アルギニンとオルニチンの投与はラット心筋においてVEGF発現を促進し毛細血管新生を引き起こす
9
0
0
全文
(2) ? L % B R B % b R W ( $ % F + b l ) %6lf % 13 Journal of Hokkaido University of Education (Natural Sciences) Vol. 61, No.1. %E22F 8 8 August, 2010. L-Arginine and L-Ornithine Supplementation Facilitates Angiogenesis by Up-regulating VEGF Expression in Rat Cardiac Muscle SUZUKI Junichi Laboratory of Exercise Physiology, Health and Sports Sciences, Course of Sports Education, Department of Education, Iwamizawa Campus, Hokkaido University of Education. ABSTRACT The present study was designed to examine the effects of dietary L-arginine and Lornithine supplementation on capillary angiogenesis in young rats. Female (12-week-old) Wistar rats were used in the present study. Rats in the treated groups drank water containing 2.5% L-arginine and 1.2% L-ornithine (low-dose group) or 4% and 2% (high-dose group). According to histochemical identification of the capillary profile, high-dose supplementation significantly increased the capillary-to-fiber (C:F) ratio by 22%, while low-dose increased it non-significantly by 10%. Neither high- nor low-dose supplementation afFected capillary density and myofiber density. Although supplementation increased VEGF levels dosedependently, a significant increase was observed after high-dose treatment (P<0.05). Tissue eNOS expression slightly decreased after the treatment, but difference was not significant. The present results suggest that supplementation facilitates angiogenesis by promoting a NO/VEGF interrelationship in the cardiac muscle. [Keywords] Larginine, angiogenesis, endurance training, Lornithine, vascular endothelial growth factor. INTRODUCTION. Patterson & Runge 1999). Among these factors, vascular endothelial growth factor (VEGF) is. Angiogenesis, the development of new capillar-. t h o u g h t t o b e a n i m p o r t a n t r e g u l a t o r of. ies, is induced by a number of naturally occur-. angiogenesis during exercise training (Hoffner. ring growth factors, hormones, and cytokines. et al. 2003, Jensen et al. 2004, Kraus et al. 2004,. (Folkman & Klagsburn 1987, Breen et al. 1996,. W i t t w e r e t al. 2004 ), a s well a s d u r i n g.
(3) SUZUKI Junichi. embryonic development (Jakeman et al. 1993),. t a t e d angiogenesis by promoting VEGF ex-. wound healing (Brown et al. 1992), and tumor. pression.. growth (Celec et al. 2005). VEGF expression is increased in response to tissue hypoxia (Schweiki et al. 1992, Gold-. MATERIALS AND METHODS. berg & Schneider 1994). Recently, regulation of. This study was approved by the Animal Care. VEGF gene expression induced by NO has been. and Use Committee of Hokkaido University of. demonstrated in numerous cell types, including. Education and performed in accordance with. v a s c u l a r s m o o t h muscle cells (Dulak et al.. the "Guiding Principles for the Care and Use of. 2000). N (G)-nitro-L-arginine (LNNA) , inhibitor. Animals in the Field of Physiological Sciences". of NO s y n t h a s e (NOS), abolished exercise-. of the Physiological Society of Japan.. induced capillary angiogenesis in skeletal mus-. Animals, experimental conditions and cardiac samples. cle (Hudlicka et al. 2000). I n cardiac muscle,. Thirty- two (10-week-old) female Wistar rats. neonatal endothelial NOS-deficient NOS-'-). were purchased from Charles River Japan Inc.. mice had significantly less capillary density and. (Tokyo, Japan). After the rats were kept for 2. VEGF mRNA expression than neonatal wild-. weeks to allow adaptation to the new environ-. t y p e mice (Zhao et al. 2002). T h e s e findings. ment, they were randomly divided into the fol-. s t r o n g l y d e m o n s t r a t e t h a t endogenous NO. lowing three groups according to their treat-. plays a critical role in VEGF-mediated. ment conditions: non-treated (Cnt, n=8), low-. angiogenesis.. dose L-arginine and L-ornithine treated (Low,. Supplementation of L-arginine, a substrate. n=8), and high-dose t r e a t e d (High, n=8). T h e. for NOS, facilitated endothelial NO synthesis. amino acid administration started a t 12 weeks. during exercise and enhanced aerobic capacity. of a g e a n d l a s t e d for 6 weeks. All r a t s w e r e. (Maxwell et al. 2001). Although NO is a major. given commercial laboratory chow (CE-2, Clea. mediator of VEGF-mediated angiogenesis, diet-. Japan Inc.) ad libitum. Rats in the non-treated. ary L-arginine administration failed to induce. groups drank t a p water, whereas r a t s in the. angiogenesis in skeletal as well as cardiac musc-. treated groups drank water containing 2.5%. les (Suzuki 2005). While L-arginine is metabol-. (w/v) L-arginine (A5006, Sigma-Aldrich, St.. ized to NO, it is catalyzed to Lornithine by argi-. Louis, MO, USA) and 1.2% (w/v) L-ornithine. nase. Thus L-arginine administration alone can. (02375, Sigma-Aldrich) for the Low group and. not enhance NO production. I t seems possible. 4% and 2%, respectively, for the High group.. that administration of L-ornithine in addition to. Under light anesthesia with ether, the rats. L-arginine helps to metabolize L-arginine more. were anaesthetized with alpha-chloralose (0.06. favorably to NO, thereby facilitating VEGF-. g/Kg i. p.) and urethane (0.7 g/kg i. p.). A toe. induced angiogenesis.. pinch response was used to validate adequate. In the present study, experiments were de-. anesthesia. Then, the left ventricle (LV) were. signed to examine whether L-arginine and L-. excised and weighed. T h e tissues were placed. ornithine supplementation facilitates capillary. in e m b e d d i n g medium, a n Optimal C u t t i n g. angiogenesis in cardiac muscle. The present re-. T e m p e r a t u r e (O.C.T.) compound (Sakura. sults demonstrate that the administration facili-. Finetechnical, Tokyo, Japan), and frozen rapidly.
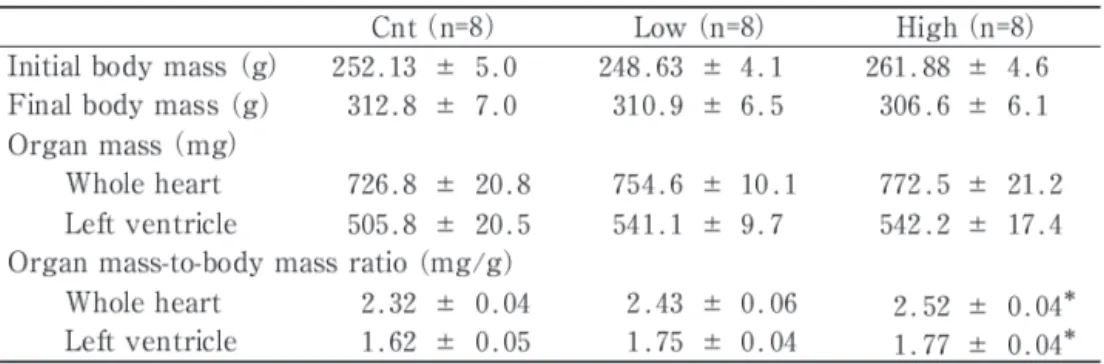
(4) LArginine and LOrnithine Supplementation Facilitates Cardiac Angiogenesis. in i s o p e n t a n e cooled t o i t s melting point. tissue homogenized with three interrupted 15-s. (-160°C) with liquid nitrogen. The apex half of. bursts with a Polytron homogenizer in ice-cold. the LV was used for biochemical analyses, while. medium (10 mM HEPES buffer, pH 7.3; 0.1%. the other half was used for histochemical analy-. Triton X-100 ; 11.5% (w/v) sucrose; and 5% (v. ses. All LV samples were treated in a similar. /v) protease inhibitor cocktail (P2714, Sigma) ) .. fashion. T h e t i s s u e s a m p l e s w e r e s t o r e d a t. After centrifugation a t 1,500 x g for 10 min a t. -80°C until analysis.. 4°C , t h e s u p e r n a t a n t w a s u s e d for protein Tissue cross-sections,. analysis. Total protein w a s m e a s u r e d using. 10-pm-thick, were obtained using a cryotome. PRO-MEASURE protein measurement solution. (CM-1500; Leica Japan, Tokyo, Japan) at -20°C.. (iNtRON Biotechnology, Korea). A sample was. To determine the capillary profiles, the sections. fractionated by SDS/PAGE on 7.5 or 12% (w. were double stained for alkaline phosphatase. /v) polyacrylamide gels, electrophoretically. (AP) and dipeptidyl peptidase IV (DPP IV) in. transferred to a polyvinylidene fluoride mem-. the capillary endothelium. Although the original. b r a n e , a n d blocked with 1% (w/v) bovine. s t a i n i n g protocol w a s d e s c r i b e d by Lojda. serum albumin for 1 h. The blots were exposed. (1979), a slightly modified protocol was used, as. t o specific p r i m a r y antibodies (Santa C r u z. described previously (Suzuki J 2002). T h e im-. Biotech, Santa Cruz, CA, USA) against VEGF. ages of incubated sections were digitized using. (1 :1500) and eNOS (1 :1000) diluted in blocking. a digital microscope camera (PDMC le, Polaroid,. solution. After the blots were incubated with an. Cambridge, USA) attached to a light micro-. AP-conjugated secondary antibody (Santa Cruz,. scope (BX-50, Olympus, Tokyo, J a p a n ) a n d. 1: 5000) diluted in blocking solution, t h e re-. were stored on computer disk. Non-overlapping. quired proteins were detected with a. microscopic fields were selected at random from. 5-bromo-4-chloro-3-indolyl. each cardiac sample. During the measurements,. blue tetrazolium reaction. The membranes were. t h e o b s e r v e r w a s blind a s t o t h e s o u r c e. scanned (GT-8200UF, Seiko Epson Co., Tokyo,. (groups) of each slide. Morphological analyses. J a p a n ) a n d t h e densities of t h e b a n d s w e r e. were obtained from the subendocardium por-. quantified using Image-J public domain software. tion of left ventricle.. (written by Rasband W, National Institutes of. Western blot analysis Tissue homogenates were. Health, USA). T o check for loading errors, the. obtained from approximately 50 mg of frozen. density of bands was normalized to the same. Histological analyses. phosphate/nitro. Table 1 Body and heart mass values.. Cnt (n=8) Low (n=8) High (n=8) Initial body mass (g) 252.13 k 5.0 248.63 k 4.1 261.88 k 4.6 Final body mass (g) 312.8 k 7.0 310.9 k 6.5 306.6 k 6.1 Organ mass (mg) Whole heart 726.8 k 20.8 754.6 k 10.1 772.5 k 21.2 505.8 k 20.5 541.1 k 9.7 542.2 k 17.4 Left ventricle Organ mass-to-body mass ratio (mg/g) Whole heart 2.32 * 0.04 2.43 * 0.06 2.52 k 0.04* Left ventricle 1.62 k 0.05 1.77 k 0.04* 1.75 k 0.04 Values a r e means k SE. *, significantly different from t h e C n t a t P<0.05..
(5) SUZUKI Junichi. Figure 1. Micrographic images of the left ventricle stained with AP and DPP IV. (a) Control group. (b) High-dose group. All images are at the same magnification. Horizontal bar represents 50pm.. * SE. Differences between t h e t h r e e. s a m p l e t h a t w a s r u n on e v e r y gel a n d elec-. means. trophoretically transferred to every membrane.. groups w e r e analyzed using nonparametric. Negative controls without primary or secondary. Shirley-Willams test. Differences were consi-. antibodies were run to validate the results.. dered statistically significant at P<0.05.. Biochemical analyses Using the muscle homogen-. ate aliquots, metabolic enzyme activities were measured. T h e activites of c i t r a t e s y n t h a s e. RESULTS. (CS) and S-hydroxyacy 1-CoA-dehydrogenase. Mean body mass a t the end of experiment was. (HAD) were assayed according to the method. not significantly different among t h e t h r e e. of S r e r e (1969) and Bass e t al. (1969), respec-. groups (Table 1). High-dose supplementation. tively. All measurements were carried out a t. significantly enhanced both the whole heart and. 25°C with a spectrophotometer (U-2001, Hitachi. left ventricle mass-to-body mass ratio values. Co., Tokyo, Japan).. (P <O. 05). Dietary L-arginine and L-ornithine. Statistical Analyses All values are expressed as. supplementation caused cardiac hypertrophy.. I. Cnt. Low. High. I. Figure 2 Capillary density and capillary-to-fiberratio and myofiber density in the slibendocardilim of left ventricle. All values are represented as means k SE. *, significantly different from the Cnt at P<0.05..
(6) LArginine and LOrnithine Supplementation Facilitates Cardiac Angiogenesis. To investigate the effects of L-arginine and. nistration on cardiac metabolism (Fig. 4). Neith-. L-ornithine supplementation on microvascular. er high- nor low-dose supplementation affected. remodeling, enzyme histochemical identification. CS, HAD a n d HAD:CS ratio. T h e r e f o r e , L-. of capillary profiles was performed on tissue. arginine and L-ornithine supplementation did. cross-sections. Figure 1 shows representative. not afFect aerobic capacity in cardiac muscle.. micrographic images of sections that demonstrate capillary and myofiber profiles from sub-. Discussion. endocardium portion of the LV. High-dose supplementation significantly i n c r e a s e d t h e. The present study showed that L-arginine and. capillary-to-fiber (C:F) ratio by 22% (P <O. 05;. L-ornithine supplementation caused marked. Fig. 2), while low-dose increased i t non-. angiogenesis by up-regulate VEGF expression.. significantly by 10%. Neither high- nor low-dose. In a previous study, administration of L-arginine. supplementation affected capillary density and. alone, a t similar levels as in the present study,. myofiber density. T h u s , 1,-arginine and 1,-. failed to induce evident angiogenesis and to. ornithine t r e a t m e n t facilitated angiogenesis. promote VEGF expression in cardiac muscle. dose-dependently in cardiac muscle.. (Maxwell et al. 2001). Thus, the interaction of L-arginine and L-ornithine may play a major. Western blot analysis was performed to in-. role in the present results.. vestigate VEGF and eNOS protein expression levels. Although supplementation increased. I n t h e p r e s e n t s t u d y , L-arginine a n d L-. VEGF levels dose-dependently, a significant1 in-. ornithine supplementation dose-dependently in-. crease was observed after high-dose treatment. creased VEGF protein levels (Fig. 3). VEGF is. (P <O. 05). eNOS expression slightly decreased. a 45 kDa secretable homodimeric glycoprotein. after the treatment, but defference was not sig-. and its expression is increased in response to. nificant. T h e r e f o r e , supplementation dose-. hypoxia (Schweiki e t al. 1992, Goldberg &. dependently up-regulated VEGF protein in car-. Schneider 1994). Recent findings demonstrate. diac muscle.. t h a t NO facilitates VEGF gene expression in numerous cell types, including vascular smooth. Metabolic enzyme activities and t h e en-. muscle cells (Dulak et al. 2000).. zymatic ratio were determined to investigate. L-Arginine is metabolized to NO by NOS. the effects of L-arginine and L-ornithine admi-. I VEGF. Cnt. *. Low. High eNOS. I Figure 3 Western blot analysis demonstrating effects L-arginine and L-ornithine supplementation on VEGF and eNOS protein expressions. All values are represented as means * SE. *, significantly different from the Cnt at. P<0.05..
(7) SUZUKI Junichi. I. Cnt. Low. High. I. HAD. Figure 4 Citratc synthasc and 6-hydroxyacyl-CoA dchydrogcnasc activities and thcir ratio valucs in lcft vcntriclc. All valucs arc rcprcscntcd as mcans * SE.. and to 1,-ornithine by arginase. 1,-Ornithine is. which indicated that dietary 1,-arginine adminis-. converted to putrescine by ornithine decarboxy-. tration failed to induce angiogenesis (Suzuki. lase (ODC) after which putrescine is converted. 2005). Even though arginase is inhibited by NO,. to spermidine and spermine. These polyamines. dietary supplementation of L-ornithine may help. are reported to promote mammalian cell growth. to produce polyamines and thus down-regulate. (Auvinen et al. 1992, Chang et al. 2001). Over-. endostatin. Administration of L-ornithine in. expression of ODC enhanced endothelial cell. addition to L-arginine may also help to metabol-. proliferation possibly by suppressing endostatin,. ize L-arginine more favorably to NO and not to. a carboxy-terminal fragment of type XVIII col-. ornithine. Since NO is known a s both a n up-. lagen, which inhibits endothelial proliferation,. stream and downstream mediator of VEGF ex-. angiogenesis, and tumor growth (Nemoto et al.. pression (Dulak & Jozkowicz 2003, Kimura &. 2002). Recently, endostatin was shown to influ-. Esumi 2003), supplementation in the present. ence VEGF signaling pathway in endothelial. study may facilitate VEGF production. There-. cells (Schmidt et al. 2005, 2006).. fore it is possible that L-arginine and L-ornithine. NO may be an inhibitor of ODC thus limiting polyamine synthesis (Hesse et al. 2001). N-. supplementation promotes cardiac angiogenesis by facilitating NO/VEGF interrelationships.. hydroxy-arginine (NOHA), t h e principal in-. L-ornithine is also converted to L-proline,. termediate in the NOS-catalyzed conversion of. which is essential for the synthesis of collagen,. arginine to NO and citrulline, has been reported. by o r n i t h i n e a m i n o t r a n s f e r a s e in v a s c u l a r. to be a competitive inhibitor of arginase, there-. s m o o t h muscle cells (SMC) ( D u r a n t e e t al.. by limiting L-ornithine production (Buga et al.. 2000) and endothelial cells (Li et al. 2001). Ap-. 1998). Thus, administration of L-arginine alone. plication of cyclic stretch to a SMC increased L-. may lead to NO production r a t h e r t h a n 1,-. arginine uptake, 1,-proline production and col-. ornithine. Decreased L-ornithine levels may up-. lagen s y n t h e s i s (Durante e l al. 2000). T h u s ,. regulate endostatin production and thus inhibit. mechanical f o r c e s to t h e SMC by c a r d i a c. VEGF-mediated angiogenesis. This notion is. contraction/relaxation cycle may facilitate col-. consistent with the results of a previous study,. lagen synthesis around microvascular beads,.
(8) LArginine and LOrnithine Supplementation Facilitates Cardiac Angiogenesis. a n d may help to stabilize proliferating cells thereby promoting angiogenesis during the pre-. nisms. Am J Physiol 275 : R1256-R1264 Cai J, Jiang WG, Ahmed A, Boulton M (2006) Vascular endothelial growth factor-induced endothelial cell pro-. sent supplementation.. liferation i s r e g u l a t e d b y i n t e r a c t i o n b e t w e e n. I n s u m m a r y , L-arginine a n d L-ornithine s u p p l e m e n t a t i o n significantly e n h a n c e d capillary-to-fiber ratio a n d i t markedly upregulated VEGF expression in left ventricle. The present results suggest that Larginine and. VEGFR-2, SH-PTP1 and eNOS. Microvasc Res 71: 20-31 Celec P, Gardlik R, Palffy R, Hodosy J, Stuchlik S, Drahovska H, Stuchlikova M, Minarik G, Lukacs J, Jurkovicova I, Hulin I, T u r n a J, Jakubovsky J, Kopani M, Danisovic L, J a n d z i k D, Kudela M, Y o n e m i t s u Y. facilitate. (2005) T h e use of transformed Escherichia coli for. angiogenesis by promoting a NO/VEGF interre-. experimental angiogenesis induced by regulated in. L-ornithine. treatment. may. lationship in cardiac muscle.. situ production of vascular endothelial growth factora n alternative g e n e therapy. Med Hypotheses 64: 505-511 Chang CI, Liao JC, Kuo L (2001) Macrophage arginase. ACKNOWLEDGEMENT T h i s w o r k w a s s u p p o r t e d in p a r t b y grants-in-aid f r o m Hokkaido University of Education.. promotes tumor cell growth and suppresses nitric oxide-mediated tumor cytotoxicity. Cancer Res 61: 1100-1106 Dulak J , Jozkowicz A, Dembinska-Kiec A, Guevara I, Zdzienicka A, Zmudzinska-Grochot D, Florek I, Wojtowicz A, Szuba A, Cooke JP (2000) Nitric oxide induces the synthesis of vascular endothelial growth. REFERENCES. factor by rat vascular smooth muscle cells. Arterioscler Thromb Vasc Biol 20: 659-666. Auvinen M, Paasinen A, Andersson LC, Holtta E (1992). Durante W, Liao L, Reyna SV, Peyton KJ, Schafer A1. Ornithine decarboxylase activity is critical for cell. (2000) Physiological cyclic stretch directs L-arginine. transformation. Nature 360: 355-358 B a r r e t t DM, Black SM, Todor H, Schmidt-Ullrich RK, D a w s o n KS, M i k k e l s e n RB (2005) I n h i b i t i o n of protein-tyrosine phosphatases by mild oxidative stresses is dependent on S-nitrosylation. J Biol Chem 280: 14453-14461 B a s s A , Brdiczka D, E y e r P, Hofer S, P e t t e D (1969) Metabolic differentiation of distinct muscle types a t t h e level of enzymatic organization. E u r J Biochem 10:198-206 Breen EC, Johnson EC, Wagner H, Tseng HM, Sung LA,. t r a n s p o r t and metabolism to collagen synthesis in vascular smooth muscle. FASEB J 14: 1775-1783 Folkman J, Klagsbrun M (1987) Angiogenic factors. Science 2351442-447 Goldberg MA, Schneider T J (1994) Similarities between the oxygen-sensing mechanisms regulating the expression of vascular endothelial growth factor and erythropoietin. J Biol Chem 26914355-4359 Hermansen L, Wachtlova M (1971) Capillary density of skeletal muscle in well-trained and untrained men. J Appl Physiol 30: 860-863. Wagner PD (1996) Angiogenic growth factor mRNA. Hesse M, Modolell M, La Flamme AC, Schito M, Fuen-. responses in muscle t o a single bout of exercise. J. t e s JM, Cheever AW, P e a r c e EJ, Wynn T A (2001). Appl Physiol 81 :355-361. Differential regulation of nitric oxide synthase-2 and. Brown LF, Yeo KT, Berse B, Yeo TK, Senger DR, Dvor-. arginase-1 by type l/type 2 cytokines in vivo: granu-. ak HF, van de Water L (1992) Expression of vascular. lomatous pathology i s shaped by t h e p a t t e r n of L-. permeability factor (vascular endothelial growth factor) by epidermal keratinocytes during wound healing. J Exp Med 176: 1375-1379 Buga GM, Wei LH, Bauer PM, F u k u t o JM, I g n a r r o LJ. arginine metabolism. J Immunol 167 :6533-6544 Hoffner L, Nielsen JJ, Langberg H, Hellsten Y (2003) Exercise but not prostanoids enhance levels of vascular endothelial growth factor and other proliferative. (1998) NG-hydroxy-L-arginine and nitric oxide inhibit. agents in human skeletal muscle interstitium. J Phy-. Caco-2 tumor cell proliferation by distinct mecha-. siol 550: 217-225.
(9) SUZUKI Junichi. Hudlicka 0, Brown MD, Silgram H (2000) Inhibition of capillary growth in chronically stimulated rat muscles by N (G)-nitro-1-arginine, nitric oxide synthase inhibitor. Microvasc Res 59 :45-51 Jakeman LB, Armanini M, Phillips HS, Ferrara N (1993). hances endothelial proliferation by suppressing endostatin expression. Blood 99 :1478-1481 Tomanek RJ (1970) Effects of age and exercise on the extent of the myocardial capillary bed. Anat Rec 167: 55-62. Developmental expression of binding sites and mes-. O'Reilly MS, Boehm T, Shing Y, Fukai N, Vasios G, Lane. s e n g e r ribonucleic acid f o r v a s c u l a r endothelial. WS, F l y n n E , B i r k h e a d JR, O l s e n BR, F o l k m a n J. growth factor suggests a role for this protein in vas-. (1997) E n d o s t a t i n : a n e n d o g e n o u s i n h i b i t o r of. culogenesis and angiogenesis. Endocrinology 133: 848-859 Jensen L, Bangsbo J, Hellsten Y (2004) Effect of high intensity training on capillarization and presence of. angiogenesis and tumor growth. Cell 88: 277-285 Patterson C, Runge MS (1999) Therapeutic angiogenesis: the new electrophysiology? Circulation 99: 2614-2616 Schmidt A, Wenzel D, Thorey I, Sasaki T , Hescheler J,. angiogenic factors in human skeletal muscle. J Phy-. Timpl R, Addicks K, Werner S, Fleischmann BK &. siol 557 :571-582. Bloch W (2006) Endostatin influences endothelial. Kimura H, Esumi H (2003) Reciprocal regulation be-. morphology via t h e a c t i v a t e d ERKl/2-kinase en-. tween nitric oxide and vascular endothelial growth. dothelial morphology and signal transduction. Micro-. factor in angiogenesis. Acta Biochem Pol 50: 49-59 K r a u s RM, S t a l l i n g s HWSrd, Y e a g e r RC, Gavin T P. vasc Res 71 :152-162 Shweiki D, Itin A, Soffer D, Keshet E (1992) Vascular. (2004) Circulating plasma VEGF response to exercise. endothelial growth factor induced by hypoxia may. in sedentary and endurance-trained men. J Appl Phy-. mediate hypoxia-initiated angiogenesis. Nature 359:. siol 96: 1445-1450. 843-845. Kroll J , W a l t e n b e r g e r J (16999) A novel function of. Srere PA (1969) Citrate synthase. Meth Enzym 13:3-5. VEGF receptor-2 (KDR): rapid release of nitric oxide. Suzuki J (2002) Microvascular remodelling after endur-. in response to VEGF-A stimulation in endothelial. a n c e t r a i n i n g with Co2 + t r e a t m e n t in t h e r a t di-. cells. Biochem Biophys Res Commun 265 :636-639. a p h r a g m a n d hind-leg muscles. J p n J Physiol 52:. Li H, Meininger CJ, Hawker JRJr, Haynes T E , Kepka-. 409-419. L e n h a r t D, M i s t r y SK, M o r r i s S M J r , Wu G (2001). Suzuki J (2005) Microvascular angioadaptation after en-. Regulatory role of arginase I and I1 in nitric oxide,. durance training with L-arginine supplementation in. polyamine, and proline syntheses in endothelial cells.. r a t h e a r t a n d h i n d l e g m u s c l e s . E x p Physiol 90:. Am J Physiol Endocrinol Metab 280: E75-E82. 763-771. Z (1979) Studies on dipeptidyl (amino) peptidase. Wittwer M, Billeter R, Hoppeler H, Fluck M (2004) Reg-. IV (glycyl proline norphthylamidase). 11. Blood ves-. ulatory gene expression in skeletal muscle of highly. sels. Histochemistry 59 :153-166. endurance-trained humans. Acta Physiol Scand 180:. Lojda. Maxwell AJ, Ho HV, Le CQ, Lin PS, Bernstein D, Cooke. 217-227. J P (2001) L - a r g i n i n e e n h a n c e s a e r o b i c e x e r c i s e. Zhao X, Lu Z, F e n g Q (2002) Deficiency in endothelial. capacity in association with augmented nitric oxide. nitric oxide synthase impairs myocardial angiognesis.. production. J Appl Physiol 90: 933-938. Am J Physiol Heart Circ Physiol 2831H2371-H2378. Murohara T , Asahara T , Silver M, Bauters C, Masuda H, Kalka C, Kearney M, Chen D, Symes JF, Fishman MC, Huang PL, Isner JM (1998) Nitric oxide syntha s e modulates angiogenesis in response to tissue ischemia. J Clin Invest 101:2567-2578 N a k a g a m i H, Cui T X , I w a i M, S h i u c h i T , T a k e d a Matsubara Y, Wu L, Horiuchi M (2002) Tumor necrosis factor-alpha inhibits growth factor-mediated cell proliferation through SHP-1 activation in endothelial cells. Arterioscler Thromb Vasc Biol 22: 238-242 Nemoto T , Hori H, Yoshimoto M, Seyama Y, Kubota S (2002) Overexpression of ornithine decarboxylase en-.
(10)
図



関連したドキュメント
第四章では、APNP による OATP2B1 発現抑制における、高分子の関与を示す事を目 的とした。APNP による OATP2B1 発現抑制は OATP2B1 遺伝子の 3’UTR
Hugh Woodin pointed out to us that the Embedding Theorem can be derived from Theorem 3.4 of [FM], which in turn follows from the Embedding Theorem for higher models of determinacy
Nintendo Switchでは引き続きハードウェア・ソフトウェアの魅力をお伝えし、これまでの販売の勢いを高い水準
このように、このWの姿を捉えることを通して、「子どもが生き、自ら願いを形成し実現しよう
Q-Flash Plus では、システムの電源が切れているとき(S5シャットダウン状態)に BIOS を更新する ことができます。最新の BIOS を USB
「新老人運動」 の趣旨を韓国に紹介し, 日本の 「新老人 の会」 会員と, 韓国の高齢者が協力して活動を進めるこ とは, 日韓両国民の友好親善に寄与するところがきわめ
本プログラム受講生が新しい価値観を持つことができ、自身の今後進むべき道の一助になることを心から願って
を育成することを使命としており、その実現に向けて、すべての学生が卒業時に学部の区別なく共通に
