腎臓において血圧概日リズムを制御している分子の探索
日本大学大学院医学研究科博士課程 内科系内分泌代謝内科学専攻
村田 悠輔
修了年 2017 年
指導教員 相馬 正義
腎臓において血圧概日リズムを制御している分子の探索
日本大学大学院医学研究科博士課程 内科系内分泌代謝内科学専攻
村田 悠輔
修了年 2017 年
指導教員 相馬 正義
【目次】
概要……… 1
諸言……… 3
生体内時計について……… 3
高血圧と生体内時計……… 5
腎臓と生体内時計……… 6
高血圧自然発症ラット……… 7
研究目的……… 9
対象と方法……… 9
1. 実験動物……… 9
2. 全ゲノムにおけるプロモーター内に時計蛋白結合配列( E-box )を持つ遺 伝子の検索……… 10
3. DNA マイクロアレイを用いた、 WKY/Izm 、 SHR/Izm の腎臓における遺 伝子発現の比較……… 10
4. TCMK-1 細胞における候補遺伝子の発現変動の検出……… 11
5. リアルタイム PCR ……… 12
6. TCMK-1 細胞の免疫蛍光染色……… 13
7. WKY/Izm 、 SHR/Izm 腎臓の免疫染色……… 13
8. Western blot 法による WKY/Izm 、 SHR/Izm 腎髄質における FXR1 、 PPAT 蛋白発現の検討……… 14
9. 統計解析……… 15
研究結果……… 16
1. GSEA データベースによるプロモーター内に時計蛋白結合配列( E-box )
を持つ遺伝子の検索……… 16
2. DNA マイクロアレイを用いた、 WKY/Izm 、 SHR/Izm の腎臓における遺 伝子発現の比較……… 16
3. TCMK-1 細胞における生体内時計遺伝子 mRNA 発現振幅の誘導……… 17
4. デキサメサゾン添加による候補遺伝子 mRNA 発現変動の検討………… 18
5. TCMK-1 細胞における PPAT 、 FXR1 の細胞内での局在……… 19
6. WKY/Izm 、 SHR/Izm 腎臓における PPAT 、 FXR1 の局在……… 19
7. Western blot 法による WKY/Izm 、 SHR/Izm 腎髄質における FXR1 、 PPAT 蛋白発現の検討……… 20
考察……… 21
結語……… 27
謝辞……… 28
図表……… 29
参考文献……… 47
研究業績……… 50
1
【概要】
様々な生物現象に概日リズムがみられるが、中でも血圧に日内変動が見られ、
その異常が心血管合併症と相関することはよく知られている。血圧の日内変動 と生体内時計の関連性は容易に想像できるが、その機序は明らかになっていな い。腎臓は糸球体における濾過と、尿細管における電解質、水の再吸収、排泄、
さらにはレニン - アンギオテンシン系における血管収縮作用、 Na の再吸収促進 など血圧調整において極めて重要な役割を果たし、その機能が血圧の概日リズ ム形成に関わっていると考えられる。本研究では腎臓に発現し、高血圧に関連 した分子を検索し、その中で、生体内時計により概日リズムが形成される可能 性のある分子を明らかにする。
Gene Set Enrichment Analysis ( GSEA; BROAD INSTITUTE )データベー スを用いて、転写開始点から上流または下流 2000bp に生体内時計のメインルー プ の 中 心 と な る 時 計 蛋 白 で あ る BMAL1/CLOCK の 結 合 配 列 で あ る
E-box(CACGTG) を持つ遺伝子を検索し、 1032 個の遺伝子が抽出された。次に
DNA マイクロアレイを用いて 5 週齢の高血圧自然発症ラット( Spontaneously Hypertensive Rat : SHR/Izm )と Wister Kyoto Rat ( WKY/Izm )の腎臓にお ける遺伝子発現を検討した。 1032 個の E-box 配列を持つ遺伝子のうち、
SHR/Izm の腎皮質、髄質で発現が 2 倍以上増加または 50% 以下に減少している
遺 伝 子 で 、 さ ら に 脳 卒 中 易 発 症 SHR(Stroke-Prone Spontaneously
Hypertensive Rat : SHRSP/Izm) の腎皮質、髄質でも同様の発現変化がみられ
たものは、 Bcl6 、 Nr1d1 、 Ppat 、 Tef 、 Fxr1 、 Hnrnpa3 、 Npm1 、 Nptx1 、 Plagl1 、
Plbd1 、 Tbl1x 、 Trim46 の 12 遺伝子であった。これらの遺伝子は腎臓において
高血圧に関連し生体内時計に概日リズムが制御されている遺伝子であると考え
られた。
2
次いでマウス尿細管上皮細胞( TCMK-1 )に対してデキサメサゾン 0.5 μ M を 添加すると、時計遺伝子として知られている Period1(Per1) の遺伝子発現の概日 リズムが形成された。これにより TCMK-1 細胞に対するデキサメサゾン刺激は 腎尿細管の生体内時計研究において有用なツールであることが明らかとなった。
TCMK-1 細胞をデキサメサゾンで刺激し、 4 時間おきに 48 時間後まで mRNA
をリアルタイム PCR で定量し、 GSEA および DNA マイクロアレイの結果から 得られた 12 個の遺伝子のうちすでに生体内時計遺伝子として知られている
Nr1d1 を除く 11 遺伝子について、概日リズムが形成されるのか検討を行った。
得られた 11 個の遺伝子の 48 時間の mRNA 変動について周期回帰分析にて検定 を行ったところ、 Phosphoribosyl pyrophosphate amidotransferase ( Ppat ) 、 Fragile X mental retardation, autosomal homolog 1 ( Fxr1 )が有意差を持って 概日リズムを形成していることが明らかとなった。
PPAT 、 FXR1 の尿細管細胞内での局在を明らかにするために。 TCMK-1 細胞 を抗 PPAT 抗体、抗 FXR1 抗体を使用し、免疫染色を行った。 PPAT は尿細管 の核と細胞質に、 FXR1 は細胞質に染色された。次に腎での局在を明らかにする ため、 5 週齢の WKY/Izm 、 SHR/Izm 腎臓を使用し、免疫染色を行った。 PPAT は近位尿細管、遠位尿細管の核と細胞質に局在し、 FXR1 は近位尿細管、遠位尿 細管の細胞質に局在していた。
以上の結果より、 TCMK-1 細胞-デキサメサゾン系は時計遺伝子の振幅を再
現する事が可能であり腎臓における時間生物学的な解析を行う上で有用である
ことが示された。生体内時計蛋白結合配列である E-box を持ち、高血圧ラット
の腎臓で発現増加または発現減少し、培養実験系で発現振幅を形成した Ppat 、
Fxr1 は、腎臓において機能し、血圧概日リズム形成に関わる分子である可能性
が示された。
3
【緒言】
生体内時計について
様々な生物学的現象の観察から、生物が一日のリズム持つことが知られてき た。睡眠周期、心拍、血圧なども、概日リズムを示している。 18 世紀にフラン スの天文学者である de Marian はオジギソウが日中は葉を開き、夜は葉を閉じ る運動が恒常暗においても持続することを示し、生物現象を支配する生体内時 計の存在の可能性を報告した
(1)。一日のリズムは概日リズム( Circadian rhythm ) と呼ばれ、地球上の生物現象の概日リズムの周期は概ね約 24 時間である。生物 現象が概日リズムを形成するためには、正確な時を知らせる生体内時計の存在 が必要であり、この発見から現在では古細菌から人類に至るすべての生物にお いて概日リズムが存在することが推測されている
(2)。
概日リズムを形成するうえで、主となるペースメーカーが哺乳類においては 視交叉上核( Suprachiasmatic nucleus : SCN )に存在することが明らかとなり、
これを中枢時計と呼び、概日リズムの自律発振の中心となっている
(2)。その後 の研究により、多くの末梢臓器にも生体内時計が存在し、これらの時計は中枢 時計のコントロールを受けながら、それぞれの臓器に最適な時を刻むことが明 らかになっている。中枢時計は神経性因子および液性因子を介して末梢時計を コントロールしている。 SCN には自律振動をする多数の神経細胞からなり、ラ ットおよびマウスにおいてこれらの細胞を単離し分散培養を行うと、発火頻度 や遺伝子発現において若干異なる概日リズムを示したことから、 SCN における 個々の神経細胞は固有の概日振動体をもつ時計細胞といえる
(3)。
最近、生体内時計の時を刻む分子機構が明らかとなってきた。この概日リズ
4
ムを司る蛋白をコードする遺伝子群を時計遺伝子と呼ぶ。生体内時計は Brain and muscle arnt-like 1 ( BMAL1 ) 、 Circadian locomotor output cycles kaput
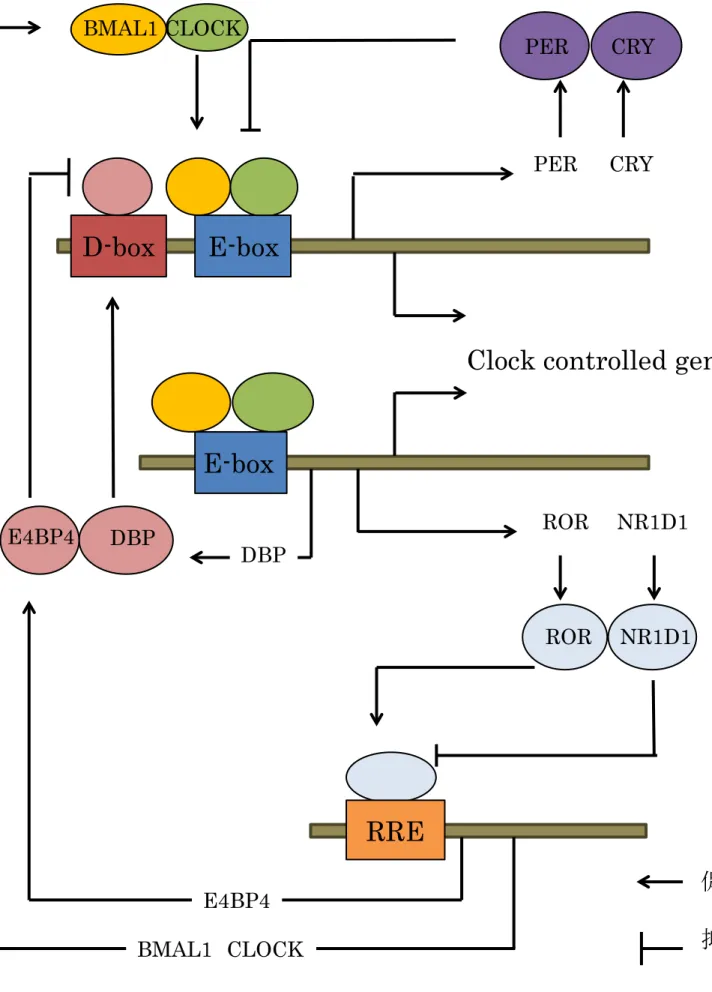
( CLOCK ) 、 Period ( PER ) 、 Cryptochrome ( CRY )がコア蛋白として働くメ インループから構成されている。細胞内において CLOCK 、 BMAL1 が二量体を 形成して転写因子として働きプロ―モーター内の E-box 配列( CACGTG )に結 合することで、多くのターゲット遺伝子の発現を促進する。ターゲット遺伝子 の中には Per や Cry 等の別の時計遺伝子も含まれている。転写、翻訳が増加し た PER 、 CRY 蛋白は核内へ移行し、 BMAL1 、 CLOCK を抑制する。 その後 PER 、 CRY はカゼインキナーゼ( CKI ε)、グリコーゲン合成酵素 (GSK-3 β ) 、 AMP 活性化プロテインキナーゼ( AMPK )などによる多段階リン酸化を経て時間を かけて分解される。これにより PER 、 CRY により BMAL1 、 CLOCK の抑制が 解かれ、再び CLOCK 、 BMAL1 による遺伝子発現の促進が始まり、時計が一周 する。このメインループのほかに、 Nuclear receptor subfamily 1 group D member 1 ( NR1D1 ) 、 Retinoic acid receptor-related orphan receptor ( ROR ) 、 D-site of albumin promoter binding protein ( DBP )、 E4 promoter binding protein 4 ( E4BP4 )が関与するサブループも存在している。 NR1D1 は RRE
( AGGTCA )配列に結合して、ネガティブフィードバックを経てメインループ
を抑制する一方で、 DBP は D-box ( TTATGTAA )配列に結合することでポジテ
ィブフィードバックを経て、メインループを促進する
(4)(図 1 )。このような生
体内時計の制御を受けて概日リズムを刻む蛋白は各組織において約 15 %にも及
ぶとの報告もあり
(5)、生体内時計は様々な生理現象の概日リズム形成に関わっ
ている。
5
高血圧と生体内時計
血圧は心拍出量と末梢血管抵抗の積であり、生体内体液量と交感神経の緊張 によって規定され、 従って血圧は元来変動するものである。 血圧が 140/90mmHg 以上は高血圧と定義され、高血圧は 30 歳以上の日本人男性の 60% 、女性の 45%
が罹患していると報告される罹患率の高い疾患であり
(6)、血圧の上昇に伴い心 血管合併症、慢性腎臓病の罹患リスクおよび死亡率を増加させる
(7, 8)。高血圧 の血圧変動に日内変動があることが知られ、夜間に血圧が低下する dipper 型と 呼ばれる変動が正常の変動であり、それに対し夜間血圧が低下しないものは
non-dipper 型、上昇するものは riser 型と呼ばれる。高血圧は多遺伝子性の本
態性高血圧と疾病に基づく血圧上昇の二次性高血圧に分けられ、二次性高血圧 では腎血管、腎実質の障害から来る腎血管性高血圧、腎性高血圧が多く、体液
貯留型の non-dipper 型高血圧を示す。最近原発性アルドステロン症の診断率が
上がり、高血圧の 6% は原発性アルドステロン症と報告され、体液貯留型の non-dipper 型高血圧を示す。 non-dipper 、 riser は dipper と比較し夜間血圧が 高値であり、心血管合併症を含めた臓器障害の独立したリスクとされている
(9)。 血圧が概日リズムを形成する事と、その乱れが臓器障害を起こすことから生体 内時計と高血圧の関連性も想定される。生体内時計を構成する遺伝子として知 られている、 Per 、 Cry 、 Bmal1 、 Clock と血圧の関係も多数報告されている。
Bmal1 ノックアウトマウスにて、血圧の低下、血圧日内変動の消失がみられ、
血管内皮細胞障害がみられる事や
(10, 11)、 Clock ノックアウトマウスでは、尿中
Na 排泄の増加、血圧低下、さらには心拍の低下と心拍の概日振幅の低下が報告
されており
(12)、生体内時計と血圧の関与が示唆される。
6
腎臓と生体内時計
腎臓が血圧調節に深く関わることはよく知られており、高血圧の病態に腎臓 は極めて重要な役割を果たしている。高血圧の腎性機序は、第一に心臓から糸 球体に至るまでの血管抵抗上昇に基づいた食塩非感受性高血圧であり、本態性 高血圧、腎血管性高血圧がこれにあたる。第二としては糸球体濾過能の低下に より、尿細管~糸球体フィードバック機構が作動し、輸入細動脈の血管抵抗の 低下にともないレニン・アンギオテンシン・アルドステロン( RAS )系が亢進 し、 Na 貯留が起こり血圧上昇を引き起こされる食塩感受性高血圧があげられ、
腎実質性高血圧がこれにあたる。第三には尿細管における Na 再吸収の増加に伴 う食塩感受性高血圧であり、本態性高血圧の塩分感受性群、原発性アルドステ
ロン症や Liddle 症候群がこれにあたる。腎臓における血圧に対する役割と、概
日リズムの関係も多数報告されている。正常の腎臓では夜間よりも日中に電解 質の排泄量や尿の産生量が多く、 Na 、 K 、 Cl 排泄に概日リズムがあることが知 られており
(13)、そのリズムの破綻と高血圧とその心血管合併症との関連が示唆 されている
(14)。生体内時計遺伝子の一つである、 Per1 ノックアウトマウスで は腎臓の集合管においてα subunit of the renal epithelial sodium channel (α ENaC )の機能低下がみられ、 Na 排泄が増加し、血圧が低下するすると報告さ れ
(15)、腎臓における生体内時計の関わりが示唆される。また、慢性腎臓病患者 において血圧の日内変動の異常( non-dipper )が見られ、 non-dipper を呈する 腎不全患者に腎移植を行うと、 non-dipper が改善されることなどから、腎臓が 血圧の概日リズム形成に関わることが強く示唆される
(16)。
腎臓における概日リズムは視交叉上核に存在する中枢時計のコントロールの
下、腎末梢時計が、腎臓に最適なタイミングを刻み、それにより概日発現変動
7
を制御された諸蛋白が血圧などの概日リズム形成に関わると考えられる。この ような理由から本研究においては高血圧において重要な役割を果たす腎臓に注 目して、生体内時計に制御を受けて血圧概日リズム形成を担う分子の同定を目 指した。
高血圧自然発症ラット
本態性高血圧の成因は未だ不明であり、ヒト本態性高血圧の分子生物学およ び遺伝学的病態や病因の解明の為には遺伝的高血圧モデル動物は必要である。
高血圧自然発症ラット (SHR) は 1964 年京都大学病理学の岡本、家森らが正常血
圧 Wister Kyoto (WKY) ラットの中から、血圧が高い個体を選択して分離し、系
統樹立した動物モデルであり、人為的な処置なしに高血圧を自然発症する
(17)。
SHR は高血圧のモデルとして長年に渡り世界中の多くの血圧研究者により使用
されてきている。 SHRSP は脳卒中が確認できた SHR の子孫を選択的に交配さ
せ、分離させた近交系である
(18)。 SHRSP も SHR と同様に、著明な高血圧を発
症し、塩分負荷により生後 25 週齢でほぼ全例が脳血管障害を発症するため、脳
卒中における唯一の自然発症モデルとして使用されている。 SHR 、 SHRSP では
4 週 齢 以 降 に 著 明 な 血 圧 上 昇 が み ら れ 、 12 週 齢 に お い て は SHR で は
180-200mmHg 、 SHRSP では 200mmHg 以上の重症高血圧を発症する。 SHR
では心肥大や血管肥厚を認め、さらに腎臓では腎硬化症が観察され、高血圧に
伴う心血管のリモデリングの研究に活用されている。 SHRSP は脳穿通枝動脈の
動脈壊死、血栓を発症し、ほぼ全例に脳卒中を伴うため、高血圧性脳血管障害
の研究に活用されている。尚、 SHR 、 SHRSP ともに、上記のような病態を示す
ため、 WKY と比較すると短命であり、 SHR は 18 か月程度、塩分負荷 SHRSP
8
では 8 か月程度である。以上の事から、 SHR 、 SHRSP はともに、本態性高血 圧症および合併症の動物モデルとして、本態性高血圧の成因となる遺伝子分析、
治療、予防、病態の研究において極めて有効である。 SHR は心拍、血圧の日内 変動のずれが加齢によって顕性化するといった報告
(19)や、 SHR から分離され、
肥 満 、 メ タ ボ リ ッ ク シ ン ド ロ ー ム を 呈 す る 肥 満 高 血 圧 自 然 発 症 ラ ッ ト
( SHR/NDmcr-cp )では non-dipper 型の血圧変動を示すことも報告され
(20)、
SHR 、 SHRSP などは血圧変動異常を研究する上で適切なモデルである。
9
【
研究目的】
様々な生物現象のリズムは生体内時計に制御されていると考えられる。血圧 の日内変動と生体内時計遺伝子の発現は密接に関与していると考えられるが、
その関連性は明らかにされていない。生体内時計の研究は経時的な検討が重要 であることから、その in vivo での検討は大きな困難を伴い、それを補う研究方 法が求められる。本研究では血圧調節に重要な役割を担う腎臓において、絶え ず電解質や水の排泄や再吸収を行う尿細管に着目し、培養尿細管細胞を用いた 生体内時計評価系の確立を目指す。さらに、遺伝子発現調節領域に生体内時計 結合配列である E-box を持つ遺伝子をスクリーニングし、その中で正常血圧の WKY/Izm に比べ高血圧の SHR/Izm 、 SHRSP/Izm 腎臓の皮質、髄質において 発現変化している遺伝子を抽出し、培養細胞を用いた生体内時計評価系により 解析して生体内時計に発現制御を受けながら血圧調節に関わる分子の同定を目 指す。
【対象と方法】
1. 実験動物
動物実験は、全て日本大学医学部動物実験委員会の指針に従って行った。 SHR 等 疾 患 モ デ ル 共 同 研 究 会 で 維 持 し て い る 純 系 の WKY/Izm ラ ッ ト お よ び
SHR/Izm 、 (雄性 4 週齢)はいずれも日本 SLC 株式会社(浜松)から購入して
実験に使用した。臓器摘出の際にはイソフルラン吸入麻酔( 2-5% in 100% 酸素)
下で体表を消毒後に腹部を正中切開し、下大静脈から採血をして失血死とした
後に、腎臓を摘出した。 (動物実験計画承認番号: AP14M049 )
10
2. 全ゲノムにおけるプロモーター内に時計蛋白結合配列( E-box )を持つ遺伝 子の検索
Gene Set Enrichment Analysis ( GSEA; BROAD INSTITUTE, USA )における Molecular Signatures Database を用いて、ヒトの転写因子ターゲットに関する遺伝 子群を検索し、遺伝子数の最も多いデータベースを用いて、転写開始点より上 流 2000bp または下流 2000bp に E-box(CACGTG) を持つ遺伝子を抽出した。
3. DNA マイクロアレイを用いた、 WKY/Izm 、 SHR/Izm 、 SHRSP/Izm の腎臓 における遺伝子発現の比較
腎臓における血圧概日リズムにかかわる分子を同定するため、高血圧自然発 症モデルである SHR/Izm と SHR/Izm から系統樹立された、高血圧を呈し、塩 分負荷で高率に脳卒中を発症する SHRSP/Izm の腎臓における遺伝子発現を DNA マイクロアレイを用いて網羅的に解析した。この結果と GSEA における解 析結果を組み合わせ、 E-box 配列を持ち、 2 つの同系統であるが異なる高血圧モ デルの腎臓において発現変動をしている遺伝子を抽出した。さらに、 SHR/Izm 、
SHRSP/Izm はともに 5-7 週齢頃から血圧の上昇がみられ、血圧上昇後はそれに
引き続いて起こる 2 次的な変化により遺伝子発現が修飾されることが予想され るため、血圧が上がる直前の 5 週齢の WKY/Izm 、 SHR/Izm 、 SHRSP/Izm を使 用した。
4 週齢の SHR/Izm 、 SHRSP/Izm と WKY/Izm の分与をうけ、 12 時間毎の明暗サ
イクル( 8 時〜 20 時が明期、 20 時〜 8 時が暗期)のもと通常食で飼育し、 5 週齢
の時点で 12 時に各々 3 頭から腎臓を摘出した。摘出した腎臓を円刃を用いて用
11
手的に皮質、髄質に分け、 TaKaRa BioMasher Standard ( Takara, Japan )でホモジ ナイズし、 Maxwell16 Tissue LEV Total RNA Purification Kit ( Promega, USA )を用 いて Total RNA を抽出した。 抽出された Total RNA 50 ng をもとに Low Input Quick Amp Labeling Kit ( Agilent Technologies, USA )を用いて蛍光ラベル化アンチセン ス相補鎖 RNA を作成し、 RNeasy mini spin columns ( Qiagen, Germany )で精製し た。ハイブリダイゼーションしたのち Agilent Technologies Microarray Scanner
( Agilent Technologies, USA )でスキャンし、 Agilent Feature Extraction 10.7.3.1
( Agilent Technologies, USA ) で 数 値 化 の う え 、 Gene Spring GX ( Agilent
Technologies, USA )でグローバルノーマライゼーションして遺伝子発現を比較検
討した。 2 倍以上または 0.5 倍以下を閾値として各ストレイン間で発現が変動し ている遺伝子を抽出した。
4. TCMK-1 細胞における候補遺伝子の発現変動の検出
( ア ) 細胞培養
マウス尿細管上皮細胞( TCMK-1 )を 10% ウシ胎児血清 (Fetal bovine serum ; FBS ; HyClone Laboratories, USA) 、ペニシリン G カリウム (Invitrogen, USA) 100 U / ml 、硫酸ストレプトマイシン 100 μ g / ml を添加した Dullbecco’ s modified eagles’ medium (DMEM ; Invitrogen, USA) で 37 度、 5%CO2 の条件 下で培養を行い、 6 well plate (2ml / well)13 枚に播種した。そして 24 時間培養 後、サブコンフルエントの状態とした。
( イ ) デキサメサゾン刺激による遺伝子発現の振幅の誘導
1 枚の 6 well plate をコントロール ( 刺激後 0 時間 ) とし、その他 12 枚の 6 well
12
plate を 0.5 μ M デキサメサゾン (Sigma-Aldrich, USA) で刺激し、 2 時間 37 ℃、
5%CO2 で培養後、 無血清 DMEM に培地交換を行った。 デキサメサゾン刺激後、
0 時間、 4 時間、 8 時間、 12 時間、 16 時間、 20 時間、 24 時間、 28 時間、 32 時 間、 36 時間、 40 時間、 44 時間、 48 時間に細胞回収を行い、 RNA を抽出し、
リアルタイム PCR 法で Per1 、 Nr1d1 、 Cry1 、 Bmal1 、 Ppat 、 Fxr1 、 B-cell lymphoma 6 ( Bcl6 ) 、 Heterogeneous nuclear ribonucleoprotein A3 ( Hnrnpa3 )
、Nucleophosmin ( Npm1 )、 Neuronal pentraxin 1 ( Nptx1 )、 Pleiomorphic adenoma gene-like 1 ( Plagl1 ) 、 Phospholipase B domain containing 1 ( Plbd1 )
、Transducin (beta)-like 1X-linked ( Tbl1x )、 Tripartite motif containing 46
( Trim46 ) 、 Thyrotroph embryonic factor ( Tef )の mRNA 発現を定量した(図 2 ) 。
5. リアルタイム PCR
TCMK-1 細胞の mRNA 定量を TaqMan プローブ法または SYBR Green 法で 行った。 TCMK-1 細胞から RNA 抽出後、 High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, USA) を用いて逆転写を行い、 cDNA を 生成した。その後、 7500 Real Time PCR System (Applied Biosystems, USA) にて、 95 ℃ 30 秒を初期変性とし、 95 ℃ 5 秒、 60 ℃ 34 秒のサイクルで 40 サイク ル 行 っ た 。 同 様 の 工 程 で 、 Glyceraldehyde-3-phosphate dehydrogenase
( GAPDH ) RNA に対して RT-PCR を行い、内部標準とした。使用したプラ
イマーは表 1 に記す。尚、各遺伝子の mRNA 発現変動を示したグラフは、各時
間の mRNA 発現量の平均値をプロットし、各点の間は線形補間しスムージング
を行い作成した。
13
6. TCMK-1 細胞の免疫蛍光染色
チャンバー付きスライドグラスに、 TCMK-1 を 10%FBS 含有 DMEM にて 37
度、 5%CO2 の条件下で培養し、 24 時間培養後、サブコンフルエントとした。
1%Phosphete buffered saline ( PBS )で洗浄後、 4% パラホルムアルデヒドで 10 分間固定し、 PBS で洗浄後、 0.1%TritonX – 100/PBS ( Sigma-Aldrich, USA ) を 用 い て 10 分 間 室 温 で イ ン キ ュ ベ ー シ ョ ン し た 。 10% goat serum 1%
Albumin/PBS で 60 分間固定後、 ウサギモノクローナル抗 PPAT 抗体 (#AV46079;
Sigma-Aldrich, USA) を 1%BSA/PBS で 250 倍に希釈し、ウサギモノクローナ ル抗 FXR1 抗体 (#AB129089; ABCAM, UK) を 300 倍に希釈し、室温で 1 時間 インキュベーションさせた。 2 次抗体として Alexa fluor 488 anti-rabbit IgG
( Invitrogen, USA )を 1000 倍希釈し、暗所で 1 時間反応させた。洗浄後、
4',6-Diamidino-2-Phenylindole を超純水で 2000 倍希釈し、 5 分間暗所室温で反 応させたのち、 Fluoromount G(Southern Biotech, USA) で封入し、 Fluoview FV10i(Olympus life science, Japan) で観察した。
7. WKY/Izm 、 SHR/Izm 腎臓の免疫染色
5 週齢の WKY/Izm 、 SHR/Izm の腎臓を摘出し、 ASP200S(Leica Biosystems,
Germany) にてパラフィン包埋後、 4 μ m に薄切後、脱パラフィン処理を行った。
0.01M クエン酸バッファー (pH 6.0) にて圧力釜で 20 分低圧で抗原賦活化を行い、
室温で冷却後、 3% 過酸化水素水で 10 分間インキュベートし、内因性ペルオキ
シダーゼの不活化を行った。 PBS で洗浄後、 10% ヤギ血清にて保湿箱内で 30 分
ブロッキ ング後、 1 次抗体 としてウ サギモノ クローナ ル抗 PPAT 抗体を
14
1%BSA/PBS で 250 倍に希釈、ウサギモノクローナル抗 FXR1 抗体を 200 倍に
希釈し、 4 ℃、 12-24 時間反応させた。反応後、 2 次抗体としてヒストファイン
シンプルステインラット MAX-PO ( MULTI ) ( #414191; ニチレイバイオサイ エンス , Japan )を使用し、保湿箱にて 30 分室温でインキュベーション後、 PBS で洗浄し、 DAB 基質キット( #425011; ニチレイバイオサイエンス , Japan )を 使用し、 3 分間室温で発色させた。マイヤーヘマトキシリン溶液に 5 分間浸し、
核染後、流水で 10 分間洗浄した。脱水、透徹処理し、マリノールで封入後、光 学顕微鏡にて観察した。
8. Western blot 法による WKY/Izm 、 SHR/Izm 腎髄質における FXR1 、 PPAT 蛋白発現の検討。
免疫染色の結果、 FXR1 、 PPAT は尿細管に発現しており、 WKY/Izm 、 SHR/Izm において、尿細管の割合が大きい腎髄質での FXR1 、 PPAT タンパクの発現を Western blot 法にて検討した。円刃にて分離した腎髄質を、 RIPA buffer ( nacalai tesque, Japan )を用いてタンパク抽出し、 Pierce BCA protein assay kit ( Themo scientific, USA )にてタンパク定量した。 2- メルカプトエタノールを含有する sample buffer を用いて 95 ℃、 5 分間熱処理を行いて変性処理し、サンプルを 1 レーンあたり 10 μ g アプライし、 20 - 30mA で電気泳動を行った。その後、 iblot
( Invitrogen, USA )にて PVDF 膜に転写し、 5% スキムミルク溶液で 1 時間室 温にてブロッキングした。 1 次抗体はウサギモノクローナル抗 FXR1 抗体、ウ サギモノクローナル抗 PPAT 抗体を使用し、 4 ℃オーバーナイトでインキュベー ションし、 2 次抗体は HRP-conjugated goat anti rabbit IgG ( Jackson immune
reseach, USA )を用いて室温で 1 時間インキュベートした。タンパク質の検出
15
は Chemi-Lumi One Super ( #02230 nacalai tesque, Japan )を使用し、
LAS3000 ( Fuji film, Japan )にてバンドを化学発光にて検出及び作成を行い、
Image J ( https://imagej.nih.gov/ij )を用いてバンドを定量化した。内部標準は GAPDH を用い、 rabbit anti GAPDH ( #G9545, Sigma-Aldrich, USA )を使用 した。
9 .統計解析
TCMK-1 細胞に対するデキサメサゾン刺激後の mRNA 変動の周期性は,コサ
インカーブを最小二乗法により最適化を行い、メサー( MESS) 、振幅( AMP )、
位相( TZ1) を求める多変量回帰分析であるシングルコサイナー法を用いて
MESS , AMP , TZ1 の推定値とその 95 %信頼区間を算出した。なお、コサイナ
ー法の解析には SAS9.4(SAS Institute Japan, Japan) を用いて行った。 Western
blot 法における WKY/Izm 、 SHR/Izm のタンパク質発現の両群間の差は、 F 検
定で P>0.05 を確認した後、 t 検定を行い、 P<0.05 を有意とした。
16
【研究結果】
1. GSEA データベースによるプロモーター内に時計蛋白結合配列( E-box )を 持つ遺伝子の検索
まず初めに、 GSEA を用いて、プロモーター内に生体内時計の中心を担う時 計 蛋 白 で あ る CLOCK 、 BMAL1 ヘ テ ロ ダ イ マ ー の 結 合 配 列 で あ る
E-box(CACGTG) 、を持つ遺伝子を全ゲノムに渡り検索した。その結果、転写開
始点から上流または下流 2000bp の間に E-box を持つ 1032 個の遺伝子が同定さ れた(表 2 ) 。
2. DNA マイクロアレイを用いた、 WKY/Izm 、 SHR/Izm 、 SHRSP/Izm の腎臓 における遺伝子発現の比較
DNA マイクロアレイを用いて 5 週齢の SHR/Izm と WKY/Izm 腎臓における 遺伝子発現の違いを検討した。腎皮質においては 363 の遺伝子が WKY/Izm と
比べて、 SHR/Izm において 2 倍以上の発現の増加がみられ、 281 の遺伝子が
WKY/Izm と比べて、 SHR/Izm において 0.5 倍以下の発現の低下がみられた。
腎髄質においては、 284 の遺伝子が WKY/Izm と比べて、 SHR/Izm において 2 倍以上の発現の増加がみられ、 203 の遺伝子が WKY/Izm と比べて、 SHR/Izm において 0.5 倍以下の発現の低下がみられた。同様に、 5 週齢の WKY/Izm と
SHRSP/Izm の腎臓における遺伝子発現の違いを検討したところ、腎皮質におい
ては 267 の遺伝子が WKY/Izm と比べて、 SHRSP/Izm において 2 倍以上の発
現の増加がみられ、 580 の遺伝子が WKY/Izm と比べて、 SHRSP/Izm において
17
0.5 倍以下の発現の低下がみられた。腎髄質においては、 639 の遺伝子が WKY/Izm と比べて、 SHRSP/Izm において 2 倍以上の発現の増加がみられ、 251 の遺伝子が WKY/Izm と比べて、 SHRSP/Izm において 0.5 倍以下の発現の低下 がみられた。 GSEA による検索で E-box を持つことが明らかになった 1032 個 の遺伝子のうち、 SHR/Izm の腎皮質において mRNA 発現が WKY/Izm 腎皮質 の 2 倍以上増加していたのは 17 遺伝子であった。このうち SHRSP/Izm の腎皮 質においても WKY/Izm 腎皮質の 2 倍以上に mRNA 発現の増加を認めたのは Bcl6 、 Nr1d1 、 Ppat 、 Tef の 4 遺伝子であった。 SHR/Izm 腎皮質において mRNA
発現が WKY/Izm 腎皮質の半分以下に減少していたのは 10 遺伝子で、このうち
SHRSP/Izm 腎皮質においても同様に半分以下に減少していたのは Fxr1 、
Hnrnpa3 、 Npm1 、 Nptx1 、 Plagl1 、 Plbd1 、 Tbl1x 、 Trim46 の 8 遺伝子であっ た。 (表 3 )
同様な解析を腎髄質でも行った。腎髄質において mRNA 発現が WKY/Izm 腎 髄質の 2 倍以上増加していたのは 10 遺伝子であった。このうち SHRSP/Izm の 腎髄質においても WKY/Izm 腎髄質の 2 倍以上に mRNA 発現の増加を認めたの は Nr1d1 、 Tef の 2 遺伝子であった。 SHR/Izm 腎髄質において mRNA 発現が WKY/Izm 腎皮質の半分以下に減少していたのは Transferrin receptor ( Tfrc ) のみで、これは SHRSP/Izm 腎髄質においては増加していた。 (表 3 )
3. TCMK-1 細胞における生体内時計遺伝子 mRNA 発現振幅の誘導
サブコンフルエントの培養 TCMK-1 細胞に 0.5 μ M デキサメサゾンを添加し、
2 時間培養後デキサメサゾンを含まない培地に交換し、デキサメサゾン添加から
4 時間毎に mRNA を回収し、 Per1 、 Nr1d1 、 Bmal1 、 Cry1 の mRNA 発現変
18
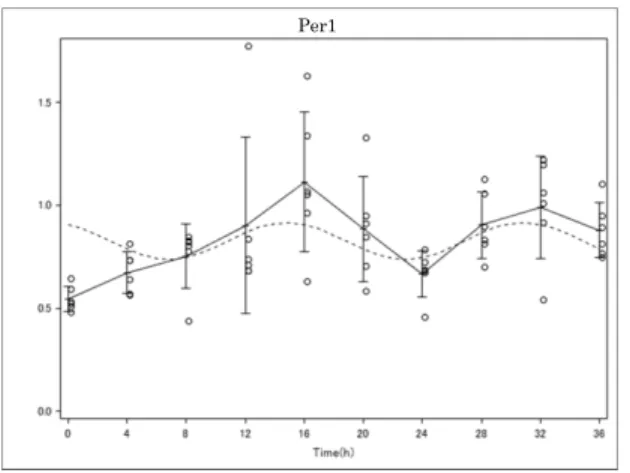
動を検討した。 Per1 、 Bmal1 、 Nr1d1 はデキサメサゾン添加 4 時間後、 28 時間 後、 44 時間後にピークを示す mRNA 発現振幅が誘導された。 Cry1 では 4 時間 後、 28 時間後、 48 時間後にピークを示す振幅であった。これによりデキサメサ ゾン刺激により尿細管培養細胞である TCMK-1 細胞においてデキサメサゾンに より生体内時計遺伝子の mRNA 発現変動が誘導されることが示された。 (図 3 、 4 )
4. デキサメサゾン添加による候補遺伝子 mRNA 発現変動の検討
GSEA 解析およびマイクロアレイ解析により同定した E-box を持ち、高血圧 モデル動物腎臓において発現変動を認めた 12 遺伝子より、すでに生体内時計の サブループを構成することが明らかになっている Nr1d1 を除いた 11 遺伝子に
ついて、 TCMK-1 細胞デキサメサゾン添加による発現振幅の誘導を検討した。
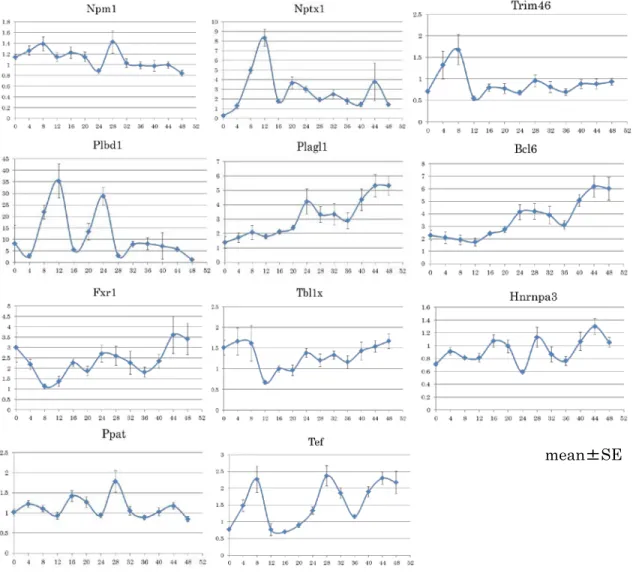
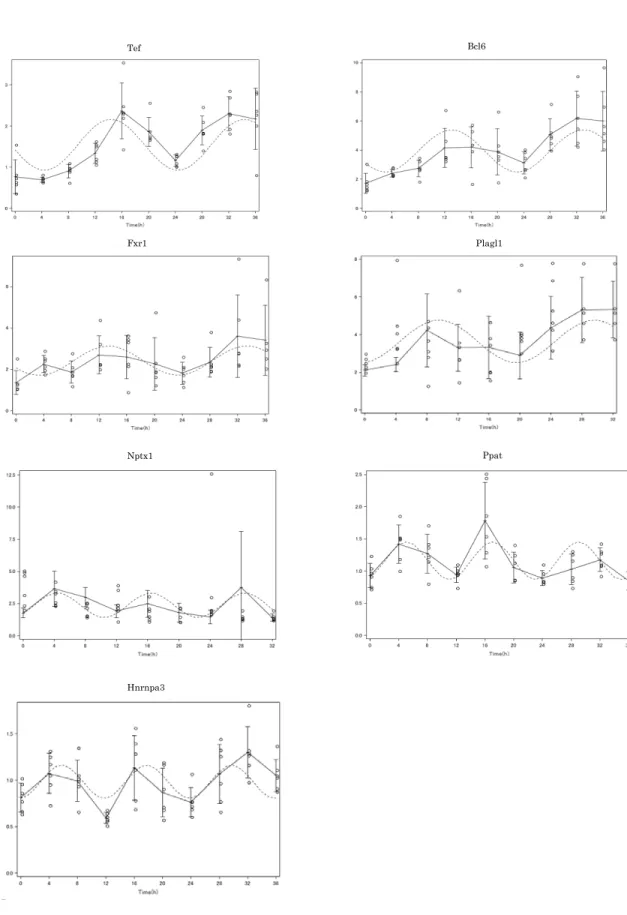
Ppat では時計遺伝子と同様にデキサメサゾン刺激後、 28 時間でピークを形成 し、その後発現減少をして、 44 時間で再び発現増加をしており、 Hnrnpa3 では 16 時間、 28 時間、 44 時間でピークを形成し、 Per1 と比較的近似した振幅を示 した。 Npm1 では Per1 と同様に、 28 時間での発現増加がみられた。 Fxr1 では 24 時間で上昇し、その後減少した後、 44 時間でピークとなり、 Per1 に近似し ており、概日リズムの形成が示唆された(図 5 ) 。
これらの変動の統計学的有意性を検討するために周期回帰分析を行った。ま ず、すでに生体内時計遺伝子として知られている、 Per1 、 Cry1 、 Nr1d1 、 Bmal1
の TCMK1 細胞におけるデキサメサゾン負荷後の発現振幅の有意性を周期回帰
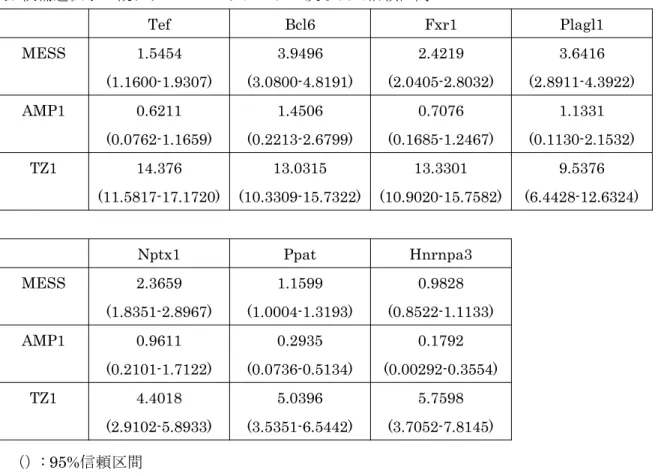
分析して解析し、 Cry1 、 Bmal1 において有意な周期性が示された( P<0.05 ) (表
4 、 5 ) (図 6 ) 。候補遺伝子である 11 の遺伝子に対して、周期関数へのフィッテ
19
ィングを試みたが、 Plbd1 、 Tbl1x 、 Trim46 、 Npm1 はフィッティングができな かった。残りの 7 遺伝子についての周期回帰分析の結果を表 7 に示す。 Fxr1 、
Ppat のみが P<0.05 の有意な回帰を示し、デキサメサゾン負荷にて周期性を持
った mRNA 日内変動が誘導されていることが示された(図 7 ) 。表 6 には周期 回帰分析で得られた概日リズムパラメータを示す。 Fxr1 では周期、振幅ともに 大きな波を示し、 Ppat は周期、振幅ともに小さく、 ultradian rhythm を示した。
5. TCMK-1 細胞における PPAT 、 FXR1 の細胞内での局在
TCMK-1 細胞を抗 PPAT 抗体、抗 FXR1 抗体を用いて、免疫蛍光染色を行っ
た。抗 PPAT 抗体を用いた染色では PPAT は細胞質と核に染色された。抗 FXR1 抗体を用いた染色では FXR1 は細胞質のみに染色された(図 8 、 9 ) 。
6. WKY/Izm 、 SHR/Izm 腎臓における PPAT 、 FXR1 の局在
5 週齢の WKY/Izm 、 SHR/Izm の腎臓組織を抗 PPAT 抗体、抗 FXR1 抗体を 用いて免疫染色を行った。 WKY/Izm 、 SHR/Izm ともに PPAT は近位尿細管、
遠位尿細管の核、細胞質に染色された。 FXR1 は近位尿細管、遠位尿細管の細胞 質で染色された。 WKY/Izm 、 SHR/Izm の間で染色性、局在の差は明らかでは なかった(図 10 、 11 )。
7 . Western blot 法による WKY/Izm 、 SHR/Izm 腎髄質における FXR1 、 PPAT
蛋白発現の検討。
20
5 週齢の WKY/Izm 、 SHR/Izm 腎髄質を用いて、 Western blot 法にて FXR1
発現量を検討した。 FXR1 は WKY/Izm に比べて SHR/Izm において有意に発現
が低下していたが、 PPAT 発現に有意差はみられなかった(図 12 、 13 ) 。
21
【
考察】
高血圧症は罹患率の極めて高い疾患であり、心血管合併症や慢性腎臓病のリ スクが上昇する。血圧は概日リズムを形成しており、その乱れが心血管合併症 や慢性腎臓病のリスクを上昇させる。慢性腎臓病患者が血圧変動異常をきたす ことも知られており、腎臓が血圧概日リズムの形成にかかわっている可能性が 示唆される。このような理由から本研究においては高血圧において重要な役割 を果たす腎臓に注目して、生体内時計に制御を受けて血圧概日リズム形成を担 う分子の同定を目指した。
生体内時計が 24 時間の時を刻む機序が明らかになっている。生体内時計は多 数の生体内時計蛋白の協調によりその概日リズムが構成される。その中心とな るのが BMAL1 、 CLOCK の 2 つの時計蛋白である。この 2 つの蛋白は転写因子 であり、ヘテロダイマーを形成して E-box 配列に結合する。この生体内時計の メインループに制御されて概日リズムを刻む遺伝子のプロモーターには E-box が必ず存在する。それゆえ、我々は GSEA 解析を行い、全ゲノム上で転写開始 点から上流 2000bp または下流 2000bp に E-box を持つ遺伝子をスクリーニング して 1032 個の候補遺伝子を得た。これらの遺伝子は BMAL1/CLOCK ヘテロダ イマーが結合する配列を持つ生体内時計により概日リズムを刻んでいる遺伝子 の候補であると考えられる。尚、 GSEA はヒトゲノムのデータベースであるが、
今回の研究成果をヒトに対して臨床応用することが目的であるため GSEA を用 いることとした。
腎臓における血圧概日リズムに関わる分子の同定のため、我々は高血圧自然
発症モデルである SHR/Izm と SHR/Izm から系統樹立されたさらなる高血圧を
呈し、塩分負荷にて高率に脳卒中を発症する SHRSP/Izm の腎臓の遺伝子発現
22
を DNA マイクロアレイを用いて網羅的に検索した。 GSEA 解析により E-box を持つことが明らかとなった 1032 遺伝子のうち、 SHR/Izm の腎臓でコントロ
ールの WKY/Izm と比較して 2 倍以上または 0.5 倍以下に発現変動していた遺
伝子を生体内時計の制御を受け、腎臓において血圧と関連している遺伝子とし て抽出した。さらに SHR/Izm と SHRSP/Izm という 2 つの同系統であるが異な る高血圧モデルの腎臓において同じ傾向で変動している遺伝子に絞り込むこと で、抽出された遺伝子が血圧に関連する可能性が高まると考えられた。こうし て、 Bcl6 、 Nr1d1 、 Ppat 、 Tef 、 Fxr1 、 Hnrnpa3 、 Npm1 、 Nptx1 、 Plagl1 、 Plbd1 、 Tbl1x 、 Trim46 の 12 遺伝子は E-box 配列を持ち、 2 つの異なる高血圧 モデルである SHR/Izm 、 SHRSP/Izm の腎臓で同じ傾向の発現変動をしている ことから、高血圧に関連し、生体内時計により発現制御されている遺伝子であ る可能性が示唆された。このうち Nr1d1 は Reverse erythroblastosis virus- α
( Rev-erb α)とも呼ばれ、すでに生体内時計のサブループを形成することが明
らかになっている。そのため、本研究の目的が生体内時計に概日リズムを詳説 し、血圧に関連している分子を同定することであるので、この時点で Nr1d1 を 除く 11 遺伝子が候補となった。
生体内時計の研究には困難を伴う。生体内時計遺伝子の概日リズムを評価す るためには個体から数時間間隔で mRNA を得なければならず、動物モデルを用 いるとしてもその数が非常に多くなり、手技的、経済的に困難な研究となる。
それゆえ、 in vitro の系において生体内時計を評価する系を確立することは生体
内時計研究を加速すると考えられる。すでに cell line の培養ラット線維芽細胞
にデキサメサゾン負荷を行うと、生体内時計遺伝子発現の振幅が再現されるこ
とが報告されている
(21)。さらに、培養細胞に高濃度ウマ血清を負荷しても同様
に生体内時計遺伝子の発現振幅が誘導されることも報告されている
(22)。これら
23
は末梢時計の振幅を再現するものと考えられている。末梢組織において時計遺 伝子発現の概日リズムを再現する方法として、上記の高濃度血清、糖質コルチ コイドが挙げられているが、再現できる理由として、何らかの血清因子が末梢 組織における遺伝子振動に影響を与える事、糖質コルチコイドの分泌そのもの が日内変動を示しており、ほぼすべての末梢細胞に Glucocorticoid receptor ( GR ) が発現しているが、中枢時計のある視交叉上核には見られず、糖質コルチコイ ドが GR に作用することで末梢組織の遺伝子振幅が引き起こされる事などが挙 げられる。肝細胞における GR を特異的に不活性化した GR
alfpCreマウスにおい て、デキサメサゾン刺激を加えた際に時計遺伝子である Per1 の発現が肝細胞に おいて発現低下し、 DBP においてはピークのずれが生じたが、 Per2 、 Per3 、
Cry1 、 Rev-erb αでは差を認めないことが示された。これらのことはデキサメサ
ゾンが遺伝子振幅を誘導するが、単一の絶対因子ではないこと示している
(22)。 本研究では、マウス尿細管上皮細胞( TCMK-1 )にデキサメサゾン 0.5 μ M を負 荷することで生体内時計遺伝子である Per1 、 Cry1 、 Nr1d1 mRNA の振幅を誘 導することに成功した。さらに、これらの発現振幅に対して周期回帰分析を行 ったところ、 Bmal1 、 Cry1 では有意な概日リズムを形成していた。すでに報告 されている培養ラット線維芽細胞にデキサメサゾンを負荷し、時計遺伝子振幅 を再現した実験系
(21)と比較すると、 Per1 では刺激後 4 時間、 20-24 時間、 44 時間での発現増加、 Cry1 では 0-8 時間、 24-32 時間で発現増加、 Nr1d1 では 0-4
時間、 16-24 時間、 40-44 時間で発現増加が見られており、本研究における結果
と近似しており
(21)、この TCMK-1 細胞に 0.5 μ M デキサメサゾン負荷系が in
vitro での腎臓末梢時計研究における有用なツールとして使用できることが示唆
された。本研究では cell line の培養マウス尿細管上皮細胞である TCMK-1 細胞
を用いた。使用する培養細胞はラットまたはヒト由来のものを検討したが最終
24
的に適切に実験に供することができる細胞がなく、 TCMK-1 細胞を選択した。
過去の報告でも cell line を用いて生体内時計の振幅を検討しており
(21)、許容 される系であると考えているが、やはり不死化した cell line は組織内の正常細 胞と異なる部分もあり、さらに他の primary culture の細胞で同様の検討が可能 であれば行うべきであると考えられる。
本研究において、 TCMK-1 − 0.5 μ M デキサメサゾン系を用いて 11 の候補遺伝 子について振幅誘導を試みたところ、 Fxr1 、 Ppat の 2 つの遺伝子において、そ の振幅の誘導に成功した。この Ppat 、 Fxr1 は高血圧モデルラットの腎臓におい て発現が大きく変動しており、 E
—box を持ち、デキサメサゾン負荷により、そ の発現振幅が誘導された。さらに免疫染色により PPAT は尿細管の核と細胞質 に、 FXR1 は尿細管の細胞質に発現していることも確認された。これらの結果は、
この 2 つの蛋白が腎臓末梢時計により制御され、血圧概日リズム形成に関わる 可能性のある候補であることを示している。さらに、 Western blot 法において、
FXR1 は SHR/Izm 腎髄質において発現低下が確認され、 FXR1 の尿細管におけ
る発現低下が血圧変動異常に起因している可能性が示唆された。 PPAT は
WKY/Izm と SHR/Izm の腎髄質において、その蛋白発現に有意な違いを認めな
かった。今回の検討は 12 時に屠殺したラットを用いて解析した。生体内時計に 制御され、発現変動する蛋白は検討する時間により発現量が異なることが予想 され、時間帯により両ストレインに発現量の違いが認められる時間帯と認めら れない時間帯があることも想定できる。今後さらに異なる時間帯による両スト レインの腎髄質における PPAT 蛋白発現を検討することも重要であると考えら れる。
Fxr1 は脆弱 X 症候群における原因遺伝子と考えられている Fragile X mental
retardation gene 1 ( Fmr1 )と同じ遺伝子ファミリーに属している。 FXR1 は
25
FMR1 、 FXR2 と相互作用する RNA 結合蛋白であり、主に 60S サブユニットと 結合する。さらに核局在化シグナルと核排出シグナルを有していることから、
細胞質、核の往復が出来ると推測されている
(23, 24, 25, 26)。 Fxr1 の具体的な機能 ははっきりわかってはいないが、心筋や骨格筋に特異的なアイソフォームが存 在しており、 Fxr1 ノックアウトマウスにおいて、生後間もなく心筋や呼吸筋の 異常発育が引き起こされ、死に至ったと報告されている
(26)。 Fxr1 の上流遺伝 子は明らかではないが、心筋における細胞接着に寄与する desmoplakin や talin2 mRNA を制御していると報告されている
(27)。ヒトにおいて、 Fxr1 の骨 格筋における異常発現が顔面肩甲上腕型筋ジストロフィーを引き起こしたとい う報告もあり
(28)、筋組織の発達に重要な役割を果たしている可能性が示唆され るが、 Fxr1 が腎臓や血圧調節に関連しているという報告はない。しかし、マウ スの血管平滑筋の細胞質に発現し、他のタンパク質と相互作用して、細胞の成 長や増殖を制御しているという報告があり
(29)、本研究において SHR/Izm 、
SHRSP/Izm の腎臓にて発現低下し、さらに尿細管細胞質に発現していることか
ら、同じ中胚葉起源の尿細管細胞質に発現し、尿細管細胞の増殖や成長に関与 することも考えられる。
Ppat にコードされる蛋白質は、プリンヌクレオチド代謝において、ホスホリ
ボシルピロリン酸を 5- ホスホリボシルアミンに変換するうえで作用するアロス
テリック酵素である。プリン代謝経路においてリボース -5- リン酸からホスホリ
ジルピロリン酸を経て、中間体であるイノシン酸を合成し、イノシン酸からグ
アニン、アデニンを合成する経路と、尿酸に分解する経路に分かれる
(30)。 PPAT
はドーナツ型のホモ四量体を形成しており、プリンヌクレオチドの結合で不活
性化ホモ四量体となり、ホスホリボシルピロリン酸が結合することで活性型ホ
モ二量体となって、プリン代謝経路における De novo 経路を制御している
(31)。
26
Ppat の 下 流 遺 伝 子 は 明 ら か で な い が 、 シ ョ ウ ジ ョ ウ バ エ に お い て DNA replication-related element binding factor (DREF) が転写因子として働き、
Ppat コード領域の上流に結合することで、 Ppat を制御していると報告されてい る
(32)。 PPAT の合成が促進されると、プリン体から分解される尿酸が増加する。
プリン体の生合成及び分解は全身の細胞で行われている。本研究において、
WKY/Izm に比べ、 SHR/Izm 、 SHRSP/Izm 腎臓において発現上昇がみられ、
PPAT は尿細管細胞の細胞質、核に局在していた。尿酸はそれ自体が抗酸化作用 をもつが、強い血管傷害性も有しており、動脈硬化の独立した危険因子となる。
本研究は生体内時計に制御され、血圧調節に関連する分子を配列解析、発現 解析、培養細胞実験にて絞り込む新しい試みである。しかしながら候補分子で ある FXR1 、 PPAT が実際にどのような機序で血圧調節に関わるのか、現在は明 らかではない。今後、細胞でのノックアウト、ノックイン実験、実験動物での 同様の研究などで明らかにすることが必要と考えられる。また、候補分子を絞 り込む過程において選択されなかった分子でも、周期回帰分析で統計学的有意 差は見られなかったものの、ある程度周期性を持った振幅が見られた分子もあ り、これらについても検討してゆくことも興味深い。
本研究において Fxr1 、 Ppat が生体内時計に制御され、血圧調節に関連するも のであることが示唆された。また今後ラット腎臓における Fxr1 、 Ppat の遺伝子 発現の継時的変動を測定して確認することがさらなる結果の検証に必要である。
これらの分子の作用機序を明らかにすることが期待され、さらにその制御によ
る、血圧変動異常の時間生物学的な治療のターゲットともなると考えられる。
27
【結語】
本研究において培養尿細管上皮細胞をデキサメサゾン刺激し、生体内時計遺 伝子および生体内時計により概日リズムが形成されている遺伝子の振幅を再現 し、高血圧症において重要な役割を果たす腎臓に関する時間生物学的な解析を 行うことが可能となった。さらに、時計遺伝子結合配列である E-box を持ち、
高血圧自然発症ラット腎臓において発現増加または発現減少をしている遺伝子
の中で、培養実験系で発現振幅を形成した Ppat 、 Fxr1 は腎臓において血圧概日
リズム形成に関与する分子であると考えられた。これらの分子の腎臓における
血圧調整の機序を解明することは、血圧概日リズム形成の機序を解明すること
に繋がると考えられる。さらに、これらの分子は時間生物学的な新しい治療タ
ーゲットにもなりうると考えられる。
28
【謝辞】
本研究にあたり、研究全般において御指導いただきました日本大学医学部内
科学系腎臓高血圧内分泌内科学分野 上野高浩先生に心から感謝致します。そ
して、研究全般を支えていただきました日本大学医学部内科学系総合内科・総
合診療分野 相馬正義教授、日本大学総合科学研究所 福田昇教授、日本大学
医学部内科学系腎臓高血圧内分泌内科学分野 阿部雅紀教授に心から感謝いた
します。また、多くの御援助をいただきました日本大学医学部内科学系腎臓高
血圧内分泌内科学分野 常見明子さん他、御協力いただきました皆様に深く感
謝いたします。
Per1 Mm00501813_m1 Bmal1 Mm00500226_m1 Cry1 Mm00514392_m1 Rev-erbα Mm00520708_m1 Npm1 Mm02391781_g1 Hnrnpa3 Mm00817148_g1 Tef Mm00457513_m1
Tbl1x forward(5' to 3') AAG ATG AGC ATT ACC AGC GAC G reverse(5' to 3') TGT GAC TTT CGA TCC CAA ACG T Plagl1 forward(5' to 3') CTG ATG AGA CAC ATG GCC ACA C
reverse(5' to 3') ATC GTC ACA CGC GTA GGA GAT C Plbd1 forward(5' to 3') CTG GTC CCC TGA ATC GAA GAA
reverse(5' to 3') CCC AGC CTG TGG TTT TAA TGG Nptx1 forward(5' to 3') AGG CAG CAT CAG CTT TGA CAA T
reverse(5' to 3') GCC GAC CCA TCA GGA AAC TTA Fxr1 forward(5' to 3') ATA ATT GGC AAC CAG AAC CGC C
reverse(5' to 3') TGG CTC TTG GTC ATT TGC TCT T Bcl6 forward(5' to 3') ATG CTG ATG CCC CAT GAC AT
reverse(5' to 3') GCA AAA GCT CTG CTC TCA CAC C Ppat forward(5' to 3') GTC AGC AGC TGG CAA TTG AAG
reverse(5' to 3') CCC ACA CTT TGT TGC ATA TCC C Trim46 forward(5' to 3') AGC TGC ATA ACA GGA TTG CCC
reverse(5' to 3') TCA ACA GCT TCA TCT CAC GCC 表1 プライマーリスト
TaqMan probe(Applied biosystems)
SYBR Green法(Invitrogen)