The Boreal Forest Society
NII-Electronic Library Service
The Boreal Forest SOCIety
日林 北支論 59, Feb
.
2011大気
C
(渥
濃度
の増
加 が
エゾ
ノ
キ
ヌ ヤ
ナ ギ
の生 理 特
性
に
与
え る
影響
所冖
支 タ 朧 輙瓢
学 合 聡 蜥 駄 森 林 埼北岡 哲 ・
上村
章・
飛田博
順・
宇都
木 玄 丸 山 温 宮城
敦子・
内宮
博 文 は じめに ヤ ナギ属 (Saiix
)の樹 木は北 半 球の暖 帯か ら寒帯に か け て広 く分 布 する(15)。
遷移 初 期の樹 木で成 長 速 度 が 速 く,
挿 し木で も 簡単に増殖 す る とい う特 長 を 持 ち,
さ ら に 再 生能力 が 高 く,
収穫後 も容 易に萌 芽 する(8,
15)。
こ の よ う な 性 質 か らバイ オ燃 料 資源 とし ての利 用 を目的に した ヤ ナ ギの 育 種 や 栽 培 実 験が行 わ れて いる(5)。
産 業 革 命以降,
CO2濃 度が急 激な速 度で増加 してい る (3)。
地 史 学 的 視 点か ら みて も,
現 代のCO2増加速 度は植 物 が 未 だ 経 験 し たことの ないほ ど 急 激で あ る(2)。
CO2は 光合 成の 基 質で あ る た め,
その増 加は植 物の分子か ら個 体レ ベ ル の成 長反応にまで,
直 接 的にも間 接 的にも様々 な影 響 を与え る(2,
7,
10)。
こ の こ と が植物,
特 に多年 生 植 物で ある樹 木のCO2増加 に 対 す る 反 応 の解明 を難 解 な ものに してい る。
こ のた めCO2 濃 度の増加に対する ヤ ナ ギ属 樹木の応 答 を 明 らか にしつ つ,
将来の高CO2濃 度に適 した育種 選 抜 を 行 うこ とは,
将 来 持 続 的にバイ オ燃 料 資源 を 供 給 す るための重 要 な 課 題であ る
。
Karp and Shield(5)はバイ オ燃 料 資源となる植 物の代 謝生 理 や 遺 伝 的 特 性 を 明 ら か にす ること が,
将 来の バ イ オ 燃 料の増産 と その持 続 的 な 利 用 に とっ て有 用 な 基礎 情報 を 提供 する と述べ て い る。
そこ で本 件 研 究は,
北 海道でバイ オ燃料資 源 とし ての 利 用 が 期 待されてい るエ ゾノキヌ ヤ ナギを用い て,
高CO2 下に お ける葉の光合 成 特 性と代 謝の解 析 を行っ た。
材 料と方 法 2010年4月 下旬に挿し穂を鹿沼 土 :赤玉土 1:1の割 合 で混 合したビニ ル ポッ ト (直 径 21cm) に 植 え 付 け た。
植 え 付 け た 苗 は 森 林 総 合 研 究 所 北海 道 支所の 自然 光人工気 象 室 (気 温 昼 25°
C1 夜 16℃)で育 成した。
CO2濃度 は 370ppm (対 照 ),
720ppm (高CO2)の 2 条件 と した。
施肥 は Kitao et al,
(6)を 参 考 に して,
極端 な養 分不 足 にな ら ない ように 与 えた。
250mgN 相 当の窒素を 固 形肥料で与 え,
残 りの 500mgN は液肥で週 1回 50mgN つつ 10週 間 与 え た。
7 月 下旬に先端 よ り7−
8枚目の成 熟し た葉 を対 象に二 酸化 炭 素一
光合 成 関 係 の 測 定を行 っ た。
測 定はLI
−
6400(Li−
cor 社 米 国)を 用 い,
測 定手II
匱は Long andBernacehi(9)に従っ た
。
最 大Rubisco活 性 (V。max )と最大電 子伝 達 速 度 (
Jmax
)(1)は Onoda et al,
(〃)を参 考に算出 し た。
光合 成 速 度 を 測 定 した葉か ら】cm2 のディス クを5枚打 ち 抜 き
,
80°
Cで 48 時 間 乾 燥させ,
窒素 含 有量 と 可 溶 性糖類
・
デンプン の分 析に 供 し た。 窒 素含有量の分 析は全自動 元 素 分 析装置 vario MAX (Elementar Analysensysteme
GmbH
,
独 国)を 用い た。
可溶 性 糖 類・
デンプン の分析 は フ ェ ノー
ル スル ホ ン酸 法 (4,
12)を 用いて行っ た。
葉の代 謝 産 物はMiyagi et al.
(〃)に 従い,
キャ ピラ リー
電気 泳 動 / 質量分析法システム (Agilent Technolegies社,独 国 ) を 用いて 定 量 し た。統 計 解 析はスチュー
デン トのt検 定 を 用 い , 高CO2 処理 の効 果の有無 を検定した。
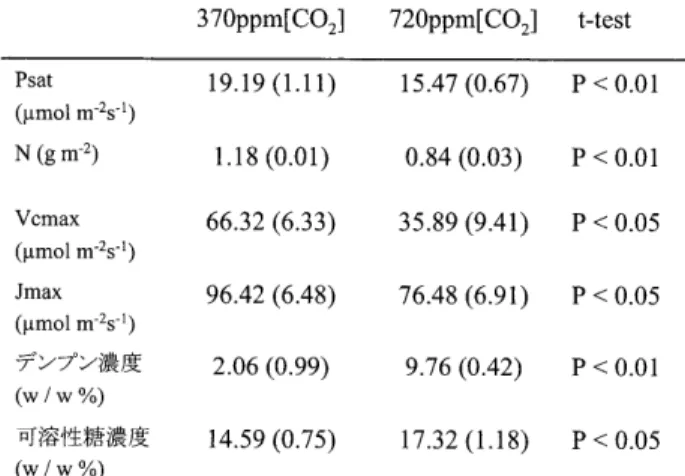
結果と考 察 1)葉の生 理 特 性:高CO2処 理 に よっ て,
生 育CO2 濃 度にお け る光飽 和の光 合 成 速 度 (Psat)の有 意 な低 下が見られた (表一
1
)。
こ の現 象は光 合 成 活 性 を示すVcmax,
Jmaxに お い て も みられ (表一
1),
光 合 成の負の制 御 現象(2,
7,
10)が 生 じた と考 え ら れ た。
葉の 窒 素 量 は 高CO2 処 理 によっ て有 意 に 減 少 し た (表一
1)。一
方,
可溶性糖 濃 度お よ びデンプ ン濃 度は高CO2処理で有 意に増加 した(表一
1)。
可溶 性 糖 類 の増 加は光 合 成 系の酵 素 やタン パ ク質を減少 させ る とい われてい る(16,
17)。
こ のた め 可溶 性 糖 類の増加 は光 合 成 の負の制 御 現 象 や窒素量の 低 下を 生 じ た一
因 で あ る と推 察 される。
2)代 謝 特 性:高COz 処 理に よっ て ア ミ ノ酸 (Arg
,
Asp,
Asn
,
Gln)の 減 少 傾 向が見ら れ た(図一
D
。
ま た ミ トコ ン ド リア のTCA 回 路 に 関 わ る 有 機 酸(Citrate
,
Iso
。itrate,
Malate,
Aconitate
,
Pruvinate),
糖リン酸系(DHAP , 3PGA , FBP,
6PG,
RuBP )の減少 が 見られた(図
一
1)。
これ らの傾 向は,
ヤ ナ ギ と同じ よ うに再生能力 が 強い多年 生 草 本 植 物(エ ゾ ノ ギ シギシ)の代 謝 解析 結果と一
致 す る(1ノ)。
可溶 性 糖 類とデンプンが光 合 成に与 える影 響は異 なる と言わ れてい る (1の。
可溶 性 糖 類の蓄 積はシグ ナル とな っ て 遺 伝 子 等 に働きかけ,
光 合 成 系のタン パ ク質 等の減 少 を招 くため(/6,
i7),
代 謝 解 析でみ られ た カル ビンサ イ クル の 糖リン 酸 (RuBP や 3PGA,
FBP )の 減少や,
光 合 成速 度の低 下,
最 大Rubisco 活性の低下 と関係が あると考え ら れ る。
デンプン の蓄 積は葉緑 体 内 に お け るCO2 の拡 散 を 阻 害 するこ と で,
間接 的に 光合 成に影 響 与 えると言 わ れ て い る(fO)。 今後 デン プン の 蓄積が ヤ ナギの光 合 成に与 え る影 響につ いて さ ら に 明 ら か に してい く必要がある。
3)お わ りに:本 研究の結果か ら,
高CO2下 で は生育CO2 濃 度下にお け る個 葉 光 合 成速度や,
光合 成 活 性,
窒 素 量 の低下 が 見 られる こと が わ かっ た。
ま た糖 類(可溶 性 糖 類 お よ びデンプン)が蓄 積し,
代 謝 物で は,
光 合 成に関わ る 糖 リン酸 系 とミトコ ン ドリア のエ ネル ギー
生 産 に関わ る Satoshi KITAOKA,
Akira UEMURA,
Hiroyuki TOBITA,
Hajime UTSUGI (Hokkaido Branch Center,
FFPRI,
Sapporo
O62
−
8516),
Yutaka MARUYAMA (Forestry Agency,
Tokyo lOO−
8950),
Atsuko MIYAGI,
Hirofumi UCHIMIYA (Institutefor
Environrnental Science and Technology
,
Saitama Univ.
Saitama 338−
8570)Leafphotosynthetic and metabolornic responses ofwillow (
SaliX
pet−
susu Kimura)to elevated CO2 concentration.
一
53
一
The Boreal Forest Society
NII-Electronic Library Service
The Boreal Forest Society
日林北 支 論59
,
Feb.
2011 TCA 回路で低下 が 見られ るこ と が わ かっ た。
今後の課 題 と してクロー
ン間に おける 生 理特 性 や 糖 類 蓄積の違い の 有 無 を 明 らかに してい く 必要が ある と考える。
特に光 合 成に負の影 響 を あた える 糖 類蓄 積の評価は 重要であるた め,
迅 速 な測定 方法の開発につ いても 取 り組 ん でい き た い と考える。
謝 辞:本 研究は 生 研セ ンター
のイノベー
シ ョ ン創 出 基 礎 的 研 究 推 進 事 業とNEDO
プロ ジェ ク トの支援を受け た。
引用文 献(」) Farquhar GD
.
von Caemmerer S.
and Berry JA.
(1980)Abiochemical model of photosynthetic CO2 assimilation
in leaves ofC3 species
.
Planta 149:78−
90.
(2) Griffin KL
.
and Seemann JR.
(1996
)Plants,
CO2 and photosynthesis in the 21st century.
Chem.
Biol,
3:245
−
254.
御 IPCC (2007 )Forth assessment report
.
一(4p Kabeya D
.
and Sakai S.
(2003)The RQIe of Roots and Cotyledons as Storage Organs in Early Stages ofEstablishment in
guercus
cris ρula; aQuantitat
正veAnalysis of the NonstructUral Carbohydrate in Cotyledons and Roots
.
Ann.
Bot.
92:537−
545,
(5) Karp A
,
and Shield 1.
(2008)Bioenergy from plants andthe sustainable yield challenge
.
Ncw Phytol.
179:15−
32.
(6) Kitao M
.
,
Koike T.
,
Tobita H.
and Maruyama Y (2005 )Elevated CO2 and limited nitrogen nutrition can restrict excitation energy dissipation in photosystem II of Japanese white birch(Betula piaり
yphylla
var.
ノaρonica )1eaves
.
Phys孟ol.
Plant.
125:64−
73.
(7) 小池 孝 良(2004 )温暖化 と植物の生 態
,
植 物 生 態学,
朝倉 書 店
,
東 京,
pp361−
391,
(8) Koike T
、
,
Kohda H.
,
Mori
S,
,
Takahashi
K,
Inoue MT.
and Lei TT
.
(1995)Growth responses of the cuttings of two willow species to elevated CO2 and temperature.
Plant Species Biol
.
10:95−
101,
C9
) Long SP,
and Bernacchi CJ.
(2003)Gas exchange measurements,
what can they tell us about the underlyinglimitations to photosynthesis?Procedures and sources of crror
,
J.
Exp.
Bot、
,54:2393
−
2401.
μo)牧 野 周 (1999)CO2と光 合 成
.
植 物の環境応 答一
生存戦 略 とその 分子機 構
一
,秀 潤 社,
東 京,
ppl34−
141.
(〃丿Miyagi A
.
,
Takahashi H,
,
Takahara K.
,
Hirabayash{ T.
,
Nishimura Y
.
Tezuka T、
.
Kawai−
Yamada M.
and Uch血iya H.
(2010) Princ孟pal component and hierarchical clustering analysis of metabolites indestructive weeds ;
polygonaceous
plants.
Metabolomics6:146
−
155,
(1 2?Ono K
,
,
Terashima I.
and Watanabe A,
(1996)Interaction between nitrogen deficit of a pEant and
nitrogen content in the old Ieaves
.
Plant Cell Physiol.
37:1083
−
1089.
(1 3)Onoda Y
.
Hikosaka K.
and Hirose T,
(2005)Seasonalchanges
in
thebalance
between capacities of RuBPcarboxylation and RuBP regeneration af正bcts CO2 response ofphotosynthesis in Po!
ygonum
cuspidatum.
J.
Exp
.
Bot.
56:755−
763.
(14)Sage RF
.
Sharkey TD,
and Seemann JR.
(1989)Acclimation
gfphotosynthesis
to elevated CO2 in five C3species
.
Plant Physiol.
89:590−
596.
(15) 酒 井 昭 (1995>植 物の分 布 と環 境 適 応
.一
熱 帯か ら極地
・
砂 漠へ一
.
朝 倉 書 店,
東 京,164pp,
(1 6)Sheen J
.
(1990)Metabolic repression oftranscription inhigher plants
,
Plant Cell 2:1027−
1038.
(1 7)Stitt M (1991)Rising CO21evels and their potential
significance fbr carbon flow in photosynthetic cells
.
Plant Cell Environ
.
14;741−
762.
表
一
1 エ ゾノ キヌヤ ナ ギの 葉の 光合成特 性 と 窒素 含 有量,
デンプン
,
可溶性糖 濃 度370ppm[CO2] 720ppm [CO2] t