九州大学学術情報リポジトリ
Kyushu University Institutional Repository
食品および環境中の免疫調節因子に関する研究
倉本, 雄一郎
九州大学農学研究科食糧化学工学専攻
https://doi.org/10.11501/3135076
出版情報:Kyushu University, 1997, 博士(農学), 課程博士 バージョン:
権利関係:
の/
食品および環境中の免疫調節因子に関する研究
倉本雄一郎
1998
食品および環境中の免疫調節因子に関する研究
日次
第 1 丘 緒論 …- … ・………一一……… …. 1 第2章 WEHI-279細胞のクローニングおよび
細胞表而免疫グロプリン分布 .……… ………. 5 第3章 食用色素の抗体産生調節機能……一一……一一……一・・… 1 7 第4章 否辛料の抗体産空調節機能………3 3 第5章 ローズベンガルの抗体産生調節機構の解析 …一一……. 46 第6章 ヒトリンパ球培養細胞株の抗体産生に及ぼす
ローズ、ベンガルの影響 .……… …63
第7章 大気汚染物質の免疫調節機能の解明 .………. 76 第8章 総括 …一一………...・H・...……・・・・……… 9 1
参考文献
• • • • • • • • • • •… … ・ ・ … … ・・ ・・ ・・ ・・ … … -……... 1 0 1
第1章 緒論
日本人のライフスタイノレは過去40年で大きく変化した。 食生活については欧 米化が進み、 脂肪の摂取が大きく増えた。 一方、 H本人をとりまく環境は工業 化の発達や臼動車が増加し、 われわれの生活は飛躍的に向上した。 その一方で、
こうしたライフスタイノレの変化はさまざまな負の影響をもたらした。 例えば、
脂肪摂取の増加はガンや動脈硬化の発症を増加させ(Niebaueret a1., 1995・1ρft
et a1., 1996)、 いくつかの食品添加物がアレルギー症状を引き起こすことが報
ばされている(Tarloet a1., 1993; Weber, 1993� Wuhrich. 1993: Nish et a1.
1991)。 また、 自動事の増加は大気汚染を悪化させ、 アレルギー患者の増加に 繋がったことが指摘されている。 ディーゼ、ル機関の排出ガス中の微粒子
(DEP)はアレルギー促進的に働くことが報告されている(Muranaka et a1.
1986; Takafuji et 81., 1987, 1989; Takenaka et 81., 1995)。 日常生活においても、
ラットをタバコの副流煙に慢性的にさらすと、 胸腺依存的および非依存的な抗 原に対する抗体産生細胞の応答が抑制され、 B細胞の機能低下に繋がることや (Savage et 81., 1991)、 タバコの煙に合まれるニコチンはヒト末梢血単核細胞 の抗体産生を調節することが報告されている(Fischer et 8人1994)。 このよう に、 H本人の生活習慣病やアレノレギーといった疾患は過去数十年のライフスタ イルの変化が主な原因とされており、 現在多くの研究者によってその解決法が 模索されているが、 発症機構が複雑であるので治療法は未だ確立されていない。
アレルギ一反応が引き起こされるとき、 一部の食品添加物はその発症に重要 な役割を演じることが知られているが、 その発症機構についてはほとんど明ら かにされていなし、。 われわれの食生活に注目すると、 多くの食品添加物や化学 物質に囲まれていることに気がつく。 われわれは食品添加物を着色、 保存、 着 香など、 われわれの利益になるように使用しているが、 それらに対する多くの 副作用も報告されている。 例えば、 亜硫酸塩は哨息を、 タートラジンはじんま しんをそれぞれ引き起こし(Tarlo et 81., 1993; Weber, 1993; Wuhrich, 1993)、
アナト一色素はアナフィラキシ一反応を引き起こすことが報告されている
(Nish et a人1991)。 タール色素は安価で安定性が高いことから多く用いられ てきたが、 他の多くの合成色素と同様に発癌性、 変異原性を有することが報告 され、 その使用量は減少しつつある。 一方、 古くから食用色素として用いられ てきた天然色素の使用量が近年増加している。 天然色素は合成色素に比べて安 定性に劣るため着色効果を高めるために使用量を多くしなくてはならず、 その ため生体に与える影響が注円されている。 さらに天然色素は着色効果に加えて、
様々な生理活性を有することが報告されている。 例えば、 Curcuma Jongair�ら 抽出されるクルクミンが、 免炭調節作用をイミすること(Liet aJ., 1993; Sui et aJ.,
1993; Yasuni et al., 1993)、 アフラトキシンにより誘導される肝障害を緩和する こと( Soni et al., 1992)、 脂質代謝調節機能を有すること(Fuj iyama -Fuj i war a
et aJ., 1992; Ammon et aJ., 1993; Srivastava et aJ., 1995)、 遺伝子発現調節機能を 有することが報告されている( H uang et a人1991; Kakar et aJ., 1994; Lu et aJ.,
1994)。 また、 紅麹的から抽出されるモナスカス色素はTPAの発婚プロモーシヨ ン効果を抑制することが報告されている(Yasukawaet aJ., 1994)。 また、 香辛 料は食品に風味を付与するほかに腐敗や細菌の繁殖を防ぐ働きがあり
(Adeg oke et al., 1994)、 様々な保存食に利用されてきた。 しかし、 天然色素 や香辛料の免疫調節機能についてはほとんど明らかにされていなし、。
ディーゼノレエンジンの排出ガス成分は大きく2種類に分けられる。 一つはガ
ス状燃焼生成物であり、 二酸化硫黄、 窒素酸化物、 二酸化炭素、 アノレデ、ヒドお よび未燃焼炭化木素が含まれる。 もう一つは粒子状燃焼生成物であり、 硫酸塩、
炭素質粒子、 有機溶媒可溶成分(多環芳香族炭化水素、 ニトロ多環芳香族類)
および抽出不能物質である(加地、 1993; Westerholm et al., 1995; Zapponi et a人 1997)。 アレノレギ一発症の原1&物質としては、 ガス状燃焼生成物よりも粒子状 燃焼生成物の方が注目されており、 例えば炭素質粒子(DEP)はアジュバント 作用を有し、 IgE産生を増強することが報告されている(Diaz-Sanchez et aJ.,
1994; Takenaka et a人1995)。 また、 多環芳香族炭化水素( PAH)が免疫抑制 作用を有すること(Holladayet aJ., ] 994)、 2,3,7,8-Tetrachlorodibenzo-p- dioxin ( TCDD)がB細胞内カルシウムイオン濃度を上昇させること(Karraset
a人1996)などが報告されている。
食品および環境アレルゲンに対するアレルギーは主としてI型アレルギ一反
応により引き起こされるが、 この反応においてはアレルゲン特異的19Eの 誘導 が重要な役割を演じる(Metcalfe, 1991)。 一方、 アレノレゲン特異的IgAはアレ ルゲンの腸管吸収を阻害することによりアレノレギーの発注を抑制する。 また、
アレノレゲン特異的IgGはIgEとの競合によりI型アレルギーを抑制する。 クラス 特異的抗体岩生調節は、 1L-4、 1L-5、 IFN巴α、 IFN-yによって誘導されることが 知られている(Elson et a1., 1994; Gauchat et a1., 1991; Ochel et a1., 1991; Pene et a1., 1988; Rousset et a1., 1991)。 同様な抗体のクラス特異的な産生調節は胞汁酸 やレクチンでも報告されている(Lim et al., 1994a, 1994b; Yamada et al.,
1993)。 胆汁酸はlmM以との濃度でラット腸間膜リンパ節(MLN)リンパ球 のIgE産生を促進し、 IgA、 IgGおよび、IgM産生を抑制した(Lim et a1.,
1994b)。 コンカ ナパリンA(Con A)はIgE産生を増強し、 アメリカヤマゴボ ウレクチン(PWM)はそれを抑制し、 それ以外のクラスには影響を及ぼさな いことが報告された(Lim et a1., 1994b)。 また、 細胞表面レセプターに結合し た抗原特異的IgEを抗原が架橋することにより肥満細胞はヒスタミンおよびロ イコトリエン(LT)などのケミカノレメディエーターを放出し、 アレノレギ一法状
を引き起こすことが知られている(Plaut et a1., 1993) 0 LTは肥満細胞の膜リン 脂質から遊離されるアラキドン酸のリポキシゲナーゼ酸化によって生成される (Dahlen et a1., 1986)。 ロイコトリエンの うちLTB4は、 好中球の走化性によっ て誘導される炎症反応を充進したり(Ford-Hutchinson et a1., 1980)、 可溶性 CD23の発現およびその遊離を増強し、 B細胞のIgE産生を促進することが知ら れている(Dugas et a1., 1990・Yamaoka et a1., 1989)。 こういった背景に立ち、
本研究においては食品および環境中の免疫調節因子の 検索を行い、 それらが免 鋭機構に及ぼす影響について検討した。
食品および環境中の抗体産生調節因子をÌn vÌtroで検索する手段として、 比較 的実験方法が容易で短時間に結果が得られ、 かっ生体に近い細胞応答を再現で きることからラット およびマウスのリンパ球が繁用されているが、 この系では
様々な細胞間相互作用を考慮する必要があることから抗体産生調節メカニズム の解明には適用しずらい面があった。 そこで樹立細胞株であるWEHI-279細胞 を選び、 抗体産生細胞に対するl直接的な最多響および抗体産生調節因子の作用機 構を検討した。
第2章 WEHI-279細胞のク口一ニングおよび 細胞表面免疫グ口プリン分布
第1節 緒論
食品および環境中の 抗体産生調節肉子を川 vitroで検索する手段として、 比較 的容易かっ生体に近い細胞応答を再現できることからラットおよびマウスのリ ンパ球が繁則されているが、 この系では様々な細胞間相互作用を考慮する必要 があることから抗体産生調節メカニズムの解明には適用しずらい面があった。
そこで本章においては、 樹立細胞株を用いた系を確立するためにWEHI-279細 胞を選んだ。 WEHI-279細胞はB細胞の分化の初期段階に位置し、 高密度の細胞 膜表面IgM(終鎖はK鎖)を発現している一方、 培養上清中には抗体が検出され ないことが報告されている(Warner et al., 1979; Gutman et al., 1981)が、 本実 験において用いられた細胞のポピュレーションなど不明な点が多い。 本章にお いてはWEHI-279細胞について、 産生される抗体、 細胞膜免疫グロプリンおよ び抗体に関する遺伝子の発現について検討を行った。
第2節 実験方法
第1 r頁 試薬
FITC標識ラット抗マウスIgE、 FITC標識ヤギ抗マウスIgGおよびFITC標識ヤ ギ抗マウスIgMはSouthern Biotechnology Associates社(Birmingham, AL)より 購入した。
酵素抗体法に用いる試薬として、 洗浄液には、 0.05%Tween 20 (和光純薬 業、 大阪)合有PBS(TPBS)を、 ブロッキング液にはBlock Ace (大日本製薬、
大阪)を用いた。 回相抗体には抗IgG抗体はヤギ抗マウスIgG(Zymed社,
Sanfrancisco, CA)を、 抗IgE抗体はラット抗マウスIgE(EIU社, BrusseIs,
Belgium)を、 抗IgM抗体はウサギ抗マウスIgM F(ab')っ断片(Zymed社)を、
抗IgAHi体はウサギ抗マウスIgA (Zymed社)を用いた。 酵素標識抗体にはベル オキシダーゼ(HRP)標識ヤギ抗マウスIgG F(ab')っ断片(Zymed社)、 HRP 標識ウサギ抗マウスIgM F(ab')っ断片(Zymed社)およびHRP標識ウサギ抗マウ スIgA (Zymed社)を用いた。 またIgE定量においてはビオチン標識ラット抗マ ウスIgE (EIU社)、 およびHRP標識ストレプトアビジン(Prozyme社, San Leandro, CA)を用いた。
目的遺伝子に対するプライマーは、 九州大学医学部付属生体防御医学研究所 から提供された。
第21頁 細胞培養および細胞染色
細胞として、 WEHI-279細胞(九州大学医学部付属生体防御医学研究所から 提供された)を用いた。 細胞は10%ウシ胎児血清(FBS; GIBCO BRL社, Grand Island, NY)を合むRPMI-1640培地を用いて、 5%COっ環境下において継代培養
した。
WEHI-279細胞の増殖曲線を作製し、 経時的に抗体産生量を測定するために、
細胞数をlxl05 cells/mlに調整し10%FBSを合むRPMI-1640培地で1、 2、 3、
4、 6、 8日間培養した。 細胞数は血球計測板を用いて計測し、 培養上清中の 抗体価は第4項に示した酵素抗体法を用いて定量した。
WEHI-279細胞の表面免疫グロプリンを解析するためには、 まず細胞を
10%FBS合有PBS溶液で洗い、 細胞懸濁液にFITC標識された抗マウスIgE、 IgG および�IgMを加えて氷中に30分放置した。 その後、 10%FBS合有PBS溶液で細 胞に結合していない標識抗体を洗い、 最後にステンレスメッシュを通してフロー サイトメーター(EPICS PROFILE II; Coulter社,Miami,FL)で解析した。
第31頁 遺伝子増幅法
的遺伝チの発現を係認するために、 RT-PCR法を用いた。 操作は次のよう に行った。 はじめに、 WEHI-279細胞から全RNAを抽出した。 107個の細胞を PBSで洗浄後、 ペレット状の細胞にTRlzol (GIBCO BRL社)を加え、 取り扱
い説明書に従って全RNAを分離した。 分離したRNAを759るエタノールで洗い、
つづいて水に溶解させた。 10μgのRNAを逆転写酵素(United States
Biochemical社, Cleveland, 0 H)を用いて分離したRNAに対して相補的なDNA (cDNA)を作製した。
つぎに、I�的遺伝子に対するプライマー(九州大学医学部付属生体防御医学 研究所から提供された)を用いて目的遺伝子を増幅した。 反応液には、 0.4 unitsのTaqポリメラーゼ(Fermentas社, Vilnius, Lithuania)、 それぞれ2.5mM
のデオキシリボヌクレオチド三リン酸を合む溶液、 200 �tMのプライマーおよび 10μMのcDNAを混合した溶液を用い、 これに緩衝液および塩化マグネシウム 溶液を加えた。 最後に、 Gene Amp PCR system 2400 (Perkin Elmer社, Foster Cit)んCA)を用い、 Fig.2-1に示したような方法で目的遺伝チを増幅した。
DNA Denaturation 94 oC, 0.5 min
�...
Primer Annealing 55 oc, 1 min
35 Cycles
Primer Extension 72 oC, 1 min
.. ...
Fig. 2-1. Temperatures and Reaction Periods Used During Amplification Cycles.
PCR産物は、 29るアガロースゲル(Sawady Technology社、 東京)を用いて電 気泳動し、 臭化エチヂウム(和光純薬工業)でDNAを染色し、 紫外線によって 得られた画像をKodak Dìgital Science (Eastman Kodak社, Rochester, NY)で撮 影した。
第4項 酵素抗体法
抗体イIfliの測定は、 Yamadaet al. (19 93)の方法に従って行った。 IgGおよび
IgMの定量には直接法を用い、 IgEの定量にはアビジンービオチン法を用いた。
まず、 96穴イムノプレートに同相抗体を加えて370Cにて1時間保温し、 TPBSで
3回洗浄した。 次に、 ブロッキング液を加え、 370Cにて1時間保温もしくは4 Cにて一晩放置した後、 洗浄した。 これ以後の洗浄操作ではアビジンービオチン 法で各4rwJ、 直接法で、各3同行った。 次に、 一次抗体(サンプル上清)を加えて 37Cで1時間保温し、 洗浄した。 IgE定量ではさらに二次抗体を加えて3TCにて 1時間保温し、 洗浄した。 最後に、 酵素標識抗体を加えて37JCで1時間保温し て、 洗浄液で洗浄後、 基質溶液を加えて37Cで15分間反応後、 反応停止液を加
えて415nmで吸光度を測定した。
第3節 結果
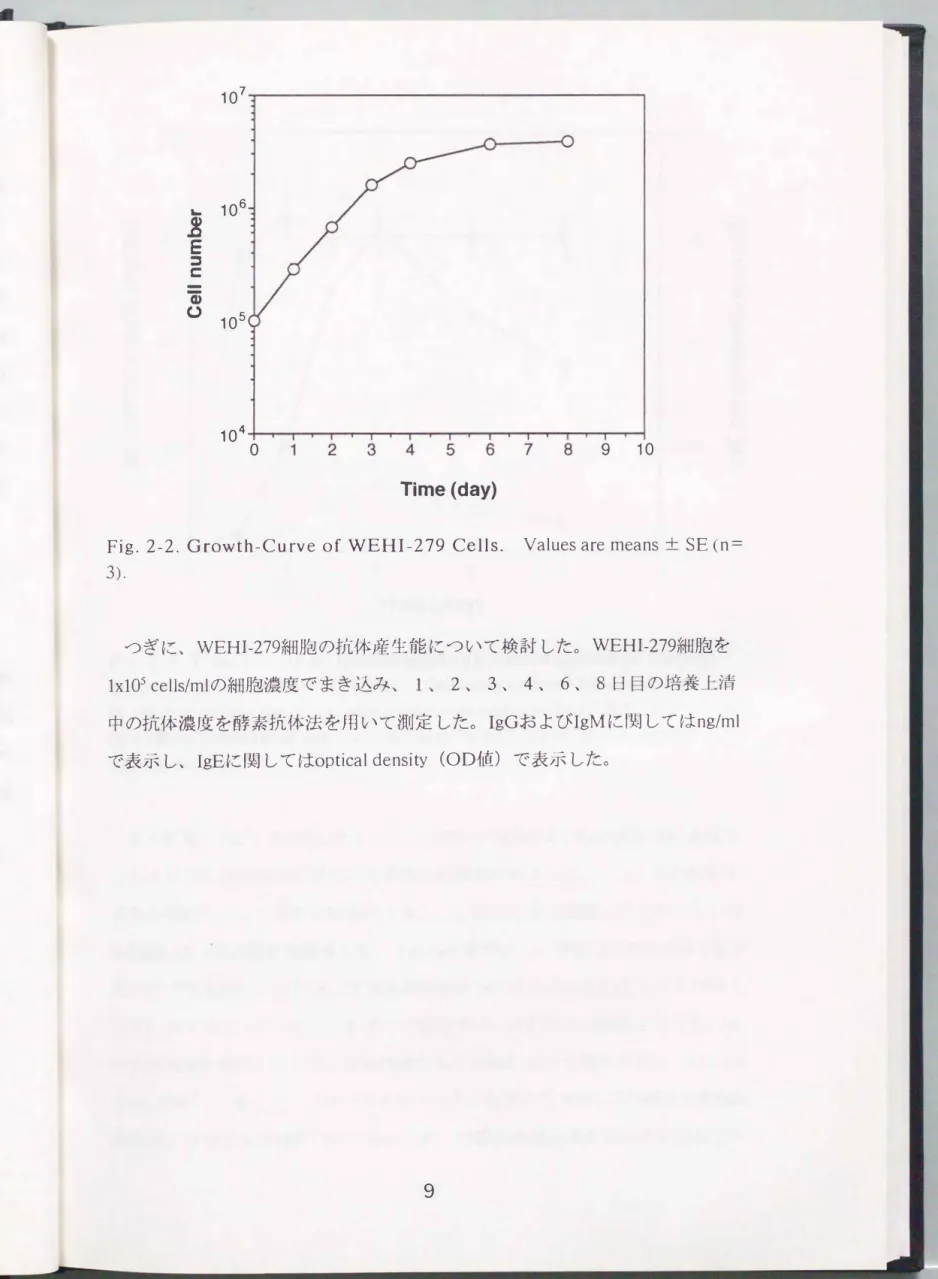
細胞の増殖速度を調ベるため、 WEHI-279細胞をlxl0S cells/mlの細胞濃度で、ま き込んだ。 細胞のまき込み後、 1、 2 、 3 、 4 、 6、 8日日の細胞濃度を血球
計測板を用いて計測した。 Fig.2-2に示したように、 WEHI-279細胞は、 まき込 み開始から 3日以内では対数増殖期にあり、 その後、 増殖速度は低下してまき 込みから 6日目には増殖は停止した。
107
104 0 106
105
』oazhコc=。υ
9 10 7 8
6
Time (day)
5 4 3 2
Values are means + SE (n=
Fig. 2-2. Growth-Curve of WEHI-279 Cells.
3).
WEHI-279細胞を WEHI-279細胞の抗体産生能について検討した。
つぎに、
清 8 1=1 の培養
6、
4、
3、
lxl05 cells/mlの細胞濃度でまき込み、 2、
中の抗体濃度を酵素抗体法を用いて測定した。 IgGおよび、IgMに関してはng/ml で表示した。
で表示し、 IgEに関してはoptical density (OD値)
400
h Ec
300、_,
。c 百'o ‘ c
d ・
200
0 0 2 0 c) 100
。
。 2 4 6 8
Time
(day)
0.20
,t-n h、
0.1 5 、《F
_, 守
0
0.05 もC 0
0.00 10
Fig. 2 -3. Time-course of Immunoglobulin Accumulation in Culture Supernatant of WEHI-279 Cells. Cells were cultured for various days and Ig concentration in the culture supernatant was measured by ELISA. IgG (0) and IgM (.) concentrations were indicated as ng/ml and that of IgE (ム)asOD
(optical density).
その結果、 Fig.2-3に示したように、 上清中のIgGおよびIgM濃度は培養開始 から4日目に最高濃度に達し、 その後抗体濃度は低下した。 一方、 IgE濃度は 培養を開始して1日白から急速に上昇し、 2 に最高濃度に達した。 その後、
IgE濃度は一定の濃度を維持した。 これらの結果から、 WEHI-279に及ぼす影響 を検討する実験においては、 まき込み開始から4日以内に細胞もしくは培養
清を回収することにした。 これまでの報告では、 羽1EHI-279細胞は高密度に細 胞表面IgMを発現しており、 抗体は産生していないという報告がある(Gutman et al., 1981)。 そこで、 フローサイトメーターを用いてWEHI-279細胞の細胞表
細胞をlxl05 cells/mlに調整し、 第2章第2節第2 項に示したように細胞を染色 した。 その結果、 Fig.2-4に示したように、 以前報告されたとおり細胞表面にお けるIgMが他のクラスに比べて最も多く発現していた。 一方、 IgGおよびIgEの 発現も認められたことから、 本実験で用いたWEHI-279細胞は、 IgM抗体のみ ならずIgGおよび�IgE抗体も産生する能力を有することが示唆された。
0% 4% 0% 0.4%
75.8% 24.1% 17.5悦 82.1哨
FITC-anti IgE FITC-anti IgG
0% I 0.8%
0.3% 98.9%
FITC-anti IgM
Fig. 2-4. Expression of Surface Immunoglobulin on W EHI-279 C ells.
Surface Igs were detected by flow-cytometer.
さらに、 WEHI-279細胞の抗体mRNA発現について解析した。 対数増殖期を 過ぎた後のWEHI-279細胞をlx107cellsに調整し、 第 2章第2節第3項に従って mRNAを分離し、 プロダクティブμmRNA (Pμ)、 Pyl、 Py2b、 Py3および、Pε、
および、IgMからIgEへのクラススイッチの際に発現が認められるε鎖の陸型転写 物(Gε)とIgMからIgG1へのクラススイッチの際に発現が認められるGylの発 現を解析するためにRT-PCR法を用いて目的遺伝子を増幅した。 ハウスキーピ
ング遺伝子にはß-actinを用いてcDNAの作製が成功したかの指標とした。 抗体 遺伝子の増幅に用いたプライマーはTable.2-1に示した。 その結果、 Fig.2-5に
ぶしたように用いたWEHI-279細胞は継代中、 常にPε、 Pμを発現していること が明らかとなった。
Table 2-1. Sequences of Primers Used in the Experiment.
Amplified RNA Primer Sequence Size 01 Amplified Product
(Position and Reference)
μa pa
382 bp
Py1
Py2b
Py3
PE
Gl:
Gyl
ß-Actin
JH4・5'-TGGACTACTGGGGTCAAGG-3' (no numbered, Gough et al., 1981) Cμ: 5'ーCATTGGGATTCATCTCTGCG-3'
(439-458, Kawakami et al., 1980) JH4: 5'-TGGACTACTGGGGTCAAGG-3'
(no numbered, Gough et al., 1981) Cy1: 5'-GACAGATGATACTTCTGGGA-3'
(884・903,Honjo et al., 1979) J H4: 5'-TGGACTACTGGGGTCAAGG-3'
(no numbered, Gough et al., 1981)
。y2b:5'-TCCACCCTCGAGGTIAGGAG-3' (954-973, Kataoka et al., 1979) JH4・5'-TGGACTACTGGGGTCAAGG-3'
(no numbered, Gough et al., 1981) Cy3: 5'ーTCCACCCAAGATGTTACCAG・2
(2811・2830, Wels et aL, 1984) J H4: 5'-TGGACT ACTGGGGTCAAGG・2
(no numbered, Gough et al., 1981) CE2: 5' -AGCGATGAATGGAGT AGC-3'
(991-1008, Ishida et a/., 1982) 1�5�ACTAGAGATTCACAACG・2
(771・778,Gerondakis, 1990)
CE2: 5' -AGCGATGAATGGAGT AGC-3' (991・1008, Ishida et a/., 1982) ly1: 5'-CGAGAAGCCTGAGGAATGTG-3'
(351-370, Honjo et a/., 1979) Cy1: 5'-GACAGATGATACTTCTGGGA-3'
(991・1008,Ishida et a/., 1982)
sense: 5'-TGGAATCCTGTGGCATCCATGAAAC・3 antisense: 5'-T AAAACGCAGCTCAGT AACAGTCCG・2
392 bp
419 bp
401 bp
365 bp
423 bp
418 bp
348 bp
隠 義 務部;制緩駒訴事山奪三
| 殻7 ユ - ‘ 、‘給料必至 公IJて - J 貯斗.' �,:�tS�}t�l�:M-��W.f.;�t持続三ょ 隣 鍛 久議議鋭機数機器義弘議長 長 : 込
I三.
..・、'�.':'., . . - -.{� ..司 、 ふ・ 1・ ぷミ1っ .,... ・ -三1
2 3 4 5
Fig. 2-5. Expression of Product ive mRNAs in WEHI-279 CelIs. Each separated mRNA from WEHI-279 cells was amplified by RT-PCR. Each lane was indicated as: Lane 1, �t mRNA (Pμ)、Lane2, Py3: Lane 3, Py1; Lane 4, Py2b; Lane
5. PL
さらに、 Fig.2-6tこぶしたように、 Gεおよび、Gylの発現も認められたことから、
本実験で山いたWEHI-279細胞はIgMからIgGlもしくはIgEへのクラススイッチ が誘導できることがぶ|唆されたD
2
Fig. 2-6. Expression of Germ-line mRNAs in WEHI-279 Cells. Each separated mRNA from WEHI-279 cells was amplified by RT-PCR. Each lane was indicated as: Lane 1, yl mRNA (Gy 1); Lane 2, Gε.
今までの結*=から、 本')ミ験でJIJし\たWEHI-279細胞はIgM、 IgGおよび、IgEf1�
生細胞が出イ1:していることが/J�II女されたため、 つづいてこの細胞をクローニン グして作グラスの抗体jjfk制11胞を作ることについて検討した。 クローニングに は限界希釈法を川いた。îifjì ì'tにI説明すると、 96穴のプレートにおいて、 1シ'(あ
たり0.8個の細胞が入るように細胞数を調整した。 このような濃度の細胞を96穴 のプレートに10枚まき込んで、 1穴に1個のみ入っている部分に印を付けて、
209るFBSを合むRPMI-1640培地中で約2週間培養した。 その後同様な操作をも
う一度繰り返し、 単クローン細胞を得た。 このようにして得られたWEHI-279 細胞の培養上清中の抗体価を酵素抗体法によって測定した。 その結果、 Table 2-2に示したように、 単クローン細胞からIgMおよび、IgG抗体の両方が検出され た。 この結果から、 本実験で用いたWEHI-279細胞にはIgM、 IgG両抗体を産生 するDouble producerが存在することが示唆された。 しかし、 今回用いた実験系 では、 酵素抗体法におけるIgM抗体とIgG抗体の交差反応を考慮する必要があ り、 Double producerの確認のために更なる検討が必要である。
Table 2-2. Immunoglobulin Concentration in the Culture Supernatant of WEHI-279 Clones.
Clone No. IgM IgG Clone No. IgM IgG Clone No. IgM IgG (μg/ml) (ng/ml) (μg/ml) (ng/ml) (μg/ml) (ng/ml)
10.2 13.7 24 9.1 11.3 47 9.9 11.4
2 9.8 14.0 25 9.1 12.1 48 8.9 5.9
3 9.8 13.1 26 9.6 10.2 49 10.1 15.8
4 7.9 7.0 27 9.2 9.7 50 10.1 9.1
5 9.7 13.3 28 8.9 10.5 51 9.6 8.0
6 9.6 11.0 29 9.1 11.3 52 6.9 4.9
7 9.8 14.1 30 9.2 8.8
8 10.0 15.0 31 9.4 11.7 9 9.6 15.5 32 9.7 13.3 10 9.6 13.6 33 9.5 20.1 11 9.8 14.5 34 9.6 15.6 12 9.1 11.3 35 9.8 10.9 13 9.9 14.6 36 9.6 12.9
14 9.0 15.7 37 9.5 11.0
15 10.2 14.2 38 9.6 12.9 16 9.4 12.0 39 9.5 14.7 17 9.2 10.0 40 9.5 8.1
18 9.6 18.0 41 9.7 11.7
19 7.9 8.0 42 10.0 12.6
20 9.7 14.0 43 9.8 14.9
21 9.3 12.5 44 7.6 7.0
22 9.0 9.5 45 9.6 11.1
23 9.4 11.9 46 9.5 9.1
第4節 考察
段々な食品および環境f 11の抗体調節附子の検索を試みるために、 これまでマ ウスおよびラットの牌臓あるいは腸間膜リンパ節(MLN)リンパ球が用いられ た。 このとき問題になるのはT制胞山米のサイトカインの関与を考慮しなけれ ばならないということである(Armitage et a1., ] 993; Collins et a1., 1995; Ling et
a1., 1995)。 そこで、 昨妓B制H胞に作川する抗体産生調節附子についての検栄を
1rうために、 マウスBリンパ服であるWEHI-279細胞を用いることにした。 本実 験においては九州大学医学部付属生体防御医学研究所より提供されたWEHI- 279細胞を)刊し、たが、 本米この細胞はWalter and Eliza Hall Institute (WEHI,
Melbourne, Australia)において樹,Jfされた細胞株である。 樹立、15初のWEHI-
279細胞はB細胞の分化の初則段附に位rf{し、 向街}�の細胞脱衣i面IgM (料鎖は K鎖)を発現しているが、 I13j委け白中には抗体は検出されないことが報告され ている(Warnerεt a1., 1979; Gutmanεt al., 1981)。 しかし、 この細胞について の情報は非常に少ないため産生される抗体、 細胞膜免疫グロプリンおよび抗体 に関する遺伝子発現について検討した。 本実験においてまず、 IgGおよびIgE抗 体が培養上清中において見出された(Fig. 2-3)。 この結果については、 B細胞
より産生したサイトカインはautocrineまたはparacnne的な作用によってB細胞 什身を刺激する(Pistoia, 1997)など、 サイトカインの作用もしくは継代途li1の 突然変異によって抗体産生細胞に分化したことが考えられる。 つぎに、 細胞膜 免疫グロプリンについて検討したところ、 本実験におけるほとんどのWEHI- 279細胞はIgMを細胞表面に発現していたが、 一部にはIgGやIgEを発現してい ることがフローサイトメーターによる分析で/示唆された。 異なるクラスの抗体 を細胞表面に16"J時に発現するとアポトーシスによって排除されるが(Kindler et
a1., 1997)、 本細胞にこのことが吋てはまるかどうかはさらに検討し、 IgGおよ
びIgE発現細胞をクローニングする必要がある。
最後に抗体に関する遺伝子発現については次のようなことが見出された。 B 細胞がIgMからIgGもしくは19Eへクラススイッチを引き起こすとき、 腕型転'手
物(GyおよびGε)の発現が11忍められることが知られているが(Ichiki et alぺ
1993; Li et aI., 1994)、 WEHI-279細胞はGy、 Gε双j]-の発現が認められたことか
ら、 この細胞はIgMからIgGおよび�IgEへクラススイッチを起こす能力を備えて いることがぷされた。 食品中には様々な抗体広生調節肉子が見111されているが (Ahmed et al., 1997; Kuramoto et aI., 1996: Verdaguer et aI., 1993)、 その調節 メカニズムはまだ不明な点が多い。 WEHI-279細胞を用いた食品成分および環 境I�Jの抗体産生調節内子の検索は、 このメカニズムの解明に対して非常に作用 であると忠われる。
第3章 食用色素の抗体産生調節機能
第1節 緒論
アレルギ一反応が引き起こされるとき、 一部の食品添加物がその発症に重要 な役割を演じることが知られているが、 その作用機構についてはほとんど明ら かにされていなし、。 日常の食生活に注目すると、 多くの食品添加物や化学物質 に閉まれていることに気がつく。 食品添加物は着色、 保存、 着香など、 われわ れの利益になるように使用されているが、 それらに対する多くの副作用も報告 されている。 例えば、 亜硫酸塩は端息を、 タートラジンはじんましんをそれぞ れ引き起こし(Tarlo et al., 1993; Weber, 1993; Wuhrich, 1993)、 アナト一色素 はアナフィラキシ一反応を引き起こすことが報告されている(Nish et a人 1991)。
タール色素は安価で安定性が高いことから多く用いられてきたが、 他の多く の合成色素と同様に発癌性、 変異原性を有することが報告され、 その使用量は 減少しつつある。 一方、 古くから食用色素として用いられてきた天然色素の使 用量が近年増加している。 天然色素は合成色素に比べて安定性に劣るため着色 効果を高めるために使用量を多くしなくてはならず、 そのため生体に与える影 響が注目されている。 さらに天然色素は着色効果に加えて、 様々な生理活性を 有することが報告されている。 例えば、 Curcuma longaから抽出されるクルク
ミンが、 免疫調節作用を有すること(Liet al., 1993; Sui et al., 1993; Yasuni et al.,1993)、 アフラトキシンにより誘導される肝障害を緩和すること(Soni et
a人1992)、 脂質代謝調節機能を有すること(Fujiyama-Fujiwaraet aJ., 1992;
Ammon et al., 1993; Srivastava et al., 1995)、 遺伝子発現調節機能を有すること が報告されている(Huang et al., 1991; Kakar et aJ., 1994・Lu et al.. 1994)。 また、
紅麹菌から抽出されるモナスカス色素はTPAの発癌プロモーション効果を抑制 することが報告されている(Yasulくawaet aJ., 1994)。 しかし、 天然色素の免疫 調節機能についてはほとんど明らかにされていない。 そこで、 本研究では食品
添加物の免疫調節機能について検討を行った。
食品および環境アレルゲンに対するアレルギーは主として I型アレノレギ一反 応により引き起こされるが、 この反応においてはアレノレゲン特異的IgEの誘導 が重要な役割を演じる(Metcalfe, 1991)。 一方、 アレノレゲン特異的IgAはアレ ノレゲンの腸管吸収を阻害することによりアレルギーの発症を抑制するo また、
アレノレゲン特異的IgGはIgEとの競合により I型アレルギーを抑制する。 そこで、
本研究ではラット牌臓および腸間膜リンパ節リンパ球の各種の抗体産生に及ぼ す天然、 および合成食用色素の影響について検討した。
第2節 実験方法
第1 r頁 器具および試薬
用いた天然食用色素のうち、 6種のノk溶性色素(ベタニン、 カカオ色素、 カー サマスイエロ一、 コチニール色素、 コーン色素およびモナスカス色素)および 4種の脂溶性色素(ビキシン、 クノレクミン、 ガーディニアイエローおよびラッ
ク色素)は全て(財)日本食品分析センター(東京)より提供された。 合成食 用色素のうち、 アマランス、 ファストグリーンFCF、 インディゴカルミン、 ター トラジン、 水溶性アナトーは和光純薬工業(大阪)から、 ブリリアントブノレー FCF、 ニューコクシン、 銅クロロフィリンナトリウム、 サンセットイエロ- FCFはキシダ化学(大阪)から購入した。 水溶性色素はリン酸緩衝液(PBS、
pH 7.4)に溶解して培養液の1/10量を添加し、 脂溶性色素はジメチルスルフオ オキシド(DMSO;ナカライテスク、 京都)に溶解して培養液の111000量を添加
した。
酵素抗体法に用いる試薬として、 洗浄液には0.05%Tween 20 (和光純薬工業) 合有PBS(TPBS)を、 ブロッキング液にはBlock Ace (大日本製薬、 大阪)を 用いた。 固相抗体には抗IgG抗体はヤギ抗ラットIgGアフイニティー精製F (ab')2断片(Cappel社, West Chester, P A)を、 抗IgE抗体はヤギ抗ラットIgE Fc 領域アフイニティー精製7S画分(Nordic Immunology社, Tilburg, The
Netherlands)を、 抗IgM抗体はヤギ抗ラットIgMアフイニティー精製F(ab')2断 片- (Zymed社, San Francisco, CA)を用いた。 酵素標識抗体はベルオキシダーゼ、
(HRP)標識ヤギ抗ラットIgGアフイニティー精製F(ab')2断片(CappeI社)お よびHRP標識ヤギ抗ラットIgMアフイニティー精製F(ab')2断片(Cappel社) を用いた。 またIgE定量においてはビ、オチン標識マウス抗ラットIgE (Zymed 社)、 およびHRP標識アビジン(DAKOPATTS社, Copenhagen, Denmark)を 用いた。
第2項 細胞調製および培養
実験動物には、 9週齢雄Brown Norwayラット(セアック吉富、 吉富)を用い、
餌はMF (オリエンタノレ酵母、 東京)を用いた。 ラットは購入後直ちに実験に 供するか、 数日予備飼育してから実験に用いた。 予備飼育の際は、 ラット用ケー ジにおがくず床(ホワイトフレーク:オリエンタル酵母)を敷いて飼育した。
リンパ球は、 Lim et a1. (1994a)の方法に従って調製した。 まず、 ラットを エーテル麻酔下で屠殺し、 牌臓および腸間膜リンパ節(MLN)を摘出した。 次 に、 RPMI-1640(日水製薬、 東京)培地中で牌臓およびMLNをすりつぶしてリ ンパ球を分離させ、 Lympholyte-Rat (Cedarlane社, Hornby, Canada)を用いて 密度勾配分離法によりリンパ球画分を得たo
ラットより分離した各リンパ球の培養では10%ウシ胎児血清(Intergen社,
Purchase, NY)合有RPMI-1640培地を用いて2x 106 cells/mIに調整し、 IgE定 においては6時間、 IgGおよび�lgM定量においては72時間、 10%C02インキュベー タ中で培養後、 上清を回収し、 抗体濃度の測定を行った。
第3項 酵素抗体法
抗体価の測定は、 Yamadaet a1. (1993)の方法に従って行った。 IgGおよび IgMの定量には直接法を用い、 IgEの定量にはアピジンービオチン法を用いた。
まず、 96穴イムノプレートに固相抗体を加えて370Cにて1時間保温し、 TPBSで 3回洗浄した。 次に、 ブロッキング液を加え、 370Cにて 1時間保温もしくは4
。Cにて一晩放置した後、 洗浄した。 これ以後の洗浄操作ではアビジンービオチン 法で各4回、 直接法で各3回行った。 次に、 一次抗体(サンプル上清)を加えて 370Cで1時間保温し、 洗浄した。 19B定量ではさらに二次抗体を加えて370Cにて 1時間保温し、 洗浄した。 最後に、 酵素標識抗体を加えて370Cで1時間保温し て、 洗浄液 で洗浄後、 基質溶液を加えて370Cで15分間反応後、 反応停止液を加 えて415nmで吸光度を測定した。
第4項 統計処理
得られた結果は、 Student'sιtestまたはDuncanの統計計算法(Duncan,
1955) に従って有意差検定を行った。
第3節 結果
第1 1頁 ラット牌臓リンパ球の抗体産生に及ぼす水溶性天然色素の影響
Table 3-1にラット牌臓リンパ球の抗体産生に及ぼす水溶性天然食用色素の影 響について検討した結果を示した。 モナスカス色素は終濃度1mMで19B濃度を 上昇させたが、 1mM以下の濃度では19B濃度を低下させた。 それ以外の色素で は19B濃度を低下させる傾向が認められた。 ベタニン、 カーサマスイエロ一、
モナスカス色素は終濃度1 mMにおいてIgGおよび、IgM濃度を強く低下させたが、
コチニール色素の低下効果は弱いものであった。 しかし、 これらの色素のIgG および�lgM濃度低下効果 は10μMおよび100μMでは認められなかった。
Tabl e 3-1. Dose-depen den t Ef f ect of Water-sol uble Color ings on Imm unoglobulin Production by Rat Spleen Ly mphocytes.
Coloring Concentration
(μM )
Conlrol 。
Cacao pigmen t 10 1∞
1αx)
Cochineal pigment 10 1α)
1αx)
Corn pigment 10 1∞
1α)()
Betanin 10
1∞
1αx)
Cartham us yellow 10 1∞
1αx)
tvlonascus pigment 10 1∞
1αx)
Relative Ig content
IgE
1 .∞:tO.07
0.74:tO.0 7**
0.88:tO.08 0.71:tO.04大大
0.76:tO. 07安 0.78:tO.11 0.80:tO.04付
O.72:tO.04犬犬 0.78:tO.07肯 1.09:tO.16
0.43:tO . 0 2 **犬 O.54:tO .0 5安肯背
0.76:tO.09安
0.69:tO.09犬 0.89:tO.05 0.83:tO.04*
0.58:tO .0 3 *大女 0.81:tO.06犬 1.26:tO .04安犬
IgG
1.∞:tO.03
0.96:tO.02 1.03:tO.02 1.02:tO.01
0.99:tO.04 1.08:tO.03 0.71:tO .04安安
O.95:tO.03 1.03:tO.04 O.85:tO.0 1 **
0.94:tO.02 0.91:tO.04 0.27:tO.0 2女六犬
0.92:tO.01 1.02:tO.04 O.∞:tO.OO女女大
0.97:tO.02 0.92:tO.03 0.09:tO.0 6村女
I gM
1.α):tO.03
O.95:tO.02 1.11:tO.01犬犬 1.18:tO.03付
1.06:tO.02 1.28:tO .0 2女安安 0.82:tO.02付
1.02:tO.01 1.07:tO.01犬 1.01:tO.02
1.12:tO.04犬 1.18:tO.04女大 O.36:tO .0 1川女
1.13:tO.03女 1. 12:tO.07 0.43:tO .00オ安安
1.07:tO.02 0.94:tO.03 0.15:tO .0 6台女大
Ig concentrati ons of the control culture were 6.81:t0.51 ng/ml for IgE, 30.5:tO.92 ng/
ml for IgG and 78.2:t2.3 ng/ml for IgM. Val ues are the mean:tSD of 4 cultures. * p く0.05 ,** pく0.01 an d *** pく0.001 compared with the control.
第21頁 ラット牌臓リンパ球の抗体産生に及ぼす非水溶性天然色素の影響
次に、 ラット牌臓リンパ球の抗体産生に及ぼす非水溶性天然色素の影響につ いて検討した。 Tabl e3-2に示したように、 ガーディニアイエロ一、 ラック色 素およびビキシンは、 今回用いた終濃度(] -100μM)ではIgE濃度を低下させ たが、 クノレクミンの効果は弱いものであった。 クルクミンは100μMではIgGお よびIgM濃度を低下させたが、 1および10μMでIgM濃度を上昇させた。 同様な
IgM濃度上昇効果はクルグミン以外の色素でも認められた。 中でも、 ラック色 素は最も高い促進活性を示し、 100μMでIgG濃度を有意に上昇させた。
Table 3-2. Do se-dependent Ef f ect of Water-insoluble Colorings on Immuno globulin Pro duction by Rat Spleen Lym phocytes.
Coloring Concen tra tion Relative Ig content
(μiv1) IgE IgG I g !\1
Control 。 1.∞:tO.04 1.∞五).02 1.∞:tO.01
Gardenia yellow 0.54:tO. 1 1安安 1.04:tO.01 1.24:tO. 04士大 安
10 0.67:tO.0 9付 1.04:tO.04 1.16:tO.04犬犬
1α) 0.83:tO.06犬 0.9 0:tO.03肯 1.11:tO.05大
Laccaic acid 1 0.4 6:tO.0 9オ安犬 1.06:tO.03 1.25:tO.0 3肯女大 10 0.45:tO.07大女大 1.16:tO.07大 1.33:tO. 04大大大 1α) O.61:tO.0 9犬士 1.40:tO.1 2犬安 1.6 2:tO. 1 1安安安
Bixin 0.54:tO.04女女大 1.04:tO.02 1.27:tO.0 2大古女
10 0.56:tO.06 安安犬 1.03:tO.03 1.25:tO.04女犬大 1∞ 0.80:tO. 04付 0.96:tO.03 1.13:tO.05安
Curcumin 1 0.92:tO.1 1 1.05:tO.02 1.3 1:tO .0 3川女
10 0.86:tO.11 1. 14:tO.10 1.29:tO.06大女
1∞ 0. 86:tO.10 0.14:tO. 0 3村女 0.24:tO. 0 2犬台女
The Ig concentrations of the control culture were 6.45:t0.25 ng/ml for 19E, 24.1:t 0.52 ng/ml for IgG and 64.7:t0.95 ng/ml for IgM. Values are the mean:tSD of 4 cul tures. * pく0.05, **pく0.01 and *** pく0.001 compared with the control.
第3項 ラット牌臓および腸間膜リンパ球の抗体産生に及ぼすアゾ色素の影響
ラット牌臓およびMLNリンパ球の抗体産生に及ぼす食用色素の影響について、
ム成色素であるアゾ色素を用いて検討した。 アゾ色素は構造中にアゾ結合(ー N=N-)を有するのでこのような名が付いている。 ここで用いた食用色素は、
アマランス、 ニューコクシン、 タートラジンおよびサンセットイエローFCFで ある。 これらの色素と共に牌臓リンパ球を72時間培養し、 上清を回収した後そ れに合まれる抗体価を酵素抗体法で定量した。 抗体価は、 PBS存在下の抗体価 (コントローノレ)に対する相対的な濃度として表した。 まず、 ラット牌臓リン
パ球の抗体産生に及ぼすアゾ色素の影響について検討した。 Fig.3-1に示したよ うに、 アゾ色素は10-1から103μMの閣の濃度では19Gおよ び�IgMレベノレに影響を 及ぼさなかった。 ニューコクシン、 タートラジンおよびサンセットイエロー FCFは100から103μMの間の濃度では19Eレベルに影響を及ぼさなかったが、 ア マランスは101 �MでIgEレベノレをコントロールの半分に低下させた。
8 5
IgE
2.0
1
Amar釧h1.5
1.0
0.5
。 。
�
New Coccine0.5
。 。
IgG IgM
g
i
回圃E羽 ::t\:: 0.5
。 。
t
Su峨tYellow FCF1.5
1.0 rC、父一--・E・--..一一一一一二一一一一一一一一--_r一一一・ぶ・一一一一一一一一一一一一一一一一ーさ一一一-三一一一一工一一一-二4・
0.5
O.�_ -1 __0 __1 __2 _�3 _� -1 _�O __1 __2 __3 ・1 __0 __, __2 __3
10-' 10v 10' 10' 10" 10-' 10v 10' 10' 10� 10-' 10v 10' 10� 10v 10
Food Coloring (μM)
Fig. 3-1. Effect of Azo Colorings on Immunoglobulin Production by Rat Spleen Lympho cytes. Spleen lymphocytes were cultured with various azo dyes at around 10-1 to 103 !J.M for 72 hr and the culture supernatant was collected to evaluate Ig concentration.
IgE 2.0
1.5
0.5 r Amaranth 0.0
C 1.5 0 窃1.01""ー
.... ‘-
z CI) 0.5
ü v.V l New Coccine
8
z 0O-m 1.5 0 〉 活1.0r
巴0.51。
0.0
Tartrazine
o.S[
Sunset Yellow FCFIgG IgM
0.0 10・1-' 10__0 v 10' __1 10' __2 10v __3 10__-' -1 10__0 v 10__1 ' 10' __2 10v __3 10__-1 ' 10v __0 __1 10' _�2 10' 10v __3 10 __4
Food Coloring (μM)
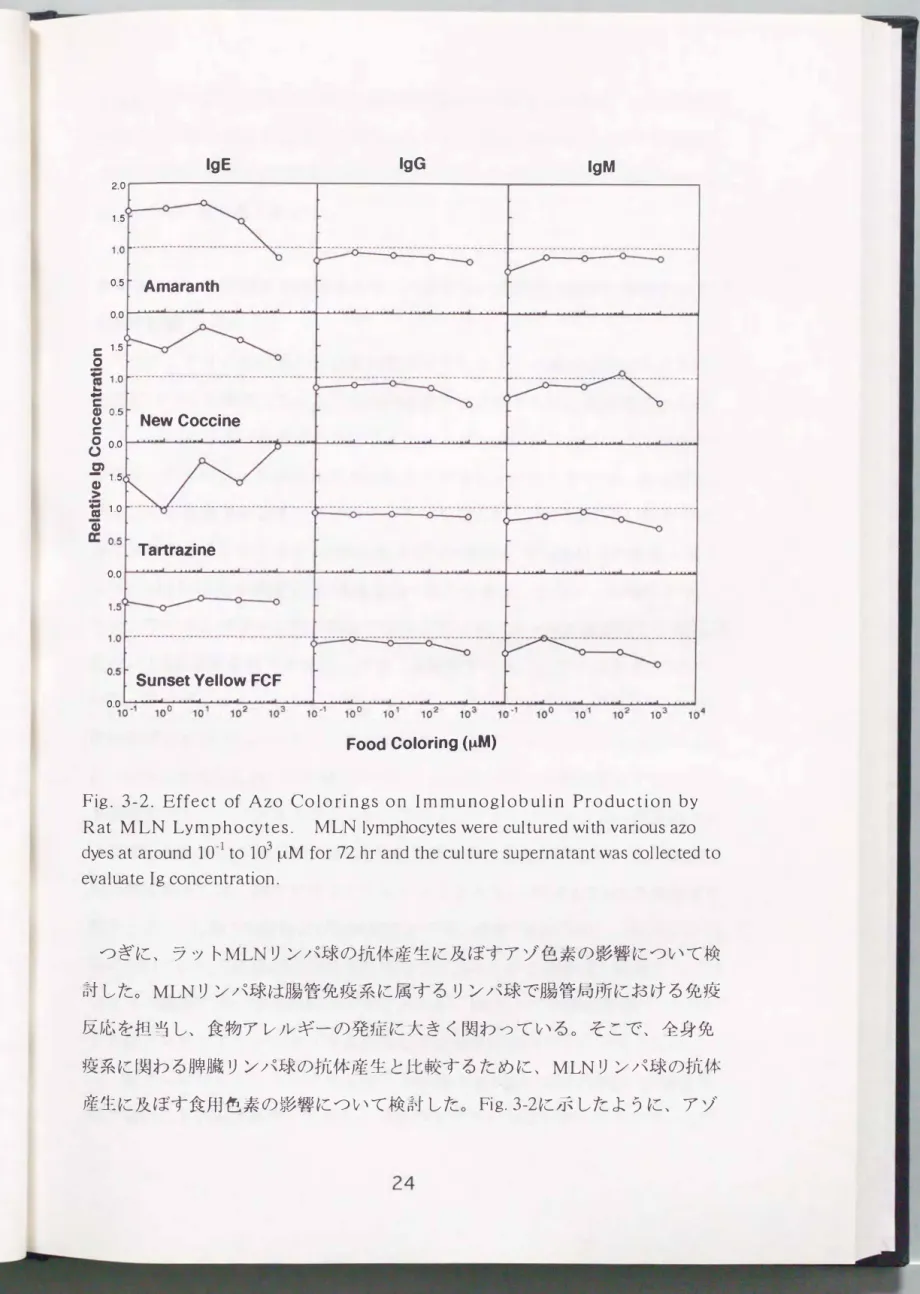
Fig. 3-2. Effect of Azo Col o rin gs on Im m u no globulin Production by Rat MLN Lymphocytes. MLN lymphocytes were cultured with various azo dyes at around 10-1 to 103 I--lM for 72 hr and the culture supern atant was collected to
evaluate Ig concentration.
つぎに、 ラットMLNリンパ球の抗体産生に及ぼすアゾ色素の影響について検 討した。 MLNリンパ球は腸管免疫系に属するリンパ球で腸管局所における免疫 反応を担当し、 食物アレノレギーの発症に大きく関わっている。 そこで、 全身免 疫系に関わる牌臓リンパ球の抗体産生と比較するために、 MLNリンパ球の抗体 産生に及ぼす食用色素の影響について検討した。 Fig.3-2に示したように、 アゾ
色素は用いた濃度領域ではIgGおよびIgM濃度に影響を及ぼさず、 この結果は 牌臓リンパ球の場合と同様であった。 しかし、 IgEレベルは全てのアゾ色素の 存在下では、 対照区の1.5倍に上昇した。 一方、 アマランスは103μMにおいて IgEレベノレを若干低下させた。
第41頁 ラット牌臓および腸間膜リンパ節リンパ球の抗体産生に及ぼす非アゾ 色素の影響
つぎ、に、 アゾ色素以外の合成食用色素がラットリンパ球の抗体産生に及ぼす 影響についても検討した。 ここで用いた非アゾ色素として、 水溶性アナト一、
トリフェニルメタン色素であるブリリアントブノレ-FCFおよびファストグリー ンFCF、 クロロフィル誘導体である銅クロロフィリンナトリウム、 およびイン
ディゴイド色素であるインディゴカルミンを選んだ。 Fig.3-3に示したように、
銅クロロフィリンナトリウムはIgGおよび、IgM濃度を102μM以上の濃度で低 させ、 10μM以上の濃度でIgEj濃度を強く低下させた。 さらに、 水溶性アナ
ト一、 ファストグリーンFCFおよびブリリアントブルーFCFは10および102μM においてIgE濃度を低下させた。 一方、 水溶性アナト一、 ファストグリーン FCF、 インディゴカルミンおよびブリリアントブ、ノレーFCFはIgGおよび�IgM濃度 には影響を及ぼさなかった。 これらの結果は、 銅クロロフィリンナトリウムは 10μM以上の濃度において免疫グロプリン産生を抑制する効果を有すること、
水溶性アナト一、 ファストグリーンFCFおよびブリリアントブルーFCFは10お よび102μMにおいて選択的にIgE産生を抑制することを示唆している。 MLNリ
ンパ球においては、 銅クロロフィリンナトリウムは1μM以上でIgE濃度を強く 低下させ、 10 !-!MでIgGおよび、IgM濃度を低下させた(Fig.3-4) 0 MLNリンパ 球において、 この色素の抗体産生抑制効果が認められる濃度は、 牌臓リンパ球 のときの濃度に比べて10倍低かったことから、 MLNリンパ球は牌臓リンパ球よ
りも銅クロロフィリンナトリウムに対して感受性が高いことが示唆された。
方、 銅クロロフィリンナトリウム以外の色素はIgGおよび、IgM濃度に影響を及 ぼさないことが示された。 しかし 水溶性アナトーおよびファストグリーン
FCFは10および102μMにおいてIgE濃度を低下させた。
IgM IgE IgG
2.0
Water-80luble Annatto
1.5
Brilliant Blue FCF
0.0
1.5
1.0
E02旬』-Fcouzoom-ω〉沼周-oz
0.0 0.5
Cu-chlorophyllln
。 。 1.5
1.0
0.5
FastG同時n FCF
1.5
1.0
Indigocarmlne
0.0
0.5
10.
103 102 101 100 10-1 103 102 101 100 103 10・1
101 102 100
0.0 0..-.:
10-'
Food Coloring (μM)
Fig. 3-3. Effect of Non-Azo Colorings o n lmmunoglobulin Production by Rat Spleen Lymphocytes. Spleen lymphocytes were cultured with various colorings such as triphenylmethane dyes (brilliant blue FCF and fast green FCF),
indigoid dye (indigocarmine), water-soluble annatto and sodium Cu-chlorophyllin at around 10-1 to 103 �M for 72 hr and the culture supernatant was collected to evaluate Ig concentration.
。
3
z z @ 〉.
IgE IgG IgM
2.0
Water-soluble Annatto
1.0
0.5
0.0
�
Brllliant Blue1.5
1.0
0.5
。 。
↑Cu-cf可lorophyllin
1.5
1.0
0.5
0.0
� Fast Green FCF
1.5
1.0
0.5
。 。
�
Indlgocarml問1.5
1.0
0.5
o.q10_ -1 -' "_0 10v 10"_1 ' "_2 10� 10" _3� 10・1-' 10"_0 v "_1 10' "_2 10� 10� "_3 "_ -1 10-' "_0 10v "_1 10' "_2 10� __3 10� "_-4 10
Food Coloring (μM)
Fig. 3-4. Eff ect of Non-Azo C olorin gs on lmmu n oglob ulin Pr oduct i on b y Rat MLN Lymp h o c yt e s. MLN Iymphocytes were cultured with various colorings such as triphenylmethane dyes (brilliant blue FCF and fast green FCF),
indigoid dye (indigocarmine), water-soluble annatto and sodi um Cu-chlorophyllin at around 10-1 to 103 !J.M for 72 hr and the culture supernatant was collected to evaluate Ig concentration.
第5項 ラットリンパ球の抗体産生に及ぼすクロロフィル誘導体の影響
銅クロロフィリンナトリウムが牌臓およびMLNリンパ球の免疫グロプリン産 生を抑制することが示されたので、 われわれはラット牌臓およびMLNリンパ球 の抗体産生に及ぼすクロロフィル誘導体の影響について検討した。 クロロフィ ノレ誘導体として、 銅クロロフィリンナトリウムの他に鉄クロロフィリンナトリ ウム、 およびマグネシウムクロロフィリン(クロロフィノレーaはメチノレおよびフィ チノレ基を有し、 クロロフィル-bはアルデ、ヒドおよびフィチノレ基を持つ)を選ん だ。 Fig.3-5に示したように、 銅および鉄クロロフィリンナトリウムは100μM の濃度で牌臓リンパ球のIgE濃度を低下させた。 マグネシウムを持つクロロフィ ノレ誘導体のうち、 クロロフィノレーbは牌臓およびMLNリンパ球のIgE濃度を上昇 させたが、 クロロフィノレーaはIgE濃度には影響を及ぼさないことが明らかとなっ
た。 これらの結果より、 ポノレフィリン環の金属配位子および、アルデ、ヒド基が抗 体産生の調節に重要な役割を演じていることが示唆された。
Spleen MLN
Control
Cu叫chlorophyllin, Na-salt
Fe剛chlorophyllin, Na-salt
Chlorophyl卜a
Chlorophylトb
。 2 3 4 。 2 3 4
IgE (nglmり
Fig. 3-5. Ef f ec t of Chlorophyl ls and their Derivatives on IgE
producion by Rat Spleen or MLN Lymphoc ytes. Cells were inc ubated for 72 hr and c ul t ure super natantwas applied to ELISA (n=4) to measure IgE
conc entration. a,bYalues i n the same group without a com mon le t ter are
5
significantly different at pく0.05.
第4節 考察
過去十数年の日本におけるアレルギー患者の 増加の原因のーっとして、 環境 中の汚染物質が挙げられる。 多くの研究者は環境中の汚染因子とアレルギー擢
患率の関係について研究してきた。 ディーゼ、ル排気ガス中の微粒子(DEP)を
オブ、アノレブミン(OVA)と共に腹腔および鼻腔に投与したとき、 DEPは免疫 グロプリン産生に対してアジュパント効果を示すことが明らかとなった
(Takafuji et a1., 1987, 1989; Takenaka et a1., 1995)。 しかし、 アレノレギー患者の
増加は、 これら大気汚染物質の増加のみで説明することは出来ない。 そこで、
食品成分とアレノレギ一発症との関係について検討した。
食餌および環境アレルゲンに対するアレノレギーは、 おもにI型アレノレギ一反応、
つまり、 アレルゲン特異的IgEの誘導が重要な役割を演じる反応によって誘導 されると考えられている(Metcalfe、 1991)。 クラス特異的な抗体産生調節で は、 胆汁酸やコンカナバリンAはラットリンパ球のIgE産生を増強する(Lim et a1., 1994)。 一方、 ポークウィードマイトジェンはその産生を抑制することが 報告されている(Lim et a人1994)。 本実験において、 種々の天然食用色素お よび合成食用色素ラットリンパ球が抗体産生に及ぼす影響について検討した。
その結果、 各天然食用色素は1から10μMにおいてIgE産生を抑制した。 これら の色素は、 様々な食物に0.1% (2-3μM)の濃度で添加されている。 クルクミン を合むCurcuma 10ngaの粉末は、 漢方薬として用いられるときその投与量は6-10 g/dayであり、 この量はクルクミン800μMに相当する。 この濃度は食品添加物
として摂食される濃度の約400倍であり、 クノレクミンの吸収率については不明 な点が多いことを考慮する必要があるが、 クノレクミンを合めた天然色素はIgE 産生を抑制することによって抗アレルギー的に作用することが示唆された。
方IgGおよび、IgMは体液性免疫において重要な働きをする抗体である。 水溶性 天然色素は1 mMで、 非水溶性天然色素のクノレクミンは100μMでIgGおよび、IgM 産生を抑制したが、 10μM以下の濃度ではその効果は認められなかった。 一方、
脂溶性天然色素はIgM産生をわずか1μMで増強した。 種々の食品成分による同 様なIgM産生増強効果が、 ヒトーヒトハイブリドーマおよびヒトリンパ球におい て報告されている(Yamadaet al., 1989, 1990a, 1990b, 1991; Maeda et al.,
1990)。 今回用いた天然食用色素の免疫調節機構は明らかでないが、 桂皮酸2 モルからなるクルクミンはその2量体構造によって、 免疫応答調節能を有する
ことが報告された(Liet al., 1993)。
即 位COOH
Caffeic Acid
Fig. 3-6. Structur e of Cu rcumin a nd Caffeic Acid.
また、 ガーディニア色素の主成分であるクロシンおよびアナト一色素の主成 分であるビキシンは抗酸化剤として用いられている(Escribanoet al., 1996;
Thresiamma et al.. 1996: Ursini et al.. 1994) ので、 ガーディニア色素およびアナ ト一色素のIgE産生抑制効果は抗酸化作用が関係していることが考えられた。
今回の研究により、 脂溶性食用色素は体液性免疫応答に対し、 増強作用を有す ることが示唆された。 さらに、 本研究で用いられた天然食用色素は主要成分以 外にも微量成分を合んでおり、 これらが免疫グロプリン産生に影響を及ぼして いることも考えられる。 一方、 合成食用色素に関しては、 アゾ色素は、 牌臓リ ンパ球の抗体産生に対しては有意な影響を与えないものの、 MLNリンパ球に対 してはIgE濃度を上昇させることが示された。 また、 胆汁酸をIL-4もしくはIL-5
の存在下でLPSと共に添加したとき、 牌臓リンパ球のIgE産生は抑制されるが、
MLNリンパ球のIgE産生は増強されることが報告されている(Lim et a1.� 1994b;
1995)。 これらの結果より、 食品成分に対する牌臓およびMLNリンパ球の応答 はそれぞれ異なることが示唆された。 さらに、 病因における食品成分および食 品添加物の役割と、 それに関連する免疫学的変化を確立するために行われた研 究で報告によると、 食品添加物(タートラジンを合む)とアレルギ一発症との 聞に相関性は認められないことが示された(Moraleset a1., 1995)。 それゆえに、
全身免疫系に及ぼすアゾ色素の影響は、 腸管免疫系に及ぼす影響より穏やかで あることが考えられる。
非アゾ色素では、 今回選んだ濃度においてIgGおよびIgMはコントロールと 同じ産生レベルであった。
川。ヒï �N'(YH5)ーC向。
Fig. 3- 7. Structure of Triphenylmethane Colo rings.
方、 トリフェニルメタン色素であるブリリアントブルーFCFおよびファス
トグリーンFCF、 さらに銅クロロフィリンナトリウムは、 牌臓リンパ球では1μ
M以上、 MLNリンパ球では10μM以上の濃度で牌臓およびMLNリンパ球のIgE 産生を強く抑制した。 また、 IgE産生抑制は銅および鉄クロロフィリンによっ て誘導されたが、 マグネシウムを配位したクロロフィリンにはその効果は認め
られなかった。
この結果に関して次のような報告がある。 銅クロロフィリンナトリウムの毒 性は遊離した二価銅イオンに由来するというものである(Wordenet a人