養殖海域の大腸菌群数をもとにした冬季マガキのノ
ロウイルス汚染の予測
著者
加藤 晃博
学位授与機関
Tohoku University
養殖海域の大腸菌群数をもとにした
冬季マガキのノロウイルス汚染の予測
専 攻 資源生物科学専攻
指導教官 高橋計介 准教授
学籍番号 A6AM1116
氏 名 加藤晃博
日付 平成 20 年 1 月 18 日
【目次】 第1 章 序論 1 第 2 章 海 水 の 大 腸 菌 群 数 と マ ガ キ の ノ ロ ウ イ ル ス 陽 性 率 の 季 節 変 動 7 第 3 章 海水の大腸菌群数をもとにしたマガキのノロウイルス汚染の予測方法の検討 29 第1節 夏季における海水の大腸菌群数と冬季マガキのノロウイルス陽性率との関係 29 第2節 リアルタイム PCR を用いたノロウイルス遺伝子数の測定と大腸菌群数との関係 39 第4章 総合考察 52 要約 56 謝辞 58 参考文献 59
第 1 章 序 論 わが国 における平 成 18 年 度 の食 中 毒 の発 生 件 数 は 1,491 件 であり、患 者 数 は 39,026 名 となっている。病 因 物 質 別 に見 ると細 菌 とウイルスが主 たる病 因 とな っており (厚 生 労 働 省 :食 中 毒 ・食 品 監 視 関 連 情 報 による)、その割 合 は件 数 で は細 菌 51.9%、ウイルス 33.8%で細 菌 の方 が多 い。しかし病 因 物 質 の中 身 を見 る と細 菌 ではカンピロバクター・ジェジュニ/コリをはじめとして様 々な種 類 の菌 が原 因 となっているのに対 し、ウイルスでは患 者 数 、件 数 ともに 99%以 上 をノロウイル スが占 めている。また、ノロウイルスによる患 者 数 は 27,616 名 となっており食 中 毒 による全 患 者 数 の約 70%を占 め、第 一 位 となっている。このようにノロウイルスは 食 中 毒 の病 因 物 質 として大 変 大 きな存 在 となっている。 ノロウイルス胃 腸 炎 の症 状 としては嘔 気 、嘔 吐 、下 痢 が主 症 状 であるが、腹 痛 、 頭 痛 、発 熱 、悪 寒 、筋 痛 、咽 頭 痛 、倦 怠 感 などを伴 うこともある。ボランティアへ の投 与 試 験 の結 果 から、潜 伏 期 は 1~2 日 であると考 えられている (入 谷 ら, 2003)。ノロウイルスはヒトの小 腸 上 皮 細 胞 の組 織 ‐血 液 型 抗 原 (ABH 抗 原 ) を レセプターとしていると考 えられ (Harrington et al., 2002)、細 胞 に接 着 し、感 染 する。ノロウイルスは電 顕 観 察 による形 態 からかつては小 型 球 形 ウイルス
(SRSV: small round-structured virus) と呼 ばれた。また 1968 年 オハイオ州 ノ ーウォークの小 学 校 で集 団 発 症 した冬 季 嘔 吐 症 についての研 究 において 、患 者 糞 便 試 料 のボランティアへの経 口 投 与 実 験 により病 気 の再 現 がなされ、さらに免 疫 電 顕 観 察 によって糞 便 試 料 中 に初 めて食 中 毒 の原 因 物 質 として小 型 球 形 ウ イルスが発 見 されたことから明 らかになった (Kapikian et al., 1972)。本 症 例 が発 生 した地 名 を冠 してこのウイルスは Norwalk virus とも呼 ばれるようになった。現
在 では、Norwalk virus とこれに似 た Norwalk-like virus はカリシウイルス属
(Family Calicividae) のノロウイルス科 (Genus Norovirus) に分 類 されている。 わが国 では 1997 年 5 月 に改 正 された食 品 衛 生 法 で、食 中 毒 原 因 物 質 に小 型 球 形 ウイルスが加 えられた。さらに 2003 年 9 月 の食 品 衛 生 法 の改 正 によって、小 型 球 形 ウイルスという名 称 がノロウイルスに変 更 された (入 谷 , 2003)。ノロウイ ルスは正 20 面 体 (直 径 27~40nm) の RNA (プラス 1 本 鎖 ) ウイルスであり、 表 面 は不 斉 の突 起 状 構 造 を有 する (Caul et al., 1982)。遺 伝 子 の全 長 は 7.5~ 7.7kb で 3 つの open reading frame (ORF) を含 み、ORF 1 は RNA 依 存 性 RNA ポリメラーゼなどの非 構 造 タンパク質 を、ORF 2 はウイルス外 皮 タンパク質 を、そ して ORF 3 は機 能 不 明 のタンパク質 を、それぞれコードしている (入 谷 ら, 2003、 Jiang et al., 1993、 Lambden et al., 1993)。ノロウイルスは RNA の配 列 から大 き く二 つの遺 伝 子 群 GenogroupⅠ (GⅠ)、GenogroupⅡ (GⅡ) に大 別 される。さ らに GⅠは 14 以 上 、GⅡでは 17 以 上 の遺 伝 子 型 (Genotype) に分 かれている。 現 在 このウイルスの培 養 実 験 系 は確 立 されておらず、ウイルスの活 性 測 定 がで きない。このことがノロウイルス研 究 において大 きな問 題 となっている。本 ウイルス の検 出 は電 顕 観 察 か、RT-PCR、Real-time PCR といったウイルスの遺 伝 子 を検 出 する方 法 が行 われている (Jiang et al.,1992、Kageyama et al., 2003)。
食 品 から病 因 物 質 を検 出 ・特 定 することは容 易 ではないが、特 にノロウイルス は食 品 からの検 出 が難 しい。そのため、ノロウイルスを原 因 とする 食 中 毒 事 案 の 70%近 くにおいて原 因 食 品 が特 定 されていない。そのような状 況 ではあるが、食 中 毒 の原 因 食 品 として特 定 された食 品 の中 でもっとも多 いのは二 枚 貝 である。その 理 由 は二 枚 貝 がろ過 食 性 の水 生 動 物 であり 、環 境 水 中 のウイルス粒 子 を餌 の 取 り 込 み と と も に 行 い 、 濃 縮 を す る か ら で あ る と 考 え ら れ て い る ( 室 賀 ・ 高 橋 ,
2005)。ノロウイルスが検 出 された二 枚 貝 としてはマガキ Crassostrea gigas 、ア メリカガキ C. verginica (Le Guyader et al., 2000、Burkhardt Ⅲ et al., 2000), ヨ ーロッパヒラガキ Ostrea edulis (Beuret et al., 2003)、ムラサキイガイ Mitilus galloprovincialis (杉 山 ら, 2002)、 ヨーロッパイガイ M. edulis (Lee, 1995)、 アサリ Ruditapes philippinarum (Kingsley, 2002) が報 告 されている。もちろん、 ノロウイルス食 中 毒 の原 因 食 品 は貝 類 だけではな い。原 因 として最 も多 いのは、 食 中 毒 による一 次 感 染 者 から糞 口 感 染 として、二 次 的 に感 染 することである。患 者 の嘔 吐 物 を 原 因 とする こと もある。 このよう に、二 枚 貝 が主 たる原 因 とは言 い 難 い面 もあるが、原 因 食 品 の1つであることは事 実 であり、二 枚 貝 のノロウイルス 汚 染 を防 止 する、あるいは汚 染 された貝 からノロウイルスを排 除 するなど対 策 が 必 要 である。 上 述 の二 枚 貝 の中 でも、マガキはノロウイルス食 中 毒 の原 因 食 品 として特 に 注 目 されている。これはろ過 食 性 という点 に加 えて、マガキは生 もしくは十 分 加 熱 せずに食 べられることが多 いこと、さらに、マガキは他 の多 くの二 枚 貝 と異 なり、 内 臓 ごと丸 ごと摂 食 される場 合 が多 いことが理 由 であるとされる (Cliver ,1997、 Parashar, 2001)。また、ノロウイルス胃 腸 炎 患 者 が増 加 する冬 季 に出 荷 される 食 品 であるということも大 きな要 因 であると考 えられる。カキ類 が海 水 中 に拡 散 し たノロウイルスを取 り込 むことは実 証 されており (Schwab et al., 1998)、また、河 川 水 、海 洋 におけるノロウイルス汚 染 経 路 解 明 のための調 査 ・研 究 において、胃 腸 炎 患 者 -河 川 -海 域 -カキという汚 染 経 路 も実 証 されている (植 木 , 2003、植 木 , 2004)。これらのことから、程 度 の違 いはあれ養 殖 場 から取 り上 げたカキの中 には、 ノロウイルスを取 り込 んだものが含 まれる可 能 性 は否 定 できない。したがって、そ のまま出 荷 してしまうことがないようにと生 産 現 場 での対 応 が求 められ、例 えば宮
城 県 ではノロウイルスの出 荷 前 検 査 が行 われている。しかし、これは抜 き取 り検 査 であり、ノロウイルス汚 染 カキの出 荷 を完 全 に防 ぐことは難 しい。 カキの食 中 毒 への対 策 として浄 化 処 理 (depuration) がある。これは漁 獲 し たカキを水 槽 などに収 容 し、清 浄 な海 水 を1~2日 ほどかけ流 すことにより、カキ に含 まれる病 原 微 生 物 を除 去 あるいは減 尐 させることを目 的 とした方 法 である。 浄 化 処 理 に関 して様 々なウイルスで研 究 がなされている。例 えば、アデノウイルス (Adenovirus) をヨーロッパイガイとヨーロッパヒラガキに取 り込 ませた実 験 では、 水 温 18℃においても RT-PCR により 42 日 間 後 に検 出 されている (Hernroth and Allard, 2007)。また、アメリカガキにおいて 22℃で 48 時 間 浄 化 したときに、消 化 盲 嚢 において大 腸 菌 は 95%減 尐 したのに対 して、ノロウイルスは 7%しか減 尐 しなか った (Schwab et al., 1998)。ノロウイルスの代 替 ウイルスとして用 いられるネコカ リシウイルス (Feline calicivirus: FCVF4) を用 いたカキの浄 化 実 験 では、カキ から検 出 されなくなるまで 96 時 間 を要 している (山 木 ら, 2003)。すなわち、これら の結 果 から浄 化 処 理 は細 菌 を減 尐 させるためには有 効 であっても、ウイルスに対 しては細 菌 と同 様 の減 尐 効 果 を期 待 するのは難 しいと考 えられる。 現 在 、食 品 としてのカキの安 全 性 の指 標 として、養 殖 海 域 の大 腸 菌 群 数 が用 いられている。わが国 ではカキ養 殖 海 域 の基 準 は海 水 100 mLあたりの大 腸 菌 群 数 (最 確 数 Most Probable Number: MPN) が 1000 以 下 であり、生 食 用 カキの 養 殖 環 境 は MPN が 70 以 下 であることが条 件 として定 められている (水 産 用 水 基 準 ; 2000)。さらに生 食 用 カキに対 しては、食 品 衛 生 法 でカキの成 分 規 格 と して一 般 細 菌 数 50000/g 以 下 、大 腸 菌 群 数 MPN 230/100g 以 下 、腸 炎 ビブリ オ MPN 100/g 以 下 という基 準 が定 められている。しかし、このように細 菌 を用 い た基 準 は細 菌 性 の食 中 毒 を防 ぐ効 果 があるが、ノロウイルス胃 腸 炎 やその他 の
ウイルス性 の感 染 症 に対 しては、必 ずしも有 効 なものではないとする多 くの報 告 がある (Lee, 2000、Cook et al., 1986、Richerds, 1988)。したがって、ノロウイルス による食 中 毒 を避 けるためには、ノロウイルス汚 染 そのものを評 価 する必 要 であ ると考 えられる。しかし現 在 、ノロウイルスの培 養 法 は確 立 されておらず、生 きた (感 染 力 を持 った) ウイルスの定 量 はできない。そこでノロウイルスに代 わる指 標 が求 められる。条 件 としては、ノロウイルスの動 態 とよく一 致 し、定 量 性 があること などが求 められる。候 補 としてはバクテリオファージ (Doré, 1998) が挙 げられて いるが、決 定 的 な代 替 指 標 はまだ確 立 していない。 本 研 究 では、養 殖 漁 場 におけるノロウイルス汚 染 の指 標 として大 腸 菌 群 数 を 用 いることができるのかを検 討 した。上 で述 べたように、大 腸 菌 群 数 はノロウイル スや他 の感 染 性 ウイルスの汚 染 を知 るための指 標 には適 さないともいわれる。そ れにもかかわらず、あえて大 腸 菌 群 数 を取 り上 げた理 由 は以 下 の通 りである。 漁 場 へのノロウイルス流 入 は糞 便 汚 染 とともに生 じると考 えられるため、下 水 の影 響 の大 きさを知 ることがノロウイルス汚 染 の程 度 を知 ることにつながる。そこで、 糞 便 汚 染 の指 標 として用 いられるのは大 腸 菌 群 数 であり、これを調 べることは間 接 的 にせよノロウイルス汚 染 の状 況 を知 ることになると考 えられるからである。ま た、前 述 の通 り、大 腸 菌 群 数 は、カキ漁 場 の評 価 基 準 として定 められており、測 定 機 会 が多 く、大 量 のデータがこれまでに集 積 され、今 後 も集 められていくことに なる。すなわち、養 殖 海 域 における微 生 物 (細 菌 、ウイルスを含 めた) のデータ として最 も充 実 したものであり、生 かし方 によってはノロウイルス汚 染 の指 標 にな り得 ると考 えたからである。本 研 究 では具 体 的 に、宮 城 県 内 の 8 箇 所 のカキ養 殖 場 において RT-PCR を用 いて養 殖 マガキのノロウイルス汚 染 の現 状 を把 握 すると ともに、養 殖 海 域 の汚 染 度 を大 腸 菌 群 数 により評 価 し て両 者 の関 係 性 を調 べ、
大 腸 菌 群 数 がマガキ養 殖 海 域 における冬 季 ノロウイルス汚 染 リスクの指 標 となり 得 るか評 価 することを試 みた。
第 2 章 大 腸 菌 群 数 とノロウイルス陽 性 率 の季 節 変 動 食 中 毒 原 因 ウイルスとして問 題 となっているノロウイルスはヒトで感 染 、増 殖 し たのちに糞 便 とともに排 出 される。下 水 処 理 を経 て不 活 化 されなかった一 部 のウ イルスがマガキなどのろ過 食 性 の二 枚 貝 などに餌 とともに取 り込 まれて蓄 積 、濃 縮 することが知 られている (Ueki et al., 2005; 室 賀 ・高 橋 , 2005)。そのため、宮 城 県 で盛 んなマガキ養 殖 業 にとってノロウイルス汚 染 は深 刻 な問 題 となっている。 ノロウイルスは 85℃、1 分 の加 熱 により不 活 化 されるものの、宮 城 県 産 マガキは 生 食 用 として出 荷 することが多 く、ノロウイルス汚 染 状 況 の把 握 が強 く求 められる。 海 水 中 に低 濃 度 で拡 散 している状 態 のノロウイルスを検 出 することは困 難 である。 また、ノロウイルスは現 在 培 養 系 が確 立 されていないので代 替 指 標 が必 要 となっ てい る。現 在 までにバクテ リ オファージ などが指 標 とし て提 案 されてい るものの、 現 在 のところ決 定 的 な指 標 は定 められていない。現 在 マガキの食 品 衛 生 上 の安 全 性 の指 標 としては、大 腸 菌 群 数 が養 殖 海 域 の水 質 の糞 便 汚 染 の指 標 として 用 いられているが、これはノロウイルス汚 染 の指 標 として 定 められているものでは ない。 本 章 では天 然 、養 殖 マガキの体 内 からノロウイルスを RT-PCR により検 出 する とともに、マガキ生 息 海 域 の海 水 の大 腸 菌 群 数 を測 定 し、季 節 変 動 を見 ることで 糞 便 汚 染 の指 標 である大 腸 菌 群 数 がマガキのノロウイルス汚 染 の指 標 となるか を検 討 した。 【材 料 と方 法 】 マガキ
マガキ (Crassostrea gigas thunberg 1793) のサンプリング地 点 として宮 城 県 沿 岸 の 3 箇 所 (Bay X、Farm A、Farm B)を設 定 し、そのうち Bay X では天 然 マガ キを、その他 場 所 では養 殖 マガキを採 取 した。Bay X、Farm A、Farm B では 2005 年 4 月 から 2006 年 3 月 にかけて各 月 1 回 ずつマガキを採 取 した (各 月 8~30 個 体 )。採 取 したマガキの平 均 殻 高 (mm)、殻 長 (mm)、殻 幅 (mm)、消 化 盲 嚢 重 量 (g) は Bay X: 68.3±17、42.6±10、26.1±6、0.5±0.3、Farm A: 120.8 ±13、53.8±7、32.6±6、1.3±0.5、Farm B: 126.6±15、60.8±8、33.9±5、2.1 ±0.9 であった。採 取 したマガキはクーラーボックス、またはコンテナに入 れ輸 送 し、 48 時 間 以 内 に RNA 抽 出 を行 った。当 日 RNA 抽 出 ができなかった個 体 は水 槽 に 入 れず、4 ℃に保 った低 温 室 に保 管 した。RT-PCR の陽 性 対 照 用 のウイルスとし て、広 島 大 学 から分 与 されたノダウイルス科 の striped jack nervous necrosis virus (SJNNV) を用 いた。なお SJNNV はシマアジのウイルス性 神 経 壊 死 症 の原 因 ウイルスであり、ノロウイルスと同 じくプラス 1 本 鎖 RNA を持 っている。 海 水 上 記 のマガキのサンプリング地 点 と同 じところから表 層 の海 水 を採 取 した。採 取 した海 水 はクーラーボックスに入 れて運 搬 し、採 取 後 6 時 間 以 内 に試 験 に供 し た。海 水 の水 温 と塩 分 濃 度 も同 時 に測 定 した。 1. マガキ組 織 からのノロウイルスの検 出 方 法 ノロウイルスの検 出 は、組 織 の摘 出 、RNA 抽 出 、DNase 処 理 、RT 反 応 、1st PCR、nested PCR、電 気 泳 動 の 7 段 階 の手 順 に従 って行 った。 組 織 の摘 出 カキ剥 きナイフを用 いて開 殻 し、マガキの軟 体 部 を取 り出 し、次 に乾 熱 滅 菌 し
た解 剖 ばさみを用 いて胃 を含 む消 化 盲 嚢 を摘 出 し、個 体 ごとにはさみは交 換 した。 組 織 片 ごとに約 10 mL の蒸 留 水 を加 え、ビーズ式 細 胞 破 砕 装 置 (MS-100: トミ ー精 工 社 ; 直 径 3.2 mm ステンレスビーズ 2 個 使 用 ) を用 いて 4.500 rpm、1 分 間 の条 件 で組 織 を 破 壊 し 磨 砕 液 を 得 た。最 後 に遠 心 分 離 (9.000 ×g、10 分 間 ) し、得 られた上 清 を RNA 抽 出 に用 いた。 RNA 抽 出
QIAamp Viral RNA Mini Kit (キアゲン社 ) を用 いて、プロトコールに概 ね従 って 以 下 のようにしてRNA を抽 出 した。まず1.5 mL チューブにキット付 属 のCarrier RNA を含 むBuffer AVL 560 μ L を注 入 し、上 記 の処 理 サンプル140 μ L を添 加
し、15秒 間 撹 拌 し、室 温 で10分 間 静 置 した。100%エタノール560 μ L をサンプルに 添 加 し15秒 間 撹 拌 した。溶 液 の半 量 (630 μ L) をキット付 属 のスピンカラムに注 入 し、遠 心 分 離 (6,000 ×g 、1 分 間 ) を行 った。このスピンカラムをコレクション チューブから外 し、濾 液 を廃 棄 し、残 りのサンプル溶 液 (630 μ L) を加 え、再 度 同 様 に遠 心 分 離 を行 った。濾 液 を廃 棄 後 、キット付 属 のBuffer AW 1 を500 μ L を 添 加 し、遠 心 分 離 (6,000 ×g 、1 分 間 ) し濾 液 を廃 棄 し、続 いてBuffer AW 2 を500 μ L を加 え、遠 心 分 離 (18,000×g 、3 分 間 ) を行 った。なお、これらの遠 心 操 作 のたびにコレクションチューブを新 しいものに交 換 した。スピンカラムを高 温 加 圧 滅 菌 した容 積 1.5 mL マイクロチューブに移 し、濾 液 は廃 棄 した。スピンカラム を開 き、室 温 に戻 したBuffer AVE 60 μ L を添 加 し、キャップを閉 めて室 温 で1 分 間 静 置 した後 、遠 心 分 離 (6,000 ×g 、1 分 間 ) を行 い、高 温 加 圧 滅 菌 した0.5 mL のPCR チューブに分 注 しRNA 抽 出 物 を得 た。このRNA 抽 出 物 をDNase 処 理 するまで-30℃ で保 存 した。
DNase 処 理
抽 出 したRNA サンプルとRQ 1 DNase (プロメガ社 ) を混 合 したDNase reaction mixture(表 1)をPCR チューブ (0.2 mL 容 、以 下 同 様 ) に入 れ37℃ 、30 分 間 静 置 しDNase 処 理 を施 した。次 にTerminate reaction mixture (表 1) を別 のPCR チューブに入 れ、75℃ 、10 分 間 静 置 し、DNaseを失 活 させたあと4℃に保 ち速 や かに以 下 に示 すRT 反 応 に移 行 した。なお、加 熱 冷 却 処 理 はサーマルサイクラー (TaKaRa PCR Thermal Cycler Dice Gradient: タカラ社 、あるいはGeneAmp® PCR System: アプライドバイオシステムズ社 ) を用 い、これ以 降 の熱 処 理 も同 機 材 を用 いて行 った。なお、ネガティブコントロールとして、蒸 留 水 を用 いたサンプルを 作 成 した。
RT 反 応
DNase 処 理 後 のTerminate reaction mixture に逆 転 写 酵 素 であるSuperscript Ⅱ (インビトロジェン社 ) を添 加 し、表 2 に示 したRT mixture をPCR チューブ中 に作 製 した。次 に42℃ 、60 分 間 の熱 処 理 で逆 転 写 反 応 を行 い、その後 99℃ 、5 分 間 加 熱 し、RT 酵 素 を失 活 させた後 4℃ に静 置 し、次 の1st PCR に備 えた。
1st PCR
上 のRT 反 応 で得 た鋳 型 DNA からDNA ポリメラーゼ (EX Taq HotStart Version: タカラ社 ) を用 いてPCR により目 的 のDNA を増 幅 した。すなわち、表 3 に示 す1st PCR mixture をPCR チューブ中 に用 意 し、94℃ ・3分 間 を1サイクル、 94℃ ・1分 間 、50℃ ・1分 間 、72℃・2分 間 を40サイクル行 った後 72℃・15 分 間 反
応 させ、次 のnested-PCR を行 うまで4℃ で静 置 した。 Nested PCR 表 3 に示 したnested PCR mix を新 しいPCR チューブの中 に用 意 し、1st PCR と同 じ条 件 で1st PCR 産 物 から目 的 DNA を増 幅 した。 プライマー ノロウイルスには2つの主 要 遺 伝 子 群 GⅠおよびGⅡ が知 られており、それぞれ の遺 伝 子 群 を別 々に検 出 する必 要 がある。RT 反 応 、1stPCR および nested-PCR にはそれぞれの遺 伝 子 群 に固 有 のプライマーを用 い、すべてのプラ イマーは高 い保 存 性 があるとされるORF 1 とORF 2 の結 合 領 域 に位 置 している (Kageyama et al., 2003)。最 初 にプライマーは厚 生 労 働 省 医 薬 食 品 局 食 品 安 全 部 監 視 安 全 課 (食 安 監 ) 発 第 1105001号 に示 されたノロウイルスの検 出 法 に基 づ き遺 伝 子 群 GⅠ の検 出 に対 して、1st PCR ではプライマーセットCOG1F-G1SKR、 GⅡの検 出 に対 してはCOG2F-G2SKR を用 いた。また、nested-PCR においては プライマーセットG1SKF-NEWG1R (GⅠ用 )、G2SKF-NEWG2R (GⅡ用 ) を用 いた (前 川 , 2006)。これらのプライマーの塩 基 配 列 を表 4 に示 した。なお、1stPCRで生 じるPCR 産 物 に期 待 される塩 基 数 は381 bp (GⅠ)、387 bp (GⅡ) であり、 nested-PCRの産 物 に期 待 される塩 基 数 は252 bp(GⅠ)、216 bp(GⅡ)であった。 陽 性 対 照 として用 いたSJNNV に関 してはNishizawa et al.(1994、1997) に従 い、 1st PCR では外 被 膜 タンパク遺 伝 子 領 域 のTergetregion 4 を標 的 とするプライマ ーセットを用 いた (PCR 産 物 の予 想 塩 基 数 は約 430 bp)。さらにnested-PCR で はさらにその内 側 を標 的 とするプライマーセット (Forward primer:
5’-CTTGAGACACCTGAGGACACCAC-3’、 Reverse primer: 5’-CCCAACTGTGAATGTCTTGTTAAAGT-3’) を用 いた(PCR 産 物 の予 想 塩 基 数 は約 310 bp)。 PCR 産 物 の観 察 5×TBE (ニッポンジーン社 ) を蒸 留 水 で10 倍 に希 釈 し、0.5×TBEとして電 気 泳 動 用 のバッファーとした。PCR 産 物 10 μ L は6×Loadingbuffer (タカラ社 ) 2 μ L と混 合 し、SYBR Safe™ DNA gel stain (0.5×TBE)(インビトロジェン社 ) を用
いて作 製 した1.5%アガロースゲルにて電 気 泳 動 し、トランスイルミネーター (Vilber Lourmat社 ) によるUV 照 射 を行 い、紫 外 線 吸 収 フィルター (SC 50:富 士 フィルム 社 ) を通 して観 察 し、PCR 産 物 のバンドの有 無 を確 認 した。電 気 泳 動 の結 果 より マガキのノロウイルス陽 性 率 を算 出 した。 [ノロウイルス陽 性 率 =(ノロウイルス陽 性 個 体 )/(ノロウイルス検 査 個 体 )× 100] ノロウイルス陽 性 個 体 : ノロウイルスG1、G2どちらかが検 出 された個 体 PCR産 物 のシークエンス Nested PCR 産 物 を 電 気 泳 動 に か け 予 想 塩 基 数 付 近 の バ ン ド を 切 り 出 し 、 MinEluteT M Gel Extraction Kit (キアゲン社 ) を用 いて精 製 し、バイオマトリック
ス研 究 所 (千 葉 ) に分 析 を依 頼 した。
2. 海 水 の大 腸 菌 群 数 の測 定 方 法
大 腸 菌 群 数 の測 定 は 『沿 岸 環 境 調 査 マニュアルⅡ(水 質 ・微 生 物 篇 )』 (日 本 海 洋 学 会 編 、 恒 星 社 厚 生 閣 ) に従 い、乳 糖 ブイヨン培 地 「ニッスイ」 (日 水 製 薬 ) による MPN 法 を行 った。
滅 菌 した希 釈 水 (リン酸 緩 衝 液 : PBS、pH 7.0) を用 いて試 料 (海 水 ) の 10 倍 希 釈 系 列 (100~10- 4) を作 製 し、各 希 釈 液 1 mL ずつを 5 本 (または 3 本 ) の
培 地 とダーラム管 を入 れた発 酵 管 に接 種 した。接 種 した試 験 管 を 35±1℃で培 養 し、48±3 時 間 後 に観 察 し、培 地 の色 が黄 ないし黄 緑 色 に変 化 し、ガスが発 生 したものを陽 性 とした。試 料 の希 釈 段 階 と大 腸 菌 群 陽 性 の試 験 管 数 から試 料 中 の菌 数 を最 確 数 (Most Probable Number: MPN) 計 数 表 より求 めた。
【結 果 】 マガキのノロウイルス陽 性 率 の季 節 的 変 動 今 回 、マガキの消 化 盲 嚢 を切 り出 しウイルスの RNA を抽 出 し、RT-PCR を用 いて ノロウイルスを検 出 することでノロウイルス汚 染 状 況 の季 節 的 変 動 を調 べた (図 1、 表 5)。調 査 した 3 つのサンプリング場 所 における結 果 で、7 月 から 11 月 までの夏 季 を中 心 とした時 期 におい てマガキのノ ロウイルス汚 染 は ほとんど 確 認 さ れず、 12 月 から翌 年 3 月 までの冬 季 には程 度 の差 はあるものの、3 箇 所 全 てにおいて マガキからノ ロウイルスが検 出 さ れるという共 通 し た結 果 が得 られた。 冬 季 の陽 性 率 を見 ると、Farm A、Bay X は高 く、それぞれ検 査 個 体 が全 て陽 性 となる月 も 存 在 するが、一 方 で Farm B ではわずかしか検 出 されておらず、検 出 される期 間 も 他 の 2 箇 所 が 12 月 から 3 月 まで検 出 されたのに対 して、Farm B は 12 月 から 2 月 までと短 かった。表 5 に G1、G2 ごとの季 節 変 動 の結 果 を示 したが、これを見 る と、G1 よりも G2 がより多 くの個 体 から検 出 された。Farm A、Bay X では G2 の陽 性 率 が 100 % になる月 (Farm A:1 月 、Bay X:2 月 、3 月 ) があるなど冬 季 のノロ ウイルス陽 性 率 は高 かった。それに対 して G1 では 1 月 の Bay X の 40 % (6 個 体 /15 個 体 ) が最 も高 い値 であり、Farm A でも検 出 されるものの、5 月 の 10 % (2/20)、2 月 の 3% (1/26) というように G2 の陽 性 率 と比 べると低 く、Farm B で は冬 季 においても G1 は全 く検 出 されないという結 果 であった。 海 水 の大 腸 菌 群 数 の季 節 変 動 毎 月 1 回 マガキを採 取 する際 に表 層 の海 水 を汲 み取 り、乳 糖 ブイヨンを用 いて大 腸 菌 群 数 を測 定 し、マガキとともに季 節 変 動 を調 べた (図 2、表 6)。調 査 した 3
つのポイントにおいて、6 月 から 9 月 を中 心 に値 が高 く、特 に 7 月 に一 年 で最 も値 が高 くなり、マガキからノロウイルスが検 出 される 冬 場 にかけて減 尐 するという傾 向 が共 通 して見 られた。冬 季 にマガキからノロウイルス陽 性 率 が高 かった Bay X と Farm A は夏 季 を中 心 に大 腸 菌 群 数 が多 く、Farm A では 7 月 に MPN が 16000 と高 い値 が得 られた。Bay X は 7 月 の値 が Farm A ほど高 くないものの、5 月 から 10 月 にかけて Farm A よりも高 い値 が検 出 され、他 の 2 点 では検 出 限 界 以 下 と なった 2 月 、3 月 でも MPN が 490、1700 と高 い値 が得 られた。一 方 、冬 季 マガキ のノロウイルス陽 性 率 が他 の 2 点 と比 べて低 かった Farm B では 6 月 から 10 月 にかけて菌 は検 出 されるものの、最 も高 くても 7 月 の 490 と低 い値 であり、それ以 外 の月 では検 出 限 界 以 下 であった。
【考 察 】 ノロウイルスはヒトの糞 便 とともに排 出 され 、下 水 処 理 場 を経 て不 活 化 されず に残 った一 部 が海 域 に放 出 され、カキなどの二 枚 貝 類 が汚 染 されると考 えられて い る。そのためカキのノ ロウイルス汚 染 と 糞 便 汚 染 の指 標 である大 腸 菌 群 数 の 季 節 変 動 を調 べ両 者 の関 係 を見 た。 海 水 の大 腸 菌 群 数 は水 温 の上 昇 する夏 季 に増 加 し、今 回 調 査 した 3 点 のす べてにおいて 7 月 に最 も高 い値 を示 し、その後 水 温 の低 下 する秋 には数 が減 尐 し、Bay X を除 く 2 地 点 は 11 月 以 降 検 出 限 界 以 下 となった。Bay X では季 節 を 問 わず多 くの大 腸 菌 群 数 が多 く検 出 されたが、夏 季 に最 も高 い値 を示 す点 は他 の 2 地 点 と同 様 であった。Bay X は近 くに下 水 処 理 場 があり、そこから流 入 する 水 の影 響 が大 きく、高 い大 腸 菌 群 数 が検 出 されたと考 えられる。Farm A も同 様 に近 くの河 川 の上 流 に下 水 処 理 場 が設 置 されているため、夏 季 には高 い値 を示 したが、Bay X と比 較 すると沿 岸 の人 口 も尐 ないことから流 入 する大 腸 菌 群 数 自 体 が尐 なかったと考 えられる。一 方 で大 腸 菌 群 数 の尐 なかった Farm B は沿 岸 の 人 口 が尐 なく、大 きな下 水 処 理 場 も近 くにないことが低 い汚 染 率 と関 連 している と考 えられた。以 上 のように、今 回 調 査 した 3 つの海 域 における大 腸 菌 群 の変 動 は周 辺 環 境 の違 い、下 水 処 理 場 の有 無 などを反 映 していると思 われた 。 マガキにおけるノロウイルス汚 染 の季 節 変 動 をみると 3 点 全 てにおいて汚 染 が見 られるのが 12 月 から 2 月 にかけての冬 季 であり、特 に大 腸 菌 群 数 の多 くな る夏 季 にはほとんどノロウイルスは検 出 されなかった Farm A、Bay X では 12 月 か ら翌 年 3 月 にかけて高 率 で陽 性 個 体 が検 出 された。また同 時 期 に行 った研 究 に お い て、 Bay X にお い ては マガキと 同 様 にム ラサ キイ ガイ M. galloprovincialis
からもノ ロウイルスが検 出 され、 同 海 域 に多 くのノロウイルスが存 在 すること、 海 水 に放 出 されたノロウイルスがマガキ以 外 の二 枚 貝 にも取 り込 まれていることが 示 されている (三 浦 , 2006)。これらのことから、3 つの調 査 海 域 において陽 性 率 の違 いはあるものの、マガキ体 内 のノロウイルスは冬 季 (12 月 から翌 年 3 月 ) に検 出 され、それ以 外 の時 期 はほとんど検 出 されないことが示 された。 以 上 のよう に糞 便 汚 染 の指 標 である大 腸 菌 群 数 は夏 季 多 く検 出 さ れるが冬 季 にはほと んど検 出 さ れなかった。その一 方 で、マガキのノ ロウイルス汚 染 は 冬 季 を 中 心 に起 きており 、夏 季 にはほと んど生 じ ていなかった。 すなわち、ノ ロウイ ルスが多 数 検 出 される冬 季 に限 ってみれば、大 腸 菌 群 数 は汚 染 の指 標 とはなら ないと考 えられ、従 来 の報 告 (Lee, 2000、Cook et al., 1986、Richerds, 1988) を 裏 付 ける結 果 となった。 大 腸 菌 群 数 、ノロウイルスともにヒト糞 便 由 来 であるにもかかわらず、海 水 中 や カキ体 内 での季 節 変 動 がまったく異 なるこ とについては不 明 な点 も多 いが、これ までの研 究 で以 下 のように考 えられている。 大 腸 菌 群 はヒトの糞 便 中 には多 量 に存 在 するがいったん外 界 に排 出 されると 比 較 的 短 時 間 に消 滅 する (『沿 岸 環 境 調 査 マニュアルⅡ(水 質 ・微 生 物 篇 )』 、 1990)。水 温 の高 い夏 季 は菌 の生 存 に適 しているので、外 界 において大 腸 菌 群 は長 期 にわたり多 量 に存 在 すると考 えられる。その一 方 で水 温 の低 い冬 季 は菌 の生 存 が抑 制 され、すぐに死 滅 していると考 えられる。 冬 季 にノロウイルス汚 染 が起 こる原 因 としては、冬 季 の環 境 水 中 にウイルスが 多 く存 在 するということが考 えられる。ノロウイルス食 中 毒 、感 染 症 が多 く見 られ る時 期 が冬 季 であり、患 者 から多 くのウイルスが海 水 に放 出 される。厚 生 労 働 省 の食 中 毒 ・食 品 監 視 関 連 情 報 によると、平 成 17 年 12 月 から 18 年 3 月 にかけて
の冬 季 においてノロウイルス食 中 毒 の患 者 数 は多 く、全 国 で 7000 人 を超 えてお り 、多 くのウイルスが糞 便 と と もに放 出 され、その一 部 が沿 岸 水 中 に流 入 し たと 考 えられる。したがって、本 実 験 の結 果 もノロウイルス感 染 患 者 からのノロウイル ス汚 染 が大 きく影 響 していると考 えられる。 その後 、ろ過 食 性 のマガキが餌 の摂 取 とともにウイルスを取 り込 み濃 縮 していると考 えられる。カキ自 体 の季 節 変 化 と ウイルス蓄 積 の関 係 性 については、 Burkhardt Ⅲ and Calci (2000) が低 水 温 時 にマガキにおける F. coliphage の蓄 積 が高 くなった実 験 結 果 に基 づき、低 水 温 時 によるウイルス生 き残 りと 、アメリ カガキのウイルス蓄 積 能 力 の季 節 的 変 化 を あげてい る。Lee (2000 ) は冬 季 の低 水 温 により カキの生 理 活 性 が低 下 し 、異 物 排 除 機 能 が十 分 に発 揮 されない可 能 性 も指 摘 している。 現 在 のマガキの食 品 として安 全 基 準 として、大 腸 菌 群 数 が用 いられている。マ ガキ養 殖 場 の基 準 は海 水 中 の大 腸 菌 群 数 1000 以 下 、生 食 用 カキは 70 以 下 で あることが定 められている (水 産 用 水 基 準 ; 2000)。宮 城 県 ではカキの出 荷 が始 まるのは 9 月 末 から 10 月 はじめであるが、その時 期 の大 腸 菌 群 数 は 1000 を下 回 っており 、出 荷 シ ーズンを 通 し て基 準 を満 たした。前 述 の通 り、本 研 究 では 大 腸 菌 群 数 の下 がる冬 季 にノロウイルス汚 染 が強 ま っているという結 果 が得 られた。 したがって、今 回 調 べた ノ ロウイルスと 大 腸 菌 群 数 は 、季 節 変 動 が大 きく異 なる ため、同 じ時 期 の両 者 の数 の間 には 相 関 は認 められなかった。すなわち冬 季 の 大 腸 菌 群 数 をノロウイルス汚 染 の指 標 として用 いることができないという結 果 とな った。これは海 外 での報 告 (Cook et al., 1986、Lee, 2000)とも一 致 していた。
しかし、今 回 調 べた 3 点 の結 果 を場 所 ごとに比 較 してみると、Bay X、Farm A は大 腸 菌 群 数 、ノロウイルス陽 性 率 の値 が大 きく出 る場 所 であり、Farm B は両 者 の値 がピーク時 でも他 の 2 点 と比 べて低 く出 る場 所 であった。以 上 のような場
所 ごとの比 較 の結 果 から、夏 季 に検 出 された大 腸 菌 群 数 の多 寡 と冬 季 における マガキのノロウイルス陽 性 率 の高 低 には、相 関 がある可 能 性 が考 えられた。
T
er
mina
te
r
eac
tio
n
mi
xt
ure
・
・
React
io
n
mi
xt
ure
V
o
lume
・
e
xt
ract
ed
RN
A
・
10
×
React
io
n
B
uf
fer
fo
r
RQ
1
RN
as
e
-F
ree
DNa
se
・
RQ
1
RN
as
e
-F
ree
DN
as
e
To
tal
C
o
mp
o
nen
ts
・
・
To
tal
React
io
n
mi
xt
ure
V
o
lume
・
e
xt
ract
ed
RN
A
・
10
×
React
io
n
B
uf
fer
fo
r
RQ
1
RN
as
e
F
ree
DNa
se
・
F
ree
DN
To
tal
C
o
mp
o
nen
ts
2μ
L
DNa
se
reac
tio
n
mi
xt
ure
表 1 D Na se 反応液の組成16
μ
L
2μ
L
20
μ
L
2μ
L
20
μ
L
22
μ
L
DNa
se
reac
tio
n
mi
xt
ure
Sto
p
So
lut
io
n
fo
r
RQ
1
DNa
se
T
er
mina
te
r
eac
tio
n
mi
xt
ure
・
・
React
io
n
mi
xt
ure
V
o
lume
・
e
xt
ract
ed
RN
A
・
10
×
React
io
n
B
uf
fer
fo
r
RQ
1
RN
as
e
-F
ree
DNa
se
・
RQ
1
RN
as
e
-F
ree
DN
as
e
To
tal
C
o
mp
o
nen
ts
・
・
To
tal
React
io
n
mi
xt
ure
V
o
lume
・
e
xt
ract
ed
RN
A
・
10
×
React
io
n
B
uf
fer
fo
r
RQ
1
RN
as
e
F
ree
DNa
se
・
F
ree
DN
To
tal
C
o
mp
o
nen
ts
2μ
L
DNa
se
reac
tio
n
mi
xt
ure
表 1 D Na se 反応液の組成16
μ
L
2μ
L
20
μ
L
2μ
L
20
μ
L
22
μ
L
DNa
se
reac
tio
n
mi
xt
ure
Sto
p
So
lut
io
n
fo
r
RQ
1
DNa
se
20
μ
L
React
io
n
mi
xt
ur
e
Vo
lume
Co
mp
o
nent
s
3
μ
L
2.
83
μ
L
1
μ
L
1
μ
L
0.
67
μ
L
1
μ
L
0.
5
μ
L
10
μ
L
・
5
×
RT
P
CR
B
uf
fer
・
RNa
se
D
N
as
e
fr
ee
w
at
er
・
0.
1
M
D
TT
・
dN
TP
(
10
mM
)
・
RNa
se
O
U
T
TM・
Supe
rScr
ip
t
TMⅡ
RT
・
R
ever
s
P
ri
mer
(
25
μ
M
)
・
ter
m
inat
e
react
io
n
m
ix
tur
e
Tot
al
RT
m
ix
tur
e
20
μ
L
React
io
n
mi
xt
ur
e
Vo
lume
Co
mp
o
nent
s
3
μ
L
2.
83
μ
L
1
μ
L
1
μ
L
0.
67
μ
L
1
μ
L
0.
5
μ
L
10
μ
L
・
5
×
RT
P
CR
B
uf
fer
・
RNa
se
D
N
as
e
fr
ee
w
at
er
・
0.
1
M
D
TT
・
dN
TP
(
10
mM
)
・
RNa
se
O
U
T
TM・
Supe
rScr
ip
t
TMⅡ
RT
・
R
ever
s
P
ri
mer
(
μ
)
・
Tot
al
RT
m
ix
tur
e
表 2 逆転写反応液の組成20
μ
L
React
io
n
mi
xt
ur
e
Vo
lume
Co
mp
o
nent
s
3
μ
L
2.
83
μ
L
1
μ
L
1
μ
L
0.
67
μ
L
1
μ
L
0.
5
μ
L
10
μ
L
・
5
×
RT
P
CR
B
uf
fer
・
RNa
se
D
N
as
e
fr
ee
w
at
er
・
0.
1
M
D
TT
・
dN
TP
(
10
mM
)
・
RNa
se
O
U
T
TM・
Supe
rScr
ip
t
TMⅡ
RT
・
R
ever
s
P
ri
mer
(
25
μ
M
)
・
ter
m
inat
e
react
io
n
m
ix
tur
e
Tot
al
RT
m
ix
tur
e
20
μ
L
React
io
n
mi
xt
ur
e
Vo
lume
Co
mp
o
nent
s
3
μ
L
2.
83
μ
L
1
μ
L
1
μ
L
0.
67
μ
L
1
μ
L
0.
5
μ
L
10
μ
L
・
5
×
RT
P
CR
B
uf
fer
・
RNa
se
D
N
as
e
fr
ee
w
at
er
・
0.
1
M
D
TT
・
dN
TP
(
10
mM
)
・
RNa
se
O
U
T
TM・
Supe
rScr
ip
t
TMⅡ
RT
・
R
ever
s
P
ri
mer
(
μ
)
・
Tot
al
RT
m
ix
tur
e
表 2 逆転写反応液の組成50 μ L R e act ion m ix tu re V ol um e Co mpo n e nts 3 3 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 5 μ L T ot al 1 st P CR m ix tu re ・ D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ R・ e ve rs e p rime r ・ RT m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ F o rw ar d p rim er ( 2 5 μ M ) ・ 1・ st P CR m ix tu re 3 6 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 2 μ L n e st ed -P CR mi xt u re 50 μ L 50 μ L R e act ion m ix tu re V ol um e Co mpo n e nts 3 3 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 5 μ L T ot al 1 st P CR m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ F o rw ar d p rim er (2 5 μ M ) ・ R e ve rs e p rime r (2 5 μ M ) ・ RT m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ ・ R e ve rs e p ri me r ( 2 5 μ M ) ・ 1 st P CR m ix tu re T ot al 3 6 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 2 μ L 50 μ L 表 3 1 st P C R と n es ted -P C R 反 応 液 の組 成 50 μ L R e act ion m ix tu re V ol um e Co mpo n e nts 3 3 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 5 μ L T ot al 1 st P CR m ix tu re ・ D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ R・ e ve rs e p rime r ・ RT m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ F o rw ar d p rim er ( 2 5 μ M ) ・ 1・ st P CR m ix tu re 3 6 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 2 μ L n e st ed -P CR mi xt u re 50 μ L 50 μ L R e act ion m ix tu re V ol um e Co mpo n e nts 3 3 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 5 μ L T ot al 1 st P CR m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x T aq b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ F o rw ar d p rim er (2 5 μ M ) ・ R e ve rs e p rime r (2 5 μ M ) ・ RT m ix tu re ・ R Na se D Na se fr e e w at er ・ 10 × E x b uf fe r ・ dN TP ( 2 .5 mM ) ・ E x T aq Hot Sta rt ・ ・ R e ve rs e p ri me r ( 2 5 μ M ) ・ 1 st P CR m ix tu re T ot al 3 6 .75 μ L 5 μ L 4 μ L 0 .2 5 μ L 1 μ L 1 μ L 2 μ L 50 μ L 表 3 1 st P C R と n es ted -P C R 反 応 液 の組 成
S eq ue n ce ( 5 P rim e r G e no gr oup G -G1 -S KF NEW G 1 R CO G 1F G1 -S K R G2 -S KF NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -S K R 1 st P CR G2 -S K R n e st ed PC R G -G -G -G -G -S eq ue n ce ( 5 ’ → 3’ ) P rim e r G e no gr oup G -Ⅰ G1 -NEW G 1 R CO G 1F G1 -G2 -NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -1 st P CR G2 -n e st ed PC R G -Ⅱ G -Ⅰ G -Ⅱ G -Ⅰ G -Ⅱ 表 4 ノ ロ ウ イ ルス 検 出用 RT -P C R プ ラ イマ ー の塩 基配 列 IU B c o de : R = A o r G ; B = C , G o r T ; Y = C o r T ; H = A , C o r T ; N = a Ny b as e S eq ue n ce ( 5 P rim e r G e no gr oup G -G1 -S KF NEW G 1 R CO G 1F G1 -S K R G2 -S KF NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -S K R 1 st P CR G2 -S K R n e st ed PC R G -G -G -G -G -S eq ue n ce ( 5 ’ → 3’ ) P rim e r G e no gr oup G -Ⅰ G1 -NEW G 1 R CO G 1F G1 -G2 -NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -1 st P CR G2 -n e st ed PC R G -Ⅱ G -Ⅰ G -Ⅱ G -Ⅰ G -Ⅱ 表 4 ノ ロ ウ イ ルス 検 出用 RT -P C R プ ラ イマ ー の塩 基配 列 S eq ue n ce ( 5 P rim e r G e no gr oup G -G1 -S KF NEW G 1 R CO G 1F G1 -S K R G2 -S KF NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -S K R 1 st P CR G2 -S K R n e st ed PC R G -G -G -G -G -S eq ue n ce ( 5 ’ → 3’ ) P rim e r G e no gr oup G -Ⅰ G1 -NEW G 1 R CO G 1F G1 -G2 -NEW G 2 R CO G 2F G2 -S K R C AR G A R B CN AT G T T YA G R T G G A TG A G CG Y TG G A TG CGN T TY C AT G A CN TG G G AG G G CG ATC G CA A G G CT TG T ACA AA AT T ATT T CT AA CC AACCC AR CC A TT R T ACA CC R CC NGC A TR HC CR T TR T AC AT C TG CC CG AA T T YG TA AAT G A A CA TCACC G G G G G T A TT A T T TG G St ep R T r eact io n G1 -1 st P CR G2 -n e st ed PC R G -Ⅱ G -Ⅰ G -Ⅱ G -Ⅰ G -Ⅱ 表 4 ノ ロ ウ イ ルス 検 出用 RT -P C R プ ラ イマ ー の塩 基配 列 IU B c o de : R = A o r G ; B = C , G o r T ; Y = C o r T ; H = A , C o r T ; N = a Ny b as e
F ar m A F ar m B Ba y X G1 G2 G 1 o r G 2 G 1 G2 G 1 o r G 2 G1 G2 G 1 o r G 2 2 0 05 A pr 1 0 ( 3 /2 9 ) 4 8 ( 1 4/ 29 ) 5 5 ( 1 6/ 29 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 1 7 ( 4 /2 3 ) 9 1 ( 2 1/ 23 ) 9 1 ( 2 1/ 23 ) M ay 1 0 ( 2 /2 0 ) 0 ( 0/ 20 ) 1 0 ( 2 /2 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 8 ) 1 3 ( 1 /8 ) 1 3 ( 1 /8 ) Ju n 0 ( 0/ 15 ) 1 3 ( 2 /1 5 ) 1 3 ( 2 /1 5 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) Ju l 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) A u g 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) Se p 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) O ct 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) N ov 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) D e c 3 ( 1/ 30 ) 4 7 ( 1 4/ 30 ) 4 7 ( 1 4/ 30 ) 0 ( 0/ 10 ) 2 0 ( 2 /1 0 ) 2 0 ( 2 /1 0 ) 2 0 ( 3 /1 5 ) 4 0 ( 6 /1 5 ) 4 7 ( 7 /1 5 ) 2 0 06 Jan 0 ( 0/ 30 ) 1 0 0 (3 0 /3 0 ) 1 0 0 (3 0 /3 0 ) 0 ( 0/ 13 ) 8 ( 1/ 13 ) 8 ( 1/ 13 ) 4 0 ( 6 /1 5 ) 8 7 ( 1 3/ 15 ) 8 7 ( 1 3/ 15 ) F e b 3 ( 1/ 26 ) 3 8 ( 1 0/ 26 ) 3 8 ( 1 0/ 26 ) 0 ( 0/ 15 ) 7 ( 1/ 15 ) 7 ( 1/ 15 ) 1 3 ( 2 /1 5 ) 1 0 0 (1 5 /1 5 ) 1 0 0 (1 5 /1 5 ) M ar 0 ( 0/ 30 ) 7 7 ( 2 3/ 30 ) 7 7 ( 2 3/ 30 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 3 0 ( 3 /1 0 ) 1 0 0 (1 0 /1 0 ) 1 0 0 (1 0 /1 0 ) 表 5 2005 年 4 月から 2006 年 3 月に おけ る 宮城県内 3 箇所( B ay X 、 F ar m A 、 F ar m B ) での ノ ロ ウイ ル ス陽性率( %) の季節変動 〔()内は(陽性個体/検査個体) 〕 F ar m A F ar m B Ba y X G1 G2 G 1 o r G 2 G 1 G2 G 1 o r G 2 G1 G2 G 1 o r G 2 2 0 05 A pr 1 0 ( 3 /2 9 ) 4 8 ( 1 4/ 29 ) 5 5 ( 1 6/ 29 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 1 7 ( 4 /2 3 ) 9 1 ( 2 1/ 23 ) 9 1 ( 2 1/ 23 ) M ay 1 0 ( 2 /2 0 ) 0 ( 0/ 20 ) 1 0 ( 2 /2 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 8 ) 1 3 ( 1 /8 ) 1 3 ( 1 /8 ) Ju n 0 ( 0/ 15 ) 1 3 ( 2 /1 5 ) 1 3 ( 2 /1 5 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) Ju l 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) A u g 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) Se p 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) O ct 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) N ov 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) D e c 3 ( 1/ 30 ) 4 7 ( 1 4/ 30 ) 4 7 ( 1 4/ 30 ) 0 ( 0/ 10 ) 2 0 ( 2 /1 0 ) 2 0 ( 2 /1 0 ) 2 0 ( 3 /1 5 ) 4 0 ( 6 /1 5 ) 4 7 ( 7 /1 5 ) 2 0 06 Jan 0 ( 0/ 30 ) 1 0 0 (3 0 /3 0 ) 1 0 0 (3 0 /3 0 ) 0 ( 0/ 13 ) 8 ( 1/ 13 ) 8 ( 1/ 13 ) 4 0 ( 6 /1 5 ) 8 7 ( 1 3/ 15 ) 8 7 ( 1 3/ 15 ) F e b 3 ( 1/ 26 ) 3 8 ( 1 0/ 26 ) 3 8 ( 1 0/ 26 ) 0 ( 0/ 15 ) 7 ( 1/ 15 ) 7 ( 1/ 15 ) 1 3 ( 2 /1 5 ) 1 0 0 (1 5 /1 5 ) 1 0 0 (1 5 /1 5 ) M ar 0 ( 0/ 30 ) 7 7 ( 2 3/ 30 ) 7 7 ( 2 3/ 30 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 3 0 ( 3 /1 0 ) 1 0 0 (1 0 /1 0 ) 1 0 0 (1 0 /1 0 ) F ar m A F ar m B Ba y X G1 G2 G 1 o r G 2 G 1 G2 G 1 o r G 2 G1 G2 G 1 o r G 2 2 0 05 A pr 1 0 ( 3 /2 9 ) 4 8 ( 1 4/ 29 ) 5 5 ( 1 6/ 29 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 1 7 ( 4 /2 3 ) 9 1 ( 2 1/ 23 ) 9 1 ( 2 1/ 23 ) M ay 1 0 ( 2 /2 0 ) 0 ( 0/ 20 ) 1 0 ( 2 /2 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 8 ) 1 3 ( 1 /8 ) 1 3 ( 1 /8 ) Ju n 0 ( 0/ 15 ) 1 3 ( 2 /1 5 ) 1 3 ( 2 /1 5 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) Ju l 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) A u g 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) Se p 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) O ct 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) N ov 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) D e c 3 ( 1/ 30 ) 4 7 ( 1 4/ 30 ) 4 7 ( 1 4/ 30 ) 0 ( 0/ 10 ) 2 0 ( 2 /1 0 ) 2 0 ( 2 /1 0 ) 2 0 ( 3 /1 5 ) 4 0 ( 6 /1 5 ) 4 7 ( 7 /1 5 ) 2 0 06 Jan 0 ( 0/ 30 ) 1 0 0 (3 0 /3 0 ) 1 0 0 (3 0 /3 0 ) 0 ( 0/ 13 ) 8 ( 1/ 13 ) 8 ( 1/ 13 ) 4 0 ( 6 /1 5 ) 8 7 ( 1 3/ 15 ) 8 7 ( 1 3/ 15 ) F e b 3 ( 1/ 26 ) 3 8 ( 1 0/ 26 ) 3 8 ( 1 0/ 26 ) 0 ( 0/ 15 ) 7 ( 1/ 15 ) 7 ( 1/ 15 ) 1 3 ( 2 /1 5 ) 1 0 0 (1 5 /1 5 ) 1 0 0 (1 5 /1 5 ) M ar 0 ( 0/ 30 ) 7 7 ( 2 3/ 30 ) 7 7 ( 2 3/ 30 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 3 0 ( 3 /1 0 ) 1 0 0 (1 0 /1 0 ) 1 0 0 (1 0 /1 0 ) F ar m A F ar m B Ba y X G1 G2 G 1 o r G 2 G 1 G2 G 1 o r G 2 G1 G2 G 1 o r G 2 2 0 05 A pr 1 0 ( 3 /2 9 ) 4 8 ( 1 4/ 29 ) 5 5 ( 1 6/ 29 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 1 7 ( 4 /2 3 ) 9 1 ( 2 1/ 23 ) 9 1 ( 2 1/ 23 ) M ay 1 0 ( 2 /2 0 ) 0 ( 0/ 20 ) 1 0 ( 2 /2 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 8 ) 1 3 ( 1 /8 ) 1 3 ( 1 /8 ) Ju n 0 ( 0/ 15 ) 1 3 ( 2 /1 5 ) 1 3 ( 2 /1 5 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) 0 ( 0/ 12 ) Ju l 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 1 0 ( 1 /1 0 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) 0 ( 0/ 13 ) A u g 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) Se p 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) 0 ( 0/ 9 ) O ct 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) N ov 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) 0 ( 0/ 10 ) D e c 3 ( 1/ 30 ) 4 7 ( 1 4/ 30 ) 4 7 ( 1 4/ 30 ) 0 ( 0/ 10 ) 2 0 ( 2 /1 0 ) 2 0 ( 2 /1 0 ) 2 0 ( 3 /1 5 ) 4 0 ( 6 /1 5 ) 4 7 ( 7 /1 5 ) 2 0 06 Jan 0 ( 0/ 30 ) 1 0 0 (3 0 /3 0 ) 1 0 0 (3 0 /3 0 ) 0 ( 0/ 13 ) 8 ( 1/ 13 ) 8 ( 1/ 13 ) 4 0 ( 6 /1 5 ) 8 7 ( 1 3/ 15 ) 8 7 ( 1 3/ 15 ) F e b 3 ( 1/ 26 ) 3 8 ( 1 0/ 26 ) 3 8 ( 1 0/ 26 ) 0 ( 0/ 15 ) 7 ( 1/ 15 ) 7 ( 1/ 15 ) 1 3 ( 2 /1 5 ) 1 0 0 (1 5 /1 5 ) 1 0 0 (1 5 /1 5 ) M ar 0 ( 0/ 30 ) 7 7 ( 2 3/ 30 ) 7 7 ( 2 3/ 30 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 0 ( 0/ 15 ) 3 0 ( 3 /1 0 ) 1 0 0 (1 0 /1 0 ) 1 0 0 (1 0 /1 0 ) 表 5 2005 年 4 月から 2006 年 3 月に おけ る 宮城県内 3 箇所( B ay X 、 F ar m A 、 F ar m B ) での ノ ロ ウイ ル ス陽性率( %) の季節変動 〔()内は(陽性個体/検査個体) 〕
1700 1 1 Mar 490 1 480 Feb 1 1 1 Jan 2006 790 1 1 Dec 80 1 20 Nov 790 20 490 Oct 1300 20 1100 Sep 2400 50 490 Aug 5430 490 16000 Jul 3500 70 790 Jun 330 1 170 May 50 1 50 Apr 2005 Bay X Farm B Farm A 表6 2005 年 4 月から 2006 年 3 月における宮城県内 3 箇所 (Bay X、Farm A、Farm B) での大腸菌群数の季節変動
1700 1 1 Mar 490 1 480 Feb 1 1 1 Jan 2006 790 1 1 Dec 80 1 20 Nov 790 20 490 Oct 1300 20 1100 Sep 2400 50 490 Aug 5430 490 16000 Jul 3500 70 790 Jun 330 1 170 May 50 1 50 Apr 2005 Bay X Farm B Farm A 表6 2005 年 4 月から 2006 年 3 月における宮城県内 3 箇所 (Bay X、Farm A、Farm B) での大腸菌群数の季節変動
2006 2005 salinity temperature salinity temperature salinity temperature Bay X Farm B Farm A
表7 2005 年 4 月から 2006 年 3 月における宮城県内 3 箇所 (Bay X、Farm A、Farm B) の海水の水温 (℃)、塩分 (‰) の季節変動 13.5 20 26 24.1 26 24 15.5 10.5 3.5 1 4 5.5 30.5 28.6 27.5 26.8 25.8 28.1 34 35 32 35 35 34 9.2 11.5 16.8 19.5 23 20 19.5 15.3 10.5 8.7 6 5 28 33.4 31.3 30.8 32.2 32.3 35 36 35 36 36 36 12 17.5 24 20.5 25 24 19 15.5 8 3 3 5 30.5 30.1 29.5 21.1 27.5 29.9 33 34 34 34 33 25 Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar 2006 2005 salinity temperature salinity temperature salinity temperature Bay X Farm B Farm A
表7 2005 年 4 月から 2006 年 3 月における宮城県内 3 箇所 (Bay X、Farm A、Farm B) の海水の水温 (℃)、塩分 (‰) の季節変動 13.5 20 26 24.1 26 24 15.5 10.5 3.5 1 4 5.5 30.5 28.6 27.5 26.8 25.8 28.1 34 35 32 35 35 34 9.2 11.5 16.8 19.5 23 20 19.5 15.3 10.5 8.7 6 5 28 33.4 31.3 30.8 32.2 32.3 35 36 35 36 36 36 12 17.5 24 20.5 25 24 19 15.5 8 3 3 5 30.5 30.1 29.5 21.1 27.5 29.9 33 34 34 34 33 25 Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
第 3 章 大 腸 菌 群 数 をもとにしたマガキのノロウイルス汚 染 の予 測 方 法 の検 討 第 2 章 において、夏 季 における大 腸 菌 群 数 と冬 季 マガキのノロウイルス汚 染 との 間 には関 係 のあることが示 唆 された。そこで本 章 では、第 2 章 で調 べた 3 箇 所 の 他 に新 たに宮 城 県 内 のマガキ養 殖 場 5 箇 所 を調 査 海 域 と定 め、海 水 の大 腸 菌 群 数 とマガキのノ ロウイルス陽 性 率 の測 定 を行 って両 者 の関 係 性 を 明 らかにす ることを目 的 とした。また、第 2 章 ではノロウイルス汚 染 の程 度 を陽 性 個 体 (陽 性 率 ) で評 価 したが、本 章 ではノロウイルス遺 伝 子 の定 量 評 価 を目 的 として、リ アルタイム RT-PCR を行 い、大 腸 菌 群 数 とノロウイルス遺 伝 子 数 の量 的 関 係 を 明 らかにすることを試 みた。 第 1 節 夏 季 における海 水 の大 腸 菌 群 数 と冬 季 マガキのノロウイルス陽 性 率 と の関 係 【材 料 と方 法 】 マガキと海 水 マガキ、および養 殖 海 域 付 近 の海 水 のサンプリング地 点 として前 述 3 箇 所 と は別 の 5 箇 所 (Farm C、Farm D、Farm E、Farm F、Farm G) の養 殖 場 を設 定 し た。今 回 の 5 箇 所 には、同 一 湾 内 にある Farm C、D と Farm F、G (C,D と F,G は 異 なる湾 である) を設 定 し、近 接 する海 域 において大 腸 菌 群 数 やノロウイルスの 動 態 が異 なるかどうか調 べることにした。採 取 したマガキの平 均 殻 高 (mm)、殻 長 (mm)、殻 幅 (mm)、消 化 盲 嚢 重 量 (g) はそれぞれ、Farm C: 104.8±1.3、 49.3±0.6、27.6±0.5、0.9±0.0、Farm D: 136.4±2.0、63.4±0.8、32.6±0.5、1.6 ±0.1、Farm E: 116.4±1.0、61.0±0.8、36.5±0.6、1.1±0.0、Farm F: 129.2±1.8、
53.8± 0.9 、32.6 ±0.7、0.9 ±0.0 、Farm G: 123.1 ±1.7 、 60.9±0.7 34.0± 0.6 、 1.0±0.0 であった。Bay X、Farm A、Farm B に関 して 2005 年 12 月 から 2006 年 2 月 のマガキのサイズは Bay X: 62.4、39.1、24.6、0.4、Farm A: 116.7、50.9、 31.7、1.4、Farm B: 119.1、59.2、32.3、2.4 であった。Farm C、D、E、F、G では 2006 年 12 月 から 2007 年 3 月 にかけて各 月 1 回 30 個 体 ずつのマガキを採 取 し た。 マガキ組 織 からの ノ ロウイルス検 出 方 法 お よび海 水 中 の大 腸 菌 群 数 の測 定 方 法 第 2 章 と同 様 の方 法 で行 った。 なお、大 腸 菌 群 数 とノロウイルス陽 性 率 の相 関 関 係 は t-test により有 意 性 の判 定 を行 った。
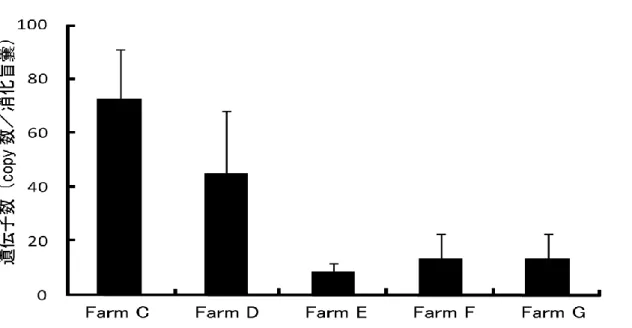
【結 果 】 5 箇 所 の調 査 地 点 において 2006 年 12 月 から 2007 年 3 月 にかけて冬 季 マガキ のノロウイルス陽 性 率 を測 定 した結 果 、全 ての海 域 においてノロウイルス G1、G2 両 方 が検 出 された (表 8)。月 別 にみると、12 月 と 1 月 にかけて全 ての海 域 にお いて陽 性 率 が高 く、特 に Farm C、D においては 1 月 に G1、G2 ともに 50 % 前 後 の陽 性 率 となった。Farm E は Farm C、D と比 較 するとピーク時 の陽 性 率 がやや 低 いが、今 回 調 査 した 4 ヶ月 の間 、常 に陽 性 率 が 20 % 以 上 となっておりノロウイ ルス汚 染 が長 く続 いているという特 徴 があった。Farm F、G は他 の 3 点 と比 べると 汚 染 度 は低 く、12 月 が最 も陽 性 率 が高 く出 た後 は低 い陽 性 率 で推 移 した。また、 Farm C、D と Farm F、G はそれぞれ同 一 湾 内 に設 定 した 2 点 であり陽 性 率 の高 さ、その変 化 の挙 動 はほぼ同 様 であった。
2005 年 12 月 から 2006 年 2 月 にかけての Bay X、Farm A、B のノロウイルス 陽 性 率 のデータと併 せて計 8 箇 所 における冬 季 (12 月 、1 月 、2 月 ) マガキのノ ロウイルス陽 性 率 とその平 均 値 を表 10 に示 した。平 均 値 の最 も高 い値 は Bay X の 78 % で、最 も低 い値 は Farm B の 11 % あった。 マガキのノロウイルス陽 性 率 を測 定 した 5 箇 所 において、2006 年 7 月 から 10 月 にかけて海 水 の大 腸 菌 群 数 を測 定 した(表 9)。今 回 の 5 点 での測 定 では 7 月 、 8 月 に高 い値 が得 られた。最 も値 が高 かったのは Farm C であり、8 月 を中 心 に他 の 4 点 よりどの月 も高 い値 が出 ていた。一 方 、同 一 湾 に存 在 する Farm D は Farm C 比 べると値 は小 さかった。Farm C、D とは異 なる湾 である Farm E、F、G は先 ほ どの 2 点 と比 べて値 は小 さく、第 2 章 で示 した Farm B の値 と同 等 であった。ノロ ウイルス陽 性 率 と同 様 に 2005 年 の調 査 点 と併 せて計 8 箇 所 において、夏 季 (7
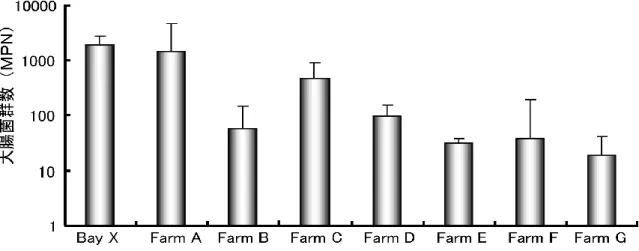
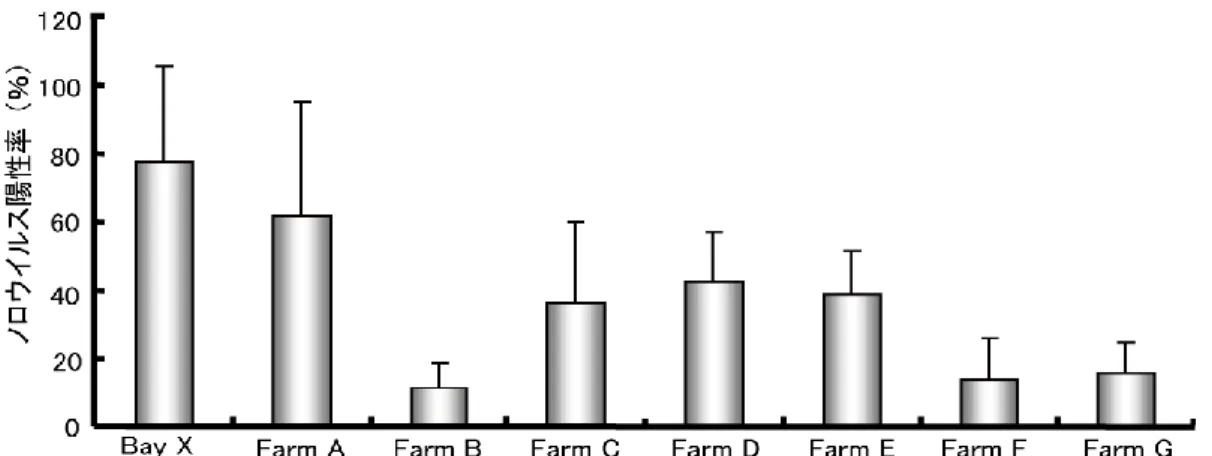
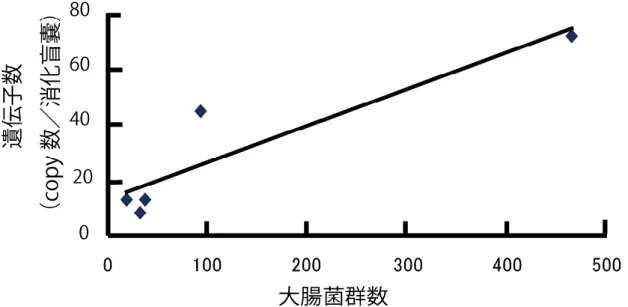
月 ~10 月 ) の海 水 中 の大 腸 菌 群 数 の値 (MPN) とその平 均 値 を示 した。平 均 値 が最 も高 い地 点 は Bay X の 1912.7 であり、最 も低 い値 は Farm G の 19 であっ た。 表 9、10 に示 した結 果 からマガキのノロウイルス陽 性 率 の平 均 値 と海 水 の大 腸 菌 群 数 の平 均 値 の相 関 関 係 を図 5 に示 した。結 果 は、相 関 係 数 R=0.884 (p < 0.01 N=8) であり両 者 には相 関 が認 められた。
0 (0/29) 7 (2/30) 17 (5/30) 24 (7/29) G1or G2 0 (0/29) 3 (1/30) 13 (4/30) 14 (4/29) G2 0 (0/29) 3 (1/30) 3 (1/30) 14 (4/29) G1 Farm G 3 (1/30) 10 (3/30) 3 (1/29) 28 (8/29) G1or G2 3 (1/30) 3 (1/30) 0 (0/29) 21 (6/29) G2 0 (0/30) 7 (2/30) 3 (1/29) 10 (3/29) G1 Farm F 30 (9/30) 43 (13/30) 48 (14/29) 24 (7/29) G1or G2 10 (3/30) 23 (7/30) 28 (8/29) 17 (5/29) G2 20 (6/30) 37 (11/30) 28 (8/29) 17 (5/29) G1 Farm E N.D 26 (7/27) 53 (16/30) 48 (14/29) G1or G2 N.D 15 (4/27) 33 (10/30) 38 (11/29) G2 N.D 19 (5/27) 50 (15/30) 24 (7/29) G1 Farm D 10 (3/30) 10 (3/30) 57 (16/28) 41 (12/26) G1or G2 0 (0/30) 7 (2/30) 39 (11/28) 31 (9/26) G2 10 (3/30) 7 (2/30) 57 (16/28) 21 (6/26) G1 Farm C Mar Feb Jan Dec 表8 2006年12月から2007年3月における宮城県内5箇所の養殖海域(Farm C~G)での マガキのノロウイルス陽性率 (%) 〔()内は(陽性個体/検査個体)〕 0 (0/29) 7 (2/30) 17 (5/30) 24 (7/29) G1or G2 0 (0/29) 3 (1/30) 13 (4/30) 14 (4/29) G2 0 (0/29) 3 (1/30) 3 (1/30) 14 (4/29) G1 Farm G 3 (1/30) 10 (3/30) 3 (1/29) 28 (8/29) G1or G2 3 (1/30) 3 (1/30) 0 (0/29) 21 (6/29) G2 0 (0/30) 7 (2/30) 3 (1/29) 10 (3/29) G1 Farm F 30 (9/30) 43 (13/30) 48 (14/29) 24 (7/29) G1or G2 10 (3/30) 23 (7/30) 28 (8/29) 17 (5/29) G2 20 (6/30) 37 (11/30) 28 (8/29) 17 (5/29) G1 Farm E N.D 26 (7/27) 53 (16/30) 48 (14/29) G1or G2 N.D 15 (4/27) 33 (10/30) 38 (11/29) G2 N.D 19 (5/27) 50 (15/30) 24 (7/29) G1 Farm D 10 (3/30) 10 (3/30) 57 (16/28) 41 (12/26) G1or G2 0 (0/30) 7 (2/30) 39 (11/28) 31 (9/26) G2 10 (3/30) 7 (2/30) 57 (16/28) 21 (6/26) G1 Farm C Mar Feb Jan Dec 表8 2006年12月から2007年3月における宮城県内5箇所の養殖海域(Farm C~G)での マガキのノロウイルス陽性率 (%) 〔()内は(陽性個体/検査個体)〕
19 37.9 31.6 93.4 464.2 56 1433.8 1912.7 geometrical mean 50 20 50 20 50 20 490 790 Oct 20 1 20 50 490 20 1100 1300 Sep 1 130 20 230 2400 50 490 2400 Aug 130 790 50 330 790 490 16000 5430 Jul Farm G Farm F Farm E Farm D Farm C Farm B Farm A Bay X 19 37.9 31.6 93.4 464.2 56 1433.8 1912.7 geometrical mean 50 20 50 20 50 20 490 790 Oct 20 1 20 50 490 20 1100 1300 Sep 1 130 20 230 2400 50 490 2400 Aug 130 790 50 330 790 490 16000 5430 Jul Farm G Farm F Farm E Farm D Farm C Farm B Farm A Bay X
表9 宮城県内 8 箇所 (Bay X、Farm A~G) における夏季(7月~10月)における海水の 大腸菌群数 (MPN) の値とその平均値 (相乗平均) 19 37.9 31.6 93.4 464.2 56 1433.8 1912.7 geometrical mean 50 20 50 20 50 20 490 790 Oct 20 1 20 50 490 20 1100 1300 Sep 1 130 20 230 2400 50 490 2400 Aug 130 790 50 330 790 490 16000 5430 Jul Farm G Farm F Farm E Farm D Farm C Farm B Farm A Bay X 19 37.9 31.6 93.4 464.2 56 1433.8 1912.7 geometrical mean 50 20 50 20 50 20 490 790 Oct 20 1 20 50 490 20 1100 1300 Sep 1 130 20 230 2400 50 490 2400 Aug 130 790 50 330 790 490 16000 5430 Jul Farm G Farm F Farm E Farm D Farm C Farm B Farm A Bay X
表9 宮城県内 8 箇所 (Bay X、Farm A~G) における夏季(7月~10月)における海水の 大腸菌群数 (MPN) の値とその平均値 (相乗平均)
B ay X Fa rm A Fa rm B Fa rm C Fa rm D Fa rm E Fa rm F Fa rm G Dec 47 ( 7/15) 47 ( 14/30) 20 ( 2/10) 41 ( 12/26) 48 ( 14/29) 24 ( 7/29) 28 ( 8/29) 24 ( 7/29) Jan 87 ( 13/15) 100 ( 30/30) 8 ( 1/13) 57 ( 16/28) 53 ( 16/30) 48 ( 14/29) 3 ( 1/29) 17 ( 5/30) Feb 100 ( 15/15) 38 ( 10/26) 7 ( 1/15) 10 ( 3/30) 26 ( 7/27) 43 ( 13/30) 10 ( 3/30) 7 ( 2/30) av era ge va lue 78 62 11 36 43 39 14 16 表 10 宮城県内 8 箇所 ( B ay X 、 F ar m A ~ G ) に おけ る 冬季 (12 月~ 2 月)に おける マガ キの ノ ロ ウ イ ルス陽 性率 (% ) と そ の 平均値 〔 ()内は (陽性 個体 /検 査個 体) 〕
図 3 宮城県内8箇所 (Bay X、Farm A~G) の海域における、夏季(7月~10月)の海水の大腸菌群数の平均値 図 3 宮城県内8箇所 (Bay X、Farm A~G) の海域における、夏季(7月~10月)の海水の大腸菌群数の平均値
図 4 宮城県内 8 箇所 (Bay X、Farm A~G) の海域における、冬季 (12月~2月) マガキのノロウイルス陽性率の平均値 図 4 宮城県内 8 箇所 (Bay X、Farm A~G) の海域における、冬季 (12月~2月) マガキのノロウイルス陽性率の平均値
図 5 宮城県内 8 箇所 (Bay X、Farm A~G) の海域における冬季マガキのノロウイルス陽性率の
平均値と、夏季の海水の大腸菌群数の平均値の相関関係 (R= 0.884p<0.01N = 8)
図 5 宮城県内 8 箇所 (Bay X、Farm A~G) の海域における冬季マガキのノロウイルス陽性率の