brackish-water locality. Meddelelser fra Danmarks Fiskeri-og HavundersØgelser, 4, 8, 215-270. 内山雄介(2005):干潟の流れと地形変化.ながれ, 24, 57-66.