細部課題1 トラフグ再生産効果の遺伝的な解析とその検証 細部課題2 トラフグの放流効果を判定するための遺伝情報データベースの構築 細部課題3 ヒラメの再生産効果を検証するための評価手法の検討 細部課題4 マダイの放流事業および遺伝的知見に関わる情報収集
◎細部課題 1 トラフグ再生産効果の遺伝的な解析とその検証
【 目 的 】 栽培漁業における種苗放流の効果は、主に放流魚の混入率や回収率などの一代回収型での 評価がなされてきた。既存の標識放流による追跡調査では天然海域において放流魚が再生産 を行うことによる資源増大効果の検証は不可能であり、種遺伝情報の分析により放流魚の子 孫を探索、定量することが必要である。本課題は、トラフグを対象として、両親の遺伝情報 が引き継がれる核DNA のマイクロサテライト(ms)DNA1)および母親のみの遺伝情報が引 き継がれるミトコンドリア(mt)DNA2)の 2 種類の DNA マーカーを整備して鑑定能力を評 価する。続いて、それらのマーカーを用いて種苗生産用親魚の遺伝情報を蓄積しデータベー ス化を実施する。さらに、データベース化した遺伝情報に基づき、これらの種苗生産用親魚 から生産され、放流された人工種苗を天然海域において検出することについても検討する。 【 研 究 方 法 】 1 ) DNA マ ー カ ー 整 備 の た め の ト ラ フ グ サ ン プ ル の 選 定 DNA マーカーの整備には、2011 年 3 5 月に若狭湾、瀬戸内海中央部の走島沿岸および伊 勢湾において漁獲された3 標本集団(いずれも成魚)を用いた(図 1)。この漁獲サンプルは DNA マーカーの整備及び鑑定能力を調べることを目的とすることから、標識(胸鰭切除、背 部への焼印、胸鰭基部へのイラストマー注入または耳石のアリザリンコンプレクソン(ALC) による染色)や外部形態の異常(尾鰭変形または鼻孔隔皮欠損)から人工種苗由来の可能性 のある個体は標本集団から除外した。 2 ) DNA マ ー カ ー の 整 備 2 ) ̶1 mtDNA マ ー カ ー の 整 備 昨年度は調節領域 3)前半部(523 塩基)を対象としたが、個体識別能力を強化するため調 節領域全配列(820∼822 塩基)を読み取り、その配列を鑑定の対象とした(図 2)。 2 ) ̶2 整 備 し た マ ー カ ー の 天 然 で の 応 用 と ハ プ ロ タ イ プ 多 型 の 検 出 の 検 討 DNA マーカーとしての遺伝的変異性や鑑定能力の評価では、上記1)の若狭湾、瀬戸内海 中央部の走島沿岸および伊勢湾海域の3 標本集団を対象に各個体の塩基配列(ハプロタイプ) を決定し、標本集団内の遺伝的変異性(ハプロタイプ数、ハプロタイプ多様度4))を求めた。 2 ) ̶3 msDNA マ ー カ ー の 整 備 msDNA 分析には、過度なスタッタリング 5)がなく、明瞭な遺伝子型決定が可能と判断さ れたKai et al. (2005)が開発した 11 ローカス6)(f61、f100、f60、f112、f86、f1770、f178、 f160、f153、f204、f65)、Cui et al. (2005)が開発した 3 ローカス(Cst-6、Cst-7、Cst-8)Furukawa et al. (2004)が開発したfms13、Takagi et al. (2001)が開発した Tru-17 の計 16 ローカスを選定し、マルチプレックス法 7)により PCR8)増幅した。これらのローカスは図 3 に示すように独立した染色体上に位置している。 2 ) ̶4 整 備 し た マ ー カ ー の 天 然 で の 応 用 と 遺 伝 子 型 の 決 定 、 ア リ ル 多 様 度 、 ヘ テ ロ 接 合 体 率 等 の 検 討 DNA マーカーとしての遺伝的変異性や鑑定能力の評価では、上記1)の 3 標本集団を対象 に各個体の遺伝子型を決定し、標本集団内の遺伝的変異性(アリル9)多様度10)、ヘテロ接合 体率(HE)11)、多型情報含有値(PIC)12)およびヌルアリル13)の頻度の推定値を求めた。 3 ) 種 苗 生 産 用 親 魚 の 遺 伝 情 報 デ ー タ の 蓄 積 種苗生産用親魚の遺伝情報データの蓄積については、独立行政法人水産総合研究センター 瀬戸内海区水産研究所屋島庁舎(屋島庁舎)で養成している種苗生産用親魚(以下、屋島親魚と する)を対象とした。これらの親魚は、香川県三豊市仁尾町沖の定置網で漁獲されたトラフ グ天然1 歳魚を養成したものである(図 4)。種苗生産用親魚には PIT タグ 14)が装着され、 個体識別されている。サンプルとした種苗生産用親魚の年齢は、2011 年 5 月 1 日時点で 6 11 歳であった。また、生存個体だけでなく、現在は死亡しているが 過去に種苗生産に用い られていた親魚のエタノール固定された鰭の標本も分析に供した。整備した DNA マーカー を用いて各個体のmtDNA および msDNA 分析を行い、データベースに登録した。 4 ) 山 口 県 埴 生 沿 岸 域 に お け る 0 歳 魚 での DNA マーカーを用いた鑑定試験 DNA マーカーを用いた鑑定試験は、山口県瀬戸内海沿岸の埴生沿岸で 2011 年 7 月 6 11 日に底曵網で漁獲された平均全長 78.03mm(61.32∼92.80mm)、平均体重 11.46g(5.27∼ 19.46g)の 0 歳の人工種苗および天然魚 22 個体をを対象とした。2011 年 6 7 月に埴生沿 岸では複数の群から構成された19.9 万尾の人工種苗が放流された。そのうち約 3 割に当たる 6 万尾は屋島庁舎で養成していた親魚に由来(以下、屋島由来とする)する。さらに、屋島 由来の6 万尾のうち 1.5 万尾には右胸鰭切除もしくは耳石の ALC 染色の標識が施されていた。 人工種苗の識別は、胸鰭切除、背部への焼印および外部形態の異常に加え、耳石の形状異常 (割れるなど)に基づいて行った。 4 ) ̶1 mtDNA 分 析 山口県瀬戸内海沿岸の埴生沿岸で漁獲された0 歳魚 22 個体の mtDNA 調節領域の塩基配 列を分析し、屋島親魚と比較した。 4 ) ̶2 msDNA 分 析 山口県瀬戸内海沿岸の埴生沿岸で漁獲された0 歳魚 22 個体の msDNA を分析し、屋島親 魚と比較した。
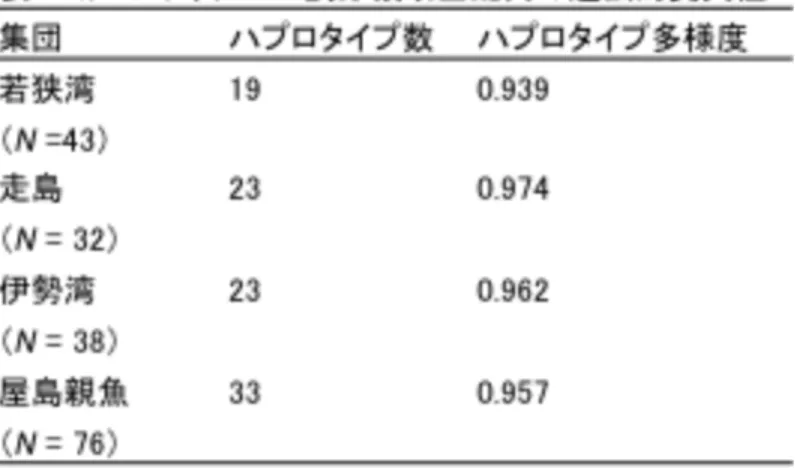
試験 設 定 の フ ロ ー 図 【 研 究 成 果 の 概 要 】 1 ) mtDNA 分 析 トラフグ野生集団である若狭湾、走島および伊勢湾の標本集団について遺伝的変異性を求 めた結果、分析個体数43、32、38 に対して検出されたハプロタイプ数はそれぞれ 19、23 お よび23、ハプロタイプ多様度は 0.939 0.974 であった(表 1)。一方、屋島親魚 76 個体を 分析した結果、33 のハプロタイプが検出された(表 1)。昨年度、調節領域前半部を用いた際 は、屋島親魚から 15 のハプロタイプが検出されたが、本年度、対象を約 300 塩基増やすこ とでハプロタイプは増加した。 2011 年に山口県埴生で漁獲された 0 歳の人工種苗 21 個体、天然魚 1 個体の調節領域全配 列を決定し、屋島親魚の塩基配列と比較した結果、12 個体の人工種苗が雌の種苗生産用親魚 4 個体のいずれかと同一の塩基配列を有していた(図 5)。一方、それら人工種苗のうち 5 個 体はmsDNA 分析では親子関係は否定された(図 5)。 2 ) msDNA 分 析 トラフグ野生集団におけるmsDNA16 ローカスの遺伝的変異性を若狭湾(43 個体)、走島 (32 個体)および伊勢湾(38 個体)の標本集団について求めた結果、アリル多様度は 17.2 19.0、HEは0.667 0.952、PIC は 0.637 0.950 を示した(表 2)。また、いずれのローカ スにおいてもヌルアリルの存在確率は有意ではなかった(P>0.05)(表 2)。また、個体判別
率をPI 値(任意に選んだ 2 個体間で全ローカスの遺伝子型が一致する確率)により評価した ところ、若狭湾で8.2×10-28、走島で 2.5×10-27、伊勢湾で 7.3×10-29となり、高い個体判別力 を示した。 屋島庁舎における種苗生産用親魚76 個体のマーカー型を決定した(表 2、3)。 2011 年に埴生で漁獲された 0 歳の人工種苗 21 個体、天然魚 1 個体のマーカー型を決定し、 親子鑑定を行った結果、人工種苗21 個体のうち、7 個体の人工種苗の両親を特定することが できた(表4)。それ以外の 14 個体の人工種苗については屋島親魚とは合致しなかった。 【 考 察 】 mtDNA 分析については、トラフグ野生集団の調節領域全塩基配列を分析することで昨年 度得られた結果よりも鑑定能力を改善することができた。しかし、漁獲された人工種苗と屋 島親魚との塩基配列が一致してもmsDNA 分析で親子関係否定された場合も認められ、トラ フグのmtDNA 調節領域の塩基配列の多様性はヒラメと比べてもやや低いことから(細部課 題3 を参照)、mtDNA のみを用いた親子鑑定はトラフグでは不確実性を含むことも明らかと なった。このことから、本種の親子鑑定は調節領域のみだけではなく、msDNA 分析と組み 合わせることが必須である。 msDNA 分析については、確実にかつ容易に遺伝子型決定することが可能なローカスの選 抜と効率よく遺伝子型決定を行うことのできるマルチプレックス化を図ることができた。そ して、トラフグ野生集団の遺伝的変異性を評価することで DNA マーカーとして高い鑑定能 力を有すると判断された。さらに、予備的試験において、複数の群から構成される人工種苗 や天然稚魚の中から正確に特定の種苗生産用親魚群から生産された人工種苗を検出できるこ とを実証した。 本年度の結果より、放流魚の子孫を探索するためには、まず mtDNA 分析を行い、絞り込 まれた候補についてmsDNA 分析を行うことが効率的であると考えられる。このことにより、 msDNA 分析のみと比較してPI 値は約 104倍高くなると考えられ、本事業の目的に沿うもの であると期待される。 【 次 年 度 以 降 に 向 け た 提 言 】 今回整備したmsDNA16 ローカスならびに mtDNA 調節領域全塩基配列は高い変異性を 有しており、各個体について全ての情報を取得することで漁獲物の中から特定の種苗生産用 親魚から生産された人工種苗を検出することが可能であることが示唆された。今後、これら のマーカーを用いて人工種苗が天然海域で再生産にどの程度寄与しているかについて評価し ていく必要がある。 【 参 考 文 献 】
Cui JZ, Shen XY, Yang GP, Gong QL, Gu QQ (2005) Characterization of microsatellite DNAs in Takifugu rubripes genome and their utilization in the genetic diversity analysis of T. rubripes and T. pseudommus. Aquaculture 250: 129-137.
Nishida M, Watabe S (2004) Isolation of microsatellite markers by in silico screening implicated for genetic linkage mapping in Japanese pufferfish Takifugu rubripes. Fish. Sci. 70: 620-628.
Kai W, Kikuchi K, Fujita M, Suetake H, Fujiwara A, Yoshiura Y, Ototake M, Venkatesh B, Miyaki K, Suzuki Y (2005) A Genetic Linkage Map for the Tiger Pufferfish, Takifugu rubripes. Genetics 171: 227-238.
Reza S, Furukawa S, Mochizuki T, Matsumura H, Watabe S (2008) Genetic comparison between torafugu Takifugu rubripes and its closely related species karasu Takifugu chinensis. Fish. Sci. 74: 743-754.
Shields GS, Kocher TD (1991) Phylogenetic relationship of North American Ursids based on analysis of mitochondrial DNA. Evolution 45: 218-221.
Takagi M, Sakai H, Takei T, Kaneko Y (2001) Mechanism of inbreeding and genetic diversity of the Tiger puffer Takifugu rubripes. In: Japanese Fisheries Resource Conservation Association Report (in Japanese). Japanese Fisheries Resource Conservation Association, Tokyo, pp 41-47.
図 2. トラフグの mtDNA. 赤線で囲まれた領域が調節領域 図 3. 各染色体とそこに座乗する msDNA ローカスのイメージ図. 黄緑の棒は染色体、ピンク の横線は msDNA ローカスを表す.染色体上の番号は染色体の番号、ピンクの線の左側に msDNA のローカス名を表現している.
図 4. (独)水産総合研究センター瀬戸内海区水産研究所屋島庁舎の位置および種苗生産用 親魚の漁獲された地点. mt DNA 調節領域全配列の系 統が同一のグループ DNA msDNA 分析から親子関係と認識は○、一 方、mtDNA 調節領域の塩基配列は同じだ けれども、msDNA 分析から親子関係が否 定されたものは●とした.
図 5. 2011 年 7 月に山口県瀬戸内海側の埴生周辺で漁獲された 0 歳の人工種苗、天然稚魚お よび屋島親魚の mtDNA 調節領域全配列(820 822 塩基)の系統関係.同一の塩基配列 を持つ個体は赤線で囲んだ。調節領域の塩基配列が屋島親魚と同一で、msDNA 分析から も親子関係と認識された人工種苗には○を、一方、調節領域の塩基配列は同一だが、 msDNA 分析からは親子関係が否定された人工種苗には●を付記している.
◎細部課題 2 トラフグの放流効果を判定するための遺伝情報データベースの
構築
【 目 的 】ミトコンドリア(mt)DNA 及びマイクロサテライト(ms)DNA 等の DNA マーカーを用 いて放流効果を算出するには、正確な親子判定を行い、放流魚やその子孫を効率的に検出す ることが重要である。特に、msDNA では、一組の両親から生まれた子供でもそれぞれ異な る遺伝子の組み合わせを持つため、親子判定を行う場合には親魚の遺伝情報データベースの 構築と、コンピュータープログラムによる組み合わせ計算が不可欠である。また、mtDNA についても親魚や漁獲物から得られた多数の DNA 配列を正確に比較するうえでデータベー スの利活用が必須である。以上のことから、本課題ではトラフグの親魚を対象とした DNA 情報データベースの構築を目的とする。 【 研 究 方 法 】 1 ) 親 魚 デ ー タ ベ ー ス の 作 成
昨年度、市販の表計算ソフト(Microsoft 社製, Microsoft ExcelTM)上に試作したデータベー
グのmsDNA 及び mtDNA の多型情報を登録し、再生産効果の評価に向けた親魚データベー スの構築を図る。また、内蔵のプログラミング言語(Microsoft 社製, Excel VBATM)によって
データ抽出機能等を付加し、実際にデータベースとして十分に機能するかどうか検討する。 2 ) 再 生 産 シ ミ ュ レ ー シ ョ ン ソ フ ト の 作 成 再生産個体の検出及び再生産評価法を検討するため、放流種苗と天然集団との交配シミュ レーションプログラムを作成し、放流種苗の再生産に伴う天然海域での遺伝的組成の変化に ついて検討する。また、プログラムは、Perl 等のより高速な計算を可能とするプログラミン グ言語を用いて作成する。 【 研 究 成 果 の 概 要 】 1 ) 親 魚 デ ー タ ベ ー ス の 作 成 昨年度入力したトラフグ親魚76 尾の msDNA 及び mtDNA の情報に加え、山口県埴生沖 で再捕された放流種苗21 尾及び天然稚魚 1 尾の DNA 情報を登録した。また、個体識別能力 を強化するため、昨年度入力した親魚について新たにPIC が 0.9 以上の高い個体識別能を有 する6 ローカスの msDNA(f100, f160, f60, f1770, f204, f153)情報を追加するとともに、 親魚の各個体について、mtDNA の調節領域前半部の塩基配列情報を約 300bp 追加し、合計 800 塩基(一部塩基の挿入により 822 塩基となる)とした。これらにより、親魚と放流種苗及 び天然稚魚をあわせた 98 個体について、16 ローカスの msDNA 情報と 820∼822 塩基の mtDNA 塩基配列情報が蓄積された。また、新たに追加した mtDNA 情報については、デー タベースに付加しているハプロタイプ分析機能によって再度解析し、新たなハプロタイプ番 号を登録した。この結果、ハプロタイプ数は昨年度の15 種類から 36 種類に増加した(表 1)。 2 ) 再 生 産 シ ミ ュ レ ー シ ョ ン ソ フ ト の 作 成 放流種苗の再生産効果を把握するには、再生産個体の遺伝子型を予測し、実際に天然集団 中でそれらの頻度を調査する必要がある。また、より実用的な手法を確立するには、調査の 規模やマーカー数等について具体的に検討しなければならない。しかし、これまで、再生産 個体の検出を目的とした水産生物の調査のみならず検討は非常に少ない。このため、本課題 では、放流種苗と天然個体が交配を行った場合に、どのような遺伝子型がどの程度集団中に 存在しうるか予測することを目的として再生産シミュレーションソフトの作成に取り組んだ。 一般に、放流尾数に対して天然の資源量は非常に多いと考えられる。このため、シミュレ ーションでは、資源量が大きくアリル頻度が安定している天然集団と放流種苗の各個体との 間で各ローカスのどちらかのアリルを交換するものとした(図 1)。また、天然集団の各アリル が交配によって放流種苗の子世代に採用されるかどうかは、天然集団のアリル頻度に従って 決定されるものとした。ただし、天然集団と放流種苗の遺伝的交流によって、天然集団の遺 伝子頻度も影響を受けるものとし、この際、放流種苗の資源量は任意に設定可能とした(実 際の操作では相対的な大きさとして入力)。 今年度は、高速での計算が可能なプログラミング言語Perl を用いてソフトウェアを作成し、 様々な放流種苗のマイクロサテライトDNA 及びミトコンドリア DNA データに対して、上記 の設定に基づく計算が可能となった(図 2)。
【 考 察 】 親魚データベースにおいては、データを追加しても検索及び抽出機能は有効に働いており、 基本的に現在のプラットフォームでデータ数の増加にも対応できるものと考えられた。今後 は、他魚種についても登録を進め、問題点の抽出を行うべきと思われる。また、シミュレー ションにおいては、数式モデルによる予測とは異なり、世代交代の各試行において個体毎の アリル型を予測することができた。このことは、放流種苗の再生産の影響を把握するための 解析法を検討する上で非常に効果的であり、今後、放流集団や天然集団の資源量及びマーカ ー数等、様々な条件下でのシミュレーションを行うことで、実際の分析に有効な解析法が開 発できるものと考えられた。 【 次 年 度 以 降 に 向 け た 提 言 】 データベースについては、さらに多くのデータを登録するとともに、各県や自治体での試 験的な運用を行い、より利用しやすいものに改善していく必要がある。また、再生産シミュ レーションソフトについては、放流種苗の実データを用いたシミュレーションを実施する等、 再生産効果の調査手法の確立に向けてより具体的な検討を行う必要がある。
図 1. 再生産シミュレーションの基本概念 図 2. 再生産シミュレーションソフトの起動画面 Pwij: 天然集団j世代でのi番目のアリル頻度,Psij: 放流種苗j世代でのi番目のアリル頻度. Nw: 天然集団の資源量,Ns: 放流種苗の資源量,N=Nw+Ns *天然集団、放流種苗ともに 0 世代目は実データ