アカパンカビの細胞壁構築を制御するシグナル伝達
経路に関する研究
著者
亀井 誠之
学位授与大学
東洋大学
取得学位
博士
学位の分野
生命科学
報告番号
32663甲第364号
学位授与年月日
2014-03-25
URL
http://id.nii.ac.jp/1060/00006736/
2013年 度
東 洋 大 学 審 査 学 位 論 文
ア カ パ ン カ ビ の 細 胞 壁 構 築 を 制 御 す る
シ グ ナ ル 伝 達 経 路 に 関 す る 研 究
生 命 科 学 研 究 科 生 命 科 学 専 攻 博 士 後 期 課 程
第 3学 年
4910110002
亀 井 誠 之
ア カ パ ン カ ビ の 細 胞 壁 構 築 を 制 御 す る
シ グ ナ ル 伝 達 経 路 に 関 す る 研 究
生 命 科 学 研 究 科 生 命 科 学 専 攻 博 士 後 期 課 程
目 次
序 論 3 第 1 章 M A P キ ナ ー ゼ M A K - 1 と M A K - 2 の 細 胞 壁 構 築 へ の 関 与 1 - 1 . 緒 言 9 1 - 2 . 材 料 と 方 法 1 3 1 - 3 . 結 果 と 考 察 1 - 3 - 1 . M A P キ ナ ー ゼ 破 壊 株 の 細 胞 壁 ス ト レ ス 感 受 性 2 9 1 - 3 - 2 . M A K - 1 と M A K - 2 の リ ン 酸 化 解 析 3 4 1 - 3 - 3 . 細 胞 壁 合 成 酵 素 遺 伝 子 の 発 現 解 析 3 8 1 - 3 - 4 . M A K - 1 と M A K - 2 に よ り 制 御 さ れ る 遺 伝 子 の 同 定 4 1 1 - 3 - 5 . M A K - 1 と M A K - 2 の 細 胞 内 局 在 解 析 4 3 第 2 章 浸 透 圧 応 答 経 路 の M A P キ ナ ー ゼ OS -2 に よ る 細 胞 壁 関 連 遺 伝 子 の 発 現 制 御 2 - 1 . 緒 言 4 9 2 - 2 . 材 料 と 方 法 5 2 2 - 3 . 結 果 と 考 察 2 - 3 - 1 . f l u d i o x o n i l 処 理 に よ り 誘 導 さ れ る C W I 関 連 遺 伝 子 の ア レ イ 解 析 5 5 2 - 3 - 2 . O S 経 路 で 制 御 さ れ る C W I 関 連 遺 伝 子 の 同 定 5 6 2 - 3 - 3 . b e t a - 1 , 3 - g l u c a n o s y l t r a n s f e r a s e g e l 遺 伝 子 の 探 索 と 相 同 性 解 析 5 7 2 - 3 - 4 . g e l 破 壊 株 の 形 質 解 析 6 0 2 - 3 - 5 . g e l 破 壊 株 の 細 胞 壁 損 傷 剤 に 対 す る 感 受 性 6 2 2 - 3 - 6 . g e l 遺 伝 子 の 基 礎 発 現 量 の 比 較 6 32 - 3 - 7 . g e l 破 壊 株 に お け る 細 胞 壁 合 成 遺 伝 子 の 発 現 解 析 6 4 2 - 3 - 8 . g e l の f l u d i o x o n i l 誘 導 性 と O S 経 路 依 存 性 6 5 2 - 3 - 9 . g e l の m i c a f u n g i n 誘 導 性 と M A K - 1 及 び M A K - 2 依 存 性 6 7 第 3 章 M A P キ ナ ー ゼ M A K - 1 が 制 御 す る 転 写 調 節 因 子 の 同 定 3 - 1 . 緒 言 6 9 3 - 2 . 材 料 と 方 法 7 1 3 - 3 . 結 果 と 考 察 3 - 3 - 1 . 破 壊 株 を 利 用 し た M A K - 1 制 御 転 写 調 節 因 子 の ス ク リ ー ニ ン グ 7 3 3 - 3 - 2 . m s n - 1 及 び r l m - 1 破 壊 株 の 細 胞 壁 損 傷 に 対 す る 感 受 性 7 9 3 - 3 - 3 . 転 写 調 節 因 子 M S N - 1 が 制 御 す る 遺 伝 子 の 同 定 8 0 3 - 3 - 4 . 転 写 調 節 因 子 R L M - 1 が 制 御 す る 遺 伝 子 の 同 定 8 3 総 合 考 察 及 び 結 論 8 5 参 考 文 献 9 4 謝 辞 11 6
序 論
糸 状 菌 の 細 胞 は 強 固 な 細 胞 壁 を 有 す る 。糸 状 菌 の 細 胞 壁 は 、a l p h a -1 , 3 -グ ル カ ン 、 b e t a -1 , 3 - グ ル カ ン 、 キ チ ン や マ ン ナ ン な ど の 多 糖 類 と 糖 タ ン パ ク 質 な ど が 複 雑 に 入 り 組 ん で 構 成 さ れ た 構 造 体 で あ る ( Fi g . 1 ) ( Kl i s , 1 9 9 4 ; Kl i s e t a l . , 2 0 1 0 ; H a r t l a n d e t a l . , 1 9 9 4 ; B o w m a n e t a l . , 2 0 0 6 ; L a t g e , 2 0 0 7 ; F r e e , 2 0 1 3 ) 。 真 核 生 物 の 細 胞 壁 は 、 細 胞 の 保 護 や 細 胞 形 態 の 維 持 の 役 割 だ け で な く 、 細 胞 外 の 栄 養 環 境 、 浸 透 圧 や 細 胞 壁 損 傷 な ど の 環 境 ス ト レ ス に よ る 刺 激 を 感 知 し 、 細 胞 内 へ 情 報 を 伝 達 す る こ と に も 寄 与 す る ( S e n g a r e t a l . , 1 9 9 7 ; Le vi n , 2 0 0 5 ) 。 細 胞 壁 は 動 物 に は 存 在 し な い 構 造 体 で あ り 、 糸 状 菌 の 細 胞 壁 は セ ル ロ ー ス や リ グ ニ ン を 主 成 分 と す る 植 物 の 細 胞 壁 と は 異 な る こ と か ら 、 病 原 糸 状 菌 に 対 す る 農 薬 や 抗 真 菌 剤 の 新 た な 創 薬 標 的 と し て 注 目 さ れ て い る 。 F i g . 1 . C e l l w a l l s t r u c t u r e o f f i l a m e n t o u s f u n g u s .糸 状 菌 類 は 、 出 芽 や 分 裂 に よ る 増 殖 を 繰 り 返 す 酵 母 類 と は 大 き く 異 な り 、 無 性 胞 子 の 発 芽 管 形 成 、 菌 糸 の 伸 長 ・ 分 岐 ・ 融 合 、 気 中 菌 糸 形 成 、 子 嚢 胞 子 形 成 な ど の 多 岐 に 亘 る 多 様 な 形 態 形 成 を 伴 う 生 活 環 を 有 す る ( F u e t a l . , 2 0 11 ; B o r k o v i c h e t a l . , 2 0 0 4 ; B o r k o v i c h , E b b o l e . , 2 0 1 0 ) 。 こ の よ う な 形 態 形 成 過 程 に お い て 、 糸 状 菌 は 細 胞 壁 の 多 糖 構 造 を 部 分 分 解 と 生 合 成 に よ っ て 再 構 築 す る 必 要 が あ り 、 そ の 調 節 は 厳 密 に 制 御 さ れ て い る と 考 え ら れ る 。 本 研 究 で は 、 ア カ パ ン カ ビ を 材 料 と し て 、 細 胞 壁 の 合 成 と 再 構 築 の 制 御 機 構 の 解 明 を 目 的 と し て 、 M A P キ ナ ー ゼ お よ び 細 胞 壁 関 連 遺 伝 子 の 発 現 解 析 を 中 心 に 研 究 を 展 開 し た 。 細 胞 壁 の 再 構 築 に は 、 M AP キ ナ ー ゼ が 重 要 な 役 割 を 果 た す こ と が 出 芽 酵 母 で 報 告 さ れ て い る こ と か ら 、 ま ず 、 M AP キ ナ ー ゼ と そ の シ グ ナ ル 伝 達 経 路 に つ い て 概 説 す る 。 生 物 は 多 種 多 様 な ス ト レ ス 環 境 に 曝 さ れ る が 、 外 環 境 か ら の 刺 激 に 適 応 す る た め に 、 細 胞 外 ス ト レ ス の 認 識 と そ れ に 対 応 し た 反 応 を つ な ぐ 制 御 系 と し て 、 様 々 な 細 胞 内 シ グ ナ ル 伝 達 経 路 を 有 す る 。 m i t o g e n - a c t i va t e d p r o t e i n k i n a s e ( M A P K , M A P キ ナ ー ゼ ) は 、 哺 乳 類 か ら 植 物 や 真 菌 類 に 至 る ま で 、 真 核 生 物 に お い て 広 く 保 存 さ れ た セ リ ン / ス レ オ ニ ン キ ナ ー ゼ で あ る 。 M AP キ ナ ー ゼ は 、 M A P K K キ ナ ー ゼ 及 び M A P K キ ナ ー ゼ と 共 に M A P キ ナ ー ゼ カ ス ケ ー ド を 構 成 し て お り (K o s a k o e t a l . , 1 9 9 2 ; M a t s u d a e t a l . , 1 9 9 2 )、 そ の リ ン 酸 化 ( 活 性 化 ) は 厳 密 に 調 節 さ れ て い る 。 細 胞 は 栄 養 環 境 や 細 胞 外 か ら の 様 々 な 刺 激 あ る い は 環 境 変 化 か ら く る ス ト レ ス な ど の シ グ ナ ル を 受 容 す る と 、 M AP キ ナ ー ゼ が リ ン 酸 化 さ れ て 細 胞 核 へ と 移 行 し 、転 写 調 節 因 子 を 活 性 化 し て 、外 環 境 の 変 化 に 応 じ た 様 々 な 遺 伝 子 の 発 現 を 制 御 す る 。 こ の よ う に 、 M AP キ ナ ー ゼ を 構 成 要 素 と す る シ グ ナ ル 伝 達 経 路 は 、 細 胞 の 恒 常 性 維 持 、 ス ト レ ス 応 答 、 細 胞 増 殖 や 細 胞 分 化 の 制 御 に お い て 重 要 な 役 割 を 果 た し て い る( R a y e t a l . , 1 9 8 7 ; P o s a d a e t a l . , 1 9 9 2) 。 真 菌 類 の M AP キ ナ ー ゼ に 関 す る 研 究 は 、 出 芽 増 殖 を 繰 り 返 す シ ン プ ル な 形 態
形 成 を 示 す 酵 母 細 胞 を 用 い た 研 究 が 最 も 進 ん で お り 、 M AP キ ナ ー ゼ が 酸 化 ス ト レ ス や 浸 透 圧 ス ト レ ス な ど の ス ト レ ス 応 答 に 深 く 関 与 す る こ と が 知 ら れ て い る ( M i l l a r 1 9 9 9 ; S t a l e v a e t a l . , 2 0 0 4 ; R o b e r t s o n e t a l . , 2 0 0 8 ; C h e n e t a l . , 2 0 0 7 ; C o r r e i a e t a l . , 2 0 1 0 ) 。 ま た 、 細 胞 の 分 化 に 伴 う 細 胞 壁 構 造 の 変 動 や 物 理 的 あ る い は 化 学 的 分 解 に よ る 細 胞 壁 構 造 の 変 化 を 生 育 環 境 や 損 傷 に 応 じ た 適 切 な 構 造 体 へ と 保 持 す る「 細 胞 壁 の 完 全 性 」( c e l l wa l l i n t e g r i t y: C W I) の 維 持 も 、M AP キ ナ ー ゼ が 深 く 関 与 し て い る ( R i s p a i l e t a l . , 2 0 0 9 ; G a r c i a e t a l . , 2 0 0 9 ; Le v i n , 2 0 11 ) 。 出 芽 酵 母 ( S a c c h a ro m y c e s c e re v i s i a e ) で は 、栄 養 成 長 、低 浸 透 圧 、高 温 環 境 や 細 胞 壁 の 損 傷 に よ っ て C W Iが 損 な わ れ る と 、C W I 経 路 と 呼 ば れ る M AP キ ナ ー ゼ カ ス ケ ー ド を コ ア と し た シ グ ナ ル 伝 達 経 路 が 、 細 胞 壁 生 合 成 と 再 構 築 を 亢 進 さ せ る こ と に よ っ て 、 CW Iを 厳 密 に 制 御 し て い る ( Fi g . 2 ) 。 出 芽 酵 母 の C W I 経 路 で は 、細 胞 壁 に 異 常 が 生 じ る と 、細 胞 膜 に 存 在 す る ス ト レ ス セ ン サ ー ( W s c 1 -3 , M i d 2 , C wh 4 3 ) が こ れ を 感 知 し て 、 細 胞 内 下 流 の R h o 1 ( s m a l l G - p r o t e i n ) を 介 し て P r o t e i n k i n a s e C ( P K C ) が 活 性 化 さ れ る 。活 性 型 P K C は 、 F i g . 2 . C e l l w a l l i n t e g r i t y p a t h w a y i n S . c e re v i s i a e .
下 流 の B c k 1 ( M AP KK キ ナ ー ゼ ) 、 M k k 1 / 2 ( M AP Kキ ナ ー ゼ ) と M p k 1 ( M A P キ ナ ー ゼ ) か ら な る M p k 1 M AP キ ナ ー ゼ カ ス ケ ー ド に 細 胞 壁 損 傷 シ グ ナ ル を 入 力 す る 。 リ ン 酸 化 さ れ た M p k 1 は 活 性 型 と な り 、転 写 調 節 因 子 R l m1 を 活 性 化 す る 。活 性 型 R l m 1 は 細 胞 核 内 へ と 移 行 し て 、 細 胞 壁 関 連 遺 伝 子 群 ( b e t a - 1 , 3 - g l u c a n s y n t h a s e F K S 1 , F K S 2 ; c h i t i n s y n t h a s e C H S 3 ; e n d o - t y p e b e t a - 1 , 3 - g l u c a n a s e B G L 2 ; a l p h a - m a n n o s i d a s e D F G 5 な ど ) の 発 現 を 制 御 す る 。 発 現 誘 導 さ れ た 細 胞 壁 の 生 合 成 に 関 与 す る 酵 素 や 補 強 を 担 う 細 胞 壁 糖 タ ン パ ク に よ っ て 、 細 胞 壁 の 損 傷 に 応 答 し た 細 胞 壁 構 築 が な さ れ る こ と で 、C WI が 厳 密 に 維 持 さ れ て い る ( J u n g e t a l . , 1 9 9 9 ; L e v i n . , 2 0 0 5 , 2 0 1 1 ) 。 こ の よ う に 、 出 芽 酵 母 で は 、 M p k 1 M A P キ ナ ー ゼ を コ ア と す る C W I 経 路 で 細 胞 壁 ス ト レ ス 応 答 が ほ ぼ 完 全 に 説 明 で き る と 考 え ら れ て い る 。 糸 状 菌 類 の C W Iに 関 し て は 、 A s p e rg i l l u s n i d u l a n s の M p k A M AP キ ナ ー ゼ ( 出 芽 酵 母 の C W I 経 路 の M p k 1 M AP キ ナ ー ゼ オ ル ソ ロ グ ) が 糸 状 菌 特 有 の a l p h a 1 , 3 -グ ル カ ン 合 成 酵 素 遺 伝 子 ( a g s A , a g s B ) の 発 現 を 制 御 し て い る こ と が 明 ら か に な っ て い る ( Fu j i o k a e t a l . , 2 0 0 7 ) 。 し か し 、 A . n i d u l a n s の M p k A M AP キ ナ ー ゼ は 、 酵 母 の C W I 経 路 で 制 御 さ れ て い る 細 胞 壁 合 成 の 主 要 酵 素 で あ る キ チ ン 合 成 酵 素 や b e t a -1 , 3 - グ ル カ ン 合 成 酵 素 な ど の 遺 伝 子 群 の 発 現 制 御 に は 関 与 し な い と 報 告 さ れ て い る ( Fu j i o k a e t a l . , 2 0 0 7 ) 。 糸 状 菌 で の 研 究 報 告 は 限 ら れ て い る が 、 糸 状 菌 の C W I の 制 御 機 構 は 出 芽 酵 母 の C W I 経 路 と は 異 な っ て い る 可 能 性 が 示 唆 さ れ て い る 。 一 方 で 、 病 原 糸 状 菌 の 研 究 で は 、 医 真 菌 A s p e rg i l l u s f u m i g a t u s の M k k A ( 出 芽 酵 母 の C W I 経 路 の M k k 1 / 2 オ ル ソ ロ グ ) の 破 壊 株 が 病 原 性 を 低 下 さ せ る こ と ( D i r r e t a l . , 2 0 1 0 ) や 、 植 物 病 原 菌 F u s a r i u m o x y s p o r u m の b e t a - 1 , 3 - g l u c a n o s y l t r a n s f e r a s e の 遺 伝 子 破 壊 が 病 原 性 の 消 失 を 招 く こ と ( C a r a c u e l e t a l . , 2 0 0 5 ) な ど が 報 告 さ れ て い る 。 こ の よ う に 糸 状 菌 の C W I に 関 与 す る 因 子 や 細 胞 壁 合 成 に 関 与 す る 酵 素 を コ ー ド す る 遺 伝 子 の オ ル ソ ロ グ の 欠 損 が 病 原 性 低
下 や 基 礎 生 育 の 減 衰 を 引 き 起 こ す 事 象 が 報 告 さ れ て い る 。し か し 、糸 状 菌 の C W I を 制 御 す る シ グ ナ ル 伝 達 の 全 体 像 に つ い て は 、 ほ と ん ど 解 明 さ れ て い な い 。 そ こ で 本 研 究 で は 、 糸 状 菌 の 細 胞 壁 構 築 を 制 御 す る シ グ ナ ル 伝 達 経 路 を 明 ら か に す る こ と を 目 的 と し た 。 研 究 材 料 と し て 、 糸 状 菌 の モ デ ル 生 物 で あ る ア カ パ ン カ ビ ( N e u ro s p o r a c r a s s a ) を 使 用 し た 。 ア カ パ ン カ ビ は 同 じ 子 嚢 菌 類 に 属 す る 出 芽 酵 母 や 糸 状 菌 A s p e rg i l l u s 属 と 比 べ て 、 植 物 病 原 菌 と 進 化 的 に よ り 近 縁 な 関 係 に あ る 。 遺 伝 解 析 が 可 能 で あ る ア カ パ ン カ ビ は 、 植 物 病 原 菌 で 困 難 な 多 重 遺 伝 子 破 壊 株 の 作 製 が 単 一 破 壊 株 同 士 間 の 交 配 に よ っ て 作 製 す る こ と が で き 、 古 く か ら 遺 伝 学 や 生 化 学 的 研 究 が 盛 ん に 行 わ れ て き た ア カ パ ン カ ビ に は 、 形 態 形 成 や ス ト レ ス に 対 し て 高 い 感 受 性 を 示 す 変 異 株 が 存 在 す る 。 さ ら に 、 ゲ ノ ム 情 報 が 公 開 さ れ て い る こ と か ら 、 遺 伝 子 の 検 索 や 比 較 解 析 を 容 易 に 行 う こ と が で き る ( G a l a g a n e t a l . , 2 0 0 3 ) 。 ま た 、 網 羅 的 破 壊 株 作 製 プ ロ ジ ェ ク ト に よ っ て 、 シ グ ナ ル 伝 達 経 路 の 構 成 因 子 や 転 写 調 節 因 子 を 始 め と す る 遺 伝 子 破 壊 株 の 作 出 が 行 わ れ て い る 。 こ の 網 羅 的 な 遺 伝 子 破 壊 株 の 作 製 に は 、 I n o u e H . ら の 研 究 ( N i n o m i y a e t a l . , 2 0 0 4 ) に よ っ て 見 出 さ れ た k u 変 異 株 を 親 株 と し て 用 い る 高 頻 度 の 相 同 組 み 換 え 法 の 寄 与 す る と こ ろ が 非 常 に 大 き い 。 こ れ ら の 変 異 株 や 破 壊 株 は Fu n g a l G e n e t i c s S t o c k C e n t e r ( FG S C , Ka n s a s C i t y, M O , U S A ) か ら 入 手 す る こ と が 可 能 で あ る 。 こ れ ら の 観 点 か ら 、 植 物 病 原 菌 の 病 原 性 や 形 態 形 成 に 関 与 す る 因 子 の 研 究 に は 、 出 芽 酵 母 や A s p e rg i l l u s と 同 様 に ア カ パ ン カ ビ を 研 究 材 料 と す る こ と の 有 用 性 が 高 い 。 こ れ ら の 状 況 を 踏 ま え 、 本 研 究 は 糸 状 菌 の 細 胞 壁 構 築 を 制 御 す る シ グ ナ ル 伝 達 経 路 を 解 明 す る た め 、 ア カ パ ン カ ビ の M AP キ ナ ー ゼ を 中 心 に 解 析 を 進 め た 。 第 1 章 で は 、 ゲ ノ ム に 存 在 す る 3 種 類 の M AP キ ナ ー ゼ の 細 胞 壁 ス ト レ ス に 対 す る 応 答 を 解 析 し 、M A K- 1 と M A K-2 が C W Iに 関 与 す る こ と を 明 ら か に し た 。さ ら に 、 そ れ ら の 下 流 で 制 御 さ れ て い る 遺 伝 子 の 同 定 を 行 っ た 。第 2 章 で は 、高 浸 透 圧 が
細 胞 壁 に 与 え る 影 響 に 着 目 し 、 浸 透 圧 応 答 を 担 う O S 経 路 の M A P キ ナ ー ゼ O S -2 も ま た 、 M A K -1 と M A K-2 と は 独 立 に C W I に 貢 献 し て い る こ と を 明 ら か に し た 。 第 3 章 で は 、 糸 状 菌 で は こ れ ま で 報 告 が な い 、 MA K-1 が 制 御 す る 転 写 調 節 因 子 の ス ク リ ー ニ ン グ を 行 い 、 M S N -1 を 同 定 し た 。 ア カ パ ン カ ビ を 用 い た 本 研 究 は 、 糸 状 菌 が 共 通 に も つ 3 種 類 の M AP キ ナ ー ゼ の C W Iへ の 貢 献 と 細 胞 壁 関 連 酵 素 遺 伝 子 の 調 節 機 構 を 明 ら か に し た も の で あ り 、 植 物 病 原 菌 を 始 め と す る 糸 状 菌 特 有 の C W I経 路 に よ る 細 胞 壁 構 築 の 制 御 機 構 の 解 明 や 病 原 性 に 関 与 す る 因 子 の 探 索 に 寄 与 す る も の で あ る と 考 え ら れ る 。
第 1章
MAPキ ナ ー ゼ MAK-1 と MAK-2 の 細 胞 壁 構 築 へ の 関 与
1-1. 緒 言
出 芽 酵 母 の c e l l wa l l i n t e g r i t y ( C W I) 経 路 に お い て は 、 細 胞 壁 の 損 傷 に よ っ て M p k 1 ( M A P キ ナ ー ゼ ) が リ ン 酸 化 さ れ て 活 性 型 と な り 、 転 写 調 節 因 子 R l m 1 を 活 性 化 す る 。 活 性 型 R l m1 が 細 胞 壁 関 連 遺 伝 子 群 の 発 現 を 誘 導 し て 、 細 胞 壁 損 傷 に 応 答 し た 細 胞 壁 の 生 合 成 と 再 構 築 が な さ れ る こ と に よ り 、 C W Iは 厳 密 に 制 御 さ れ て い る ( J u n g e t a l . , 1 9 9 9 ; Le v i n , 2 0 11 ) 。 こ れ を も と に 、す で に 様 々 な 糸 状 菌 で CW Iに 関 す る M p k 1 - l i k e M AP キ ナ ー ゼ の 遺 伝 子 破 壊 株 が 作 出 さ れ 、 解 析 が 行 わ れ て い る 。 例 え ば 、 イ ネ い も ち 病 菌 ( M a g n a p o r t h e g r i s e a ) で は 、 M p k 1 - l i k e M A P キ ナ ー ゼ で あ る M P S 1 の 破 壊 株 は 植 物 へ の 侵 入 が 出 来 な く な り 、 感 染 能 を 消 失 す る ( X u e t a l . , 1 9 9 8 ) 。 ま た 、 赤 か び 病 菌 ( F u s a r i u m g r a m i n e a r u m ) の M p k 1 - l i k e M A P キ ナ ー ゼ は 、 ト リ コ テ セ ン 系 か び 毒 の d e o x yn i va l e n o l の 蓄 積 に 関 与 し て い る ( H o u e t a l . , 2 0 0 2 ) 。 動 物 へ の 感 染 能 を 有 す る A s p e r g i l l u s f u m i g a t u s で は 、 m p k A ( S . c e r e v i s i a e M P K 1 ホ モ ロ グ ) の 欠 損 は 、 生 育 遅 延 や 形 態 形 成 の 劇 的 な 変 化 だ け で な く 、 細 胞 壁 損 傷 剤 に 対 す る 感 受 性 を 高 め る こ と が 示 さ れ て い る ( V a l i a n t e e t a l . , 2 0 0 8 ) 。 こ れ ら の 病 原 菌 の 研 究 は 、 C W I 経 路 が 病 原 性 あ る い は 二 次 代 謝 制 御 に 強 く 影 響 す る こ と を 示 し て い る が 、 C W I 経 路 の 機 構 の 解 明 に は 至 っ て い な い 。 糸 状 菌 の C W I 経 路 の 解 明 を 目 指 し た 研 究 と し て 、 東 北 大 学 ( 阿 部 敬 悦 研 究 室 ) を 中 心 と し た A s p e rg i l l u s n i d u l a n s を 用 い た 研 究 が あ る 。A. n i d u l a n s の M p k 1 - l i k e M AP キ ナ ー ゼ で あ る M p k A は 、そ の 欠 損 に よ っ て 分 生 子 発 芽 率 や 極 性 生 長 が 減 衰 す る ( B u s s i n k e t a l . , 1 9 9 9 ) 。 さ ら に 、 M p k A は そ の 下 流 の 転 写 調 節 因 子 R l mA ( S . c e re v i s i a e R l m1 ホ モ ロ グ ) を 介 し て 、 糸 状 菌 特 有 の a l ph a -1 , 3 - グ ル カ ン 合 成 酵 素 遺 伝 子 ( a g s A , a g s B ) の 発 現 を 制 御 し て い る 。一 方 で 、細 胞 壁 合 成 に 主 要 な キ チ ン 合 成 酵 素 遺 伝 子 ( c h s C ) や b e t a 1 , 3-グ ル カ ン 合 成 酵 素 遺 伝 子 ( f k s A ) な ど の 細 胞 壁 合 成 遺 伝 子 の 発 現 に は 関 与 し て い な い ( Fi g . 3 ) ( Fu j i o k a e t a l . , 2 0 0 7 ) 。 こ の こ と は 、 出 芽 酵 母 と A . n i d u l a n s で は 、 M p k 1 - l i k e M A P キ ナ ー ゼ の 制 御 す る 遺 伝 子 や そ の 制 御 機 構 に 大 き な 違 い が 存 在 す る こ と を 示 唆 し て い る が 、そ の 全 容 に つ い て は 不 明 の 点 が 多 く 残 さ れ て い る 。 モ デ ル 糸 状 菌 で あ る ア カ パ ン カ ビ は 、 出 芽 酵 母 と の 比 較 ゲ ノ ム 解 析 か ら 、 3 種 類 の M AP キ ナ ー ゼ 経 路 ( O S 、 MA K-1 、 M A K-2 経 路 ) を も つ こ と が 明 ら か に な っ て い る ( Fi g . 4 ) ( G a l a g a n e t a l . , 2 0 0 3 ; B o r k o v i c h e t a l . , 2 0 0 4 ; Ka me i e t a l . , 2 0 1 3 a ) 。 ア カ パ ン カ ビ の M AP キ ナ ー ゼ 経 路 の 各 構 成 因 子 は 、 出 芽 酵 母 の 各 M AP キ ナ ー ゼ 経 路 と 高 い 相 同 性 を 示 し 、真 菌 類 に お い て 高 度 に 保 存 さ れ て い る 。O S 経 路 は O S - 4 ( M A P K K K ) − O S - 5 ( M A P K K ) − O S - 2 ( M A P K ) か ら 構 成 さ れ て お り 、 出 芽 酵 母 の 高 浸 透 圧 応 答 経 路 S s k 2 / 2 2 − P b s 2 − H og 1 M AP K カ ス ケ ー ド の ホ モ ロ グ で あ る 。 同 様 に 、ア カ パ ン カ ビ の M A K-2 経 路 の N R C -1 − M E K-2 − M A K-2 は 、出 芽 酵 母 に お い て 交 配 能 や 形 態 形 成 を 制 御 す る S t e 11 − S t e 7 − Fu s 3 / Ks s 1 と 高 い 相 同 性 を 示 す 。 ま た 、ア カ パ ン カ ビ の M A K-1 経 路 の M I K- 1 − M E K-1 − M A K-1 は 、出 芽 酵 母 の C W I 経 路 の B c k 1 − M k k 1 / 2 − M p k 1 と そ れ ぞ れ 高 い 相 同 性 を 有 す る 。 以 下 に 、 3 種 類 の F i g . 3 . C e l l w a l l i n t e g r i t y p a t h w a y i n A . n i d u l a n s .
M A P キ ナ ー ゼ 経 路 に つ い て の こ れ ま で の 研 究 を 概 説 す る と と も に 、 細 胞 壁 と の 関 係 に つ い て 述 べ る 。 ア カ パ ン カ ビ の O S -2 M AP キ ナ ー ゼ 経 路 は 、 3 種 類 の M AP キ ナ ー ゼ 経 路 の 中 で 最 も 研 究 が 進 ん で お り 、 浸 透 圧 ス ト レ ス 応 答 に お い て 重 要 な 役 割 を 担 う こ と が 明 ら か に な っ て い る ( Zh a n g e t a l . , 2 0 0 2 ; Fu j i mu r a e t a l . , 2 0 0 3 ) 。 O S 経 路 の 3 種 類 の 変 異 株 (o s - 4 , o s - 5 , o s - 2 ) は 浸 透 圧 に 高 い 感 受 性 を 示 し 、 ジ カ ル ボ キ シ イ ミ ド や フ ェ ニ ル ピ ロ ー ル 系 殺 菌 剤 に 対 し て 耐 性 を 示 す 。O S 経 路 は 様 々 な 遺 伝 子 群 を 発 現 制 御 す る こ と に よ り 浸 透 圧 応 答 や 概 日 リ ズ ム の 出 力 経 路 と し て の 一 端 を 担 っ て い る ( N o g u c h i e t a l . , 2 0 0 7 ; V i t a l i n i e t a l . , 2 0 0 7 ; W a t a n a b e e t a l . , 2 0 0 7 ; Y a m a s h i t a e t a l . , 2 0 0 7 ; K a m e i e t a l . , 2 0 1 3 b ) 。 O S 経 路 が C W I に 関 与 す る と い う 報 告 は こ れ ま で な い が 、 浸 透 圧 ス ト レ ス に 応 答 し て 膨 圧 を 制 御 し て お り 、 細 胞 壁 に 物 理 的 ス ト レ ス を 生 じ る 可 能 性 が 考 え ら れ る 。 ア カ パ ン カ ビ の M A K-2 経 路 は 、 m a k - 2 の 遺 伝 子 破 壊 株 が 雌 性 不 稔 か つ 子 嚢 胞 F i g . 4 . T h r e e M A P K c a s c a d e s i n N . c r a s s a .
子 が 発 芽 不 能 で あ る こ と か ら 、 交 配 能 を 制 御 し て い る ( P a n d e y e t a l . , 2 0 0 4 ; Li e t a l . , 2 0 0 5 ) 。 こ の こ と は 、 本 経 路 が 基 本 的 に 出 芽 酵 母 と 同 様 、 有 性 生 殖 過 程 に 関 与 す る こ と を 示 し て い る 。 ア カ パ ン カ ビ に は c o n i d i a l a n a s t o m o s i s t u b e s ( C A T s ) と 呼 ば れ る 発 芽 し た 分 生 子 の 先 端 間 が 融 合 す る 現 象 が 知 ら れ て い る ( R o c a e t a l . , 2 0 0 5 ) 。こ れ は 、受 精 の よ う な 異 な る 交 配 型 遺 伝 子 を も つ 細 胞 間 の 融 合 で は な く 、 無 性 増 殖 過 程 に お い て 、同 じ 遺 伝 的 背 景 を も つ 細 胞 同 士 が 融 合 す る 現 象 で あ り 、 多 核 の 糸 状 菌 で は 一 般 に 認 め ら れ る 現 象 で あ る 。C A T s 先 端 間 の 細 胞 融 合 部 位 で は 細 胞 間 で シ グ ナ ル 交 換 が 行 わ れ て お り 、 M A K-2 は こ の 役 割 を 担 う こ と が 示 さ れ て い る ( Fl e i s s n e r e t a l . , 2 0 0 9 ) 。 細 胞 壁 を も つ 菌 糸 が 融 合 す る た め に は 、 融 合 部 位 に お け る 細 胞 壁 の 部 分 分 解 と 再 合 成 が 必 要 で あ り 、 C W I へ の 関 与 の 可 能 性 が 考 え ら れ る 。 一 方 、 M A K-1 は 、 出 芽 酵 母 と の 比 較 ゲ ノ ム 解 析 か ら C W Iを 制 御 す る こ と が 推 定 さ れ て い る が 、 C W Iへ の 関 連 性 に つ い て は 、 こ れ ま で 明 ら か に な っ て い な い 。ア カ パ ン カ ビ の M A K-1 は t y r o s i n a s e 遺 伝 子 を 負 に 発 現 調 節 す る こ と に よ っ て 二 次 代 謝 系 を 制 御 す る こ と 、 そ し て 概 日 リ ズ ム の 出 力 経 路 と し て の 役 割 を 担 う こ と が 見 出 さ れ て い る ( P a r k e t a l . , 2 0 0 8 ; B e n n e t t e t a l . , 2 0 1 3 ) 。 し か し 、 こ れ ら は 細 胞 壁 構 築 と は 異 な る 現 象 で あ る と 考 え ら れ る 。 ま た 、 M AK- 1 下 流 の 転 写 調 節 因 子 と そ の 制 御 遺 伝 子 に つ い て も 同 定 さ れ て い な い 。 従 っ て 、 ア カ パ ン カ ビ の M A K -1 が 出 芽 酵 母 の よ う に C W Iを 制 御 し て い る か に つ い て は 不 明 で あ る 。 そ こ で 、 本 章 に お い て は 、 ア カ パ ン カ ビ の 3 種 類 の M AP キ ナ ー ゼ の 各 遺 伝 子 破 壊 株 の 評 価 、M AP キ ナ ー ゼ の リ ン 酸 化 解 析 、遺 伝 子 発 現 解 析 に よ り 、3 種 類 の M AP キ ナ ー ゼ の C W Iへ の 関 連 性 に つ い て 検 証 を 行 っ た 。
1-2. 材 料 と 方 法
ア カ パ ン カ ビ の 菌 株 、 培 地 及 び 感 受 性 試 験
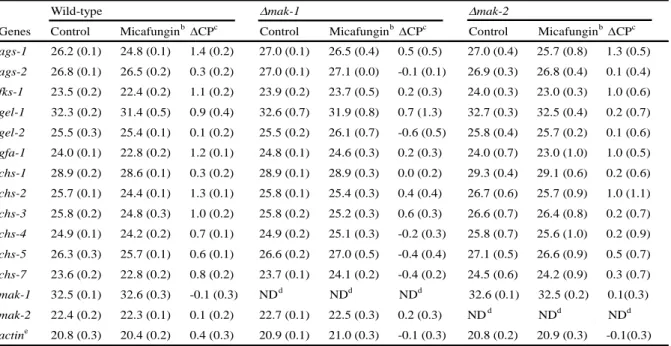
本 章 で 使 用 し た ア カ パ ン カ ビ の 菌 株 を Ta b l e 1 に 記 し た 。 野 生 株 と し て C 1 - T 1 0 - 3 7 株 ( Ta m a r u e t a l . , 1 9 8 9 ) を 使 用 し 、 各 破 壊 株 は 、 網 羅 的 遺 伝 子 破 壊 株 作 成 プ ロ ジ ェ ク ト に よ り 作 出 さ れ た 破 壊 株 ラ イ ブ ラ リ ー か ら 入 手 し た( Funga l G e n e t i c s S t o c k C e n t e r ( F G S C , K a n s a s C i t y, M O , U S A ) ) 。Table 1. The Neurospora crassa strains used in this study.
Strain Genotype Reference or sourse C1-T10-37 A mat A Tamaru et al., 1989 C1-T10-19 a mat a Tamaru et al., 1989 FGSC #11326 mat A; mik-1::Hygr FGSCa FGSC #11327 mat a; mik-1::Hygr FGSCa FGSC #11318 mat a; mek-1::Hygr FGSCa FGSC #11319 mat A; mek-1::Hygr FGSCa FGSC #11320 mat A; mak-1::Hygr FGSCa FGSC #11321 mat a; mak-1::Hygr FGSCa FGSC #18202 mat a; os-4::Hyg; mus-51::Barr FGSCa FGSC #18203 mat a; os-5::Hyg; mus-51::Barr FGSCa FGSC #6041 mat a; os-4 (Y256M223) FGSCb FGSC #4576 mat a; os-5 (NM216o) FGSCb FGSC #17933 mat A; os-2::Hygr FGSCa FGSC #1509 mat A; os-2 (ALS10) FGSCc FGSC #18162 mat a; nrc-1::Hyg; mus-51::Barr FGSCa FGSC #11524 mat a; mek-2::Hyg; mus-51::Barr FGSCa FGSC #11482 mat a; mak-2::Hygr; mus-51::Barr FGSCa FGSC #6103 mat A; his-3 FGSC
a)
Neurospora knockout strains from the functional genomics program (Colot et al., 2006) were
obtained from Fungal Genetics Stock Center.
b, c)
Neurospora mutant strains that were characterized by Perkins D.D. (1974) were obtained
ア カ パ ン カ ビ の 菌 株 の 維 持 と 継 代 培 養 に は 、Vo g e l ’s me d i u m N 最 少 ( V M ) 培 地 (1.2 w/v% suc rose, 1 ×Vogel’s solution )を 使 用 し た (Vogel., 1956)。 分 生 子 (無 性 胞
子)形 成 は 、 グ リ セ ロ ー ル 完 全 培 地 (1.0 v/v% glycero l, 1 ×Vogel’s solu tion, 1.0
v / v % v i t a m i n s o l u t i o n , 2 . 5 g / l y e a s t e x t r a c t , 1 g / l c a s a m i n o a c i d , 5 g / l m a l t e x t r a c t ) を 用 い て 、 植 菌 後 1 0 日 間 培 養 し 、 形 成 さ れ た 分 生 子 を 滅 菌 水 に 懸 濁 後 、 茶 こ し を 使 用 し て 菌 糸 を 除 い た 。 さ ら に 滅 菌 水 で 数 回 繰 り 返 し て 洗 浄 後 、 再 度 、 滅 菌 水 に 懸 濁 し た も の を 分 生 子 溶 液 と し た 。 ま た 、 菌 糸 体 は 上 述 の 方 法 で 調 製 し た 分 生 子 を 、 6 . 6 × 1 05 c e l l s / m l と な る よ う に V M 液 体 培 地 に 植 菌 し 、 室 温 で 1 8 時 間 培 養 後 、 生 育 し た 菌 体 を 吸 引 濾 過 に よ り 回 収 し た 。 集 菌 し た 菌 体 は 、 液 体 窒 素 に よ り 凍 結 し 、 使 用 す る ま で -8 0 ℃ で 凍 結 保 存 し た 。 感 受 性 試 験 に 用 い た 細 胞 壁 損 傷 剤 は 、 b e t a -1 , 3 - グ ル カ ン 合 成 酵 素 阻 害 剤 m i c a f u n g i n ( A s t e l l a s P h a r m a , T o k y o , J a p a n ) 、 キ チ ン 合 成 阻 害 剤 p o l y o x i n D ( K a k e n P h a r m a c e u t i c a l C o . , L t d , T o k y o , J a p a n ) 、キ チ ン 合 成 阻 害 剤 C a l c o f l u o r W h i t e ( C F W ) ( S i g m a A l d r i c h , S t . L o u i s , M O ) 、 グ ル カ ン 合 成 阻 害 剤 C o n g o R e d ( C R ) ( S i g m a A l d r i c h ) 、 細 胞 壁 損 傷 剤 S D S ( s o d i u m d o d e c y l s u l p h a t e ) ( Wa k o P u r e C h e m i c a l , O s a k a , J a p a n ) の 作 用 の 異 な る 5 種 類 の 薬 剤 を 使 用 し た 。 な お 、 m i c a f u n g i n 及 び p o l y o x i n D は そ れ ぞ れ グ ル カ ン や キ チ ン の 生 合 成 阻 害 剤 で あ り 、 C R と C F W は キ チ ン や グ ル カ ン に そ れ ぞ れ 結 合 す る こ と に よ っ て 細 胞 壁 損 傷 を 与 え る 剤 で あ る 。 ま た 、 S D S は 界 面 活 性 剤 で あ り 、 細 胞 壁 成 分 の 溶 解 に よ り 損 傷 を 引 き 起 こ す 。 V M 培 地 を 用 い た 菌 糸 生 育 阻 害 試 験 は 、前 培 養 し た 菌 糸 の 先 端 を コ ル ク ボ ー ラ ー で く り 抜 き 、 そ の 菌 糸 デ ィ ス ク を 薬 剤 含 有 培 地 上 に 接 種 し 、28℃ 、 18時 間 培 養 し た 。 感 受 性 の 判 定 は 、 薬 剤 含 有 培 地 上 で の 生 育 菌 糸 長 を 、 無 処 理 の 菌 糸 生 育 長 と 比 較 し て 生 育 阻 害 率 を 算 出 し た 。 ま た 、 胞 子 希 釈 法 に よ る 感 受 性 試 験 は 、 s o r b o s e 含 有 V M 平 板 培 地 ( 0 . 2 w / v % s u c r o s e , 1 . 0 w / v % s o r b o s e , 1 × Vo g e l ’s s o l u t i o n ) 上 に 、 1 06 個 の 分 生 子 か ら 1 / 1 0 倍 ご と に 段 階 希 釈 し た 胞 子 液 を 5 lず つ ス ポ ッ ト
し 、28℃ 、 48時 間 培 養 し た 。
タ ン パ ク 質 の 抽 出 及 び 定 量
0 . 5 m m 径 ガ ラ ス ビ ー ズ を 加 え た 2 m l ス ク リ ュ ー チ ュ ー ブ に 、 t o t a l c e l l e x t r a c t i o n b u f f e r ( 5 0 m M H E P E S b u ff e r ( p H 7 . 6 ) , 1 0 % g l y c e r o l , 1 3 7 m M N a C l , 2 5 m M N a F, 1 0 m M N a4P2O7, 2 m M N a3V O4, c o m p l e t e p r o t e a s e i n h i b i t o r c o c k t a i l 1 t a b l e t / 5 0 m l ( R o c h e ) ) を 7 0 0 ~ 1 0 0 0l 加 え て 、 ア カ パ ン カ ビ の 凍 結 サ ン プ ル (無 性 胞 子 あ る い は 菌 糸 体 ) を 添 加 し た 。 ビ ー ズ ビ ー タ ー ( M P B i ome d i c a l s , J a p a n , F a s t P r e pT M F P 1 2 0 / B I O 1 0 1 ; 設 定 条 件 - S p e e d : 6 . 0 m / s , Ti m e : 2 0 秒 ) に セ ッ ト し て 菌 体 を 破 砕 し た 。 菌 体 破 砕 サ ン プ ル は 、 遠 心 分 離 ( 1 3 , 0 0 0 g , 1 0 分 間 , 4 ℃ ) を 行 い 、 得 ら れ た 上 清 を 再 度 遠 心 分 離 し 、 そ の 上 清 を t o t a l c e l l e x t r a c t と し た 。 タ ン パ ク 質 濃 度 は 、 B r a d f o r d 試 薬 ( P r o t e i n A s s a y Ki t I / B IO -R A D La b o r a t o r i e s , H e r c u l e s , C A ) を 用 い て 定 量 し た 。ウ ェ ス タ ン ブ ロ ッ テ ィ ン グ 解 析
菌 体 由 来 の t o t a l c e l l e x t r a c t 5 0 gを 2×sa mp le bu ffe r (0.1 M Tris-HCl (pH6.8), 4 % S D S , 1 . 6 9 2 M 2 - m e r c a p t o e t h a n o l , 1 % s u c r o s e , 0 . 0 1 % B P B ) を 用 い て 9 5 ℃, 5分 間 の 熱 処 理 に よ り タ ン パ ク 質 を 変 性 さ せ 、S D S -ポ リ ア ク リ ル ア ミ ド ゲ ル 電 気 泳 動 ( PA G E ) 用 サ ン プ ル と し た 。同 サ ン プ ル を 1 0 % ポ リ ア ク リ ル ア ミ ド ゲ ル に よ り 分 離 ( 電 気 泳 動 条 件 : 2 5 mM Tr i s , 1 9 2 m M g l y c i n e , 0 . 1 % S D S , 定 電 圧 2 0 0 V ) 後 、 n i t r o c e l l u l o s e m e m b r a n e ( A d v a n t e c h C o . , L t d . , To k y o , J a p a n ) 上 に ブ ロ ッ テ ィ ン グ し た ( ブ ロ ッ テ ィ ン グ 条 件 : 2 5 mM Tr i s , 1 9 2 mM g l yc i n e , 2 0 % m e t h a n o l , 定 電 圧 1 0 0 V, 電 流 上 限 3 5 0 m A , 6 0 分 間 ) 。メ ン ブ レ ン を w a s h b u ff e r ( T B S - T: 1 × T B S , 0 . 2 % Tw e e n - 2 0 ) で 5 分 間 洗 浄 後 、 b l o c k i n g b u ff e r ( w a s h b u f f e r, 1 0 % B S A ) を 用 い て 室 温 で 6 0 分 間 、 ま た は 4 ℃ , オ ー バ ー ナ イ ト で 振 盪 し 、 ブ ロ ッ キ ン グ 処 理 を 行 っ た 。
ウ ェ ス タ ン ブ ロ ッ テ ィ ン グ 法 に お け る M AP キ ナ ー ゼ の 免 疫 染 色 は 、 ブ ロ ッ キ ン グ 終 了 後 の メ ン ブ レ ン を 一 次 抗 体 溶 液 ( 各 種 一 次 抗 体 , 1 × wa s h b u ff e r ) に 浸 し 、 室 温 で 2 時 間 あ る い は 、 4 ℃ で オ ー バ ー ナ イ ト 処 理 し た 。 一 次 抗 体 と し て 、 t o t a l M A K - 2 の 検 出 に は a n t i - M A P k i n a s e ( E R K 1 , E R K - 2 ) a n t i b o d y p r o d u c e d i n r a b b i t ( S i g m a A l d r i c h , M 5 6 7 0 ) を 2 0 0 0 0 倍 希 釈 し 、 リ ン 酸 化 M A K - 1 及 び M A K - 2 の 検 出 に は p h o s p h o -p 4 4 / 4 2 M A P K ( E r k 1 / 2 ) ( T h r 2 0 2 / Tyr 2 0 4 ) a n t i b o d y ( C e l l S i g n a l i n g Te c h n o l o g y, D a n v e r s , M A , # 9 1 0 1 ) を 1 0 0 0 倍 希 釈 し て 使 用 し た 。同 様 に 、t o t a l O S - 2 の 検 出 に は H o g 1 a n t i b o d y ( S a n t a C r u z B i o t e c h n o l o g y I n c , y- 2 1 5 , 2 5 0 0 倍 希 釈 ) 、 リ ン 酸 化 O S -2 の 検 出 に は p h o s p h o -p 3 8 M A P K a n t i b o d y ( C e l l S i g n a l i n g T e c h n o l o g y, D a n v e r s , M A , # 9 2 1 1 , 1 0 0 0 倍 希 釈 ) を そ れ ぞ れ 一 次 抗 体 と し て 使 用 し た 。 一 次 抗 体 反 応 処 理 後 、 wa s h b u ff e r で 3 回 洗 浄 し 、 二 次 抗 体 処 理 ( a n t i - r a b b i t Ig G A P c o n j u g a t e a n t i b o d y ( P r o m e g a , F i t c h b u r g , W I , S 3 7 3 1 ) 7 5 0 0 倍 希 釈 , 1 × w a s h b u f f e r ) を 室 温 1 時 間 行 っ た 。 二 次 抗 体 処 理 後 の メ ン ブ レ ン を wa s h b u ff e r で 3 回 洗 浄 し 、 メ ン ブ レ ン の p H を 約 9 . 5 と す る た め 、 AP 発 色 b u ff e r ( 1 0 0 m M Tr i s -H C l ( p H 9 . 5 ) , 1 0 0 m M N a C l , 5 0 m M M g C l2・ 6 H2O ) で 置 換 後 、 遮 光 条 件 下 で A P 発 色 液 ( A P 発 色 b u ff e r, 0 . 3 4 m g / m l N B T ( n i t r o - b l u e t e t r a z o l i u m c h l o r i d e ) , 0 . 1 8 m g / m l B C I P ( 5 - b r o m o - 4 - c h l o r o - 3 ' - i n d o l y l p h o s p h a t a s e p - t o l u i d i n e s a l t ) に よ る 発 色 に よ っ て 各 タ ン パ ク 質 を 検 出 し た 。 適 切 な 発 色 が 得 ら れ た 後 、 氷 冷 し た イ オ ン 交 換 水 で 十 分 に 洗 浄 し 、 発 色 反 応 を 停 止 し た 。
ア カ パ ン カ ビ の RNA抽 出 及 び 調 製
2 m l ス ク リ ュ ー チ ュ ー ブ に 0 . 5 m m 径 ガ ラ ス ビ ー ズ を 2 0 0 μ l 入 れ 、 R N A p r o s o l u t i o n ( M P B i o m e d i c a l s , J a p a n ) を 1 m l 加 え た 。 ア カ パ ン カ ビ の 分 生 子 や 菌 糸 体 な ど の 凍 結 菌 体 サ ン プ ル を 加 え 、 ビ ー ズ ビ ー タ ー ( M P B i o me d i c a l s , J a p a n , F a s t P r e pT M F P 1 2 0 / B I O 1 0 1 ; 設 定 条 件 - S p e e d : 6 . 0 m / s , Ti m e : 4 0 秒 ) で 菌 体 サ ン プ ルを 破 砕 し た 。 菌 体 破 砕 サ ン プ ル は 1 3 , 0 0 0 g で 5 分 間 ( 4 ℃ ) 遠 心 分 離 し 、 上 層 を 新 し い マ イ ク ロ チ ュ ー ブ へ 移 し た 。 さ ら に 、 室 温 で 5 分 間 イ ン キ ュ ベ ー ト し た 後 、 ク ロ ロ ホ ル ム 3 0 0 μ l を 添 加 し て ボ ル テ ッ ク ス で 1 0 秒 間 混 合 し た 。 再 度 、 室 温 で 5 分 間 イ ン キ ュ ベ ー ト し 、遠 心 分 離 ( 1 3 , 0 0 0 g , 5 分 間 , 4 ℃ ) 後 の 上 清 を 新 し い マ イ ク ロ チ ュ ー ブ に 移 し た 。 こ の 操 作 を 再 度 繰 り 返 し た R NA を 含 む 溶 液 に 、 9 9 . 5 % エ タ ノ ー ル を 2 . 5 倍 量 添 加 し た 。5 回 転 倒 混 和 後 、-2 0 ℃ 下 に 3 0 分 間 以 上 静 置 し 、1 3 , 0 0 0 g で 1 5 分 間( 4 ℃ ) 遠 心 分 離 し た 。 デ カ ン テ ー シ ョ ン で 上 清 を 除 去 後 、 8 0 % エ タ ノ ー ル で リ ン ス し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 2 分 間 、 4 ℃ ) 後 、 ピ ペ ッ ト マ ン を 用 い て 上 清 を 捨 て 、 残 存 す る エ タ ノ ー ル を 除 去 後 、 チ ュ ー ブ の フ タ を 空 け て 室 温 で 風 乾 し た 。 N u c l e a s e - F r e e Wa t e r ( Q i a g e n , 1 2 9 11 5 ) で R N A を 溶 解 し 、 R e c o m b i n a n t D N a s e I ( Ta k a r a - B i o , 2 2 7 0 A ) に よ る ゲ ノ ム D N A の 消 化 を 3 7 ℃ , 3 0 分 間 行 っ た 。 3 M 酢 酸 ナ ト リ ウ ム 、 エ タ 沈 メ イ ト ( Wa k o P u r e C h e m i c a l , 3 1 2 - 0 1 7 9 1 ) を そ れ ぞ れ 加 え て 、 エ タ ノ ー ル 沈 殿 を 行 い 、 精 製 R NA サ ン プ ル と し た 。 R NA の 純 度 、 濃 度 測 定 は 微 量 分 光 光 度 計 ( N a n o D r o p 2 0 0 0 / T h e r mo S c i e n t i f i c J a p a n ) を 用 い て 行 っ た 。
cDN A合 成 (定 量 RT-PCR用 )
定 量 RT-P CR に 用 い る c D NA は 、 P r i m e S c r i p t RT R e a g e n t k i t ( Ta k a r a -B i o , R R 0 3 7 ) を 使 用 し 、 t o t a l R N A 1 gか ら mRNAの 特 異 的 な 逆 転 写 反 応 (37℃ 、 15分 間 )に よ っ て 取 得 し た 。 逆 転 写 酵 素 の 失 活 は 8 5 ℃ で 5 秒 間 の 熱 処 理 に よ り 行 っ た 。定 量 RT-PCR
遺 伝 子 転 写 量 の 解 析 は Li g h t C yc l e r S ys t e m ( R o c h e D i a g n o s t i c s ) を 用 い て 行 っ た 。 こ の シ ス テ ム は P C R 産 物 の 増 幅 を 蛍 光 シ グ ナ ル で モ ニ タ リ ン グ す る こ と に よ り 、 定 量 的 か つ リ ア ル タ イ ム に 検 出 す る こ と が 可 能 で あ る 。 本 研 究 に お け る 遺 伝 子発 現 量 の 解 析 で は 、 U n i ve r s a l P r o b e Li b r a r y ( R o c h e D i a g n o s t i c s ) 由 来 の Ta q M a n プ ロ ー ブ と 称 さ れ る 加 水 分 解 プ ロ ー ブ を 使 用 し た 。 こ の プ ロ ー ブ は 、5’末 側 に 蛍 光 レ ポ ー タ ー 色 素 、3’末 側 に ク エ ン チ ャ ー が そ れ ぞ れ ラ ベ ル さ れ て い る ( Fig. 5)。 5’末 側 の 蛍 光 レ ポ ー タ ー 色 素 に よ る 蛍 光 シ グ ナ ル は 、 完 全 状 態 で は 3’末 側 の ク エ ン チ ャ ー に よ り ほ ぼ 完 全 に 抑 制 さ れ て お り 、蛍 光 シ グ ナ ル は 皆 無 に 等 し い 。 し か し 、Ta q D NA ポ リ メ ラ ー ゼ の 5 ’ → 3 ’ エ キ ソ ヌ ク レ ア ー ゼ 活 性 に よ り 分 解 さ れ る と 、蛍 光 色 素 が ク エ ン チ ン グ さ れ な く な る 。こ れ に よ り 、P C R サ イ ク ル 中 に 、 5 ’ 末 側 か ら 切 断 さ れ た 蛍 光 色 素 が 蓄 積 さ れ 、 蛍 光 シ グ ナ ル が 増 大 す る ( F i g . 6 ) 。 蛍 光 強 度 が 検 知 さ れ た P C R の 蛍 光 増 殖 曲 線 の 立 ち 上 が り 点 ( C yc l e 数 ) が C r o s s i n g F i g . 5 . T a q M a n p r o b e . F i g . 6 . Ta q M a n プ ロ ー ブ を 用 い た リ ア ル タ イ ム 定 量 P C R 法 の 概 略 .
P o i n t ( C P ) と し て 示 さ れ る 。 C P 値 は 、 鋳 型 と な る D N A の 初 期 濃 度 に 依 存 し て い る た め 、 CP 値 の 差 異 が 遺 伝 子 転 写 量 の 差 と し て 求 め る こ と が で き る ( Fi g . 7 ) 。
定 量RT-PCR 反 応 に は 、 Ligh tC yc ler 480 ( Roche Diagnostic s) を 使 用 し た 。 PCR反
応 液 に は 、 Li g h t C yc l e r P r o b e s M a s t e r ( R o c h e D i a g n o s t i c s ) を 使 用 し 、 1 反 応 あ た り 、 以 下 の 組 成 で 行 っ た ; 1 × LC 4 8 0 p r o b e s M a s t e r M i x , 0 . 5M Prime r L, 0.5 M
P r i m e r R , 0 . 1M TaqMan probe, cDNA 1 l。リ ア ル タ イ ム 定 量 PCRの 反 応 条 件 は 、
P r e - i n c u b a t i o n ( × 1 c y c l e ) : 9 5 ℃ , 1 0 分 間 ; A m p l i f i c a t i o n ( × 4 5 c y c l e s ) : 9 5 ℃ , 1 0 秒 間 - 6 0 ℃ , 3 0 秒 間 - 7 2 ℃ , 1 秒 間 で 行 っ た 。 ま た 、 Ta q M a n プ ロ ー ブ の 選 択 及 び プ ラ イ マ ー の 設 計 は 、 R o c h e A s s a y D e s i g n C e n t e r ( h t t p : / / w w w. u n i v e r s a l p r o b e l i b r a r y. c o m ) を 利 用 し た 。 本 章 に お い て 使 用 し た プ ラ イ マ ー 及 び Ta q M a n プ ロ ー ブ の リ ス ト を Ta b l e 2 に 示 し た 。な お 、内 部 標 準 遺 伝 子 と し て a c t i n 遺 伝 子 を 使 用 し た 。 F i g . 7 . P C R a m p l i f i c a t i o n c u r v e i n r e a l - t i m e P C R .
マ イ ク ロ ア レ イ
マ イ ク ロ ア レ イ 解 析 に 使 用 す る ア カ パ ン カ ビ の mR N A の 抽 出 に は 、 mR N A D i r e c t K i t ( I n v i t r o g e n ) を 使 用 し 、 プ ロ ト コ ル を 一 部 改 変 し て 行 っ た 。 ま ず 、 2 m l
ス ク リ ュ ー チ ュ ー ブ に 0 . 5 m m径 ガ ラ ス ビ ー ズ 2 0 0 l, lysis/bin ding buffer (100
m M Tr i s - H C l , 5 0 0 m M L i C l , 1 0 m M E D TA , p H 8 . 0 , 1 % L i D S , 5 m M D T T ) 1 m l を 用 意 し 、菌 体 サ ン プ ル ( 分 生 子 o r 菌 糸 体 ) を 加 え た 。ビ ー ズ ビ ー タ ー ( M P B i o me d i c a l s , J a p a n , F a s t P r e pT M F P 1 2 0 / B I O 1 0 1 ; 設 定 条 件 - S p e e d : 6 . 0 m / s , Ti m e : 4 0 秒 ) で 菌 体 を
破 砕 後 、 遠 心 分 離(13,000g, 4℃ , 5分 間 )し た 。 新 し い マ イ ク ロ チ ュ ー ブ に
D y n a b e a d s 2 0 0 lを 用 意 し 、同 チ ュ ー ブ を マ グ ネ ッ ト に セ ッ ト 後 、約 30秒 経 過 後 、
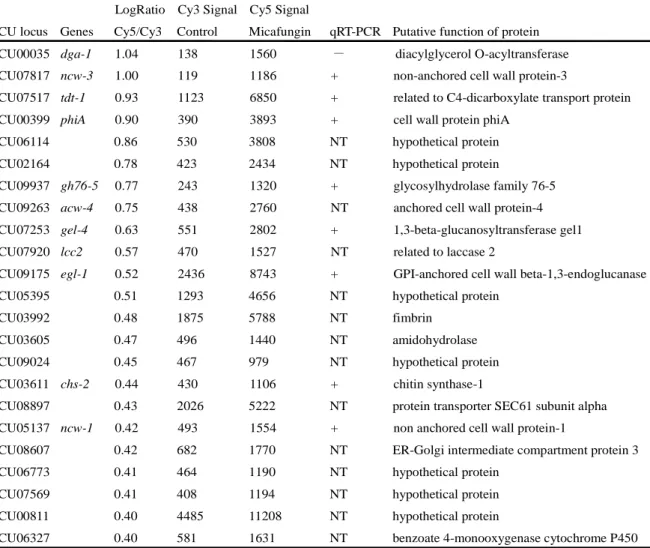
Table 2. List of primer sets and probes used for quantitative real-time RT-PCR in this study.
Target gene NCU locus Forward primer (5′→3′) Reverse primer (5′→3′) Probe #
ags-1 NCU02478 GGGTTGAGATGTGGAACCAG GGCCAGTATTAGGGTTGACG 9
ags-2 NCU08132 CCGTCCTCTCCAGGTGTCT CGACGCCCTGGATGTTAT 5
fks-1 NCU06871 TACGGCATGGACTACGGTAA GGGATACGGTTCCTTGGAG 4
gel-1 NCU01162 TCGGGAGGTCTTATGTACGAG GAGCTTGGCAATTCCGTATC 25
gel-2 NCU06781 CTCCGTCCCCGTCTTCTACT GTGAACATACGGGGCTTCA 46
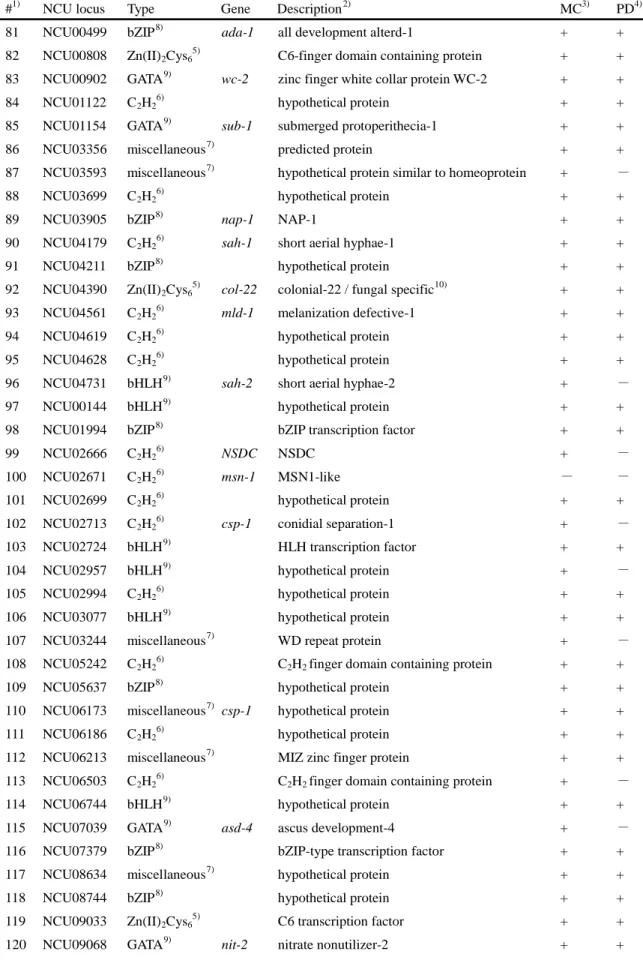
gel-4 NCU07253 AGAGATTCTACGTTCGCGGTA GATCGAGGTTGGCAGAAGAA 64
gfa-1 NCU11350 TCGACTGCCTTCAGGGTATC CCAACCAATAGGCGATCAAC 6
chs-1 NCU05239 AGTATTTCAAGGGCGAGACG ACATGTTCGCCGTAAACACA 44
chs-2 NCU03611 TCAGGATCAGCAGGTGTCAT GCCAAGAAGTAAAGGCCGTA 49
chs-3 NCU04251 CACCAGCAAGGCTTCGATA TAGCGCATCTCCAGGACTGT 81
chs-4 NCU09324 TCGATTGCAATGACGGACTA ACCATCGCAAATAACGAGGA 5
chs-5 NCU04352 TCGACGAGGTCAGCGAGT CTTGTTCCGATCCCTTGCT 3
chs-7 NCU04350 GTGGTTTGGGATTCCAAGAAG GAACCACGGGTGATTCCA 4
mak-1 NCU09842 CATTGTCTGGTACGTCCACCT CAAAAGCACAAGCGGCTAA 15
mak-2 NCU02393 GCATCACGGTCGAAGAGG ATCATCCGGGTCGTGGTA 1
phiA NCU00399 CTCGCAGCACCTCACCTT ATGCGGTACGAGGAGTCG 43
ncw-1 NCU05137 CCTGGCTCAACCAGATCG GGATTAGACCGAGGGGAGAA 25
egl-1 NCU09175 GGCTTCAACTCGGGTAACAC GCACCCAGTCCTTCTTGAAC 28
ncw-3 NCU07817 GGTGGTGGCTTCGTCAAT CTGAGGAGGGCAAGCAAC 45
gh76-5 NCU09937 ATTTGCAACACGGACATGC ATCTTGGAGGCGTAGACGAA 7
tdt-1 NCU07517 TTGTGTTGGGCAACCATGT CCCTCCCCGGATAACAAATA 77
dga-1 NCU00035 CTTTTCTCGGCATGTTCCTT TCTTTTCCAAAGGCTTGGTG 34
溶 液 が ク リ ア に な っ た と こ ろ で 上 清 を 除 去 し た 。 同 チ ュ ー ブ を マ グ ネ ッ ト か ら 外 し て 、 l ys i s / b i n d i n g b u ff e r で D yn a b e a d s を 洗 浄 し た 。 前 述 の 破 砕 菌 体 サ ン プ ル
の 上 清 7 5 0lを wash 済 Dynab eads入 り マ イ ク ロ チ ュ ー ブ に 加 え 、 転 倒 混 和 後 、
R o l l e r M i x e r を 使 用 し て 、 室 温 、 5 分 イ ン キ ュ ベ ー ト し た 。 磁 気 分 離 後 、 上 清 を 捨 て 、 w a s h i n g b u ff e r A ( 1 0 mM Tr i s -H C l , 1 5 0 mM Li C l , 1 m M E D TA , p H 8 . 0 , 0 . 1 % L i D S ) 及 び w a s i n g b u f f e r B ( 1 0 m M Tr i s - H C l , 1 5 0 m M L i C l , 1 m M E D TA , p H 8 . 0 ) で B e a d s / m R N A c o m p l e x を 室 温 で そ れ ぞ れ 2 回 ず つ 洗 浄 し た 。 洗 浄 後 の B e a d s か ら m R N A を 遊 離 ・ 溶 解 す る た め 、 e l u t i o n b u ff e r ( 1 0 m M Tr i s - H C l , p H 7 . 5 ) 2 0lを 加 え て 8 0 ℃ 、2 分 イ ン キ ュ ベ ー ト し 、磁 気 分 離 後 の 上 清 を 新 し い マ イ ク ロ チ ュ ー ブ に 移 し 、 mR N A サ ン プ ル と し た 。 解 析 に 使 用 し た マ イ ク ロ ア レ イ ス ラ イ ド は 、Fu n g a l G e n e t i c s S t o c k C e n t e r か ら 購 入 し た 。購 入 し た ア レ イ ス ラ イ ド の G ALフ ァ イ ル 及 び 遺 伝 子 リ ス ト の 情 報 は 、 h t t p : / / w e b . u c o n n . e d u / t o w n s e n d / L i n k s / f f d a t a b a s e / d o w n l o a d s . h t m l か ら 取 得 し た 。 ア レ イ ス ラ イ ド は 、 U V ク ロ ス リ ン ク ( 6 0 0 mJ ) に よ る D N A プ ロ ー ブ の 固 定 を 行 っ て か ら 使 用 し た 。 マ イ ク ロ ア レ イ 解 析 に 用 い た mR N A は 、 野 生 株 を 1 g/ml m i c a f u n g i n で 2 時 間 処 理 し た 菌 体 か ら 抽 出 し 、 調 製 し た m R N A 1 gを マ イ ク ロ ア レ イ 解 析 に 用 い た 。 マ イ ク ロ ア レ イ 解 析 は 、 3 D NA A r r a y 5 0 Ki t ( G e n i s p h e r e , H a t f i e l d , P A ) の プ ロ ト コ ル を 一 部 改 変 し て 行 っ た 。 c D NA 合 成 時 に 、 C y3 蛍 光 色 素 及 び C y5 蛍 光 色 素 が そ れ ぞ れ 特 異 的 に 結 合 す る プ ラ イ マ ー と 、 逆 転 写 酵 素 と し て S u p e r S c r i p t I I ( I n v i t r o g e n ) を 使 用 し た 。 野 生 株 無 処 理 の m R N A サ ン プ ル を C y 3 、 野 生 株 m i c a f u n g i n 処 理 の m R N A サ ン プ ル を C y 5 標 識 し た 。 c D N A 濃 縮 に は 、 M i c r o c o n Y M - 3 0 ( M i l l i p o r e ) を 用 い た 。 ハ イ ブ リ ダ イ ゼ ー シ ョ ン の バ ッ フ ァ ー に は 、 2 × S D S - B a s e d H y b r i d i z a t i o n B u ff e r を 使 用 し 、 c D N A ハ イ ブ リ ダ イ ゼ ー シ ョ ン を 6 0 ℃ で オ ー バ ー ナ イ ト 、 3 D NA ハ イ ブ リ ダ イ ゼ ー シ ョ ン を 5 5 ℃ で 3 時 間 行 っ た 。 各 ハ
イ ブ リ ラ イ ゼ ー シ ョ ン 後 の ア レ イ ス ラ イ ド の wa s h に は 、 予 め 4 2 ℃ に 加 温 し た 2 × S S C , 0 . 2 % S D S w a s h b u ff e r を 用 い た 。 な お 、 こ こ に 記 載 し た 実 験 方 法 は 、 マ イ ク ロ ア レ イ と 定 量 P CR の 発 現 差 が 、最 も 高 い 相 関 ( R2 = 0 . 9 9 ) を 示 す こ と が 山 下 和 宏 博 士 ( 2 0 1 0 年 度 東 洋 大 学 大 学 院 生 命 科 学 研 究 科 生 命 科 学 専 攻 博 士 学 位 論 文 ) に よ り 示 さ れ て い る 。 蛍 光 シ グ ナ ル は D u a l - La s e r D N A M i c r o a r r a y S c a n n e r ( A g i l e n t T e c h n o l o g i e s , S a n t a C l a r a , C A , G 2 5 6 5 A A ) で 検 出 し た 。 次 に 、 F e a t u r e E x t r a c t i o n S o f t w a r e v e r s i o n 8 . 5 ( A g i l e n t T e c h n o l o g i e s , S a n t a C l a r a , C A ) を使 用 し て 、 ア レ イ ス ポ ッ ト の シ グ ナ ル 強 度 と バ ッ ク グ ラ ウ ン ド に 有 意 に 差 が あ る か を 判 定 し 、 ノ イ ズ レ ベ ル の ス ポ ッ ト を 除 い て 数 値 化 し た 。
ア カ パ ン カ ビ の ゲ ノ ム DNA抽 出 及 び 調 製
2 m l ス ク リ ュ ー チ ュ ー ブ に 2 0 0 lの ガ ラ ス ビ ー ズ (0.5 mm径 )を 加 え 、抽 出 バ ッ フ ァ ー ( 5 0 mM Tr i s - H C l ( p H 8 . 0 ) , 1 0 0 mM E D TA , pH 8 . 0 , 0 . 5 % SD S ) 5 0 0 μ l を 加 え た 。 菌 体 サ ン プ ル(分 生 子 や 菌 糸 )を 滅 菌 済 み 竹 串 で 採 取 し 、 抽 出 バ ッ フ ァ ー 中 に 懸 濁 し た 。 細 胞 を ビ ー ズ ビ ー タ ー で 破 砕 し た ( M P B i o me d i c a l s , J a p a n , F a s t P r e pT M F P 1 2 0 / B I O 1 0 1 ; 設 定 条 件 - S p e e d : 6 . 0 m / s , Ti m e : 4 0 秒 ) 。 菌 体 由 来 の D N a s e を 失 活 さ せ る た め 、 直 ち に ウ ォ ー タ ー バ ス に 移 し て 6 5 ℃ 、 7 分 間 イ ン キ ュ ベ ー ト し 、 7 . 5 M 酢 酸 ア ン モ ニ ウ ム を 3 0 0 l加 え て 、 十 分 に 転 倒 混 和 後 、 氷 上 で 1 0 分 間 以 上 静 置 し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 5 分 間 、 4 ℃ ) 後 の 上 清 を 新 し い マ イ ク ロ チ ュ ー ブ に 移 し 、 等 量 の P C I ( p h e n o l : c h l o r o f o r m : i s o a m yl a l c h o l = 2 5 : 2 4 : 1 ) を 加 え て 、 ボ ル テ ッ ク ス を 用 い て 混 合 し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 5 分 間 、 4 ℃ ) 後 、 水 層 ( 上 層 ) の み を 新 し い チ ュ ー ブ に 移 し 、 9 9 . 5 % エ タ ノ ー ル を 上 清 の 2 . 5 倍 量 添 加 し 、 軽 度 に 転 倒 攪 拌 し た 。 再 度 、 同 条 件 で の 遠 心 分 離 後 、 上 清 を 捨 て 、 7 0 % エ タ ノ ー ル で リ ン ス し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 2 分 間 、 4 ℃ ) し た 後 、 上 清 を 除 去 し 、 M i c r o Va c ( T O M Y ) を 用 い て 真 空 乾 燥 さ せ た 。 T E b u ff e r ( p H 8 . 0 ) 5 0 lに 溶 解 後 、R N a s e 処 理 を 行 い ( 1 m g / m l R N a s e , 1 l)、 精 製 ゲ ノ ム DNAサ ン プ ル と し た 。
大 腸 菌 か ら の プ ラ ス ミ ド 調 製
基 本 的 な 操 作 、手 順 は ア ル カ リ-SDS法 に よ る 方 法 に 準 じ て プ ラ ス ミ ド の 調 製 を 行 っ た 。LB 培 地 で 8時 間 ~ 一 晩 培 養 し た 大 腸 菌 の 培 養 液 を 10,000g, 1分 間 、遠 心 分 離 し て 大 腸 菌 を 沈 殿 さ せ 、 培 地 ( 上 清 ) を 除 去 後 、 S o l u t i o n I ( 5 0 mM グ ル コ ー ス , 2 5 mM Tr i s -H C l ( p H 8 . 0 ) , 1 0 mM E D TA ( p H 8 . 0 ) ) を 添 加 し て 細 胞 を ボ ル テ ッ ク ス を 用 い て 懸 濁 し た 。 S o l u t i o n I I ( 0 . 2 M N a O H , 1 % SD S ) を 加 え て 3 回 転 倒 混 和 後 、 4 分 間 氷 冷 後 、 S o l u t i o n I I I ( 3 M 酢 酸 カ リ ウ ム , 5 M 酢 酸 ) を 添 加 し て 3 回 転 倒 混 和 後 、 5 分 間 氷 冷 し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 5 分 間 、 4 ℃ ) 後 の 上 清 を マ イ ク ロ チ ュ ー ブ に 移 し 、 等 量 の P C I ( P h e n o l : C h l o r o f o r m : I s o a m y l a l c o h o l = 2 5 : 2 4 : 1 ) を 加 え て 、ボ ル テ ッ ク ス を 用 い て 混 合 し た 。遠 心 分 離 ( 1 3 , 0 0 0 g 、5 分 間 、4 ℃ ) 後 、 水 層 ( 上 層 ) の み を 新 し い チ ュ ー ブ に 移 し 、 9 9 . 5 % エ タ ノ ー ル を 上 清 の 2 . 5 倍 量 添 加 し 、 軽 度 に 転 倒 攪 拌 し た 。 再 度 、 同 条 件 で の 遠 心 分 離 後 、 上 清 を 捨 て 、 7 0 % エ タ ノ ー ル で リ ン ス し た 。 遠 心 分 離 ( 1 3 , 0 0 0 g 、 2 分 間 、 4 ℃ ) し た 後 、 上 清 を 可 能 な 限 り 除 去 し 、M i c r o Va c ( TO M Y ) を 用 い て 真 空 乾 燥 さ せ た 。T E b u ff e r ( p H 8 . 0 ) 5 0 lに 溶 解 し た 後 、 RNase処 理 を 行 い (1 mg /ml RNase, 1 l)、 精 製 プ ラ ス ミ ド サ ン プ ル と し た 。pMAK-1-sGFP の 構 築
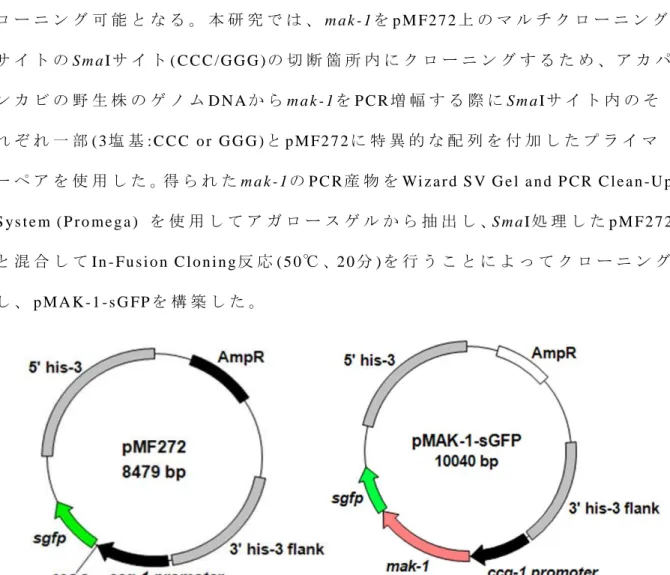
p M A K - 1 - s G F P ( F i g . 8 ) の 構 築 に 使 用 し た プ ラ イ マ ー を Ta b l e 3 に 記 し た 。ま た 、 ア カ パ ン カ ビ 用 発 現 ベ ク タ ー は FG S C か ら 入 手 し た p M F2 7 2 ( FG S C # 6 0 9 , G e n B a n k : AY 5 9 8 4 2 8 , F i g . 8 , F r e i t a g e t a l . , 2 0 0 4 ) を 使 用 し た 。 p M F 2 7 2 に は 、 c c g - 1 p r o m o t e r が 含 ま れ て お り 、 構 成 的 に 下 流 の 遺 伝 子 を 発 現 さ せ る こ と が 出 来 る 。 ク ロ ー ニ ン グ 反 応 に お い て は 、 In - Fu s i o n C l o n i n g e n z ym e ( Ta k a r a / C l o n t e c h ) を 使用 し た 。こ の 酵 素 は 3 ’ → 5 ’ e n d o n u c l e a s e 活 性 を 有 し て お り 、そ れ ぞ れ 異 な る D NA 断 片 の 3 ’ 末 端 か ら 1 5 塩 基 以 下 を 分 解 す る 。 そ の 結 果 生 じ た 相 同 な 突 出 し た 5 ’ 末 端 間 が 核 酸 間 の 相 互 作 用 に よ っ て ハ イ ブ リ ダ イ ゼ ー シ ョ ン す る こ と に よ り 、 ク ロ ー ニ ン グ 可 能 と な る 。 本 研 究 で は 、 m ak - 1 を p M F2 7 2 上 の マ ル チ ク ロ ー ニ ン グ サ イ ト の S m a Iサ イ ト ( C C C / GG G ) の 切 断 箇 所 内 に ク ロ ー ニ ン グ す る た め 、 ア カ パ ン カ ビ の 野 生 株 の ゲ ノ ム D NA か ら m a k - 1を P C R 増 幅 す る 際 に Sm a Iサ イ ト 内 の そ れ ぞ れ 一 部 ( 3 塩 基 : C C C o r G G G ) と p M F2 7 2 に 特 異 的 な 配 列 を 付 加 し た プ ラ イ マ ー ペ ア を 使 用 し た 。得 ら れ た m a k - 1 の P C R 産 物 を Wi z a r d S V G e l a n d P CR C l e a n -U p S y s t e m ( P r o m e g a ) を 使 用 し て ア ガ ロ ー ス ゲ ル か ら 抽 出 し 、S m a I 処 理 し た p M F 2 7 2 と 混 合 し て In - Fu s i o n C l o n i n g 反 応 ( 5 0 ℃ 、2 0 分 ) を 行 う こ と に よ っ て ク ロ ー ニ ン グ し 、 p M A K -1 - s G FP を 構 築 し た 。
Table 3. Primers for pMAK-1-sGFP construction Primer name Primer sequence (5′→3′)
mak-1-gfp F2 aactagtggatccccATGGCTGATCTCGTGGGTCG mak-1-gfp R2 catgttaattaacccCATGCCACCAAGCTCGGCCT pMF272 gfp F1 acgtaaacggccacaagttc
pMF272 gfp R1 ttctcgttggggtctttgct
mak-1 sequence from wild-type genome DNA were amplified with the 2 primers, mak-1-gfp F2 and mak-1-gfp R2, using PCR.
The underlined sequences are 15 bp overlaps required for recombination (pMF272 specific sequence and partial SmaI-site) that were added to allow for the directional insertion of the PCR products into the pMF272 vectors using In-Fusion HD Cloning reaction. pMF272 gfp F1 and pMF272 gfp R1 were used to confirm N. crassa transformant.
F i g . 8 . M a p o f p M F 2 7 2 a n d p M A K - 1 - s G F P
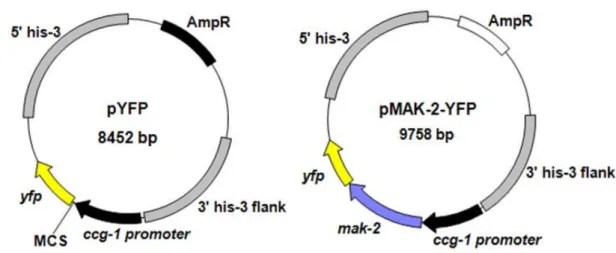
pMAK-2-YFPの 構 築
p M A K - 2 - Y F P ( F i g . 9 ) の 構 築 に 使 用 し た プ ラ イ マ ー を Ta b l e 4 に 記 し た 。 ま た 、 発 現 ベ ク タ ー は FG S C か ら 入 手 し た p Y FP ( FG S C # 6 7 5 , G e nB a n k : E F6 6 1 0 3 0 , Fi g . 9 ) を 使 用 し た 。 p Y FP も p M F2 7 2 と 同 様 に 、 c c g - 1 p r o mo t e r が 含 ま れ て い る 。 m a k - 2 を p Y FP 上 の マ ル チ ク ロ ー ニ ン グ サ イ ト の S m a Iサ イ ト ( C C C / G GG ) の 切 断 箇 所 内 に ク ロ ー ニ ン グ す る た め 、ア カ パ ン カ ビ の 野 生 株 の ゲ ノ ム D NA か ら m a k - 2 を P CR 増 幅 す る 際 に S m a Iサ イ ト 内 の そ れ ぞ れ 一 部 ( 3 塩 基 : C C C o r G GG ) と p Y FP に 特 異 的 な 配 列 を 付 加 し た プ ラ イ マ ー ペ ア を 使 用 し た 。 得 ら れ た m ak - 2 の P C R 産 物 を Wi z a r d S V G e l a n d P C R C l e a n - U p S y s t e m ( P r o m e g a ) を 使 用 し て ア ガ ロ ー ス ゲ ル か ら 抽 出 後 、 S m a I処 理 し た p Y FP と 混 合 し 、 In - Fu s i o n C l o n i n g 反 応 ( 5 0 ℃ 、 2 0 分 ) に よ り ク ロ ー ニ ン グ し た 。Table 4. Primers for pMAK-2-YFP construction Primer name Primer sequence (5′→3′)
mak-2-yfp F2 aactagtggatccccATGAGCAGCGCACAA mak-2-yfp R2 catgttaattaacccCCTCATAATCTCCTG pYFP F1 acgtaaacggccacaagttc
pYFP R1 ttctcgttggggtctttgct
mak-2 sequence from wild-type genome DNA were amplified with the 2 primers, mak-2-yfp F2 and mak-2-yfp R2, using PCR.
The underlined sequences are 15 bp overlaps required for recombination (pYFP specific sequence and partial SmaI-site) that were added to allow for the directional insertion of the PCR products into the pYFP vectors using In-Fusion HD Cloning reaction.
pYFP F1 and pYFP R1 were used to confirm N. crassa transformant.
F i g . 9 . M a p o f p Y F P a n d p M A K - 2 - Y F P
ア カ パ ン カ ビ の 形 質 転 換 法
形 質 転 換 に お い て は G e n e P u l s e r I I E l e c t r o p o r a t i o n S ys t e m ( B IO - R AD L a b o r a t o r i e s , H e r c u l e s , C A ) を 用 い た エ レ ク ト ロ ポ レ ー シ ョ ン 法 に よ り 、 遺 伝 子 断 片 を 導 入 し た 。 ま ず 、 ア カ パ ン カ ビ の hi s - 3 株 ( FG S C # 6 1 0 3 ) を 0 . 2 mg / m l ヒ ス チ ジ ン 含 有 グ リ セ ロ ー ル 完 全 培 地 で 室 温 条 件 下 、 1 0 日 間 培 養 し 、 大 量 の 分 生 子 を 回 収 し た 。 1 M ソ ル ビ ト ー ル で 3 回 洗 浄 後 、 1 M ソ ル ビ ト ー ル 5 0 0 l に 懸 濁 し 、 マ イ ク ロ チ ュ ー ブ に 移 し 、 遠 心 分 離 ( 1 0 , 0 0 0 g , 4 ℃ , 1 分 間 ) し た 。 沈 殿 し た 分 生 子 と 上 清 の 比 が 2 : 1 と な る よ う に 上 清 を 捨 て 、再 度 、十 分 に ボ ル テ ッ ク ス で 混 合 し 、 濃 度 の 高 い 分 生 子 懸 濁 液 を 準 備 し た 。そ の 胞 子 液 か ら 4 0 lの 胞 子 液 を 新 し い マ イ ク ロ チ ュ ー ブ に 採 取 し て 、遺 伝 子 導 入 断 片 ( D r a I処 理 に よ り 線 状 化 し た プ ラ ス ミ ド ) を 3 0 0 n g ( 添 加 量 : 3 ~ 5 l)加 え て 混 合 し た 後 、 5分 間 氷 冷 し た 。 Gene P ulserの エ レ ク ト ロ ポ レ ー シ ョ ン 条 件 を C APA C ITA NC E : 5 0 F, RESOSTANCE: 200
o h m n s , S E T V O LT S : 7 . 5 k V / c m に 設 定 し 、 胞 子 液 4 0 lと 遺 伝 子 導 入 断 片 の 混 合 物 を エ レ ク ト ロ ポ レ ー シ ョ ン 専 用 キ ュ ベ ッ ト に 全 量 加 え て 、 エ レ ク ト ロ ポ レ ー シ
ョ ン を 行 っ た 。氷 冷 1 M ソ ル ビ ト ー ル 9 6 0 lを 同 キ ュ ベ ッ ト に 加 え て 穏 や か に ピ
ペ ッ テ ィ ン グ で 混 合 し て 1 M ソ ル ビ ト ー ル が 入 っ て い た マ イ ク ロ チ ュ ー ブ に 全
量 を 回 収 し て 1 0 分 間 氷 冷 し た 。 こ の う ち 、 2 5 0 l~ 1 mlを Top agar (2% So rbose,
1 M S o r b i t o l , 1 × Vo g e l ’s S o l u t i o n , 1 . 5 w / v % A g a r ) に 混 ぜ て 、 ソ ル ボ ー ス 含 有 Vo g e l ’s 寒 天 培 地 上 に 展 開 し 、 3 0 ℃ で 3 日 間 培 養 し た 。 パ ス ツ ー ル ピ ペ ッ ト を 使 用 し て 、 生 育 し て き た コ ロ ニ ー を V M 寒 天 培 地 に ピ ッ ク ア ッ プ し 、 1 週 間 程 度 培 養 し た 。 本 章 で 使 用 し た p M F2 7 2 , p Y FP は ア カ パ ン カ ビ の h i s - 3 座 に 2 箇 所 の 相 同 組 み 換 え に よ り 組 み 込 ま れ る よ う 、 h i s - 3 座 が 一 部 改 変 さ れ た ゲ ノ ム 配 列 が 使 用 さ れ て い る ( M a rg o l i n e t a l . , 1 9 9 7 ) 。 こ れ に よ り 、 ヒ ス チ ジ ン 要 求 株 の ヒ ス チ ジ ン 要 求 性 が 回 復 す る た め 、 ヒ ス チ ジ ン 無 添 加 の 培 地 中 か ら 形 質 転 換 体 を 選 抜 す る こ と が 可 能 で あ る 。 得 ら れ た 形 質 転 換 体 の 候 補 株 は 、 遺 伝 子 診 断 に よ り h i s - 3
遺 伝 子 座 へ の 導 入 断 片 の 挿 入 を 確 認 後 、 S yn t h e t i c c r o s s ( S C ) 培 地 ( 0 . 1 w / v% G l u c o s e , 1 × S C s o l u t i o n ) ( We s t e rg a a r d e t a l . , 1 9 4 7 ) 上 で 野 生 株 と 交 配 す る こ と に よ り ホ モ カ リ オ ン 化 し 、 目 的 の 形 質 転 換 体 を 取 得 し た 。
GFP or YFP融 合 タ ン パ ク 質 発 現 株 の 観 察 方 法
観 察 に は 共 焦 点 レ ー ザ ー ス キ ャ ン 顕 微 鏡 ( C a r l Ze i s s LS M - 5 1 0 ) を 使 用 し た 。 菌 糸 の 観 察 で は V M 固 体 培 地 ( 3 w/ v % 寒 天 ) 上 に G FP 融 合 タ ン パ ク 質 発 現 株 の 分 生 子 を 接 種 し 、 2 8 ℃ 、 1 4 ~ 1 8 時 間 、 前 培 養 し た 。 伸 長 し た 菌 糸 先 端 を ス パ ー テ ル や メ ス 刃 を 用 い て 培 地 ご と 切 り 取 り 、 滅 菌 水 を 1 0 l滴 下 し た ス ラ イ ド ガ ラ ス ( M AT S U N A M I N E O , 2 5 × 6 0 m m ) 上 に 、 菌 体 面 が 下 向 き に な る よ う に 乗 せ て キ ム ワ イ プ で 余 分 な 水 分 を 除 い た 。 G FP 蛍 光 の 検 出 は 、 励 起 光 及 び 蛍 光 フ ィ ル タ ー を そ れ ぞ れ E x c i t a t i o n 4 8 8 n m / E m i s s i o n 5 3 0 n m に 設 定 し て 観 察 し た 。 Y FP 蛍 光 の 検 出 は 、 励 起 光 及 び 蛍 光 フ ィ ル タ ー E x c i t a t i o n 5 1 4 n m / E mi s s i o n 5 2 7 n m に 設 定 し て 観 察 し た 。細 胞 核 染 色
細 胞 核 の 染 色 に は 、 D AP I ( 4 ' , 6 - d i a m i d i no - 2 - p h e n y l i n d o l e - H C l ) を 使 用 し た 。 上 述 と 同 様 に 、培 地 ご と 切 り 出 し た 菌 糸 を 固 定 液 ( 3 . 7 % パ ラ ホ ル ム ア ル デ ヒ ド , 0 . 1 M P B S ( p H 7 . 4 ) ) に 3 0 秒 間 浸 し 、 P B S で 1 5 秒 間 洗 浄 後 、 0 . 5 ~ 1 g/ml DAP Iで 2 分 間 、遮 光 条 件 下 で 染 色 し た 。新 た に 準 備 し た P B S で 2 回 洗 浄 し た 。D AP I蛍 光 の 検 出 は 、 共 焦 点 レ ー ザ ー ス キ ャ ン 顕 微 鏡 ( C a r l Ze i s s LS M - 5 1 0 ) を 使 用 し 、 励 起 光 及 び 蛍 光 フ ィ ル タ ー E x c i t a t i o n 3 6 4 n m / E m i s s i o n 4 5 4 n m に 設 定 し て 観 察 し た 。細 胞 壁 染 色
細 胞 壁 の 染 色 に は C a l c o f l u o r W h i t e ( C FW ) を 使 用 し た 。 上 述 と 同 様 に 、 伸 長 した 菌 糸 先 端 を ス パ ー テ ル や メ ス 刃 を 用 い て 培 地 ご と 切 り 取 り 、 ス ラ イ ド ガ ラ ス ( M AT S U N A M I N E O , 2 5 × 6 0 m m ) 上 に 0 . 1 ~ 1 . 0 m g / m l C F W を 1 0 l滴 下 し 、 そ こ に 菌 体 面 が 下 向 き に な る よ う ス ラ イ ド ガ ラ ス 上 に 乗 せ て キ ム ワ イ プ で 余 分 な 水 分 を 除 い た 。 C FW 蛍 光 の 検 出 は 、 共 焦 点 レ ー ザ ー ス キ ャ ン 顕 微 鏡 ( C a r l Ze i s s L S M - 5 1 0 ) を 使 用 し 、 励 起 光 及 び 蛍 光 フ ィ ル タ ー を そ れ ぞ れ E x c i t a t i o n 4 0 5 n m / E m i s s i o n 4 7 5 n m に 設 定 し て 観 察 し た 。
Polymerase chain reaction (PCR)
ベ ク タ ー 構 築 に お い て は 、 P h u s i o n® H i g h - F i d e l i t y D N A P o l y m e r a s e ( N E B ) を 使 用 し た 。 遺 伝 子 破 壊 株 の 遺 伝 子 破 壊 の 検 出 に お い て は 、 KAPA Ta q E X t r a D N A P o l y m e r a s e ( 日 本 ジ ェ ネ テ ィ ク ス ) を 使 用 し た 。 こ れ ら の D N A p o l y m e r a s e を 使 用 し た P C R 反 応 は 、各 付 属 の マ ニ ュ ア ル に 従 っ て 、P C R 反 応 液 を 調 製 し 、P C R 反 応 装 置 ( i C yc l e r / B IO -R A D La b o r a t o r i e s , H e r c u l e s , C A ) を 使 用 し た 。1-3. 結 果 と 考 察
1-3-1. MAPキ ナ ー ゼ 破 壊 株 の 細 胞 壁 ス ト レ ス 感 受 性
ア カ パ ン カ ビ は 3 種 類 の M AP キ ナ ー ゼ ( M AP K, M A K-1 , M A K- 2 , O S -2 ) を も っ て い る 。 ま ず 、 こ れ ら の 各 遺 伝 子 破 壊 株 (m a k - 1 , m a k - 2 ,o s - 2 ) の 示 す 形 質 を Vo g e l ’s 最 少 寒 天 培 地 で 生 育 さ せ て 野 生 株 と 比 較 し た 。 な お 、 菌 糸 形 態 の 異 常 と 生 育 遅 延 が 認 め ら れ た 破 壊 株 が 存 在 し た た め 、 菌 糸 形 態 は 各 菌 株 が 培 養 シ ャ ー レ に 十 分 に 生 育 し た 状 態 で 比 較 し 、 生 育 遅 延 は 野 生 株 の 2 4 時 間 培 養 後 の 菌 糸 生 育 長 に 対 す る 各 破 壊 株 の 菌 糸 生 育 比 率 で 示 し た ( Fi g . 1 0 ) 。 野 生 株 を Vo g e l ’s 最 少 寒 天 培 地 上 で 生 育 さ せ る と 、 接 種 源 か ら 円 形 状 に 菌 糸 が 生 育 し 、 培 養 シ ャ ー レ に 菌 糸 が い っ ぱ い に な る ま で ( 約 2 4 時 間 ) は 、 栄 養 成 長 を 続 け 、無 性 生 殖 に あ た る 分 生 子 形 成 は ほ ど ん ど 認 め ら れ な い 。こ れ に 対 し て 、 出 芽 酵 母 の C W I経 路 の M p k 1 の オ ル ソ ロ グ で あ る M A K-1 の 破 壊 株m a k - 1 は 、 F i g . 1 0 A に 示 し た よ う に 、 分 生 子 形 成 と 菌 糸 伸 長 が 高 頻 度 で 繰 り 返 さ れ 、 そ の 菌 叢 は 特 徴 的 な 形 態 を 示 し た 。 ま た 、 2 4 時 間 後 の 野 生 株 の 生 育 度 を 1 0 0 % と す る と 、 m a k - 1 の 生 育 度 は 2 0 % 以 下 で あ り 、3 種 類 の 破 壊 株 の 中 で 最 も 著 し い 生 育 遅 延 が F i g . 1 0 . C o m p a r i s o n o f m o r p h o l o g y o f w i l d - t y p e , m a k - 1 , m a k - 2 , a n do s - 2 .認 め ら れ た ( Fi g . 1 0 B ) 。 出 芽 酵 母 に お い て 交 配 能 を 制 御 す る こ と で 知 ら れ る F u s 3 / K s s 1 の オ ル ソ ロ グ M A K - 2 の 破 壊 株m a k - 2 は 、 培 養 開 始 直 後 ( 培 地 上 へ の 接 種 地 点 ) か ら 分 生 子 形 成 を 伴 い な が ら 菌 糸 を 伸 長 し 、菌 糸 生 育 は 、m a k - 1 よ り は 早 い 生 育 度 を 示 し た が 、野 生 株 の 4 0 % 程 度 の 生 育 遅 延 を 示 し た ( Fi g . 1 0 A , B ) 。最 後 に 、 出 芽 酵 母 の 浸 透 圧 応 答 経 路 の H o g 1 M AP キ ナ ー ゼ の オ ル ソ ロ グ で あ る O S - 2 の 破 壊 株o s - 2 は 、最 少 寒 天 培 地 上 で の 菌 糸 生 育 や 形 態 形 成 は 、野 生 株 と 顕 著 な 差 は 認 め ら れ な か っ た ( Fi g . 1 0 A , B ) 。 各 M A P キ ナ ー ゼ 破 壊 株 の 分 生 子 形 成 を 調 べ る た め に 、 分 生 子 形 成 培 地 ( グ リ セ ロ ー ル 完 全 培 地 )に 接 種 後 、1 0 日 間 培 養 後 に 形 成 さ れ た 分 生 子 数 を 比 較 し た 。 野 生 株 で は 、 培 養 あ た り 約 1 . 4 × 1 08個 / m l の 分 生 子 が 回 収 さ れ た が 、 o s - 2 及 び m a k - 2 で は 、 野 生 株 よ り も や や 分 生 子 形 成 が 抑 制 さ れ て い た も の の 、 1 . 0 × 1 08 個 / ml 以 上 の 分 生 子 が 回 収 さ れ た 。一 方 、m a k - 1 は 3 種 類 の 破 壊 株 の 中 で 最 も 顕 著 に 分 生 子 形 成 が 抑 制 さ れ 、 野 生 株 の 半 数 以 下 で あ っ た ( Fi g . 11 A ) 。 そ こ で 、 m a k - 1 の 分 生 子 を 顕 微 鏡 で 観 察 し た 。 野 生 株 、o s - 2 及 びm a k - 2 で は 、 菌 糸 か ら 直 接 形 成 さ れ る 小 型 分 生 子 ( mi c r o c o n i d i a ) と 分 生 子 柄 上 に 形 成 さ れ る 大 型 分 生 子 ( ma c r o c o n i d i a ) が 認 め ら れ た 。 一 方 、m a k - 1 は 正 常 な 2 種 類 の 分 生 子 に 加 え て 、 一 般 に は ほ と ん ど 見 ら れ な い 菌 糸 様 分 生 子 a r t h r o c o n i d i a が 数 多 く 形 成 さ れ F i g . 11 . C o n i d i a l m o r p h o l o g y i n m a k - 1 .
て い た ( Fi g . 11 B ) 。 こ れ ら の こ と か ら 、 M A K-1 は 菌 糸 生 育 や 分 生 子 形 成 に 必 須 で は な い も の の 、 3 種 類 の M A Kキ ナ ー ゼ の う ち 基 礎 生 育 に 最 も 影 響 の 大 き い M A P キ ナ ー ゼ で あ る と 考 え ら れ た 。 次 に 、 3 種 類 の M AP キ ナ ー ゼ が 細 胞 壁 損 傷 の 修 復 に ど の 程 度 関 与 し て い る か を 検 証 す る た め に 、 各 遺 伝 子 破 壊 株 の 細 胞 壁 損 傷 剤 に 対 す る 感 受 性 を 調 べ た 。 ま ず 、 酵 母 や A . n i d u l a n s の 研 究 で C W I経 路 の 活 性 化 を 促 す こ と が 知 ら れ て い る b e t a - 1 , 3 - グ ル カ ン 合 成 酵 素 阻 害 剤 に 着 目 し 、 本 研 究 で は 、 細 胞 壁 を 損 傷 さ せ る 薬 剤 と し て ア カ パ ン カ ビ に 高 い 抗 菌 活 性 を 示 し た m i c a f u n g i n を 採 用 し た 。 M i c a f u n g i n に 対 す る 感 受 性 の 評 価 は 1 / 1 0 に 段 階 的 に 希 釈 し た 分 生 子 懸 濁 液 を 、 薬 剤 含 有 培 地 に ス ポ ッ ト し て コ ロ ニ ー 形 成 の 有 無 を 観 察 し た ( Fi g . 1 2 ) 。 そ の 結 果 、 3 種 類 の 遺 伝 子 破 壊 株 の う ち 、o s - 2 は 、 野 生 株 と 同 等 の 感 受 性 を 示 し た 。 一 方 、m a k - 1 とm a k - 2 は 、 b e t a - 1 , 3 - グ ル カ ン 合 成 酵 素 阻 害 剤 m i c a f u n g i n に 対 し て 、 野 生 株 よ り も 高 い 感 受 性 を 示 す こ と が 明 ら か に な っ た 。 両 者 の 比 較 で は 、 当 初CWI経 路 に 最 も 関 与 し て い る と 予 測 し て い たm a k - 1 よ り も 有 性 生 殖 に 関 与 す るm a k - 2 の 方 が よ り 顕 著 な m i c a f u n g i n 感 受 性 を 示 し た ( F i g . 1 2 ) 。 M A P キ ナ ー ゼ は M A P K K キ ナ ー ゼ と M A P K キ ナ ー ゼ か ら な る M A P キ ナ ー ゼ カ ス ケ ー ド を 形 成 す る 。M A P キ ナ ー ゼ の 活 性 化 に は こ の 上 流 に あ た る M AP KKキ ナ F i g . 1 2 . S e n s i t i v i t y o f 3 M A P K d i s r u p t a n t s t o m i c a f u n g i n .