ミツバチ科学15(3):107-114 HoneybeeScience(1994)
マル- ナバ チの利 用
- その現状
と将来-昆虫の もつ様 々な機能が注 目され,その利用 の可能性が論議 されている. その中において, 送粉昆虫の受粉能力 は農作物 の生産性 に多大 な 影響 を及ぼす重要 な行動形質 として位置づけ ら れている (Free,1993). 農作物の花粉媒介用 に実用化 されている送粉昆虫 には, セイヨウ ミ ソバチ (Apismelliferla)を初め とす る ミツパ テ類,マメコパテ (Osmiacornifrons)の属す る ツ ツ- ナ パ テ頬, そ して シマ ハ ナ ア ブ (Eristaliscerealis)などが知 られ,大 きな経済 効果を生 み出 している.真社会性昆虫であ り大 きなコロニーを基本単位 と して生活 しているセ イヨウ ミツバチは,飼育管理技術 も確立 されて お り,従来 より,養蜂業の一・環 として様 々な農 作 物 の 花 粉 媒 介 に 利 用 さ れ て い る (Jay, 1986). 日本で も, - ウスイチゴの生産 にはな くて はな らな い存在 とな って いる (Matsuka alldSakai,1989). しか しなが ら, ミツバチは トマ ト(Lyc
opersiconesculentum)のよ うに花 蜜を分泌 しないナス科 (Solanaceae)植物 には 訪花せず, また紫外線 を除去するフィルムで被 覆 された- ウス内では十分 な受粉効果が期待で きないなどの使用上 の限界 も指摘 されている. それ らの施設園芸上 の問題点 を解決 したのが, ミツバチとは異 なる生活様式 を もつマル-ナバ チ頬である. ヨーロッパでは既 にダーウィンの 時代か ら, マル-ナバチの もっ高 い花粉媒介能 力が注 目され, 1800年代後半 にイギ リスか ら ニュージーラン ドに船で運 ばれた女王蜂の子孫 たちが レッ ドクローバーな どの牧草 の種子生産 に大活躍 してお り,同国の主要 な経済基盤 とし ての酪農 を支える縁 の下 の力持 ちとして きわめ て大切 にされ,人々の生活 において も身近 な も小野 正人
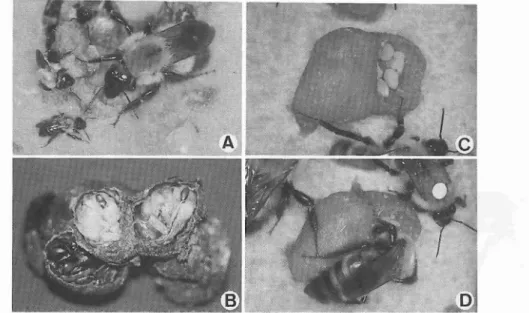
のとして位置づけ られている.筆者 は日本産 マ ル-ナバチを室内周年飼育 し,受粉能力 に関す る研究 を進めているが,本稿ではマルハナバチ 利用技術の現状 と問題点 などについて概略を述 べたい. マ ルハ ナバ チの分 類 と生 態 マル-ナパテ (英名;bumblebee)は,昆虫 網, ハ チ 目 (Hymenoptera), ミ ツ バ チ 科 (Apidae),マル-ナパテ属 (Bombus)に含 ま れる真社会性ハチ類である.北半球の温帯 と亜 寒帯地域 を中心 に分布す るが.南米 の熱帯圏に も生息 している.世界中で約300種が知 られ る (Pr9S-JonesandCorbet,1991).性質 は一般 的に温和であるが,熱帯地域 に分布す る種 には きわめて攻撃的な ものがある (坂上, 1970). 日本 に は,5亜 属 14種 が知 られ る (平 嶋, 1989;伊藤,1993).ヨーロッパか らの導入種, セイ ヨウオオマルハナバチ (別名 ツチマルハナ バチ,Bombusterrestris;図1) はオオマル-ナバチ亜属 に入 り, 日本 に も分類学的に近縁 な 種が3種 いる.特 にオオマルハナバチBombus (Bombus)hyPocritaとク ロマル- ナバ チB. 図 1 莞 LをT3 ; .// 1; セイヨウオオマルハナバチ 上段左:女 王蜂,右;雄蜂,下段;様々な大きさの働き蜂108 (B.)ignitusの2種 は, コロニーサイズが大 き い上 に飼いやす く,在来種の中では実用可能な 花粉媒介者 として最有力候補 と考えている.ま た,長舌型 と短舌型 という分け方をす る場合 も あり,前者の好例 としてナガマルハナバチ亜属 や トラマル-ナバチ亜属,後者ではコマルハナ バチ亜属やオオマル-ナパテ亜属が挙げ られる (図 2). トラマル-ナバチは蜜腺が花の奥深い ところにあ るツ リフネソウなどによ く訪花す る.また,早朝 にカボチ ャの花に多数の働 き蜂 が吸蜜に来ているのをみかけることがある. 温帯産マルハナバチの生活史は,少数 の例外 はあるが原則的に 1年性である.春に越冬を終 えた 1頭の女王蜂により営巣が開始 される.女 王蜂 はノネズ ミの廃巣や地下 の間隙 に入 り込 み,体か ら分泌 したロウと花粉で巣を造 り,働 き蜂 になる受精卵を数個産んで蓋をする.それ と前後 して小 さな蜜 ポッ トをそのす ぐ近 くに造 りそこに蜜を貯蔵す る.女王蜂 はあたか も鳥が 抱卵するように上か ら育児室に覆い被 さり保温 す る (図
3
-
D). 卵か ら照 った幼虫 は育児室内 で集団養育 され,やがて繭を紡 ぎ蛸になる.女 王蜂 は幼虫の生育にあわせて育児室のロウの壁 を自在に広げてい く. この育児様式には,ハチ 目の中で もこのグループに特異的に進化 したも のである.産卵か ら約1か月後に羽化す る最初 の働 き蜂 は小型であるが,今まで単独で営巣活 動 を行 って きた母親 の女王蜂 を助 け,巣 の拡 張,餌集め,巣の防衛などを担当する. このた め女王蜂 は産卵 とそれに関連する仕事 に専念で きるようになる.働 き蜂達 はさらに多 くの働 き 蜂を育て,巣は急激に大 きくなっていく. この 時,育児室内の幼虫 に花粉 を給餌す るための "ポケ ット(図4
-
C,D)
''を造 るポケ ッ トメーカ ー (トラマルハナバチなど) とそのようなもの を造 らないノンポケ ッ トメーカー (オオマルハ ナバチなど) の特徴が現れる.8
月の終わ りか ら9月頃には,雄蜂 と次の年 に女王蜂 となる新 女王蜂が多数生産 される. それと前後 して創設 女王蜂は死亡 し, コロニーは衰退の一途をたど る.新女王蜂 は巣を離れ野外で別の巣か ら巣立 った雄蜂 と交尾す る,交尾 を終えると土中や朽 ち木にもぐり込み長 い休眠期を経て,翌年の春 に 1頭で巣を創設す る. ミツパテとの違 い ハチ ミツ, ローヤルゼ リーそ して最近注E]を 集めているプロポ リスなど様々な生産物を提供 して くれる最大級の益虫である ミツバチは,ま た送粉昆虫 と して も不動 の実績 を誇 って いる (佐々木,1994).オール ラウンドプ レーヤーと もいえる ミツパテと比較 してマル-ナパテはど のような特徴をもっているのだろうか. この点 については, 松浦 (1993)の明快 な解説があ る.両種の主な相違点を表 1に示 した.応用的 な観点か らみたマルハナバチの最大の特徴 は, 振動採餌行動 (buzzforaging)である.彼等 は,病に噛みつ きそれを抱え込むように してぶ ら下が り,胸部の筋肉を振動 させ花粉を懐の体 毛 で受 け止 めて集 め ることがで きるのである (Michener,1962;Buchmann,1978;King, 1993).そ して,後脚の花粉かごに大 きな団子 状 にまとめて巣に運ぶ. トマ トの授粉 には, こ の振動採餌行動が不可欠なのである. また,秩 い空間内への適応性 もミツバチより高 い.他に も紫外線除去 フィルムの影響を受 けに くい,採 餌範既が狭 い, コロニーサイズが小 さい,攻撃 性が弱いなどの諸点 は,施設BiI芸作物の送粉者 として使用する上での大 きなメ リットといえよう.
セイ ヨウオオマルハナバチの 導入 と問題点 ヨーロッパにおけるマル-ナバチの商品化の 端 緒 は, 1987年 に ベ ル ギ ー の Roland de Jonghe博士が, セイヨウオオマル-ナバチの 温室 トマ トへの受粉 に卓越 した効果を認め, こ れを大量増殖 したBiobest社 を設立 し販売 を 開始 したことにある.その後,オランダ,イギ リス, フランスなどヨーロッパ各国を中心 にニ ュージーラン ドを含めて生産販売を行 う会社 は 十数社 に増えた(Donovan,1993).特に,北 ヨ ーロッパにおいては トマ ト栽培農家の90%(約 3,500ha)で使用 されているという (和田 ・栗109 財叩 藩 嘉 ㌃W^vB#竹 山召 通 じて輸入 された (岩崎,私信)・従来・人力で 撃 ホルモ ン剤(4-CPA:トマ トト ンなど)を処理 図2 長舌型の トラマルハナバチ (左)と短舌型のコ マル-ナバチ (右) 原, 1992). 日本へはヨー ロッパの施設園芸技 術を視察 した グループの目に最初 にとまって紹 介 され た. その後 1991年 12月 4日に Bi
o-b
e
s
t
社製 (商品名-ニー トー ン) の1
6
群が東 海物産株式会社 により, また引 き続 いて翌年1 月にはオ ランダのKo
ppe
r
t
社製 (商品名 ナチ ュポール;図5) の数群が株式会社 トーメ ンを す ることで単為結果 させ るとい う トマ トの生産 様式が 日本 の主流であ ったが, ヨーロッパか ら 導入 されたマルハナバチ利用技術 は,労働力の 大幅な削減 と自然交配 による果実の品質向上 を もた らしている (池田・忠内,1992;和田・栗原, 1992). トマ ト側か らの検討事項 もあるがその 技術 は急速 に浸透 しつつあ り,導入後3年 目で 日本 の全 トマ トハ ウスの3- 5%に普及 して い るともいわれ,現在 までの総輸入量 は15,000 群 を こえて い ると推定 されて い る (和 田,私 信).その後,日本では数社 が海外のマル-ナパ テ生産会社 と販売契約,あ るいは技術提携 を結 び様 々なルー トか ら輸入が開始 されている. 農産物の生産向上 という見地 に立てば, セイ ヨウオオマル-ナバチとその利用技術の導入 は 花粉媒介 とい う昆虫 と植物 の共生 システムの一 形態を人間の造 った枠組み としての農業 に取 り 入れ るという点 で,重要 な意味を もつ ものと考 え られ る. しか しなが ら, もし利用中のコロニ ーが大 き く発達 して生殖 個体 を生産 す るに至 り,新女王蜂が野外 に逃 げ出 して帰化す るよ う なことが起 きた場合の生態系 に対す る影響 を も 図3 コマル-ナバチの営巣状況A.ショ糖水を飲む女王蜂,B.営巣初期のコロニーの状態,
C.女王蜂の産卵 (複数の卵をまとめて産む),D.育児室を温める女王蜂図4 トラマル-ナバチの巣の内部 A.女王蜂と小型の働き蜂,ち.繭の中の虫
乱
C.育児室に造られた "ポケ ット"(花粉だんごが入っている),D.花粉だんごを内部に押し込む働き蜂 熟慮する必要があろう. セイヨウオオマルハナ バチはマルハナバチの中で も生活力が強 く,他 種の巣に入 り込み女王蜂 を殺 して巣をの っとっ て しま うとい う習性 もあ り (Sladen,1912; Velthuisand Cobb,1991), 日本在来のマル ハナバチに様々な形で影響 を及ぼす ことは明白 であろう. さらに, これまで形成 されて きた日 本産の-ナバチと花 との間のパー トナーシップ の崩壊 も懸念 され,動物相だけでなく植物相 に も与 え る影響 が危倶 され るのである (加藤, 1993).参考 までにセイ ヨウオオマル-ナバチ が日本に帰化するとした ら次のような道程を通 ることになるであろう. ①ハ ウス内に長期的に放置 された巣箱の中で コロニーが発達 し,新女王蜂 と雄蜂の生産が行 われる. (塾それ らの新女王蜂 と雄蜂がハ ウスの外へ逃 げ出 し交尾す る (- ウス内部で も交尾が起 こり うる). (卦新女王蜂 は交尾後 に土中で越冬す る. ④春に営巣場所を探 して巣づ くりを開始 (他 種巣をのっとる場合 もある). 6)恵 まれた環境では巣 を大 きく発達 させ,釈 女王蜂 と雄蜂を生産 し,世代が繰 り返 される. ハ ウス内の花粉量が少 ない場合は,新女王蜂 が生産 される前にコロニーが衰退 して しまうこ とも多 く,現状の使 い方が守 られていれば帰化 までには幾っかのハー ドルがある.ただ し,交 尾済みの女王蜂の大量輸入 は,それを野外に逃 が して しまった時が即④ の段階であり,帰化す る可能性が格段に高 まるので避 けなければな ら ない. また,導入種の天敵が日本 にいるか確認 されてお らず,野外での使用 は絶対 に避 けなけ ればな らない. 日本の養蜂業を支えているセイ ヨウ ミツバチも導入種であるが,オオスズメバ チと ミツバチヘギイタダニという強力な天敵が 野生化を許 していない. この2種の天敵のいな い小笠原諸島では,セイヨウ ミツバチの定着が 在来の-ナパテ相 はもとより植物相 にも影響を 及ぼ しているという (Kato,1992).オース ト ラ リアで もセイヨウ ミツバチが,土着の植物の 種 子 生産 に悪 い影響 を及 ぼ して い る ら しい (Paton,1993).さらに筆者 らの予備的研究で は,同 じ亜属内において新女王蜂の分泌す る性 フェロモ ンに種間交差活性があることが確かめ られてお り,例えば日本産のクロマル-ナバチ や コマルハナバチの新女王蜂に導入種の雄蜂が 交尾行動を起 こして しまうことも観察 されてい る.最終 的 な交尾 の成立 は確認 されていない が,交雑が起 きて しまう可能性 もないとはいえない. 導入種 が帰化 すれば様 々な障害 を もた らす こ とは必至 であろ う.具体 的 に対応策 と して考 え られ るの は有力在 来種 の利 用開発 であ る.外来 種 の導入が法的 に規制 されて い るアメ リカ合衆 国 で は在来種 の利用 が進 め られ, カナダで も同 様 の観 点 か ら試 験 が実施 され て い る (Kevan eta1.,1991). マ ル ハ ナ バ チ の 室 内 飼 育 法 春 に野外 を飛 び回 って い る女王蜂 を捕 らえて 室 内 に持 ち込 み飼育箱 内で営巣 させ る試 み は, 欧米, カナ ダの研究者 によ って熱心 に取 り組 ま れて きた (Horber,1961;PlowrightandJay, 1966).Heinrich(1979)は室 内飼育 の方法 を 紹介 して い る. 1980年 に入 ってオ ラ ンダと ド イツの研究者 によ って公表 されて い るセイ ヨウ オオマル- ナバ チの室 内飼育 法 も比較 しなが ら 述 べ る. まず春 に花 で吸蜜 して い る女王蜂 を捕 ilHl らえて飼育箱 に入 れ る.様 々な飼育箱 が考案 さ れて い るが,筆者 らは最初 150(縦)×80(横)× 65(高 さ)mmの木箱 を用 い,コロニーの成長 に ともな って大型 の箱 に移 す よ うに して いる.女 王 蜂 を 巣 箱 に入 れ る際,van Honk etal.
(1980)とDuchateauandVelthuis(1988) は2頭 の女 王 蜂 を一 緒 に いれ る ことを薦 め, vandenEijndeetal.(1991) は 1頭 の女王 蜂 に3-4頭 の セイ ヨウ ミツパ テの働 き蜂 をっ け ると営巣開始率 が高 ま ると して いる.餌 と し て新 鮮 な花粉 と30-50%の シ ョ糖 水 を与 え, 温度 を28-30℃, 相対湿度 を約60%に保 った 部屋 に飼育箱 を配 置す る. 部屋 の照 明 に赤色 ラ ンプ (波長約660人∠m)を用 いると蜂 が飛 び回 ら ず に落 ち着 く. シ ョ糖水 は自動給餌方式 と し, 花粉 は毎 日新 しい もの と交換 す るか適宜追加す る. このよ うな条件 の もとで女王蜂 は1週間前 後 で産卵 を開始 す る. その後,働 き蜂 が羽化す るとコロニーは急速 に成長 す る.働 き蜂 の生産 蓑 1 マルハナバチとミツバチの主な相違点 * マルハナバチ ミツバチ 分布 の中心 種類 生活様式 生活の最小単位 コロニーサイズ 巣の構造 巣の材料 繁殖様式 交尾 交尾時間 栄養交換 収穫 ダンス 働 き蜂のサイズ 働 き蜂の脱針性 攻撃性 採粉行動 ナス科植物への訪花性 採餌距離 活動限界温度 悪天候時活動性
U
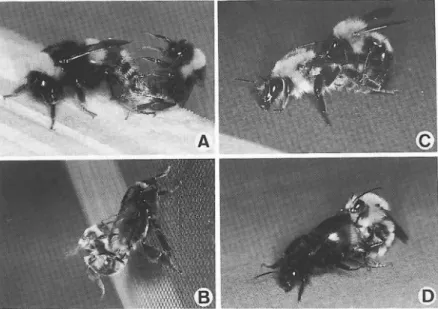
V除去 フイルムの影響 増殖管理 狭い空間への適応性 北半球の温帯,亜寒帯 約300 1年性 女王蜂 (営巣初期) 数十∼数百 不定形 蜂ろうと花粉 生殖個体の多数生産 地上 で普通1回 長 い (約 1時間) 無 無 バラツキが大 きい 無 弱 (温帯産種) 振動採粉 (能動的) . 高い 数百m以 内 6℃前後 高い 鰭 (除去波長による) 特別 な施設 が必要 極めて高い * 松浦 (1993)を参考112 図5 商品化されているセイヨウオオマル-ナバチの巣箱とその内部A.巣箱の外装,B.内部には多くの卵, 幼虫,桶を含む巣が入っている (成蜂と巣を履 う綿は除いてある).C.卵塊と若齢幼虫
,D
.女王蜂と老 熟幼虫,繭 か ら雄蜂 と新女王蜂の生産への切 り換え点 はコ ロニーによって一定 していない.切 り換えの誘 発要寓はまだよ く分か っていない.最初の産卵 か ら3-4か月が経過 した大 きなコロニーか ら は数十頭の雄蜂 と新女王蜂がそれぞれ生産 され る. 交尾 は羽化後2-3日で離巣 した雄蜂を放 飼 している大型 ケージ (約 1m3)内に 6日齢以 上 を経過 した新女王蜂を早朝 に放す ことで誘起 できる (小野 ・柿沼,1991;図 6).雄蜂の交尾 適齢 は5-20日である.交尾用のケージ内には 予 め湿 らせた水苔を入れてお くと交尾後の新女 王蜂 は数 日中に水苔の中に潜 って しまう. これ を掘 り出 して,水苔を入れた小型のガラス ビン に移 して5℃下で貯蔵する. 数か月後に取 り出 して飼育箱 に入れ,同様の処理をすれば造巣 さ せ ることが可能である.R6seler(1985)は交 尾後の新女王蜂に二酸化炭素 ガス処理をす るこ とで,休眠をスキ ップさせ ることに成功 してい る.造巣や産卵には女王蜂の体内における幼若 ホルモ ンの動 態 が関係 して いよ う (ROseler, 1977). 筆者 らはこれまで11種 の 日本産 マルハナバ チの室内飼育 を試 み9種 につ いて は生殖 力ス 卜を生産 させることに成功 している (松林 ら, 1991;小野 ら,1994;光畑 ・小野,1994).最 近 になってオオマル-ナバチの女王蜂に導入種 の働 き蜂をヘルパーとして機能 させる飼育方法 を開発 した(Onoeta1.,1994;図71A,B).ヘ ルパーをっけると営巣率,産卵率が高 まり,最 初 の働 き蜂 の羽化 が早 まる傾向があ る. さ ら に, コロニーの発達 にとって重要な営巣初期の 働 き蜂数の増加速度 も早 まるという結果が得 ら れている.少数例ではあるが実験室内で交尾 さ せた新女王蜂 にヘルパ ーをっ けることによ っ て,休眠期を経 ることな く産卵 させることも確 認できている.導入種のヘルパー効果は日本産 のマルハナバチの種によって異なり,その対合 の可能性について現在検討中である. 日本産マ ル-ナパテの周年飼育が軌道にのれば,在来種 の働 き蜂 をヘルパ ーと して有効 に利用 で きよ う. 日本産 マル- ナバ チの飼育技術 につ いて は,テ レビ,新聞などを通 じて も報道 され,マ ルハナバチに関心を もつ多 くの関係者か ら連絡 を受 け,研究室を御訪問頂 き,その意義などに ついて様々な角度か ら論議がで きたのは収穫で あった.日本産 マルハ ナバ チ利 用 の展望 マルハナバチの利用技術 は, 自然の システム を生産農業に取 り入れた素晴 らしいものと考え られる
.
「なぜ,ヨーロッパ産のマル-ナバチな のですか. 日本にも在来種がいるというのに」
とお尋ねになる方 も多いのであるが, 日本では 昆虫の機能利用 という応用面か ら, この蜂が注 目を浴びたということはほとんどなか ったよう である.確かに利用するマル-ナパテを日本在 来種 に置 き換えることに成功すればこの問題点 を解決でき,使用範囲 も広がると考え られる. 日本にはセイヨウオオマルハナバチと分類学的 に近 い種で実用化の可能性のある種 としてクロ マル-ナバチとオオマルハナバチが生息 してお り,実験室 レベルでは飼育増殖技術はほぼ成功 した段階にある.筆者 らは,彼等が優れた花粉 媒介能力を示すかという点 に関 して温室内で比 較検討 し,行動形質で選択育種することを行 っ ている.そ して, トマ トを中心 とする作物にお ける受粉能力を検証 し,企業 レベルにのせる段 階-発展 させたいと考えている. このマル-ナバチ利用技術を巡 る問題の解決 に関 して最 も重要 と思われることは,関係者全 員が一丸 とな って総合的 に協力す ることであ A 113 る.現在,既 に日本に導入 され急速に普及 して いるセイヨウオオマル-ナバチを野外 に逃 げ出 さないようにす るためにはどうすべきかを真剣 に検討す ることが必要であろう.同時に日本産 マルハナバチの実用化 にむけての試験研究を大 至急進め,将来的には在来種に置 き換えて行 く という考え方を念頭に置 くことが大切 といえよ う.そのためには大規模化 に備えた共同研究の 支援体制が是非 とも必要である. 末筆なが ら原稿を校閲 して頂いた玉川大学 ミ ツバチ科学研究施設の松香光夫教授,同大学農 学部昆虫学研究室の佐々木正己教授 に感謝の意 を表す る. さらに,数 々の貴重な御意見を賜わ った島根大学農学部の前田泰生教授 に厚 く御礼 申 し上げる.情報や資料を提供 して頂いた,農 林水産省,神奈川県農業総合研究所,マル-ナ バチ研究会, (秩)トーメン, JA全農, アピ (樵),東海物産(秩),片倉工業(樵),協和種苗 (秩),三井物産 アグロケ ミカル(秩),Koppert 礼, BBB 社の関係諸氏 にも御礼申 し上げると ともに御理解 と御協力をお願い したい. (〒194 町田市玉川学園6-ト1 玉川大学) 主 な 引 用 文 献 Donovan,B.J.1993. ミツバ チ科 学 14:145-152. Free,J.B.1993.rnsectPollinationofCrops(2nd図6 マル-ナバチの実験室内における交尾 A.セイヨウオオマル-ナバチ,B.オオマル-ナバチ,C.トラ マルハナバチ,D.コマルハナバチ
図7 日本産 マル-ナバチ2種 の室内飼育状況 A.オオマルハ ナバチ女王蜂 と導入種 のヘルパ ー,B.発達 し た コロニー,C.育児室 を温 めるクロマル-ナバチの女王晩 D.生殖 力ス ト産 出期 の コロニー ed.). AcademicPress,Londonpp. 684. Heinrich,B.(井上民二監訳.1991.マル-ナバチの 経済学.文一総合出版,東戻 pp.291,) 池 田二三高 ・忠内雄次.1992.農業 および園芸67: 1213-1216.
Kato,M,1992.Jpn.Entomo1.60:487-494. Kevan,P.G.etal.1991. Proc.Entomol.Soc.
Ontario122:15-19.
松浦誠.1993.マル- ナバチの生態的特性.第2回マ ル-ナバチ利用技術研究会発表資料.p.ト8.
小野正人 ・柿沼友紀子 1991.昆虫 ・応動昆大会. 小野正人 ら.1992.日本動物行動学会大会.
Ono,M.etal.1994.Appl.Entomol.Zool. 29: 印刷中.
Plowright,R.C.and S.C.Jay 1966. J.Apic. Res. 5:155-167.
PrヂS-Jones.0.E.and S.A.Corbet1991. Bum-blebees.Richmond Publishing Co.Ltd. ,Eng-landpp.92.
ROseler,p-F.1985. Apidologie16:165-170.
佐 々木正 己 1994.養蜂 の科学.サイエ ンスハ ウス, 東京 pp.159.
van den Eijnde, ∫. et al. 1991. Acta Horticulturae288:1541158.
VanHeemert,C.etal.1990.BeeWorld71:54 -56.
Velthuis,H.H.W.andL.Cobb 1991, Acta Horticulturae288二199-203.
和 田哲夫 ・栗原純. 1992. ミツバチ科学13:133 -136,
ONO,MASATO.Croppollinationwiththebumble -beesinJapan-itspresentandfuture-.Honeybee Science(1994) 15 (3):107-114. Fac.Agric.,
Tamagawa Univ., Machida-shi, Tokyo, 194 Japan
Bumblebee(BombusterYleStris)isaneffective pollinator of the flower like tomato
(Lycopersiconesculenlum)thatdoesnotproduce nectar. Importsofcoloniesoflaboratory-cull turedB.tenTeStrisfrom EuropetoJapanbegan in 1991.Useofthispollinatorsavesinlabour costsandproduceshigher-qualityfruit Large number of colonies of such commercialB.
terrestrishavebeen Introduced intoJapan for greenhousepollination. From theagricultural viewpoint,itseemsfruitful,howeverfrom the ecologlCalviewpoint,theseimportsmay cause seriousproblems. Thepossibleharmfulinfl u-enceofintroducedbumblebeesonbothindige -nousbeefaunaandflorashouldbeconsidered. Because ofthe problems,pollination ofcrops LithJapanesebumblebeespeciesinsteadofi n-troduced one is recommended. The authour havesucceeded ln laboratory rearingof9out of14 nativeJapaneseBombus. Hedescribes taxonomyandecology,biologicalfeature,l abo-ratory-rearingtechnique,anddiscussescurrent status and future of bumblebee-industry in Japan.